








-
Soybean (Glycine max (L.) Merrill) is a multipurpose oilseed crop widely grown in tropical, subtropical and temperate climates. The crop is mainly cultivated for its seed that contains 30%–40% quality protein and 18%–25% cholesterol-free oil[1,2]. This legume crop is very strategic in most cropping systems because it fixes atmospheric nitrogen and thereby improves soil fertility[3], important in human and livestock nutrition[4], and capable of generating income for resource-constrained farmers in the tropics[5]. In 2020, about 353,463,735 million tonnes of soybeans were produced in the world and Africa accounted for just 0.9% (3,438,611 million tonnes) as reported in FAOSTAT[6]. Soybean seed yield in Africa has remained almost stagnant in the last decade. The yield stagnancy can be attributed to multifaceted factors such as cultivation of poor performing varieties, low fertility soil status, limited or non-fertilizer application, especially phosphorus, poor agronomic management practices, erratic rainfall and distribution and lack or poor usage of inoculants on soils without the previous history of soybean production and weed interference among others[7−9].
Organic soybean production remains less than 0.1% of the total world production despite the crop being the most widely cultivated oilseed crop. According to Willer et al.[10] organic soybean accounted for 55% of oilseeds cultivated on organic lands in 2021. Yield penalty of about 15%–40% has been reported in literature for organic soybeans relative to conventional production system[11−13]. Furthermore, seed yield values often recorded on smallholder farms for soybeans are lower than the potential yields of the varieties. The actual yield reported by FAO for Africa and Nigeria in 2020 were 1.7 and 2.4 times lower than the potential yield reported for the most widely adopted soybean variety (TGx 1448-2E) in Nigeria[14]. Consequently, several research studies have been conducted to bridge the gap between actual and potential yield, and reduce yield penalty of organic soybeans in the tropics. Such efforts include use of synthetic foliar fertilisers[15,16], soil[9] and foliar applied organic fertilisers[17], crop rotation[18,19], different cropping systems[20], biofertilizers and organic manures[21] integrated use of organic and inorganic fertilization[22,23], intercropping[24,25], integrated use of organic and inorganic fertilizers in mixtures[26]. Recently, application of phosphorus at the rate of 21–30 kg P ha−1 and sowing at the appropriate time were reported to have increased soybean yield by 48% and 300%, respectively in sub Saharan Africa (SSA) by Omondi et al.[27]. Nevertheless, the yield gap is yet to be substantially bridged in the tropics despite the use of the afore mentioned approaches. Also, the use of decision support tools was recently suggested as an approach to boost the output of smallholder farmers in crop production in Africa[28].
Crop management has been described as a set of technically and environmentally sound agricultural practices usually carried out to improve the growth, development and get potential yield and high net returns in any crop or crop sequence[29,30]. If properly carried out, these management practices are expected to reduce the problems in agricultural food production and thereby boost production output. A critical analysis of the literature reveals that no single piece of literature has combined the use of management practices such as organic and conventional systems, and inoculation to quantify the productivity of soybeans in the tropics. The hypothesis of the study was that there is no difference in the productivity of soybeans irrespective of the management system adopted in the tropics. Therefore, this study was carried out to evaluate the effects of different management practices on the phenology, grain yield and yield attributes of soybeans in the humid tropics. Additionally, the research findings from this study will establish the most appropriate management practice for optimum soybean production.
