









-
Waterlogging caused by extreme rainfall is becoming increasingly common due to the deteriorating global environment, which is the most common natural hazard to agricultural production[1]. Approximately 16% of the arable areas in the world are adversely affected by waterlogging, which is predicted to increase due to the occurrence of extreme weather and heavy rains under climate change[2,3].
Tomato (Solanum lycopersicum L.) from Solanaceae is one of the most important vegetable crops cultivated extensively worldwide. Asia, Americas, Europe, and Africa accounted for 54.9%, 17.3%, 15.5% and 11.9% of total tomato production, respectively, based on the average results from the year of 1994 to 2021, where China is the biggest tomato producer (
www.fao.org/faostat/zh/#search/tomato%20production ). As the worst affected areas by waterlogging in China, North China Plain and Yangtze valley areas accounted for more than three-quarters of the affected area, which was the major tomato production area. Tomatoes are highly sensitive to waterlogging stress[2,4]. Waterlogging has negative impacts on the vegetative and reproductive organs of tomatoes[2,5,6]. Waterlogging stress for three days induced increased relative ion leakage, lipid peroxidation and hydrogen peroxide (H2O2) content but decreased chlorophyll content, together with 52 differentially expressed proteins in leaves of tomato at seedling stage[7]. Short term waterlogging (one to three days) strongly affected the flowering and fruit maturity of tomato plants, resulting in decreased fruit setting percentage in all the tomato cultivars under waterlogging than the control[5]. Ide et al.[6] found that waterlogging stress initially caused decreased fruit number and then decreased average fruit weight, which finally contributed to significantly reduced tomato yield.Usually, abiotic stresses at reproductive stage exhibited more negative effects on plants than that at generative stage, such as heat stress[8,9], drought stress[10] on tomato plants and so on. Similarly, plants were more sensitive to waterlogging stress at the reproductive stage than at the generative stage[11]. For instance, de San Celedonio et al.[11] identified that the time around anthesis was the most sensitive period to waterlogging in wheat and barley. Ploschuk et al.[12] suggested that the influence of waterlogging on soybean grain production is strongly dependent on the phenological stage at which waterlogging occurs, which showed that soybean at reproductive stages were far more sensitive to waterlogging stress as compared with the other phenological stages. Li et al., found that prior mild waterlogging or drought was conducive to the homeostasis of oxidative metabolism and better photosynthesis, and thereby to less grain yield loss of wheat during later spring with low temperature stress[13]. What's more, the response of crops to waterlogging stress at the reproductive stage was closely associated with the final yield of crops, making the effects of waterlogging stress on plants at the reproductive stage more important to study. Therefore, as one of the most important fruit vegetable crops, tomato flower number and fruit set is the key trait for evaluating waterlogging tolerance, thus evaluating waterlogging tolerance of tomato at the anthesis stage is of significant importance.
Due to the high demand and significant economic value of tomato, and its sensitivity to waterlogging stress, more efforts on elucidating the physiological and biochemical responses of tomato plants at the reproductive stage to waterlogging stress were necessary. We aimed to clarify the effects of waterlogging stress on the leaf physiology and fruit traits of different tomato genotypes. Therefore, we investigated the alterations of plant phenotypic characteristics, leaf chlorophyll content, gas exchange, activities of antioxidant enzymes, reactive oxygen species (ROS), and membrane lipid peroxidation in six tomato genotypes under waterlogging conditions. Moreover, the yield and quality of different tomato genotypes was compared between controlled and waterlogged tomato plants. Based on correlation analysis and principal component analysis (PCA), the waterlogging tolerance of six tomato genotypes was compared. Our results will provide knowledge to understand the physiological regulatory mechanism and its association with final production in tomatoes at flowering stage under waterlogging stress.
-
Six tomato genotypes (No. 1, 'MIX-002'; No. 3, 'LA4440'; No. 7, 'Fenbeibei'; No. 9, 'Hezuo 908'; No. 10, 'Jinzhu'; and No. 16, 'Ruifen 882') were used as plant materials. Three tomato genotypes (No. 1, 'MIX-002'; No. 7, 'Fenbeibei'; and No. 9, 'Hezuo 908') at seedling stage were identified as waterlogging-tolerant candidates. In contrast, three tomato genotypes (No. 2, 'LA4440'; No. 5, 'Jinzhu', and No. 6, 'Ruifen 882') at seedling stage were identified as waterlogging-sensitive candidates. Among the six genotypes, the seeds of 'MIX-002', 'Hezuo 908' and 'LA4440' were from the Laboratory of Vegetable Physiology and Ecology, Nanjing Agricultural University (Nanjing, China). The seeds of 'Fenbeibei', 'Jinzhu' and 'Ruifen 882' were from Shouguang Xinxinran Gardening Company Limited, Taiwan Farmers KNOWN YOU Seedling Company Limited and Rixwan Agriculture, Netherlands Services Limited, respectively. The seeds were sown in 72-hole cavity trays filled with substrates (the mixture of grass charcoal: vermiculite: perlite, volume ratio = 2 : 1 : 1). After irrigating the substrates thoroughly using approximately 1.5 L water, a single seed was sown in each hole on 28th Feb 2022. The trays were covered with plastic film to maintain moisture of the substrates in the plastic greenhouse, which was removed seven days after sowing. The temperature was 25 ± 6 °C and the relative humidity was 50 ± 15% in the plastic greenhouse with sunlight. The seedlings with 7−8 fully expanded leaves were transplanted in the field (Baima experimental base of Nanjing Agricultural University, Nanjing, China) on 8th Apr 2022, after when the average atmospheric temperature was generally above 10°C at night-time. Row and line spacing were 70 cm and 50 cm, respectively, during the transplanting.
Experimental treatments
-
The plants at the full flowering stage (75% of plants exhibiting open flowers) were treated under control (CK) and waterlogging (WL), respectively, on 6th May 2022, lasting 21 d. The plants under CK were irrigated normally without excess water, while the plants under WL were irrigated until the field was saturated with water. The height ponding under WL was 2−3 cm above the soil surface, which was maintained by irrigating the experimental area daily. There were five plants per treatment per genotype in three plots with 180 tomato plants in total.
Measurements
Chlorophyll content
-
The chlorophyll content of the fully expanded leaf below the second inflorescences (from bottom to top) was measured on 27th May 2022. The 0.1 g fresh leaf samples were taken, cut into pieces, and added with 10 mL 95% ethanol. The samples were completed immersed into ethanol, which was put in darkness at room temperature for 24 h. The absorbance of the extraction was measured at 665 nm, 649 nm and 470 nm using Microporous plate detecting instrument (Cytation3, BioTek, USA), with 95% ethanol as control. Total chlorophyll content was calculated by the sum of chlorophyll a and b content.
Gas exchange parameters
-
Net photosynthetic rate (PN), stomatal conductance (Gs), transpiration rate (Tr), and intercellular CO2 concentration (Ci) of the fully expanded leaf below the second inflorescences was measured using Li-6400 portable photosynthesis system (LI-COR Company, USA) on the 23rd May 2022. The temperature and light intensity were controlled at 25 ± 1 °C and 300 μmol/m2/s, respectively, in leaf chamber, where the flow rate was 500 μmol/s and the CO2 concentration was 400 ± 10 μmol/L.
