









-
β-galactosidase (BGAL; EC 3.2.1.23) is one of the oldest ubiquitous enzymes and exists widely in higher plants. BGAL catalyzes the hydrolysis of polysaccharides, glycoproteins and terminal non-reducing β-D-galactosyl residues[1]. BGALs with this hydrolytic activity exist in the glycoside hydrolase (GH) family (GH1/2/3/35/42/50/59), and plants BGAL belongs to the GH35 family[2,3]. In plants, BGALs can be divided into two types based on their substrate specificities: Class I is an enzyme consisting of exo-β-(1-4)-galactanases that acts specifically on pectin β-(1-4)-D galactan. Class II have no hydrolytic activity against β-(1-4)-galactan and acts on the β-(1-3) and β-(1-6)-galactosyl linkages of arabinogalactan proteins[4]. BGALs participate in a variety of biological processes, particularly playing a privotal role in seed germination, cell-growth and fruit maturation[5,6].
Multigene family of BGALs has been identified in a variety of plant species, including Arabidopsis[7], rice[8], tomato[9], and sweet potato[10]. Plant BGALs were widely involved in the modification of cell wall structure and intercellular connections[11,12]. In Arabidopsis, BGALs may participate in cell wall metabolism in many processes[13]. AtBGAL1/2/3/4 were involved in growth, AtBGAL12 was involved in differentiation and AtBGAL5 was involved in hairy root development. Moreover, all AtBGAL transcripts occur during the initial stage of development[14]. In tomato, 17 putative BGAL encoding isoenzymes have been identified, which have different physiological functions and subcellular localization. At least seven of these genes contributed to the total β-GAL enzyme activity, and the transcription of corresponding genes differed significantly between early and late ripening of tomato fruits[15]. In chickpeas, the expression of CanBGal-5 was associated with growth and maturation stages with high cell division rate, while CanBGal-1 and CanBGal-4 were closely associated with late epicotyl growth[16]. Moreover, the BGAL family was also regulated by biological and abiotic stresses[17]. Salt or pathogen stress induced Atbgal1 expression in Arabidopsis[18], but BGAL transcription levels in cowpea were decreased under salt treatment[19]. The transcription level of BGAL in peaches was also inhibited when subjected to water stress[20].
Longan (Dimocarpus longan Lour.) is a tropical/subtropical evergreen fruit tree belonging to the Sapindaceae Dimocarpus. Longan is an important economic crop, and its yield and fruit quality were closely related to the development of longan embryos[21]. The difficulty of sampling the early longan zygote embryo greatly limits the application of longan embryo in genetic breeding. The longan somatic embryogenesis (SE) system has the characteristics of genetic stability and high regeneration frequency, which is an ideal material for studying the cell biology and regulation mechanism of embryogenesis[22]. Chromatin accessibility reflects the contact degree of chromatin DNA with cis-acting elements and trans-acting factors, while assays for transposase-accessible chromatin sequencing (ATAC-seq) is a sequencing method to capture chromatin accessibility[23]. The study of chromatin accessibility is important for exploring the regulatory network of transcription factors (TFs) and the role of gene expression in plant growth, development and stress resistance[24,25]. Chromatin modification affects chromatin accessibility, which determines the functional state of cells[26]. The monomethylation of histone H3K4 site can be divided into three types: H3K4me1, H3K4me2 and H3K4me3, among which H3K4me1 is usually related to the enhancer[27]. H3K4me1 exhibits highly dynamic changes and is closely related to the cell type specific gene expression profile[28]. In a previous study, Chen et al.[29] revealed the gene regulatory network of cell wall thickening during early longan SE through chromatin accessibility, H3K4me1 modification, and transcriptional analysis. Modification of cell wall components has been validated as key to initiation of cellular responses related to cell fate and development. As a dynamic and complex structure, cell wall plays an important role in regulating plant growth and development, intercellular communication and defense, and determining cell shape and fate[30]. Previous studies have reported that some cell wall components may be markers of cell fate transition, including inducing somatic embryogenesis[31] and postembryonic growth[32]. The BGAL family specifically acts on the degradation of pectin, xyloglucan, or arabinogalactose proteins in cell walls, and participates in cell wall metabolism by catalyzing galactose metabolism in the large and complex side chain of the cell wall[33]. During plant maturation, BGALs play a significant role in cell wall expansion, degradation and signaling molecule conduction[5,6]. Therefore, investigating of the functional significance of BGAL associated cell wall modifications in different cell types and processes is necessary.
In this study, we identified 20 DlBGAL genes and analyzed their phylogenetic relationships, conserved motif composition, gene structure and cis-acting elements. To determine the role of DlBGAL genes in the response to stresses, we evaluated their response to different temperature treatments. DlBGAL9 was screened by multi-omics analysis and its transcription factors (TFs) regulatory network was predicted. Using luciferase assays and transient expression of DlAGL61/80 in longan, we investigated the regulatory effect of DlAGL61/80 on DlBGAL9. The regulation of cell wall thickening by DlAGL80 and DlBGAL9 was investigated using longan hairy root system. The results are expected to expand our understanding of the BGAL family and further reveal the mechanism of TF-mediated DlBGAL9 gene during early longan SE.
-
The simultaneous embryogenic culture of longan 'Honghezi' in early development consisted of embryogenic callus (EC), incomplete compact pro-embryogenic culture (ICpEC), and globular embryo (GE), which were obtained by adding 2,4-D with 1.0, 0.5 and 0.1 mg/L, respectively[22]. For heat stress (35 °C), the longan EC was transferred to Murashige and Skoog (MS) medium and incubated in dark at 25 and 35 °C, respectively. The materials were collected at 6, 9, and 12 d of treatment, frozen with liquid nitrogen and stored at −80 °C until use.