-
This experiment was conducted on the organic research plot of the Institute of Food Security, Environmental Resources and Agricultural Research, (IFSERAR), Federal University of Agriculture Abeokuta (FUNAAB) located on latitude 7°13'53'16' N and longitude 3°23'49.12'' E, and 3023'51.86' E 139 m above sea level, Abeokuta, Ogun State, South West Nigeria during the late cropping season (July–November) of 2020 and 2021. Traditionally, this area experiences bimodal rainfall distribution with two peaks in July and September and a dry spell in August termed 'august break”. However, this trend was not recorded in both years of experimentation. Rainfall only peaked in September of 2020 and August of 2021 as illustrated in Fig. 1. The late cropping season of 2020 growing season was 139 mm dryer than the average 35 year average (1985–2019) while 2021 was 12.8 mm wetter. August was markedly dry in 2020 but September recorded the highest rainfall that year. The two years of experimentation were similar in temperature and just 0.3 °C cooler than the long-term average. Overall, the weather conditions during the late cropping season were suitable for soybean production. On average, the soil of the experimental site was loamy sand in texture, slightly alkaline, low in nitrogen, high in phosphorus and low in potassium (Table 1)
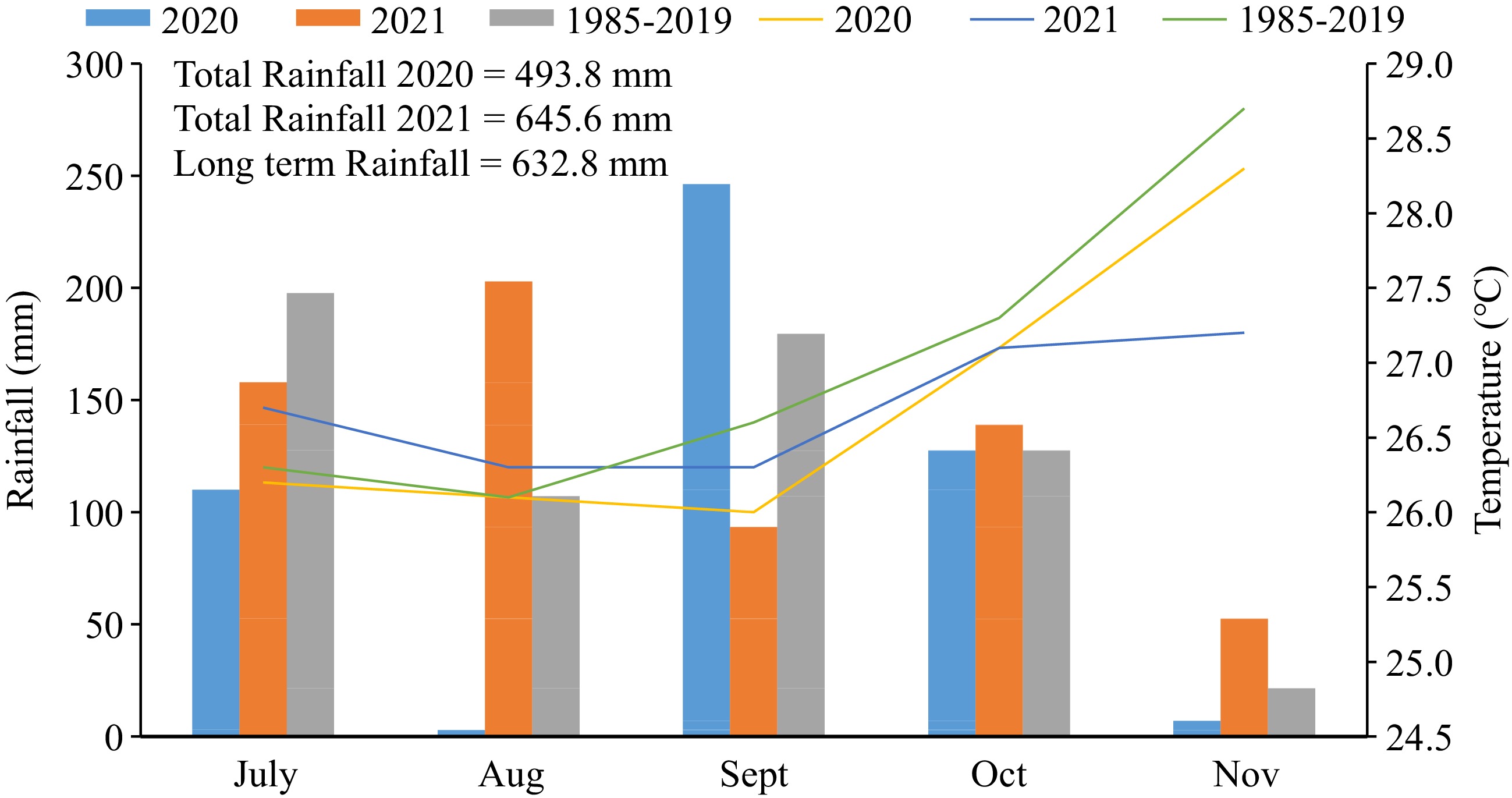
Figure 1.
Monthly rainfall and mean monthly temperature during the late cropping season (July–Nov.) of 2020, 2021 and long-term period (1985–2019). Source: Department of Agro Meteorology and Water Management Station, FUNAAB.