Activities of antioxidant enzymes
-
Leaf samples were taken from the leaves below the second inflorescence of the plants under different treatments on the last day of the treatments on 27th May 2022. The 0.2 g leaves were added with 0.05 mol/L pre-cooled phosphatic buffer solution/PBS (pH 7.8) in a pre-cooled mortar. The samples were ground into a homogenate on an ice bath, mixed thoroughly and centrifuged at 12,000 rpm and 4 °C for 20 min. Supernatant was used for measuring activities of antioxidant enzymes including superoxide dismutase (SOD), catalase (CAT), peroxidase (POD), and ascorbate peroxidase (APX). The activities of SOD and CAT were detected using the nitrogen blue tetrazole (NBT) method[14] and guaiacol method[15], respectively. The activities of APX and POD were determined using spectrophotometry[16,17]. The activities of SOD, POD, CAT and APX were determined at 560, 470, 240 and 290 nm, respectively, using UV-5500PC spectrophotometer (Metash, China).
Malondialdehyde (MDA), H2O2, proline content and O2·− production rate
-
Leaf samples were taken in the same way as the above measurements for enzyme activities.
The measurement of MDA content was based on the thiobarbituric acid (TBA) method[18] with modifications. The 0.2 g leaf samples were mixed and ground using 5% trichloroacetic acid (TCA). The supernatant was taken after centrifugation and added to 67% thiobarbituric acid (TBA). After mixing and shaking, the samples were sealed in a boiling water bath for 30 min and quickly cooled. Two hundred uL of supernatant was taken after centrifugation, which was measured at 450 nm, 532 nm and 600 nm using Microporous plate detecting instrument (Cytation3, BioTek, USA).
The H2O2 content was measured using the potassium iodide method[19]. The 0.2 g leaf samples were taken, and ground using liquid nitrogen and mixed with 0.1% TCA. The mixture was centrifuged at 3,000 rpm for 20 min. The supernatant was mixed with 1 M KI solution and 100 mM potassium sulphate, which was in darkness for 1 h. The absorbance at 390 nm was measured using Microporous plate detecting instrument (Cytation3, BioTek, USA) with 0.1% TCA as a reference.
The proline content was detected using the ninhydrin colorimetric method from Ábrahám et al.[20]. The leaf samples were mixed with 2 mL 3% sulfosalicylic acid solution and ground. The mixture was incubated in boiled water for 10 min and then centrifuged. Glacial acetic acid and acidic ninhydrin were added in supernatant, which was incubated in boiled water for 30 min. The samples after cooling were mixed with toluene, the upper layer of which was taken to measure the absorbance at 520 nm using Microporous plate detecting instrument (Cytation3, BioTek, USA).
The O2·− production rate was determined according to the method from Ke et al.[21]. Leaf samples were placed in a pre-cooled mortar and mixed 0.05 mol/L pre-cooled PBS (pH 7.8). The samples were ground into a homogenate and centrifuged at 12,000 rpm at 4 °C for 20 min. The supernatant was mixed with 0.05 M PBS (pH 7.8) and 10 mM hydroxylamine hydrochloride. The mixture was placed at 25 °C for 1 h and added with 17 mM p-aminobenzenesulfonic acid solution and 7 mM α-naphthylamine solution. The samples were mixed and vortexed, which were put at 25 °C for 20 min and then centrifuged at 3,000 rpm for 3 min. Absorbance were measured at 530 nm using Microporous plate detecting instrument (Cytation3, BioTek, USA).
Plant morphological indicators and fruit harvest
-
Plant morphological parameters were investigated on the last day of the treatments on 27th May 2022. Plant height was measured from cotyledonary node to growing point of the plant using a ruler. Stem diameter (1 cm from cotyledonary node) was measured using a vernier caliper. Leaf and inflorescence numbers of plants were counted.
Afterwards, the flower number per plant, single fruit weight, fruit number per plant, and fruit weight per plant were counted/weighed on 27th May 2022. The fruits were harvested on the same day for the following measurements. Red ripe fruits on the second inflorescence were harvested to measure transverse and longitudinal diameter, hardness, and single fruit weight. The transverse and longitudinal diameter of the fruit were measured using a vernier caliper. The hardness of the fruit was measured using a hand-held hardness tester (AGY-2, Dongguan, Guangdong Province, Three Measuring Tools Co., Ltd, Guangdong, China). Afterwards, the fruits were peeled, and the flesh was taken to determine the content of soluble solid, soluble protein, soluble sugars, and VC. Soluble solid content was detected using hand-held saccharimeter (Alto PAL-1, Japan). Soluble protein content was detected using coomassie brilliant blue G-250 staining method based on Zhang et al.[22]. The leaf samples were extracted in the same way as that for enzyme extraction. Coomassie brilliant blue solution was mixed with enzyme solution. The absorbance of the sample at 595 nm was determined after 2 min of reaction using Microporous plate detecting instrument (Cytation3, BioTek, USA). Soluble sugar content was determined using the colorimetric method as follows. The 0.2 g fruit was ground in 4 mL 80% ethanol and incubated in 80 °C water bath 40 min. The sample was centrifuged at 4,000 rpm for 15 min and the supernatant was collected. The residue samples were extracted twice using 2 mL 80% ethanol. The supernatant was mixed and added with 10 mg active carbon, and decolorized at 80 °C for 30 min. The mixture was filtered, mixed with 5 mL anthrone, and immediately incubated in a boiling water bath for 10 min. The absorbance of the samples at 625 nm were measured using Microporous plate detecting instrument (Cytation3, BioTek, USA). The VC content was detected using the red phenanthroline colorimetric method from Zhang & Kirkham[23].
Data analysis
-
All the measurements were conducted with three biological replicates. The data was analyzed in each genotype between control and waterlogging treatment using SPSS software using t-test (p < 0.05). Correlation analysis and principal component analysis of the above indicators were carried out using Origin and SPSS software, respectively.
-
The plants of all the six genotypes showed smaller size and less leaf areas under waterlogging stress as compared with the controls (Supplemental Fig. S1). The plant height of all the six tomato genotypes significantly decreased ranging from 40.4% (No. 10, 'Jinzhu') to 54.8% (No. 16, 'Ruifen 882') under waterlogging stress than the respective controls (Fig. 1a). Similarly, waterlogging stress induced significant reduction in the stem diameter of all the six genotypes ranging from 21.5% (No. 9, 'Hezuo 908') to 61.4% (No. 16, 'Ruifen 882') (Fig. 1b). The number of leaves in plants except for genotype No. 3 was significantly lower under waterlogging stress than the respective controls (Fig. 1c). More importantly, the number of inflorescences of all the six tomato genotypes significantly decreased 64.2%, 57.1%, 41.8%, 60.1%, 46.6% and 60.1%, respectively, under waterlogging stress as compared with the respective controls (Fig. 1d).