Identification of the DlBGAL gene family
-
The longan 'Honghezi' genome database was obtained from the National Center for Biotechnology Information (NCBI) database (PRJNA792504). We conducted systematic BLASTP analysis of longan whole-genome sequences using Arabidopsis BGAL protein sequences (
www.arabidopsis.org ) in BioEdit software, and confirmed whether the candidate DlBGAL family members had a complete GH35 (PF01301) domain using HMMER[34] (www.ebi.ac.uk/Tools/hmmer/ ). A total of 20 candidate longan BGAL family members were screened. Blast comparison was performed on the NCBI online website (https://blast.ncbi.nlm.nih.gov/Blast.cgi ), and members of the DlBGAL family were named with reference to Citrus. When multiple DlBGALs matched to the same Citrus member, the name was marked -1/2/3/4, according to their physical location (from top to bottom) on chromosomes. The Expasy Protparam (https://web.expasy.org/protparam/ ) was used to analyze the basic physicochemical properties of the DlBGAL protein.Phylogenetic, gene structure, and conserved motifs analysis of the DlBGAL gene family
-
Using the BGAL gene protein sequences of longan, Arabidopsis, rice and citrus, a phylogenetic tree was constructed in MEGA 11 (v. 11.0.13) by conducting 1,000 bootstrap replicates with the neighbor-joining (NJ) method[35]. The protein sequences of the Arabidopsis, rice and citrus dates were downloaded from the Ensembl Plants (
http://plants.ensembl.org/index.html ). The BGAL gene structures were constructed by longan GFF and genome files. Conserved motif analysis of DlBGAL proteins was performed with Multiple Em for Motif Elicitation (MEME) (https://meme-suite.org/meme/tools/meme ) using the default parameters. The gene structure and conserved motifs were visualized with TBtools software (Visualize Gene Structure).Chromosomal localization, synteny and cis-acting regulatory elements analysis
-
The results of chromosome localization information provided by longan GFF file were visualized by Tbtools. MCScanX[36] was used to identify syntenic gene pairs, and BLASTP results and gene location information were used for further input. Tbtools[37] was used to identify proximal, dispersed, tandem, and segmental/whole-genome duplications of DlBGALs (Circle gene view). We used PlantCARE (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) to predict the cis-acting elements of DlBGALs.Analysis of the DlBGAL gene family by RNA-seq, ATAC-seq, and ChIP-seq
-
RNA sequencing (RNA-seq), assay for transposase-accessible chromatin sequencing (ATAC-seq), and chromatin immunoprecipitation sequencing (ChIP-seq) assays during early longan SE were available in the NCBI under accession code PRJNA792504. Different temperature treatments were available in the NCBI SRA repository under accession code PRJNA889670.
Dual-luciferase reporter assays in N. benthamiana leaves
-
The DlBGAL9 promoter (2,000 bp upstream of ATG) was cloned into pGreenII 0800-LUC to produce the reporter construct, while the TFs DlAGL61_Dlo006037 and DlAGL80_Dlo004817 were cloned into the CaMV35S vector (CaMV35S-DlAGL61/80) as the effector. Primers are listed in Supplemental Table S1. The effector and reporter constructs were mediated into Agrobacterium into Nicotiana benthamiana (N. benthamiana) by agroinfiltration.
After two days of cultivation, the LUC fluorescence signal was detected using a chemiluminescence imaging system (Gelview 6000Pro II, BLT, China), and the transcriptional activity of the TFs DlAGL61/80 was determined with a Dual-Luciferase Reporter Gene Assay Kit (Yeasen, Shanghai, China) and a Multiskan Spectrum microplate spectrophotometer (BioTek, CA, USA). At least three biological replicates were performed.
Determination of β-galactosidase activity, pectin content and ROS accumulation
-
β-galactosidase activity was determined with the β-galactosidase kit (Comin Biotechnology, Suzhou, China). Briefly, 100 mg sample was ground with 1 mL extract for ice bath homogenization. After 4 °C centrifugation (15,000 rcf·min−1 for 10 min), measurement and control were set respectively. Measurement: 10 µL sample mixed 25 µL reagent I and 35 µL reagent II. Control: 10 µL sample mixed 25 µL ddH2O and reagent II. The prepared solution allowed to stand for 30 min at 37 °C before 130 µL Reagent III was added, and the absorbance at 400 nm was determined.
Pectin content was determined with the total pectin content kit (Comin Biotechnology, Suzhou, China). 0.05 g of sample was ground and 1 ml of extract solution I added. The mixed sample was placed at 90 °C for 30 min, and centrifuged at 5,000 g and 25 °C for 10 min after cooling. The supernatant was removed and 1 mL of extract solution I was added to the precipitation, repeating the operation once. After removing the supernatant, 1 mL of extract solution II was added and hydrolyzed at 90 °C for 1 h. After cooling, the sample was centrifuged at 8,000 g and 25 ° C for 15 min, and the supernatant was taken for testing. A 30 µl sample was taken and 180 µl concentrated sulfuric acid was added at 90 °C for 10 min, removed and cooled. Thirty µl of reagent III was taken and left to stand at 25 °C for 30 min, then added 60 µl of distilled water. The absorbance at 530 nm was determined.
Reactive oxygen species (ROS) accumulation was determined with the ROS test kit (Comin Biotechnology, Suzhou, China). Briefly, 0.1 g sample was ground with 1 mL reagent I and 10 µL reagent III for ice bath homogenization. After 4 °C centrifugation (600 rcf·min−1 for 5 min), the supernatant was extracted. After 4 °C centrifugation (11,000 rcf·min−1 for 10 min), the supernatant was removed and mixed with reagent II. Measurement and control were set respectively. Measurement: 20 µL sample mixed with 50 µL reagent IV, 50 µL reagent V, 50 µL reagent VI and 30 µL reagent VII. Control: 20 µL reagent II mixed 50 µL reagent IV, 50 µL reagent V, 50 µL reagent VI and 30 µL reagent VII. The mixture was placed in darkness at 37 °C for 30 min, and the absorbance values were measured at the excitation wavelength of 499 nm and the emission wavelength of 521 nm within 10 min.
Transformation of the longan embryogenic callus and hairy root
-
The genetic transformation of longan EC was mainly through freeze-thaw transformation of recombinant plasmid into Agrobacterium receptor state (GV3101), and transferred of transformed Agrobacterium into LB liquid medium containing antibiotics 100 mg·L−1 Kan and 25 mg·L−1 Rif. The culture was incubated for 18 h at a speed of 200 rcf·min−1 under constant temperature and darkness at 28 °C. Centrifuge at 6,000 rpm for 10 min before removing the supernatant. Added MS liquid medium (including 50 μmol·L−1 AS and 10 μmol·L−1 MgCl2) to resuspend the remaining bacterial solution, and adjusted the bacterial solution OD600 to 0.6−0.8. The cultured longan EC was transferred for about 15 d into the resuspension, incubated for 30 min, and transferred the filtered longan EC to MS solid medium.
According to the method of Meng et al.[38], the longan seedlings were infected with Agrobacterium rhiza. The lateral roots of longan seedlings were removed, and the culture temperature was 26 °C. The seedlings were cultured alternately under light (16 h) and darkness (8 h) for 25 d. A single gene set 100 replicates.