Table 1. Pre-planting physical and chemical properties of soil in 2020 and 2021
Parameters Conventional plot Organic plot 2020 2021 2020 2021 Physical properties Sand (%) 88.3 85.4 87.7 79.2 Silt (%) 11.0 11.4 10.9 17.7 Clay (%) 0.7 3.2 1.4 3.1 Textural class LS LS LS LS Chemical properties pH 7.25 7.55 7.34 6.98 N (g/kg) 0.60 0.61 0.62 0.49 OM (g/kg) 12.07 12.27 12.47 9.85 P (mg/kg) 44.81 39.82 29.59 48.87 K (mg/kg) 0.25 0.84 0.27 0.83 Ca (mg/kg) 671.44 905.18 631.62 682.73 Mg (mg/kg) 410.84 258.14 410.11 357.28 Na (mg/kg) 99.67 39.96 90.01 48.79 Mn (mg/kg) 4.20 3.07 4.70 4.55 Fe (mg/kg) 13.60 7.56 18.80 12.65 Zn (mg/kg) 6.80 9.75 9.70 8.33 Experimental materials
-
The test variety of soybean planted in the study was TGx 1448-2E, a late maturing variety (115–120 d), days to flowering (46 d), with good nodulation ability, good at causing suicidal germination to Striga hermonthica (parasite), resistant to lodging and pod shattering[14]. The organic fertilizer used was Aleshinloye Grade B, an abattoir based fertilizer. The fertilizer was analysed for nutrient content composition each year before any application and it contained 1.23% and 1.03% nitrogen in 2020 and 2021, respectively (Table 2). The inoculant used was NoduMax, a peat based inoculant that ensured the infection of soybean roots by symbiotic rhizobium bacteria thereby enhancing root nodulation and biological nitrogen fixation (BNF). It contains 50% culture rhizobia and 50% of the carrier material, peat. NoduMax was sourced from the International Institute for Tropical Agriculture (IITA) in Nigeria (
https://propas.iita.org ).Table 2. Nutrient composition of Aleshinloye organic fertilizer Grade B (Abattoir-based) in 2020 and 2021.
Parameters Year 2020 2021 N (g/kg) 12.30 10.30 P (g/kg) 3.40 2.90 K(g/kg) 2.00 3.50 Ca (cmol/kg) 1.90 1.00 Mg (cmol/kg) 1.10 0.50 Na (cmol/kg) 0.30 0.10 Mn(mg/kg) 53.70 44.80 Fe (mg/kg) 28.10 12.80 Zn (mg/kg) 10.30 11.00 Treatments and experimental design
-
The experiment involved the following six management practices (MP):
1. Control
2. Nodumax (inoculant)
3. Organic fertilizer
4. Nodumax + organic fertilizers
5. Conventional system (use of inorganic fertilizers and herbicides)
6. Nodumax + conventional system
The organic fertilizer (Aleshinloye Fertilizer (Grade B) contained 1.23% and 1.06% N, 33.18 and 29.08 mg/kg P, 0.20 and 0.35 mg/kg K in 2020 and 2021, respectively and was applied in quantities equivalent to 30 kg N ha–1[9]. This is line with the recommendation of starter nitrogen for soybean to boost its growth[31]. The conventional system involved the application of compounded fertilizer (30 kg N ha–1, 56 kg P2O5, and 60 kg K2O ha–1) and the spraying of pre-emergence herbicide (Satellite) immediately after sowing in both years. The six treatments were laid out in a randomized complete block design (RCBD) and replicated three times.
Crop husbandry
-
The experimental land was ploughed twice and harrowed once. Sowing of the test variety was done on July 28, 2020 and August 9, 2021 at a spacing of 60 cm × 5 cm to give a corresponding plant population of 333,000 plants per ha at the seed rate was 50 kg per hectare. The seeds were sown at a depth of 5 cm. The seeds sown under management practices (MP2, MP4 and MP6) were treated with the inoculant in the morning on the day of sowing. Weeding was done manually twice at 3 and 6 weeks after sowing (WAS). Each experimental gross plot size was 4 m × 3 m (12 m2) and net plot size 3 m × 2 m (6 m2), with walk way of 0.5 m between plots and 1 m between replicates. The total land area was 286 m2. The conventional plots that received synthetic agrochemicals were established at about 50 m away from the organic plots to avoid any commingling. Harvesting of soybeans was done annually at the harvest maturity stage (R8) according to Fehr & Caviness[32].
Data collection
-
For data collection, five randomly selected plants were tagged within the net plot at 3 WAS. Data collected on soybean phenology (number of phenological days to full bloom (R2), initial seed filling (R5), full seed (R6) and physiological maturity (R7), Grain Filling Period (GFP), nodulation (nodule count and weight). The GFP was calculated as difference between the number of phenological days to R6 and R5. Plant height at R7 was taken on the tagged plants on plot basis before harvest. After harvest, yield components such as the number of branches per plant (NBP), number of pods per plant (NPP), number of seeds per plant (NSP), pod weight (g) per plant (PWT) and seed weight (g) per plant, (SWT), and 100 seed weight, 1000 SWT (g) were measured. Seed yield was determined on plot basis after threshing all the pods and the weight per plot later converted to weight per unit land area in kg/ha. Every weight measurement was done using METTLER PJ260 Delta Range Model manufactured by Metler Instrument Ltd., High Wycombe, Switzerland.