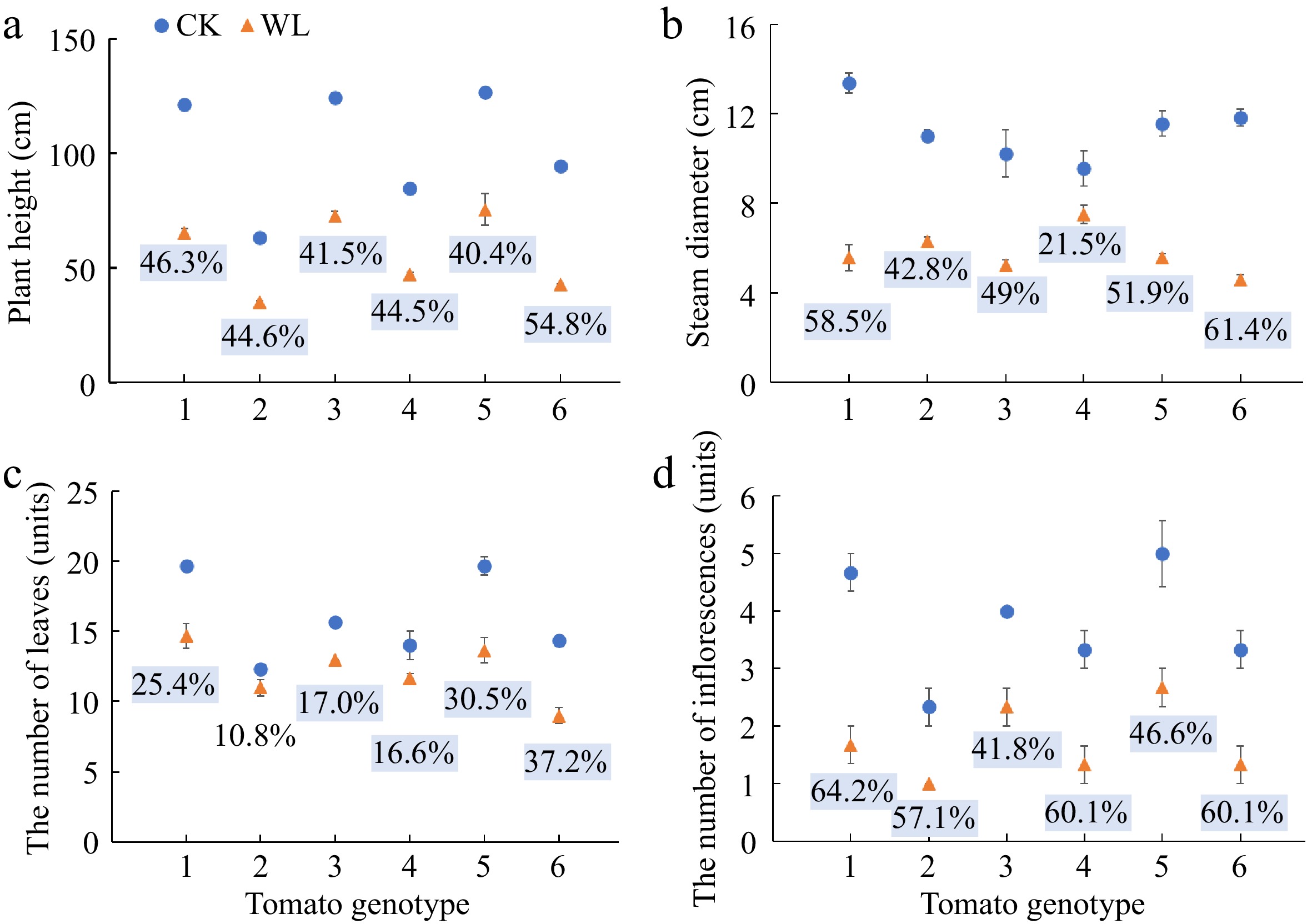
Figure 1.
(a) Plant height, (b) stem diameter, (c) number of leaves, and (d) number of inflorescences of six tomato genotypes at anthesis stage under CK and WL. The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). The percentages (blue square) referred to significantly decreased percentages of the parameters in each genotype under waterlogging treatment as compared with the respective controls (p < 0.05). The percentages without color indicated no significant difference (p < 0.05).
Chlorophyll content
-
Leaf chlorophyll a content of genotype No. 1, No. 3 and No. 10 significantly decreased 66.7%, 35.2% and 62.5%, respectively, and leaf chlorophyll b content of genotype No. 1 and No. 16 significantly decreased 31.9% and 20.0%, under waterlogging stress as compared with the respective controls (Fig. 2a, b). Waterlogging stress significantly decreased leaf total chlorophyll content of No. 1 (51.1%), No. 3 (21.0%) and No. 16 (44.5%) (Fig. 2c).
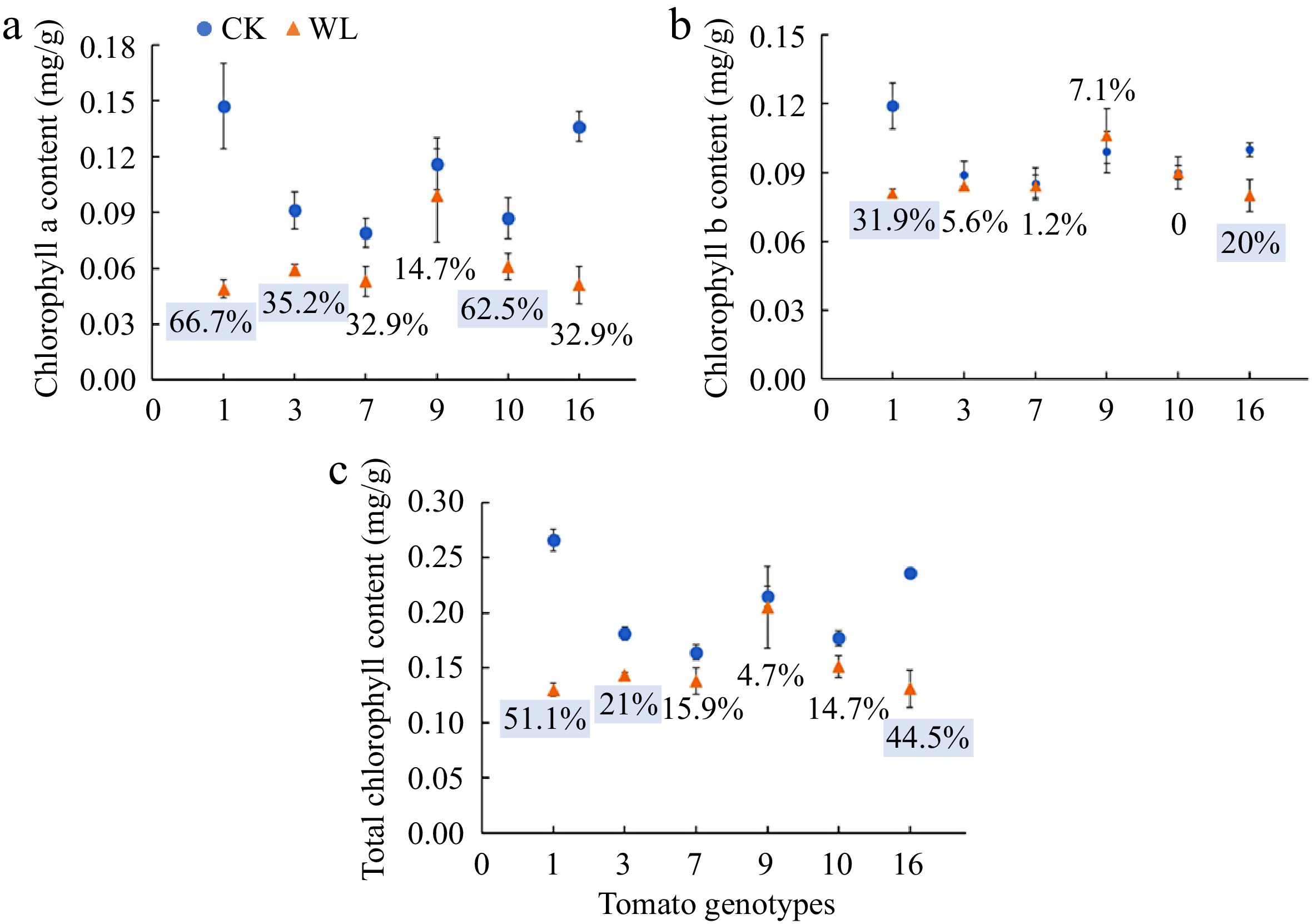
Figure 2.