Histological analysis
-
The hairy roots of longan were cut into small segments and then fixed with FAA [10% formalin, 5% acetic acid, 50% ethanol (v/v)] at ordinary temperature for 48 h. According to the methods of Yang et al.[39], the specimens were dehydrated and infiltrated in the ethanol and paraffin series. After being embedded in paraffin, the samples were cut into semi-thin (1 μm) sections using a microtome (Leica RM2125 RTS, Germany) and stained with 0.1% toluidine blue. Finally, the sections were observed under a microscope (Leica DM2500, Germany).
RNA extraction and qRT-PCR
-
Total RNA was extracted from longan SE and hairy root using TransZol Up reagent (TransGen, China) and RNAprep Pure Plant Kit (TIANGEN, China), respectively. The cDNA synthesis was performed using the SMART™ RACEcDNA Amplification Kit TransScript RNA First-Strand cDNA Synthesis SuperMix (YEASEN, China). QRT-PCR detection was performed on the Roche Light Cycler 96 instrument, using 10x diluted cDNA as the amplification template. Beta-actin (ACTB) and ELF-4α as internal reference genes for transgenic longan EC and hairy root, respectively. The relative expression was calculated by the 2−ΔΔCᴛ method[40], and significant differences were assessed by Duncan test (Student’s t-test, *p < 0.05, **p < 0.01). The data were imported into GraphPad Prism 9 software to generate graphs. Primers are listed in Supplemental Table S1.
-
To identify the BGAL genes in longan, we scanned the D. longan genome database using the known BGAL members in Arabidopsis by BLASTP and TBLASTN. After the removal of incomplete and redundant sequences, a total of 20 putative BGAL genes were identified in the D. longan genome. For the annotation of 20 DlBGAL genes, the Citrus nomenclature system was pursued with numbers representing the highest sequence similarity with the corresponding CsBGAL orthologous (Table 1). The lengths of amino acids (aa) of 20 DlBGALs range from 444 aa (BGAL13-1) to 889 aa (BGAL9). The MW of DlBGALs varies from 50.15 kDa (BGAL13-1) to 99.79 kDa (BGAL9) with an average of 86.85 kDa. The pI ranged from 5.4 (BGAL8-1) to 9.4 (BGAL13-1). Except for BGAL16-1, the remaining DlBGALs were classified as stable proteins because their instability index was less than 40 (Table 1). The predicted aliphatic index ranged from 68.75 to 85.3 in DlBGAL proteins. Due to the GRAVY value (< 0), all DlBGAL proteins were predicted to be hydrophilic.
Table 1. Basic parameter analysis of DlBGAL family.
Gene ID Gene name Size (aa) Molecular weight/kD PI Instability coefficient Hydrophilicity Dlo000174 BGAL1-1 844 93,311.98 8.25 37.76 −0.287 Dlo000176 BGAL1-2 839 92,852.64 7.96 39.56 −0.267 Dlo000290 BGAL15-1 825 92,233.81 5.56 35.42 −0.444 Dlo002805 BGAL9 889 99,793.27 6.35 35.27 −0.271 Dlo006281 BGAL17 710 79,011.06 6.25 35.45 −0.211 Dlo006300 BGAL1-3 844 93,575.84 8.4 38.83 −0.196 Dlo013472 BGAL5 734 82,104.46 8.8 36.87 −0.276 Dlo014416 BGAL16-1 716 80,274.25 9.34 41.46 −0.262 Dlo014417 BGAL16-2 832 93,191.23 8.62 36.02 −0.416 Dlo015024 BGAL16-3 838 93,942.65 6.32 39.86 −0.208 Dlo015487 BGAL13-1 444 50,149.01 9.4 38.7 −0.233 Dlo015584 BGAL3-1 843 93,502.26 7.25 38.92 −0.221 Dlo016393 BGAL6 718 81,101.64 5.46 36.14 −0.226 Dlo016396 BGAL16-4 846 94,734.74 6.92 40.98 −0.33 Dlo021540 BGAL8-1 842 93,433.6 5.4 39.83 −0.16 Dlo023146 BGAL13-2 864 96,671.78 8.37 36.43 −0.214 Dlo023292 BGAL3-2 660 73,921.74 7.3 34.86 −0.29 Dlo026229 BGAL8-2 853 92,871.32 8.07 39.94 −0.192 Dlo026230 BGAL8-3 612 67,240.37 7.56 36.87 −0.325 Dlo029560 BGAL15-2 828 93,079.31 6.56 34.06 −0.305 The phylogenetic tree of the BGAL family contains BGAL gene members from four species: D. longan, O. sativa, C. clementina and A. thaliana. To determine their evolutionary relationship, a phylogenetic tree was constructed using the neighbour-joining (NJ) method and tested by 1,000 bootstrap replicates (Fig. 1). According to phylogenetic relationships, the BGALs were clustered into seven groups (Groups I–VII), each containing 4, 2, 6, 1, 3, 3 and 1 DlBGALs, respectively. In O. sativa, A. thaliana and C. clementina, BGAL genes had been identified in cluster forms. Thus, BGAL members may be evolutionarily close in their respective species. Interestingly, BGAL proteins of O. sativa and A. thaliana and C. clementina were clustered into distinct clades (Fig. 1), suggesting that BGAL proteins were evolved after monocots and dicots divergence. Among them, most of the BGAL genes in D. longan were closely clustered with C. clementina in the groups of the phylogenetic tree, supporting the hypothesis that D. longan and C. clementina shared close and similar ancestors[41].