Data analysis
-
All the data collected on the parameters were subjected to analysis of variance (ANOVA) to test the effects of the management systems for the two field trials separately by using the MSTATC package version 1.42[33]. After the ANOVA test, significant (p < 0.05, F-test) treatment means were separated by the least significant difference method at p = 0.05.
-
Rainfall distribution and mean temperature during the periods of experimentation in 2020 and 2021 are presented in Fig. 1. Year 2021 late cropping season was wetter than that of 2020.
The late cropping season of 2020 growing season was 139 mm dryer than the 35 year average (1985–2019) while 2021 was 12.8 mm wetter. August was markedly dry in 2020 but September recorded the highest rainfall that year. The two years of experimentation were similar in temperature and just 0.3 °C cooler than the long-term average. Overall, the weather conditions during the late cropping season were suitable for soybean production. On average, the soil of the experimental site was loamy sand in texture, slightly alkaline, and low in nitrogen, high in phosphorus and low in potassium (Table 1).
Effects of management practices on the phonology of soybeans in 2020 and 2021
-
Management practices significantly (p < 0.05) affected number of phenological days to full bloom (R2) and physiological maturity (R7) of soybeans in both years, except R7 in 2021. On average, days to R2 was delayed following the application inoculant (MP2), inoculant + organic fertilizer (MP4), synthetic fertilizers (MP5) and inoculant + synthetic fertilizers (MP6) during the dry year 2020 (Table 3). However, soybeans under MP6 was the latest to mature. Management practice significantly (p < 0.05) affected the duration of Grain Filling Period only during the wetter year 2021 (Table 4). Application of inoculant (MP2) and organic fertilizer (MP3) alone and in combination (MP4) significantly (p < 0.05) prolonged grain filling period by 5–7 d in 2021. This tend however, did not translate to significantly higher grain yields among the management practices in both years (Tables 5 & 6), especially when compared to the conventional system (MP5).
Table 3. Effects of management practices on the phenology of soybeans in 2020.
Management
practicesNumber of days to Grain filling
periodR2 R5 R6 R7 MP1 59.00c 86.00a 99.00a 107.00ab 13.00a MP2 61.33b 89.00a 100.00b 106.33ab 11.00a MP3 60.33bc 89.67a 100.67a 107.33ab 11.00a MP4 61.00b 90.00a 102.00a 106.33b 12.00a MP5 61.00b 88.00a 98.67a 109.00ab 10.33a MP6 63.33a 90.33a 101.00a 110.00a 10.67a SE ± (df-10) 0.615 2.388 2.346 1.358 1.653 MP1 – Control, MP2 – NoduMax, MP3 – Organic fertilizer, MP4 – NoduMax + Organic fertilizer, MP5 – Conventional, MP6 – Conventional + NoduMax, R2 – Full bloom, R5 – Initial seed filling, R6 – Full Seed, R7 – Physiological maturity. Means with the same letter along the column are not significantly different according to Duncan's Multiple Range Test (DMRT) at 5% probability level. Table 4. Effects of management practices on the phenology of soybeans in 2021.
Management
practicesNumber of days to Grain filling
periodR2 R5 R6 R7 MP1 59.33b 80.00c 94.33c 110.67a 13.67b MP2 61.33a 80.33c 99.67ab 112.67a 19.67a MP3 59.67b 82.00b 101.33a 111.67a 20.00a MP4 61.67a 83.33a 99.67ab 109.33a 18.00a MP5 59.33b 80.67c 97.00b 110.67a 16.33b MP6 59.67b 80.67c 97.33b 110.00a 16.67b SE ± (df 10) 0.577 0.494 1.517 1.512 1.723 MP1 – Control, MP2 – NoduMax, MP3 – Organic fertilizer, MP4 – NoduMax + Organic fertilizer, MP5 – Conventional, MP6 - Conventional + NoduMax, R2 – Full bloom, R5 – Initial seed filling, R6 – Full Seed, R7 – Physiological maturity. Means with the same letter along the column are not significantly different according to Duncan's Multiple Range Test (DMRT) at 5% probability level. Table 5. Effects of management practices on seed yield and yield attributes of soybeans in 2020.