(a) Chlorophyll a content, (b) chlorophyll b content, (c) total chlorophyll content of six tomato genotypes at anthesis stage under CK and WL. The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). The percentages (blue square) referred to significantly decreased percentages of the parameters in each genotype under waterlogging treatment as compared with the respective controls (p < 0.05). The percentages without color indicated no significant difference (p < 0.05).
Leaf gas exchange
-
The PN of genotype No. 1 significantly decreased 17.5% under waterlogging stress as compared with the respective controls (Fig. 3a). Waterlogging stress induced significant drops in the Ci of No. 7 (42.6%), No. 9 (24.5%) and No. 16 (18.9%) (Fig. 3b). The Gs of No. 1 and No. 3 significantly decreased 40.6% and 20.7% respectively, and the Tr of all the six genotypes except for No. 7 significantly decreased, under waterlogging stress as compared with the respective controls (Fig. 3c, d).
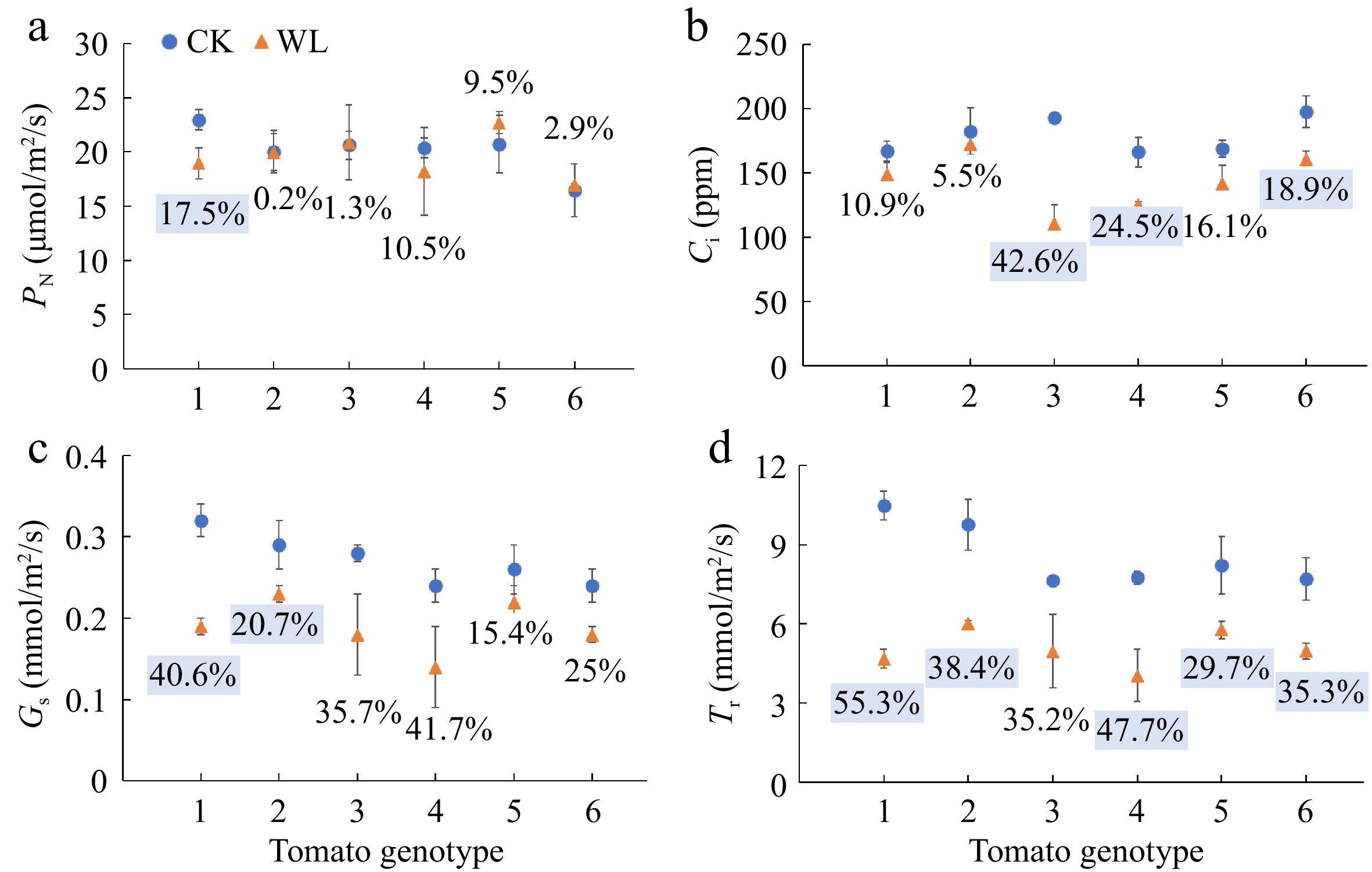
Figure 3.
(a) Net photosynthetic rate (PN), (b) intercellular CO2 concentration (Ci), (c) stomatal conductance (Gs), and (d) transpiration rate (Tr) of six tomato genotypes at anthesis stage under CK and WL. The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). The percentages (blue square) referred to significantly decreased percentages of the parameters in each genotype under waterlogging treatment as compared with the respective controls (p < 0.05). The percentages without color indicated no significant difference (p < 0.05).
Activities of antioxidant enzymes
-
The SOD activity of No.1, No. 3 and No. 16 significantly decreased 10.6%, 6.5% and 25.9%, respectively under waterlogging stress than the respective controls (Fig. 4a). As compared with the respective controls, the CAT activity of No. 9, and No. 16 significantly increased 74.9% and 159.9%, but that of No. 10 significantly decreased 67.3% under waterlogging stress (Fig. 4b). Waterlogging stress induced significant increases in the POD activity of No. 1 and No. 9, but significant drops in the POD activity of No. 3, No. 7 and No. 10 (Fig. 4c). By comparison, the APX activity of No. 1 and No. 7 significantly increased, but that of No. 10 and No. 16 significantly decreased under waterlogging stress as compared with the respective controls (Fig. 4d).