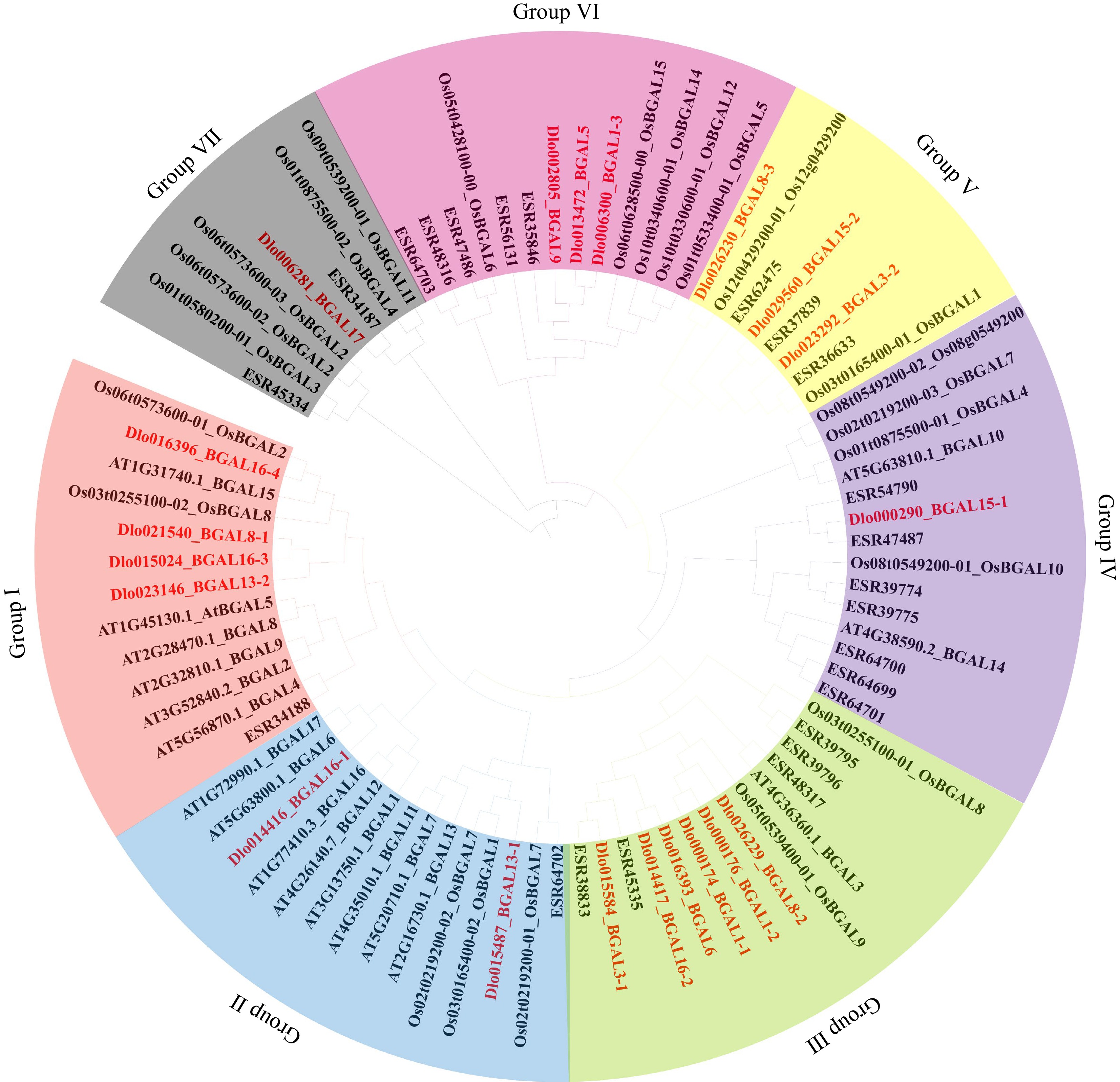
Figure 1.
Phylogenetic tree analysis of β-galactosidase (BGAL) proteins from longan, Citrus, Arabidopsis, and Oryza.
Gene structure and motif composition of longan BGAL genes
-
To further understand the evolutionary relationship of the 20 DlBGAL genes, we identified their structural characteristics and motifs (Supplemental Fig. S1). All DlBGAL genes had at least 12 exons, with 13 of the 20 genes having more than 18 exons (Supplemental Fig. S1b). The numbers of introns and exons in the genomic sequences were relatively similar in each group. In addition, ten conserved motifs of DlBGAL proteins were identified through MEME analysis (Supplemental Fig. S1c). Motifs 2 and 4, which were components of the DlBGAL domain, were found in all DlBGAL proteins. Motifs 6, 7 and 10 were detected in all DlBGAL proteins except DlBGAL13-1. For group III, IV and VI, all of the DlBGALs possessed 10 motifs (Supplemental Fig. S1c).
Chromosomal location and gene duplication events of DlBGAL genes
-
The chromosome information of DlBGAL genes was extracted and the chromosomal positions indicated that DlBGAL genes were distributed on eight of the 15 chromosomes of longan (Fig. 2a). Among them, Chr1, 6 and 7 had four DlBGAL genes. Chr3, 11 and 12 had two DlBGAL genes, while the Chr10 and 14 had the smallest numbers of DlBGAL genes (one gene each). The distribution of DlBGAL genes was uneven.
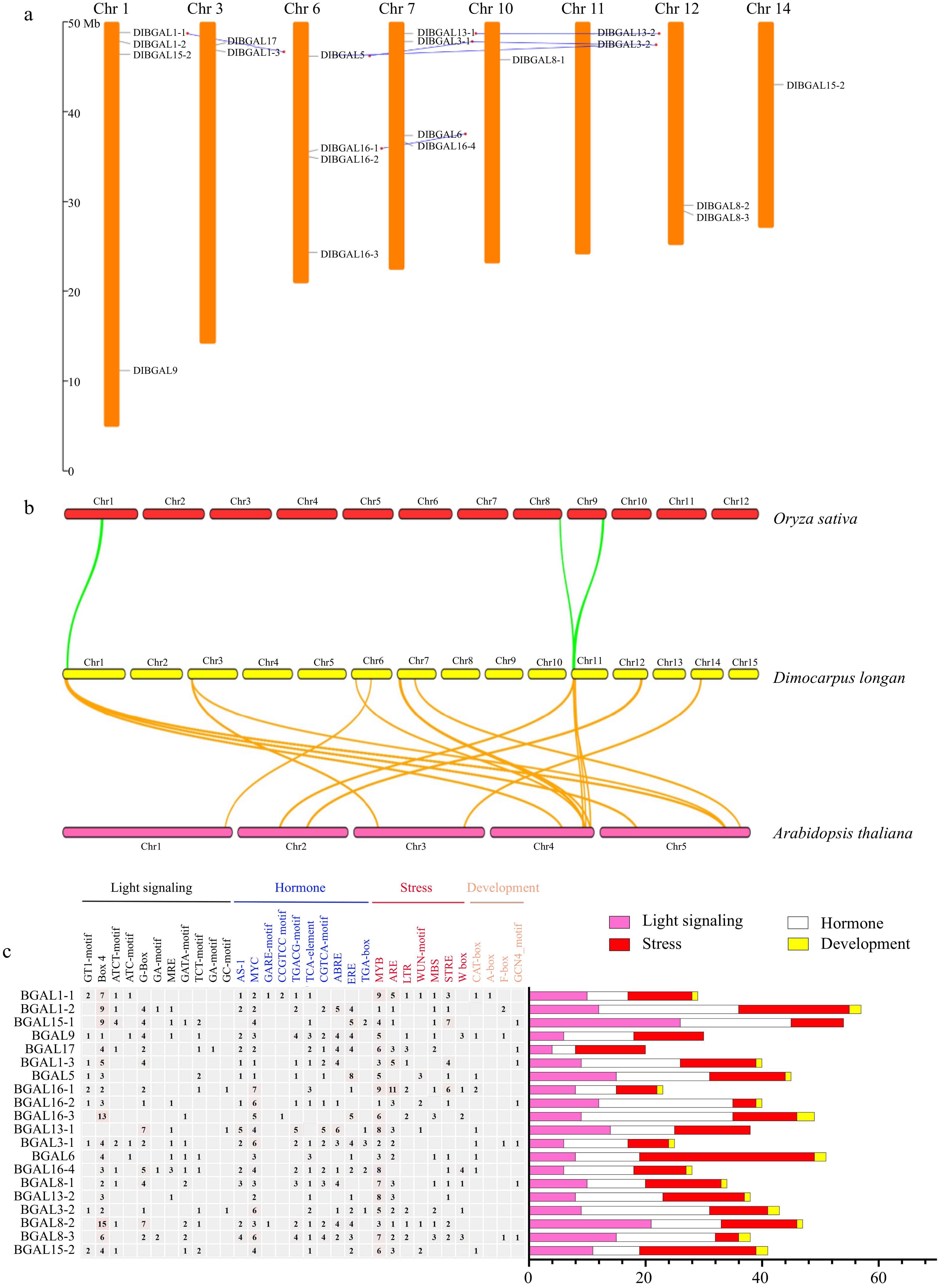
Figure 2.