Management practices PHTR7 NBR NPODS NSEEDS SWT (g) PWT (g) 100SWT (g) GYD (kg/ha) MP1 54.80ab 3.33b 40.07c 68.80d 4.70c 8.47c 6.90a 846.00c MP2 52.67ab 4.77b 79.57bc 97.93bcd 7.57b 12.83bc 7.73a 1362.00b MP3 56.77a 8.43ab 138.87a 137.20a 10.90a 24.17a 8.00a 1962.00a MP4 56.33a 8.57ab 86.87abc 126.90ab 9.37a 14.97abc 7.57a 1686.00a MP5 42.70b 11.50a 102.37ab 111.33abc 9.47a 19.53ab 6.57a 1704.00a MP6 49.43ab 5.90ab 112.63ab 79.53cd 6.47b 23.47ab 8.20a 1164.00b SE ± (df-10) 5.152 2.632 21.852 14.782 0.772 4.601 1.271 138.901 MP1 – Control, MP2 – NoduMax, MP3 – Organic fertilizer, MP4 – NoduMax + Organic fertilizer, MP5 – Conventional, MP6 – Conventional + NoduMax, PHTR7 – Plant height at physiological maturity, NBR – Number of branches, NPODS – Number of pods, NSEDDS – Number of seeds, SWT – Weight of seeds, PWT – Pod weight, 100SWT (100 seed weight), GYD – Grain yield. Means with the same letter along the column are not significantly different according to Duncan's Multiple Range Test (DMRT) at 5% probability level. Table 6. Effects of management practices on seed yield and yield attributes of soybeans in 2021.
Management practices PHTR7 (cm) NBR NPODS NSEEDS SWT (g) PWT (g) 100SWT (g) GYD (kg/ha) MP1 70.60a 4.47b 37.47a 166.13b 2.27c 6.33a 8.92a 754.80c MP2 72.00a 4.87ab 39.13a 212.30b 3.47abc 12.00a 8.12a 1155.40abc MP3 71.80a 4.03b 33.13a 189.53b 3.60ab 8.10a 7.37abc 1198.80ab MP4 67.87a 5.67ab 39.87a 241.47b 4.20ab 10.27a 7.59ab 1398.60ab MP5 59.27a 5.47ab 57.93a 413.93b 4.67a 9.00a 4.87b 1554.00a MP6 59.80a 7.13a 55.40a 819.57a 3.03b 9.33a 5.02b 999.03b SE ± (df-10) 5.677 1.012 12.089 182.192 0.548 3.532 1.121 182.836 MP1 – Control, MP2 – NoduMax, MP3 – Organic fertilizer, MP4 – NoduMax + Organic fertilizer, MP5 – Conventional, MP6 – Conventional + NoduMax, PHTR7 – Plant height at physiological maturity, NBR – Number of branches, NPODS – Number of pods, NSEDDS – Number of seeds, SWT – Weight of seeds, PWT – Pod weight, 100SWT (one hundred seed weight), GYD – Grain yield. Means with the same letter along the column are not significantly different according to Duncan's Multiple Range Test (DMRT) at 5% probability level. Effects of management practices on nodulation of soybeans in 2020 and 2021
-
Varying results were obtained on the effects of management practices on nodule count and dry weight per plant in both years (Fig. 2a, b). Higher number of nodules was recorded in 2021 than 2020 probably because of more favourable rainfall distribution. During the dry year 2020, the highest number and dry weight of nodules was recorded when organic fertilizer was applied alone (MP3) at 8WAS. Similarly, a combination of NoduMax and organic fertilizer (MP4) resulted in the highest number and dry weight of nodules at 9WAS in 2020. Comparatively high number of nodules were recorded at 8, 9 and 10WAS in 2021 when soybean seeds were inoculated with NoduMax alone (MP2) or in combination with organic fertilizer (MP4) at 9WAS or in combination with conventional practice (MP6) 8 and 10WAS (Fig. 2a).