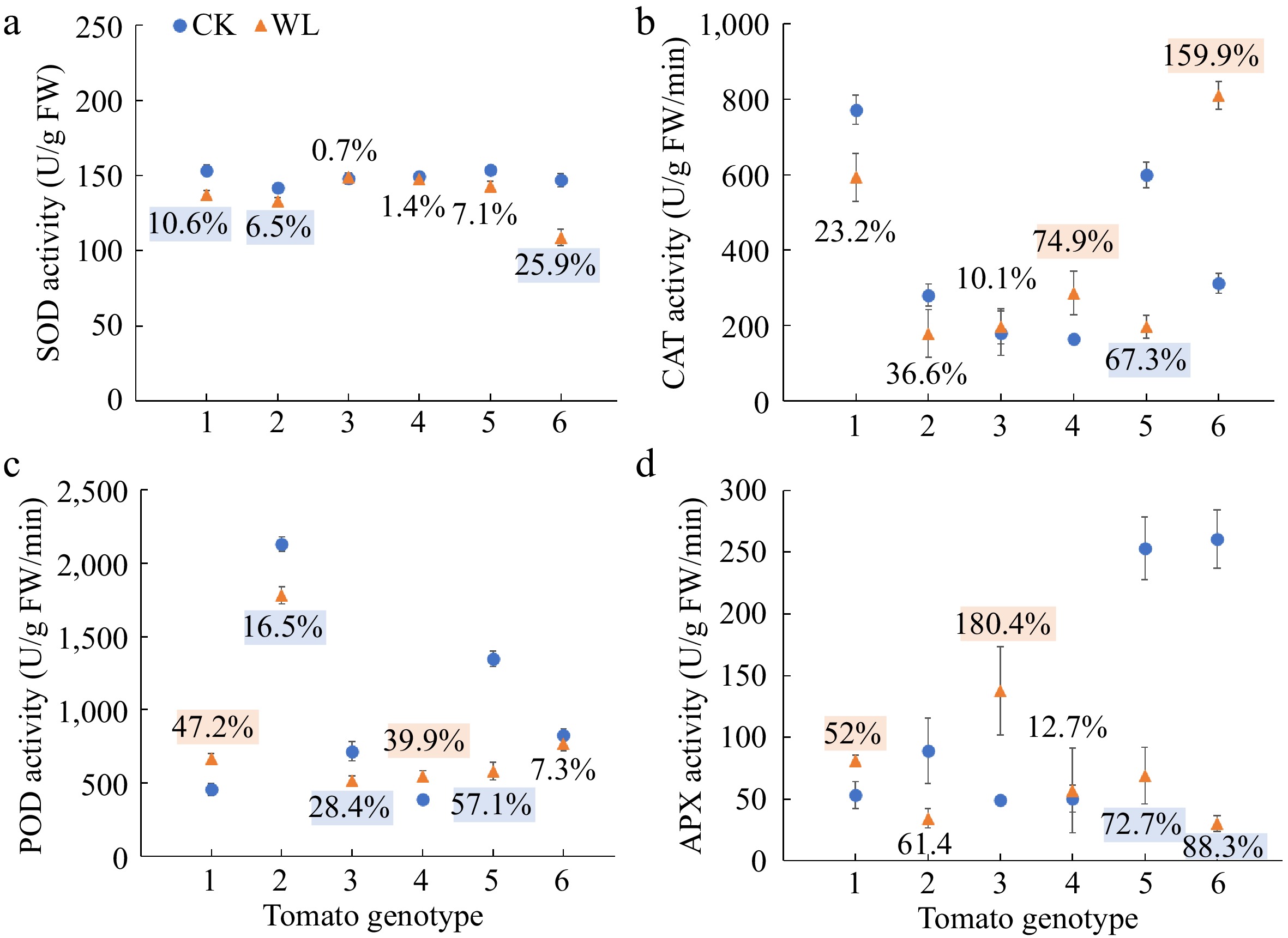
Figure 4.
Activities of (a) superoxide dismutase (SOD), (b) catalase (CAT), (c) peroxidase (POD), and (d) ascorbate peroxidase (APX) of six tomato genotypes at anthesis stage under CK and WL The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). The percentages (blue/orange square) referred to significantly decreased/increased percentages of the parameters in each genotype under waterlogging treatment as compared with the respective controls (p < 0.05). The percentages without color indicated no significant difference (p < 0.05).
Levels of leaf membrane lipid peroxidation under waterlogging stress
-
Waterlogging stress induced significant increases in the MDA content and H2O2 content of all the six genotypes (Fig. 5a, b). The proline content of No. 1 and No. 9 significantly decreased, while that of the rest four genotypes significantly increased under waterlogging stress as compared with the respective controls (Fig. 5c). Waterlogging stress significantly increased the O2·− production rate of No. 1, No. 3 and No. 16, but decreased that of No. 10 (Fig. 5d).
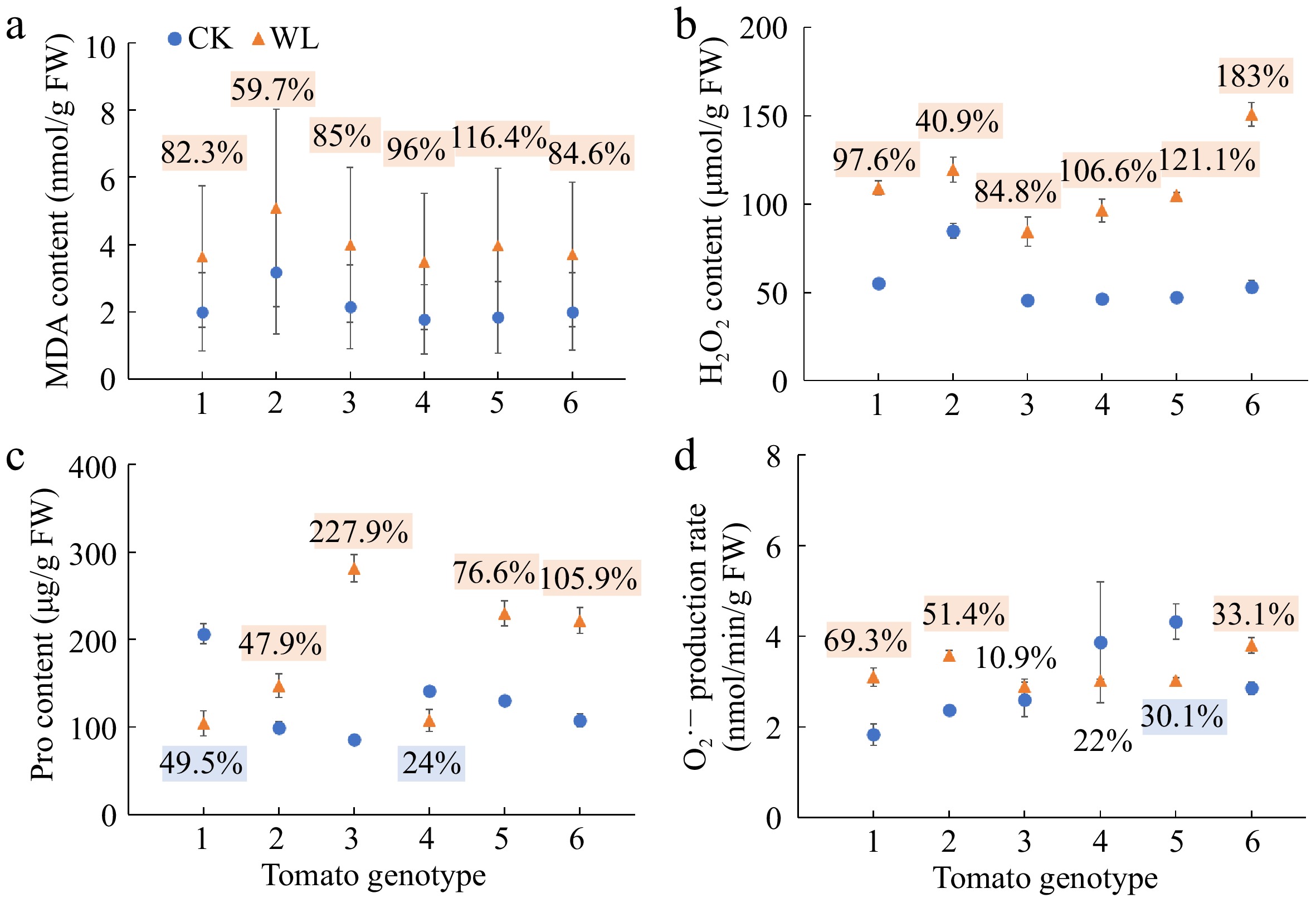
Figure 5.