Chromosomal distribution, synteny and cis-elements analysis of DlBGAL genes. (a) Distributions and duplications of DlBGAL genes along longan chromosomes. The gray and blue lines indicate chromosome locations of DlBGALs and segmentally duplicated genes, respectively. (b) The homologous relationships of BGAL genes in longan, Oryza and Arabidopsis. The green and orange lines indicate the homologous relationships of BGAL genes with Oryza and Arabidopsis, respectively. (c) Analysis of cis-elements in the promoter of DlBGAL genes. Heatmap of the numbers of cis-elements in the promoters of DlBGAL genes.
The results of collinearity analysis showed that there were six pairs of segmental duplication events in DlBGALs, corresponding to DlBGAL1-1/1-3, DlBGAL5/3-1, DlBGAL5/3-2, DlBGAL16-1/6, DlBGAL13-1/13-2 and DlBGAL3-1/3-2 (Fig. 2a). The paralogous gene pairs were distributed on different chromosomes, which may be beneficial for the evolution and expansion of the DlBGAL gene family. The expansion of the DlBGAL gene family was mainly due to segmental duplication, rather than tandem duplication. To verification the evolutionary rates and selective pressures between the DlBGAL genes and their duplicated genes, we used Ka (nonsynonymous), Ks (synonymous), and the Ka/Ks ratio for estimations. As shown in Supplemental Table S2, the Ka/Ks values of all the DlBGAL gene pairs were less than 1, indicating that the DlBGAL gene family underwent strong purifying selection. These results indicated that DlBGAL family was conservative during the domestication process of longan.
The collinearity analysis between longan and Arabidopsis thaliana or Oryza sativa showed that three DlBGAL homologue genes appeared in the three chromosomes of Oryza sativa, but 16 DlBGALs had corresponding paralogous genes on five chromosomes in Arabidopsis thaliana (Fig. 2b). The relationship between longan and Arabidopsis thaliana was closer than that between longan and Oryza sativa.
Analysis of cis-elements in the promoter region of DlBGAL genes
-
The cis-elements are involved in the regulation of gene expression, which are an important part of the response regulation network for plant growth and development, hormone response and stress response[42,43]. We isolated the 2-kb upstream promoter region of the DlBGAL genes to identify the potential cis-elements. Among them, many DlBGAL genes exhibit elements specifically related to light signaling, hormone, stress and development.
For DlBGAL genes, 34 main types of cis-elements were identified in their promoter sequences, which included 11 light responsive, 10 hormone responsive, seven stress responsive, and six development elements (Fig. 2c). The major light responsive elements were Box4 and G-box and the most common motifs were the JA-responsive elements (MYC), abscisic acid (ABA)-responsive element (ABRE), and ethylene-responsive element (ERE) of the scanned hormone responsive motifs. MYB and ARE were the most cis-elements of the stress responsive. These results suggested that DlBGAL genes were participated in complex light, hormone regulation and stress responsive networks that may involve multiple regulatory modalities.
Expression analysis of DlBGAL genes during early SE and under different temperature stress conditions
-
The expression profiles of DlBGAL genes during early SE and under temperature treatments were illustrated using our previous RNA-seq dataset. Results showed that nine DlBGALs (DlBGAL8-3/15-1/13-2/16-2/15-2/13-1/8-1/6/3-2) were almost not expressed (FPKM < 1) in the early SE RNA-seq dataset (Fig. 3a). About the remaining 11 genes, there were four genes (DlBGAL17/16-3/1-2/16-1) that showed high expression levels at EC stage and six genes (DlBGAL1-1/16-4/9/8-2/1-3/3-1) had high expression levels at GE stage. Furthermore, only DlBGAL5 was presented high expression at ICpEC stage. These results suggested that the DlBGAL genes may be involved in promoting early SE. The differentially expressed date of DlBGAL genes under different temperature conditions were derived from the RNA-seq (Fig. 3b). From the results, the expression levels of some DlBGAL genes varied obviously under high-temperature and low-temperature treatments. Compared to normal culture temperature (25 °C), seven DlBGAL genes (DlBGAL3-2/1-2/16-2/17/5/1-1/16-3) were upregulated under the high-temperature and two DlBGAL genes (DlBGAL5/1-1) were upregulated at low-temperatures.

Figure 3.
Expression profile and multi-omics analysis of DlBGAL genes. (a), (b) Expression analyses of DlBGAL genes during early longan SE and different temperature treatments. (c) Alluvial diagram of DlBGAL genes was divided into eight types (types I–VII and other) based on ATAC-seq, ChIP-seq, and RNA-seq data. The width of different colored bars represents the number of genes, and the genes included in each type were listed in the right panel. EC vs GE, the differential expression of EC vs GE; EC vs GE & ICpEC vs GE, the differential expression of EC vs GE and ICpEC vs GE; EC vs ICpEC vs GE, the differential expression of EC vs ICpEC, ICpEC vs GE and EC vs GE; ICpEC/GE, ATAC-seq or ChIP-seq (H3K4me1) peaks were detected in ICpEC/GE; EC & ICpEC, ATAC-seq or ChIP-seq (H3K4me1) peaks were detected in EC and ICpEC; EC & ICpEC & GE, ATAC-seq or ChIP-seq (H3K4me1) peaks were detected in EC, ICpEC and GE; Na, undetectable peak of ATAC-seq or ChIP-seq (H3K4me1); Ns, no difference in transcripts among the samples.