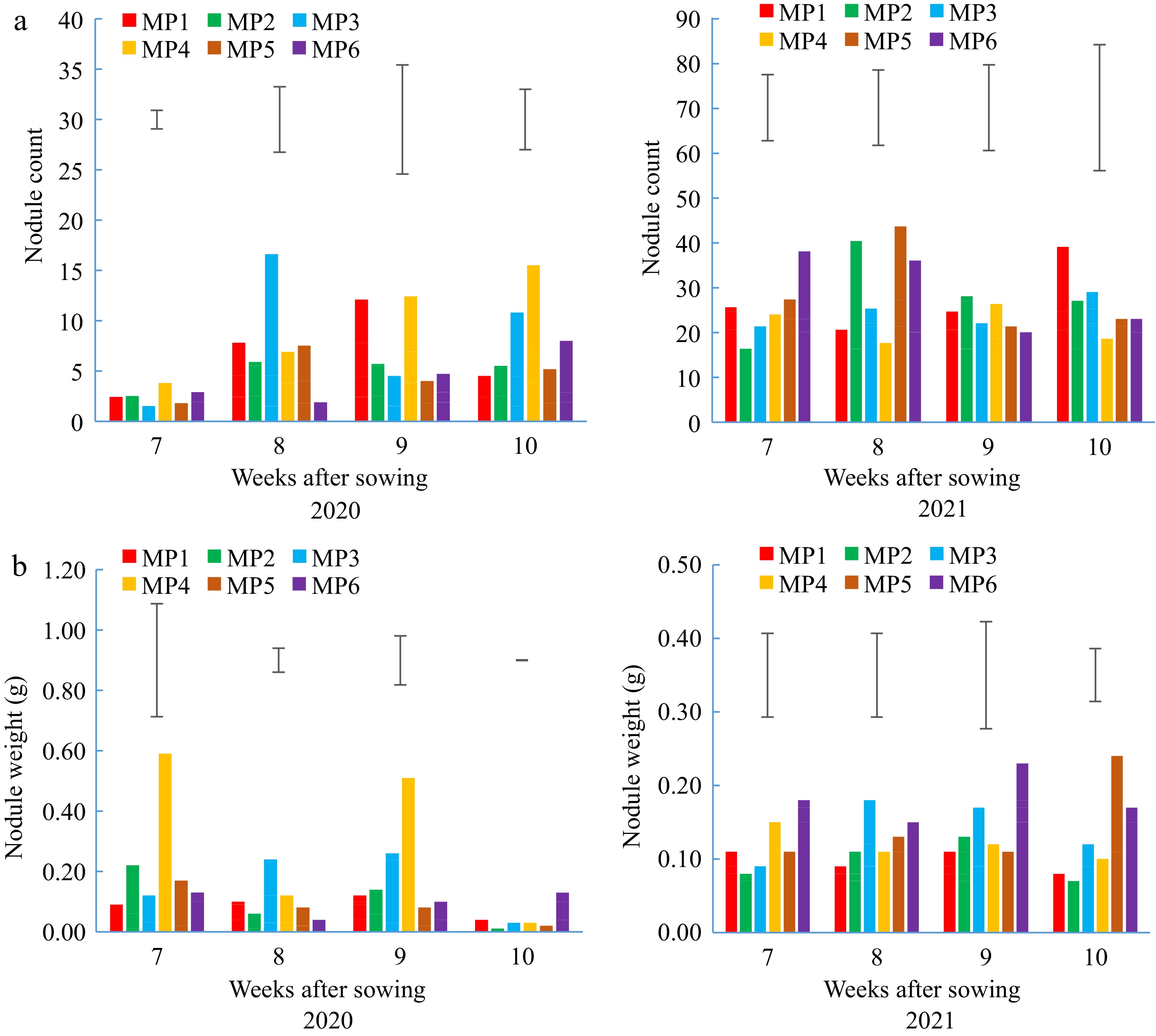
Figure 2.
(a) Nodule count and (b) nodule weight of soybean in 2020 and 2021; MP1, Control; MP2 NoduMax; MP3, Organic fertilizer; MP4, Nodumax and Organic fertilizer; MP5, Conventional system; MP6, Nodumax and Conventional system at 7, 8, 9 and 10 weeks after sowing, WAS. Capped vertical lines represent SED (df + 10).
Effects of management practices on some agronomic traits of soybeans in 2020 and 2021
-
Management practice significantly (p < 0.05) affected plant height at R7 during the drier year 2020 (Table 5) and no significant effect was recorded in 2021(Table 6). In 2020, organic fertilizer application alone (MP3) and in combination with inoculation (MP4) resulted in soybean plants that were significantly (p < 0.05) taller than soybean plants grown under conventional system (MP5). The number of branches per plant of soybeans was significantly (p < 0.05) affected by management practice in both years with conventional practice (MP5) and Conventional practice + inoculation (MP6) recording the highest number of branches in 2020 and 2021, respectively. Number of pods and seeds per plant and weight of seeds per plant were significantly (p < 0.05) affected by management practice in this study in both years, except number of pods per plant in 2021. Pod weight significantly (p < 0.05) increased following the application of organic fertilizer (PM3) in 2020 relative to the pod weight recorded under the control (PM1) and NoduMax (PM2) alone (Table 5). Inoculation with NoduMax alone (MP2) and in combination with organic fertilizer (MP4) enhanced pod weight in 2021 though not significantly (Table 6). Soybean grain yield under all the management practices where the seeds were either inoculated alone (MP2) or in combination with organic fertilizer (MP4) or conventional practice (MP6) and the conventional practice (MP5) was significantly (p < 0.05) greater than the yield from the control (MP1) in both years (Tables 5 & 6). Inoculation alone (MP2) failed to significantly enhance soybean grain yield when compared with other management practices (MP3–MP5) in 2020. A 15.1% yield advantage of soybean yield under organic practice (MP3) over the conventional practice (MP5) in the relatively dry year 2020.