(a) Malondialdehyde (MDA) content, (b) hydrogen peroxide (H2O2) content, (c) proline (Pro) content, and (d) superoxide anion (O2•─) production rate of six tomato genotypes at anthesis stage under CK and WL. The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). The percentages (blue/orange square) referred to significantly decreased/increased percentages of the parameters in each genotype under waterlogging treatment as compared with the respective controls (p < 0.05). The percentages without color indicated no significant difference (p < 0.05).
Changes in tomato yield under waterlogging stress
-
Moreover, the flower number per plant of all the genotypes except for No. 3 significantly decreased under waterlogging stress than the respective controls (Table 1). Waterlogging stress significantly decreased single fruit weight and fruit weight per plant of genotype No. 1, 7, 10 and 16, where the genotype No. 3, 9 failed to harvest tomato fruits (Table 1). The fruit number per plant of genotype No. 1 and No. 16 significantly decreased under waterlogging stress as compared with the respective controls (Table 1).
Table 1. Effects of waterlogging on yield of six tomato genotypes at the anthesis stage.
NO. Tomato genotype Treatment Flower number
per plantSingle fruit
weight (g)Fruit number per plant Fruit weight per plant (Kg) 1 MIX-002 CK 48 ± 5.0* 20.7 ± 1.22* 20 ± 1.0* 0.41 ± 0.02* WL 5 ± 1.5 9.0 ± 0.91 5 ± 3.0 0.04 ± 0.03 2 LA4440 CK 15 ± 2.8 69.7 ± 4.35 7 ± 0.3 0.46 ± 0.02 WL 4 ± 0.3 ─ ─ ─ 3 Fenbeibei CK 59 ± 10.4* 20.5 ± 0.28* 198 ± 4.0 4.06 ± 0.08* WL 21 ± 5.2 6.6 ± 0.06 70 ± 48.3 0.46 ± 0.32 4 Hezuo 908 CK 18 ± 2.3* 171.9 ± 14.52 16 ± 4.7 2.75 ± 0.81 WL 6 ± 1.2 ─ ─ ─ 5 Jinzhu CK 90 ± 14.4* 8.7 ± 0.36* 54 ± 7.2 0.47 ± 0.06* WL 24 ± 6.5 3.6 ± 0.40 26 ± 7.6 0.09 ± 0.03 6 Ruifen 882 CK 16 ± 2.2* 142.3 ± 13.71* 50 ± 2.9* 7.16 ± 0.41* WL 5 ± 2.9 16.0 ± 3.16 5 ± 0.6 0.05 ± 0.03 Note: The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). '*' indicated the significant difference of the same genotype between CK and WL (p < 0.05). Fruit characteristic of six tomato genotypes
-
The transverse and longitudinal diameter of fruits harvested from genotype No. 1, No. 7, No. 10 and No. 16 significantly decreased under waterlogging stress as compared with the respective controls (Fig. 6a, b). The fruit hardness of genotype No. 16 was lower, while the fruit soluble solid content of genotype No. 10 was higher under waterlogging stress than control (Fig. 6c, d). The content of soluble protein, soluble sugar, and VC of genotype No. 7, 10 and 16 was significantly higher under waterlogging stress than control, except the content of soluble sugar of genotype No. 16 (Fig. 6e−g).
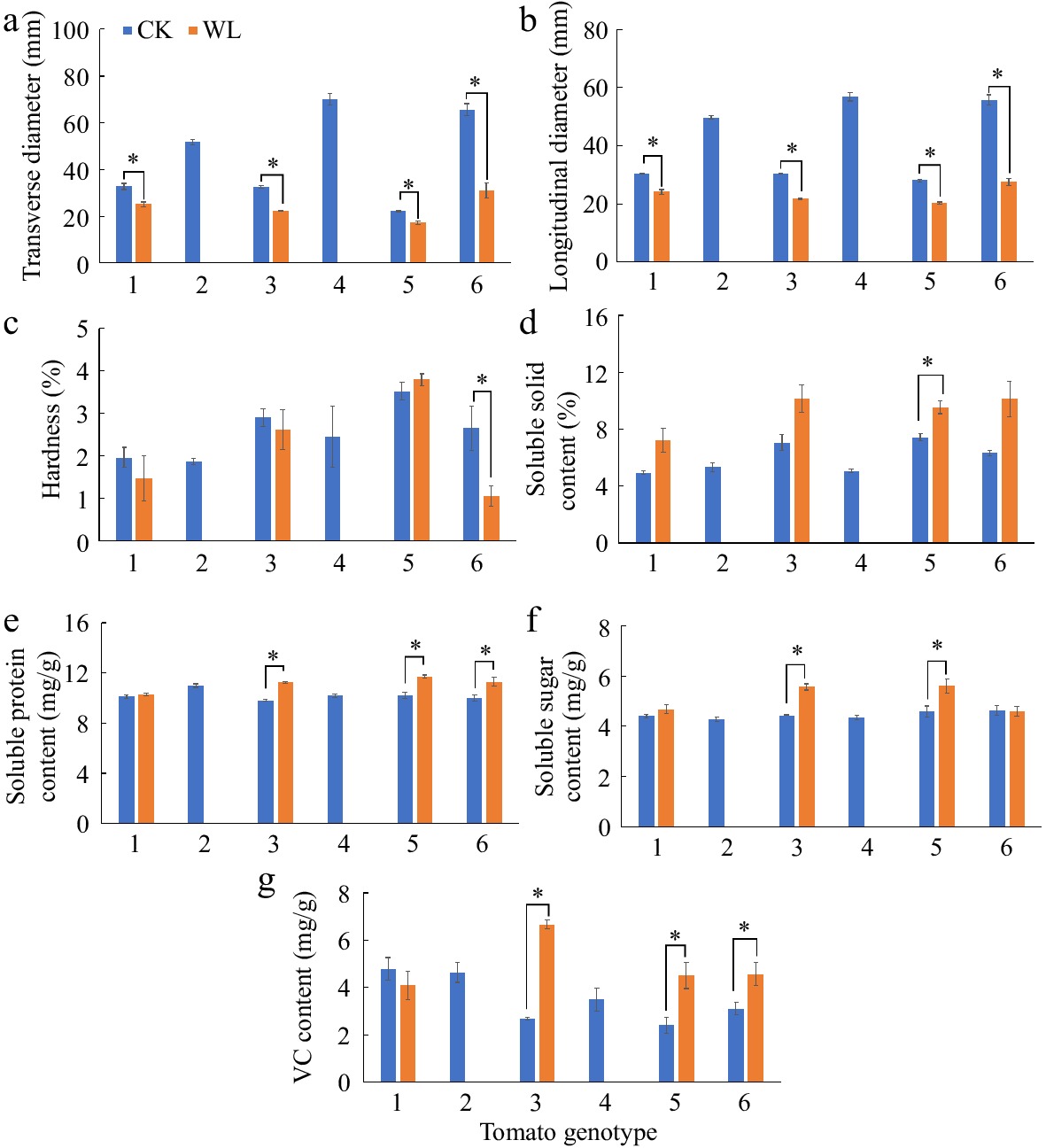
Figure 6.