The change of chromatin accessibility affects the binding of transcription factors and other protein molecules to chromatin, thereby regulating gene transcription[44]. ATAC-seq analysis revealed that chromatin accessibility of other DlBGAL genes, except DlBGAL17 and DlBGAL8-3, was open during the early SE (Fig. 3c). Histone methylation modification can activate or inhibit gene transcription by the state of methylation, in which H3K4me1 is a marker associated with enhancers[45]. Except for DlBGAL6, H3K4me1 binding peaks were detected in other DlBGAL genes during early SE, and most of them had binding peaks at three stages. Combined with transcription, chromatin accessibility, and H3K4me1 modification results, most DlBGAL genes differentially expressed during early SE could be detected in the ATAC-seq and ChIP-seq data. In summary, multi-omics analysis has implications for the potential mechanism of BGAL family involvement in early SE.
Transcription factors target the DlBGAL9 promoters and activate its transcription
-
Transcription factors (TFs) can inhibit or enhance gene expression, and promote SE formation by directly activating early embryonic model genes[24]. To understand the regulatory network of transcription factors on DlBGAL genes, the upstream 2-kb sequences of DlBGAL genes were used as TF binding regions. In total, 11 DlBGALs were predicted as targets for 10 TFs (Fig. 4a). Based on multi-omics analysis, the functional role of DlBGAL9 in early longan SE attracted further interest. Previous studies have identified an important somatic embryo marker gene, Dlo004817_AGL80, which was involved the regulatory network associated with the cell wall of longan[29]. Based on TFs and DlBGALs regulatory network, Dlo006037_DlAGL61 and Dlo004817_DlAGL80 were selected as candidate TFs for regulating DlBGAL9 (Fig. 4a).
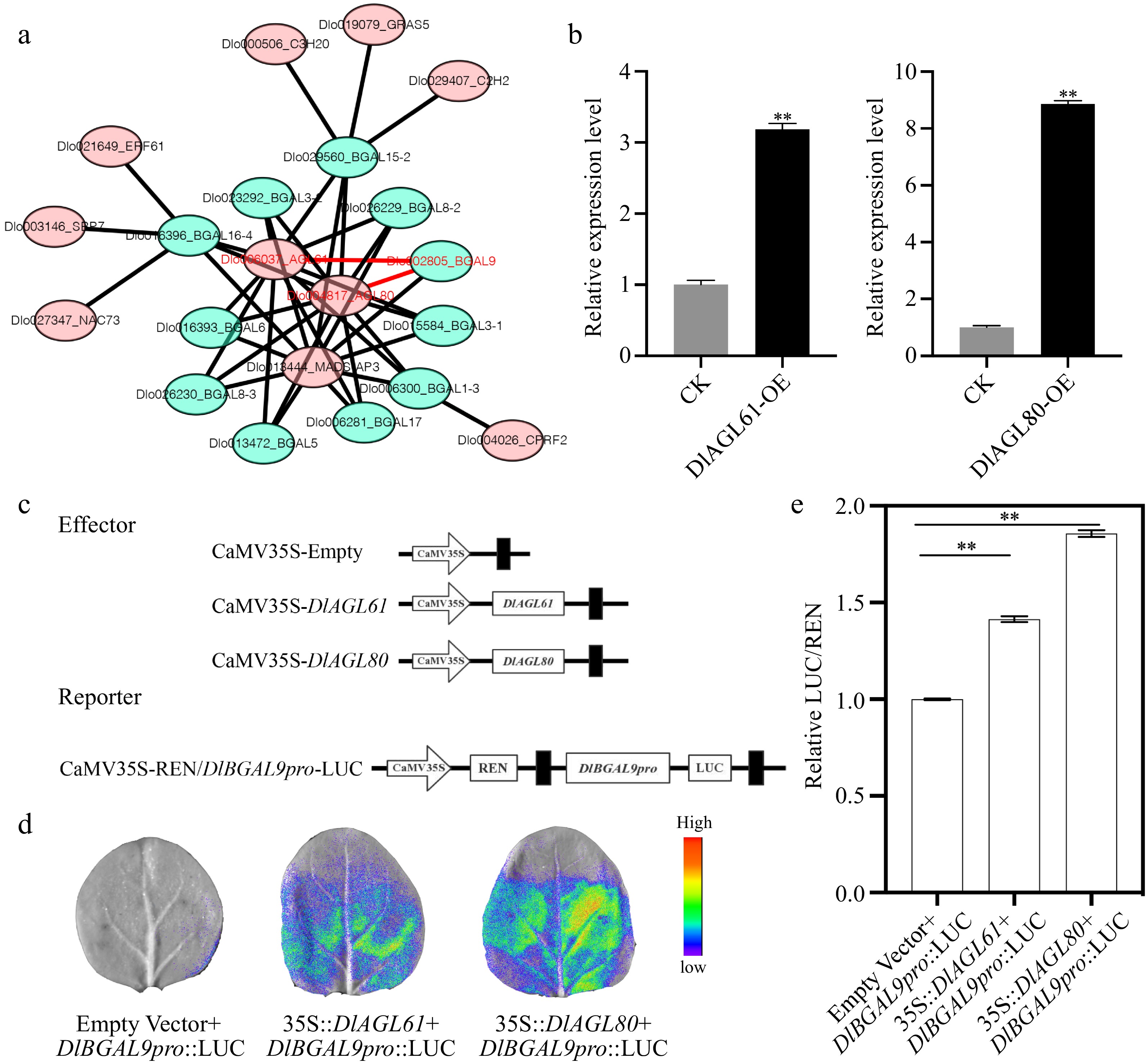
Figure 4.
Transcription factors (TFs) regulate the expression of DlBGAL9. (a) Network prediction of DlBGAL gene regulated by TFs. The red font represents the key genes. (b) The relative expression of DlBGAL9 in the TFs DlAGL61/80 transiently transformed. Beta-actin (ACTB) was used as a reference gene to normalize gene expression data. Each treatment was conducted in triplicate with three technical repeats. (c) Schematic diagrams of the effector vectors and reporter vectors in luciferase assays. (d) The interactions of DlAGL61 or DlAGL80 with the DlBGAL9 promoter that were detected in leaves of N. benthamiana through a dual luciferase reporter system. (e) Determination of the luciferase enzyme activity in N. benthamiana. Values were mean ± SD. **indicates a statistically significant difference, **p < 0.01.