-
Varying response of soybean to the different management practices in this study could partly be attributed to the varying weather conditions during the periods of experimentation. As shown in Fig. 1, rainfall distribution recorded in 2020 (493.8 mm) and 2021 (645.6 mm) compared well with the range of 500–750 mm recommended as adequate for a good seed yield of soybeans[34]. Although, the month of August in 2020 was markedly dry, rainfall was highest (246.3 mm) in September and adequate in October (127.6 mm). Rainfall distribution favoured soybean development during September and October that coincided with the critical periods (flowering – grain filling) of soybean for moisture requirement.
Attainment of full bloom (R2) was delayed following the application of inoculant (MP2), inoculant + organic fertilizer (MP4), synthetic fertilizers (MP5) and inoculant + synthetic fertilizers (MP6) during the dry year 2020. Application nitrogen has been reported to promote vegetative growth and thereby induce flowering in soybeans grown in Sudan[35]. However, in our study, the slight delay in days to R2 following the addition of nutrients could be attributed to more nutrient availability in the soil medium relative to the control. Lack of significant effect following the application of organic nutrients from different sources on days to R2, R5 and R7 of soybeans was recently reported in the humid tropics[9]. Grain Filling Period is usually determined by the interaction between genotype and environmental factors through their effects on the availability and translocation of assimilates from the leaves to the seeds as described by Patterson & Raper[36]. Application of inoculant alone (MP2) and organic fertilizer (MP3), and in combination with organic fertilizer (MP4) did not result in increased grain yield relative to the conventional system (MP5) despite prolonging GFP by 5–7 d. This finding confirmed earlier report that GFP has no direct effect on final grain yield of soybeans[37].
Application of inoculant and organic fertilizer (MP4) enhanced nodule count and weight during the dryer late cropping season of 2020 at 9WAS. This could be attributed to the ability of the organic fertilizer to improve water retention through the organic matter in it and also improve dissolution of some nutrients such as phosphorus as earlier reported by Olupot et al.[38]. These factors might have jointly improved the soil rhizosphere to make it more conducive for nodulation and plant growth[39]. Inoculation has also been reported to increase nodulation, plant growth and grain yield of soybeans in South Africa[40].
Soybean plant height is an important agronomic trait and it depends largely on the genetic makeup of the variety[41] and nitrogen availability[42]. Lack of significant effect of organic fertilizer application on soybean plant height was also recently reported in Indonesia[43]. In our study, combination of inoculant and organic fertilizer (MP4) enhanced plant height, especially during the more favourable year 2021. Similar increase in plant height in earlier studies have been attributed to the slow release of nutrients from the organic fertilizer and improved microbial activities in the soil[44] and enhanced nutrient uptake by plants and increased photosynthetic rate[45]. Application of synthetic fertilizer and inoculant (MP6) resulted in increased number of branches in this study. Increase in number of branches per plant had earlier been reported in soybean following the integrated use of organic and inorganic fertilizers in Nigeria[46]. Number of pods and seeds per plant and weight of seeds per plant have been reported as the major seed yield components of soybeans[47,48]. Increase in the values of these traits, especially seed weight per plant following the application of organic fertilizer and under conventional practice could be attributed to improved supply and availability of nutrients necessary for partitioning of assimilates to the various structural components of the plants under these management practices, especially in 2022. Similar results have been reported following the integrated use of farm yard manure and synthetic fertilizer on soybeans in Iran[49]. It was recently reported in a four year study that soybeans under organic production system produced fewer pods per plant and greater seed weight than soybeans under conventional system[50] . This trend was only consistent with our findings during the more favourable year 2021 though their differences (MP3 vs MP5) were not significant in both years. Similarly, earlier reports had also reported improvement in soybean pod weight following the application of Rhizobium japonicum alone in combination with phosphorus fertilizer[39]. The grain yield performance of the test variety in this study under different management practices (846.0–1,962.0 kg/ha in 2020) and (754.8–1,554.0 kg/ha in 2021) compared well with the average yield values in Nigeria (926.9 kg/ha) and Africa (1,348.0 kg/ha), and significantly below the world (2,784.2 kg/ha) based on FAOSTAT[6]. In our study, inoculation alone (MP2) did not significantly increase soybean grain yield more than the other management practices (MP3–MP5). Similar results were reported for soybeans in Pakistan[21]. This suggests that beyond microbial activities in the soil, other growth factors like nutrients and moisture availability are also crucial in the output any crop. The 15.1% yield advantage under organic system (MP4) over the conventional system (MP5) in 2020 confirmed earlier report that yields from organic plots tend to be greater than the conventional plots under dry conditions because of the inherent advantage of better water holding capacity under the organic system compared to the conventional system[13]. The recorded range of yield penalty is similar to values (15%–40%) reported in literature[11−13]. However, during the dryer year 2020, a very small yield penalty (1.0%) was recorded when the conventional system (MP5) was compared with the combination of organic fertilizer and NoduMax (MP4), while soybean that received only organic fertilizer application (MP3) recorded grain yield that was 15.1% superior to the yield under conventional system (MP5). The recorded trend confirmed the potential of organic soybean production in the tropics even under erratic rainfall distribution.