(a) Transverse diameter, (b) longitudinal diameter, (c) hardness, (d) soluble solid content, (e) soluble protein content, (f) soluble sugar content, and (g) vitamin C (VC) content of the fruits from six tomato genotypes at anthesis stage under CK and WL. The CK and WL indicated control and waterlogging treatment, respectively. The values indicated mean ± SEM (n = 3). '*' indicated the significant difference of the parameters in each genotype under waterlogging treatment as compared with the respective controls; no marker indicated no significant difference (p < 0.05). Plants of genotype No. 2 and No. 4 under waterlogging treatment exhibited less than three replicates due to less than three fruits, resulting in blank bars.
Correlation between the physiological traits during tomato flowering period under waterlogging stress
-
We found that the leaf MDA and H2O2 content showed extremely significantly negative correlation with yield per plant (correlation coefficient = −0.53** and −0.55**, respectively) (Fig. 7). Meanwhile, the yield per plant was extremely significantly positive correlated with transverse (0.64**) and longitudinal diameter (0.60**) (Fig. 7). The PN, POD and CAT activities, proline content, O2·− production rate and fruit soluble solid content and protein content showed weaker correlation with the rest indicators (Fig. 7), which was removed during the PCA. Six principal components (F1−F6) were extracted with 87.254% accumulative contribution percentage based on the PCA (Supplemental Table S1). The evaluation formula for overall traits of tomato plants was F = 0.313 × F1 + 0.284 × F2 + 0.166 × F3 + 0.109 × F4 + 0.069 × F5 + 0.060 × F6 (Supplemental Tables S1, S2). Based on the formula, the F value of each tomato genotype was calculated, where Jinzhu and Ruifen 882 showed the highest and lowest F value (0.814 and −0.384, respectively) (Supplemental Table S3).
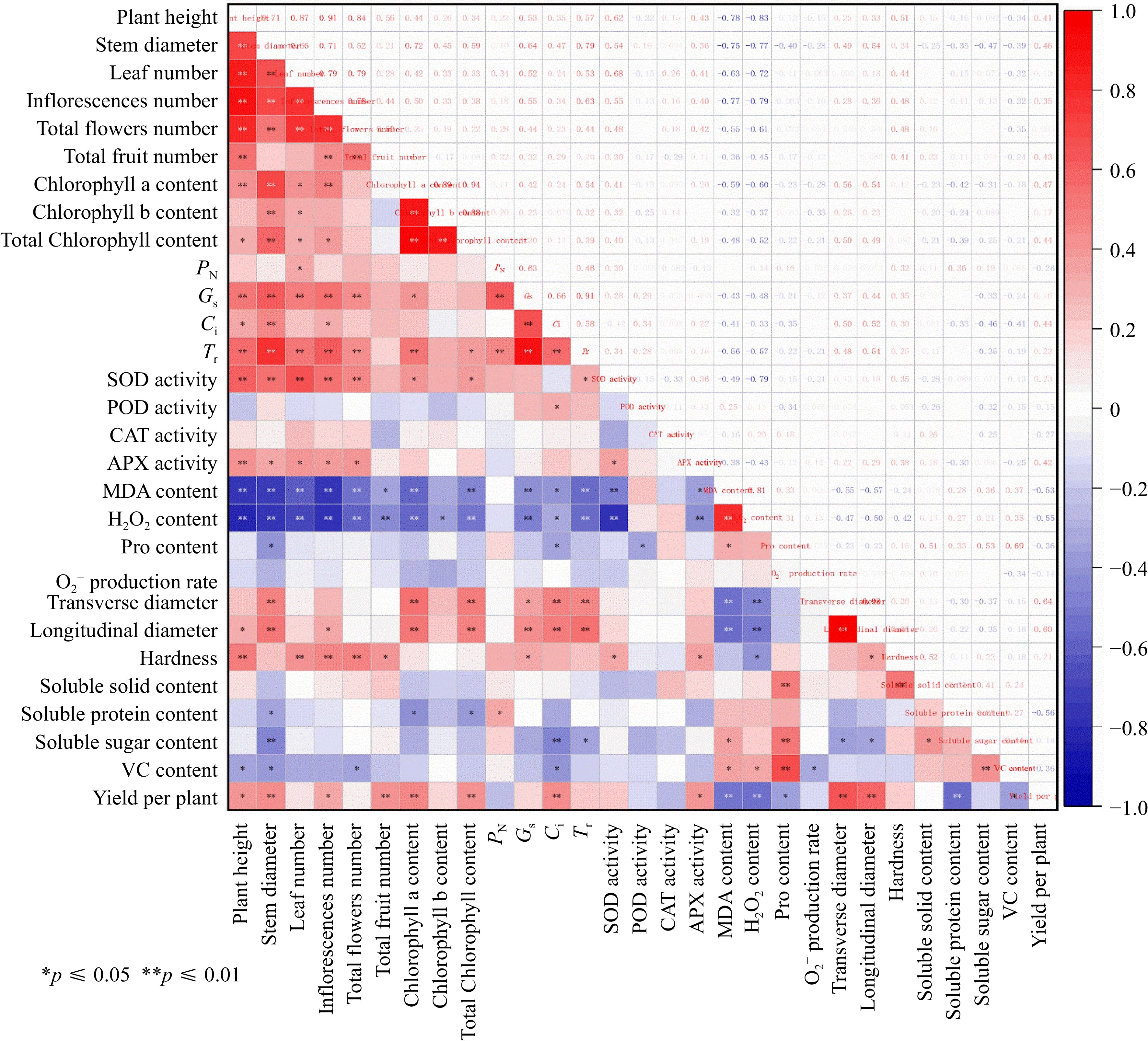
Figure 7.
Correlation analysis of all the measured parameters. PN: Net photosynthetic rate, Ci: intercellular CO2 concentration, Gs: stomatal conductance, Tr: transpiration rate, SOD: superoxide dismutase, CAT: catalase, POD: peroxidase, APX: ascorbate peroxidase, MDA: Malondialdehyde, Pro: proline, VC: vitamin C.
-
Usually, plants at the reproductive stage were more sensitive to abiotic stresses as compared with those at the seedling stage, such as high temperature[9,24], which is consistent with our findings for waterlogging stress. In our previous study, waterlogging stress for 7 d did not induce significant damage on two tomato genotypes at the seedling stage (data not shown). By comparison, the plant size and morphological traits of the six tomato genotypes at anthesis stage were all inhibited by waterlogging stress (Supplemental Fig. S1). These phenomena were accompanied by a smaller number of inflorescence and fruits and low yield in all six genotypes, corresponding to the negative effects of tomato flowering and fruit maturity by waterlogging as reported by Mohanty et al.[5]. In our previous study, high tomato fruit set under heat stress is aligned with maintained leaf carbohydrate content[9]. Here, tomato yield per plant was extremely significantly negatively correlated with the leaf MDA and H2O2 content, but significantly positively correlated with transverse and longitudinal diameter (Fig. 7). These indicated that the tomato yield under waterlogging conditions might be able to predict by leaf physiological parameters like increased MDA and H2O2 content. Moreover, smaller plant size as indicated by plant height and stem diameter, lower leaf chlorophyll content, the smaller fruit size as indicated by transverse and longitudinal diameter, and less total fruit number all together contributed to the lower tomato yield under waterlogging condition as compared with control. Using PCA, the six principal components with 87.254% accumulative contribution percentage can be applied to replace the 22 individual indicators to evaluate the waterlogging tolerance of tomato genotypes (Supplemental Table S1), which can reduce the number of variables and retain most of the original information. According to the F value from PCA, 'Jinzhu' was regarded as relatively waterlogging tolerant genotype, while 'Ruifen 882' was relatively sensitive (Supplemental Tables S1−S3).