To investigate the regulatory effect of DlAGL61/80 on DlBGAL9 transcription, we verified the expression of DlBGAL9 in longan EC that overexpressed TFs DlAGL61/80. QRT-PCR results showed that DlBGAL9 expression was significantly increased in longan EC with overexpressing of DlAGL61 (3.19-fold) and DlAGL80 (8.87-fold) (Fig. 4b). Thus, TFs DlAGL61/80 may enhance DlBGAL9 transcription by targeting its promoters. To further verify whether TFs DlAGL61/80 can activate the expression of DlBGAL9, a luciferase (LUC) reporter assay was performed in Nicotiana benthamiana leaves. Two constructs, 35S::DlAGL61 and 35S::DlAGL80 were used as effector, and DlBGAL9pro:LUC was used as reporter (Fig. 4c). Compared with control check (CK), the relative DlAGL61/80-induced firefly LUC/Renilla luciferase (REN) activity driven by the promoters of DlBGAL9 increased 1.41-fold and 1.86-fold, respectively (Fig. 4d, e). These results indicated that TFs DlAGL61/80 may target the DlBGAL9 promoters and activate their transcription.
Transcription factors and DlBGAL9 responses to heat stress and changes of β-galactosidase activity and pectin content during early longan somatic embryogenesis
-
Based on the RNA-seq data of longan EC under temperature treatments, DlBGAL genes expression detected in early longan SE responded to heat stress (Fig. 3b). Previous studies have found that heat stress affects the process of early longan SE, and EC cannot develop to GE under 35 °C treatment (heat stress). To further investigate the response of DlBGAL9 and TFs DlAGL61/80 to heat stress, we detected their expression of longan EC treated at 35 °C. The results showed that DlBGAL9 was significantly upregulated under heat stress at 6 d, and the TF DlAGL61 showed a significant downregulation trend under heat stress at 9 d (Fig. 5a). Notably, DlBGAL9 and TFs DlAGL61/80 have the highest level under heat stress at 12 d (Fig. 5a). The DlBGAL9 and TFs DlAGL61/80 showed a similar trend under heat stress, indicating that the DlAGL61/80-DlBGAL9 regulatory network may be involved in longan's response to heat stress.
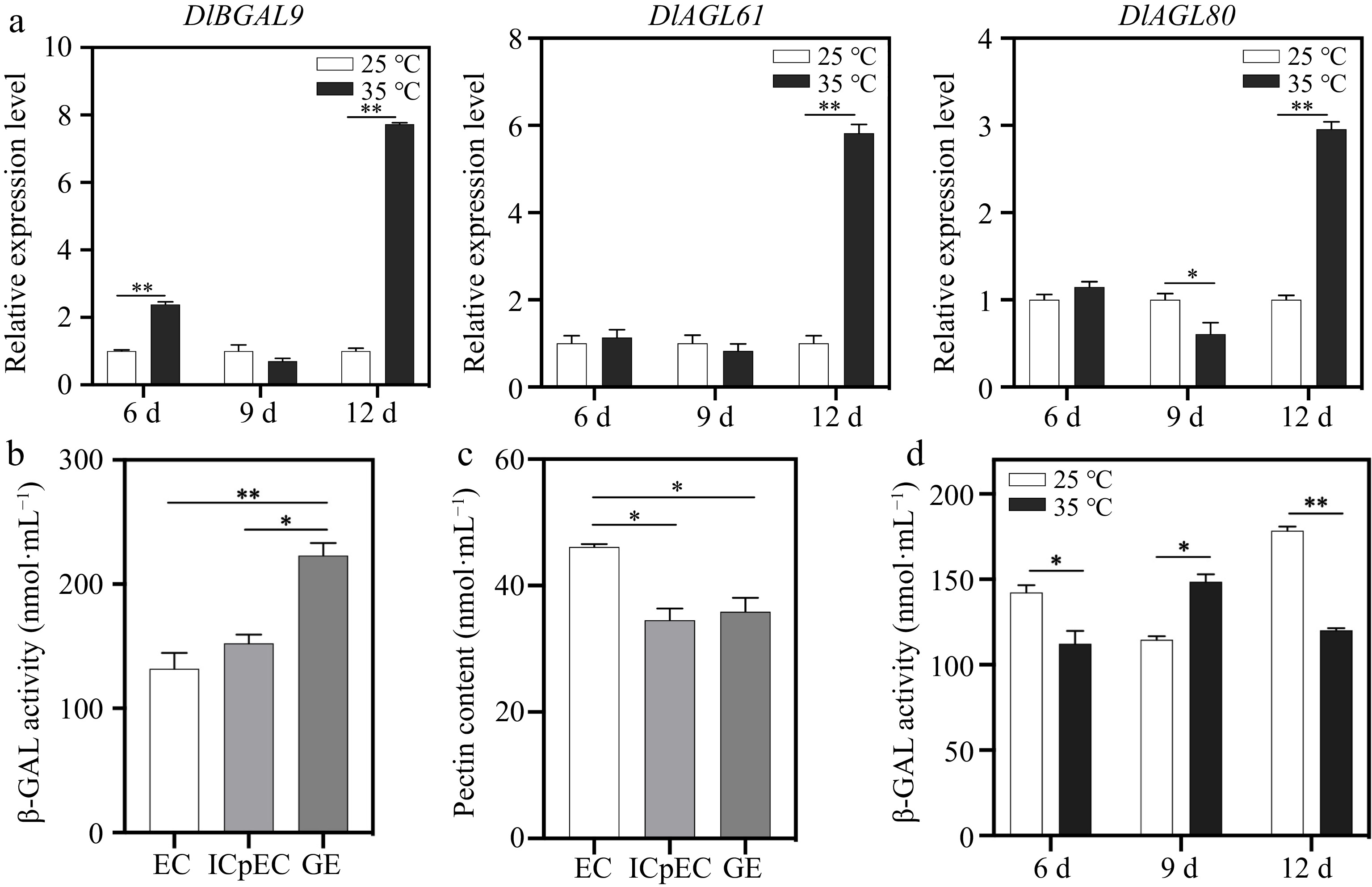
Figure 5.