-
The results of this study have revealed that prospective soybean growers in the humid tropics can cultivate organic soybean using organic fertilizer with (MP4) or without (MP3) seed inoculation and still record comparable grain yield obtainable from the conventional management system especially when the rainfall amount and distribution are optimum. These two management practices (MP3 and MP4) can be recommended for resource-constrained farmers in the tropics with limited capacity to afford the high cost of synthetic fertilizers that have access to organic fertilizers or other organic soil amendments.
-
The authors confirm contribution to the paper as follows: conceptualization: Olowe VIO; methodology: Olowe VIO, Odueme PU, Fadeyi OJ; data collection: Olowe VIO, Odueme PU, Somefun OT; data analysis: Olowe VIO; draft manuscript preparation: Olowe VIO; manuscript review and editing: Odueme PU, Fadeyi OJ, Somefun OT; field operations: Fadeyi OJ. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
This is to sincerely express our profound gratitude to the technical staff of the Institute of Food Security, Environmental Resources and Agricultural Research (IFSERAR) for their support and assistance in preparing the land for the experiment and the management of the Federal University of Agriculture, Abeokuta (FUNAAB) for providing all the farm equipment and implements used for the research.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Olowe VIO, Odueme PU, Fadeyi OJ, Somefun OT. 2023. Agronomic response of soybeans (Glycine max (L.) Merrill) to different management practices in the humid tropics. Technology in Agronomy 3:18 doi: 10.48130/TIA-2023-0018 

Agronomic response of soybeans (Glycine max (L.) Merrill) to different management practices in the humid tropics
- Received: 18 August 2023
- Accepted: 21 November 2023
- Published online: 15 December 2023
Abstract: Organic soybean (Glycine max (L.) Merrill) remains an under exploited global market niche by farmers in the tropics. Therefore, a study was conducted in two consecutive years to determine the agronomic performance of soybean under different management systems in the humid tropics. The study was carried out during the late cropping season (July–Nov.) in 2020 and 2021 on the organic research plots of the Institute of Food Security, Environmental Resources and Agricultural Research, Nigeria. The six management practices (MP) evaluated were MP1 – Control, MP2 – NoduMax, MP3 – Organic fertilizer, MP4 – NoduMax + Organic fertilizer, MP5 – Conventional, MP6 – Conventional + NoduMax in a Randomized Complete Block Design and replicated three times. Data were collected on phenology, nodulation, grain yield and yield attributes of soybeans. Number of days to full bloom (R2) and physiological maturity (R7) were significantly (p < 0.05) affected by MP in both years, except R7 in 2021. Inoculation with NoduMax (MP2) enhanced nodule number at 8 and 9 weeks after sowing (WAS) during the wetter year 2021. Number of branches and seeds per plant, seed weight per plant and grain yield were significantly (p < 0.05) affected by MP in both years. Soybean grain yield recorded under MP3 and MP4 ranged between 1,198.8–1,962.0 kg/ha in both years and compared well with the average yield values in Nigeria (926.9 kg/ha) and Africa (1,348.0 kg/ha), and significantly below the world (2,784.2 kg/ha). Therefore, MP3 and MP4 can be recommended for potential resource-constrained organic soybean farmers in the humid tropics.
-
Key words:
- Conventional system /
- Management practice /
- NoduMax /
- Organic fertilizer /
- Soybean yield