It is well-known that waterlogging induces low gas diffusion and soil aeration, resulting in low internal CO2 concentration and lacking oxygen for plant respiration[25,26]. Li et al.[27] found that post-infiltration aeration for the tomato plants grown in pot with the drip tubing, especially at fruit setting (34–57 days after sowing (DAS)) and enlargement (58–85 DAS) stages, can enhance the tomato yield, nutrition, taste, and market quality of the tomato fruits, indicating the positive effects of soil aeration on tomato yield and fruit quality. Here we found that the number of inflorescences, fruit size, single fruit weight and fruit weight per plant of all the six genotypes significantly decreased by waterlogging treatment (Fig. 1, Fig. 6, Table 1), which could be partially explained by the low gas diffusion due to excessive rhizosphere water.
As a source organ in plants, leaf is the only resource to depend on for the existence of plants[28]. The leaf as source can supply carbon, while the fruit as sink is where the carbon is transported to and utilized[29]. We previously suggested that the fact that the stable chlorophyll content, photosynthetic ability, and carbohydrate content in the mature leaf of tomato plants under heat stress could be a precondition for flower and fruit development[9]. Therefore, the leaf traits under abiotic stress were closely associated with the final yield. Here, genotype No. 1 ('MIX-002'), No. 3 ('LA4440'), No. 10 ('Jinzhu'), and No. 16 ('Ruifen 882') exhibited the decreased pigment content (Fig. 2), and the gas exchange parameters were generally down-regulated (lower PN of 'MIX-002', lower Ci of 'Hezuo 908', 'Jinzhu' and 'Ruifen 882', decreased Tr of five genotypes) (Fig. 3) under waterlogging stress. Hence, the down-regulated chlorophyll content and photosynthetic parameters, especially transpiration rate in source leaf partially contributed to the low tomato yield under waterlogging condition.
Thirdly, SOD, CAT and POD are key enzymes regulating and scavenging ROS, where SOD can catalyze O2·− to form H2O2, and H2O2 can be converted into water and molecular oxygen catalyzed by CAT and POD, thus reducing the damage caused by stress[30,31]. We found that the SOD activity of 'MIX-002' decreased, while the POD and APX activity of 'MIX-002' increased (Fig. 4). The activities of CAT, POD and APX in 'Jinzhu' decreased, while the activities of CAT and POD in 'Hezuo 908' increased (Fig. 4). What's more, waterlogging induced the accumulation of MDA and H2O2 in all the six genotypes, being closely related to the low tomato yield (Fig. 5, Fig. 7). In consequence, waterlogging stress induced oxidative damage due to the accumulated reactive oxygen species, disequilibrated the ROS dynamic balance and antioxidant system in tomato plants at anthesis stage, which is also an important reason for inhibiting the growth, flowering, and fruit setting of tomato plants.
Overall, tomato plants at anthesis stage were sensitive to waterlogging conditions, since the fruit yield of all the six genotypes significantly decreased, even though 'Jinzhu' and 'Ruifen 882' were relatively tolerant and sensitive, respectively using PCA analysis. In conclusion, waterlogging stress adversely inhibited the growth and development of tomatoes at the anthesis stage due to the combination of low gas diffusion, down-regulated chlorophyll content and transpiration rate as well as ROS disorder.
-
The authors confirm contributions to the paper as follows: study conception and design: Jiang F and Zhou R; performed the experiments: Yin J, Niu L and Li Y; methodology and data analysis: Yin J, Song X; draft preparation: Niu L, Yin J and Li Y; draft review and editing: Ottosen C, Wu Z and Zhou R. All authors reviewed the final version of this draft and granted approval for its publication.
-
All data generated during this study are included within the article or its supplementary files.
We acknowledge the help from Ge Wang and Fei Ding and the funding from earmarked fund for China Agriculture Research System (CARS-23), Jiangsu Seed Industry Revitalization project - [JBGS(2021)015], and starting grant from Nanjing Agricultural University.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Jian Yin, Lifei Niu
- Supplemental Fig. S1 Comparison of six tomato genotypes at anthesis stage under control (above) and waterlogging stress (below).
- Supplemental Table S1 Total variance explanation.
- Supplemental Table S2 Component score coefficient matrix.
- Supplemental Table S3 Score of comprehensive waterlogging tolerance evaluation index of tomato.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yin J, Niu L, Li Y, Song X, Ottosen CO, et al. 2023. The effects of waterlogging stress on plant morphology, leaf physiology and fruit yield in six tomato genotypes at anthesis stage. Vegetable Research 3:31 doi: 10.48130/VR-2023-0031 

The effects of waterlogging stress on plant morphology, leaf physiology and fruit yield in six tomato genotypes at anthesis stage
- Received: 30 July 2023
- Accepted: 21 September 2023
- Published online: 05 December 2023
Abstract: Waterlogging stress caused by concentrated and heavy rainfall has become an increasingly popular abiotic stress, especially for tomato production. The aim is to clarify the leaf physiological and biochemical responses as well as flower and fruit set of tomato plants at reproductive stage under waterlogging stress. Six tomato genotypes including 'MIX-002', 'LA4440', 'Fenbeibei', 'Hezuo 908', 'Jinzhu' and 'Ruifen 882' at anthesis stage were treated under control and waterlogging stress. We found that plant height, stem diameter, inflorescences number, single fruit weight and fruit weight per plant of all the six tomato genotypes significantly decreased under waterlogging stress as compared with the control. Chlorophyll a content of 'MIX-002', 'LA4440' and 'Jinzhu' and total chlorophyll content of 'MIX-002', 'LA4440' and 'Ruifen 882' were significantly lower under waterlogging conditions than the control. Waterlogging stress induced low net photosynthetic rate of 'MIX-002' and decreased transpiration rate of six genotypes except 'Fenbeibei'. Antioxidant enzymes activity and proline content responded to waterlogging stress by up/down regulation, resulting in high malondialdehyde and H2O2 content in leaves of six genotypes. Overall, waterlogging stress caused low gas diffusion, negatively affected pigment content, decreased transpiration rate, and induced antioxidant system disorder in tomato plants at anthesis stage. These adverse effects contributed to few buds and flowers and finally resulted in yield loss of tomato plants under waterlogging conditions. The significant correlation between malondialdehyde and H2O2 content and tomato yield indicated that the malondialdehyde and H2O2 content in leaves could be applied to predict the tomato yield under waterlogging condition.
-
Key words:
- Tomato /
- Anthesis period /
- Waterlogging stress /
- Leaf physiology /
- Fruit yield and quality