Expression pattern analysis of DlBGAL9 and TFs DlAGL61/80 in longan EC under heat stress. (a) Expression of DlBGAL9 and TFs DlAGL61/80 in longan EC under different days of heat stress. Beta-actin (ACTB) was used as a reference gene to normalize gene expression data. (b) The β-GAL activity during early longan somatic embryogenesis (EC, ICpEC, and GE). (c) The pectin content during early longan somatic embryogenesis (EC, ICpEC, and GE). (d) The β-GAL activity in longan EC under different days of heat stress. Somatic embryos were cultured on MS medium and treated at 25 °C (normal) and 35 °C (heat stress). Samples were harvested after 6, 9, and 12 d to detect the gene expression patterns and β-GAL activity. MS, Murashige and Skoog medium. EC, embryogenic callus; ICpEC, incomplete compact pro-embryogenic culture; GE, globular embryo. Each treatment was conducted in triplicate with three technical repeats. Values are mean ± SD. * Indicates a statistically significant difference, *p < 0.05, **p < 0.01.
To further analyze the role of β-GAL and pectin during early longan SE, the changes in β-GAL activity and pectin content were measured (Fig. 5b, c). The β-GAL activity was significantly upregulated during early longan SE (Fig. 5b). Pectin content was opposite to β-GAL activity, and the highest pectin content was found in the EC stage (Fig. 5c). After that, the β-GAL activity of longan ECs treated at heat stress for different durations was determined. Compared with the normal temperature (25 °C), the β-GAL activity was significantly downregulated at 6 and 12 d of heat stress (35 °C) and significantly increased at 9 d (Fig. 5d). It was suggested that β-GAL can not only participate in somatic embryonic process by negatively regulating pectin content, but also can respond to heat stress.
Overexpression of DlBGAL9/DlAGL80 thickens the root cell wall by affecting β-GAL activity
-
As the embryonic callus is composed of parenchyma cells, it poses certain obstacles to the study of cell walls. Hairy roots have the characteristics of rapid growth and hereditary stability, providing a good material for studying the impact of genes on cell walls. In this study, the constructed p1302: DlBGAL9: GFP was introduced into the Agrobacterium rhizogenes strains and transferred into longan seedlings. The GFP-signals and PCR results both confirmed that DlBGAL9 had been transferred into longan seedlings and successfully induced the hairy roots (Fig. 6a). Compared with the wild-type (WT), transgenic roots overexpressing DlBGAL9 (DlBGAL9-OE) showed a 24.20-fold increase in DlBGAL9 expression (Fig. 6a). We examined the β-GAL activity and pectin content in the transgenic hairy roots (Fig. 6b, c). The β-GAL activity of DlBGAL9-OE root was higher, while the pectin content was lower. These results suggested that DlBGAL9 influences pectin content by regulating β-GAL activity in transgenic hairy roots.
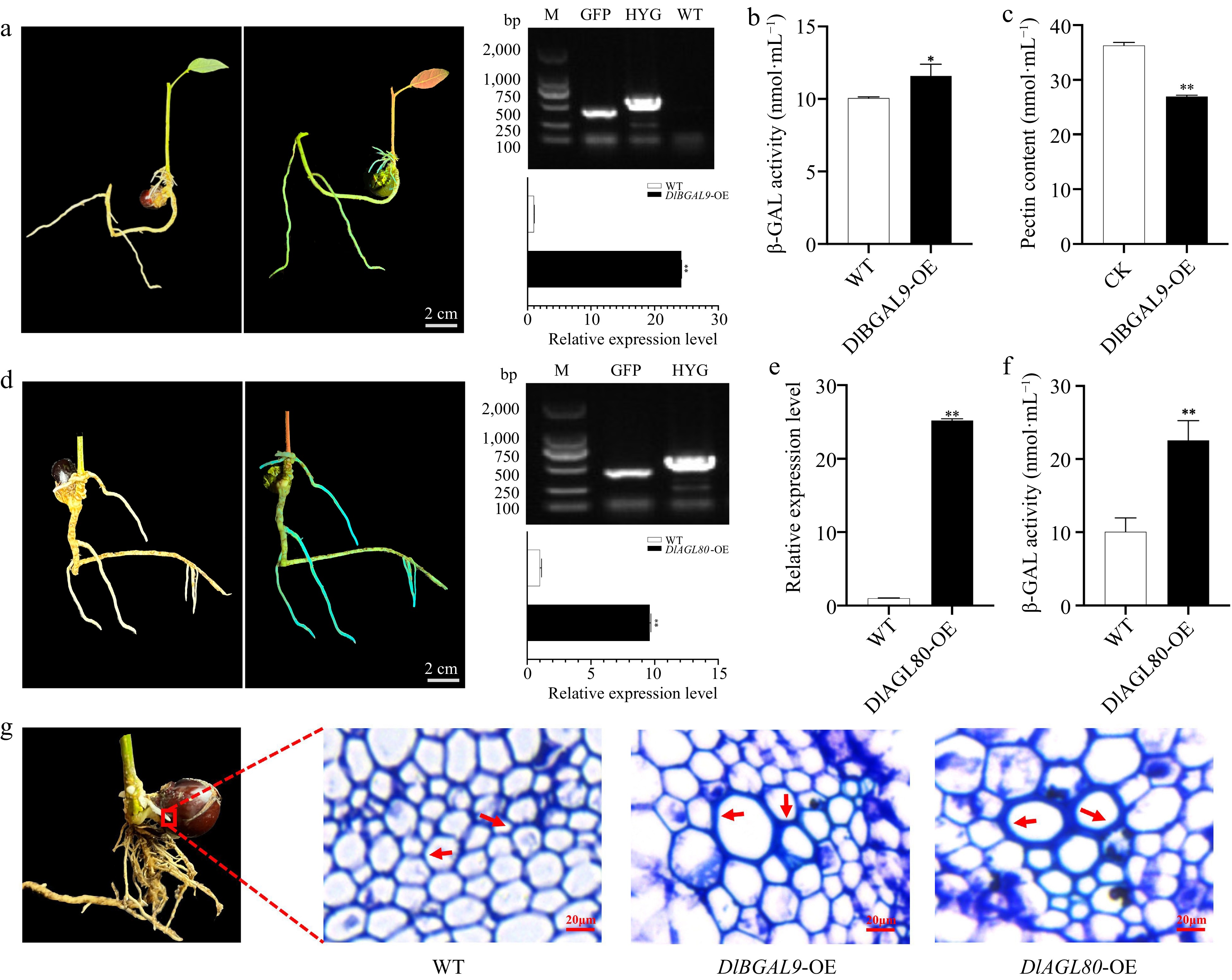
Figure 6.
Functional analysis of DlBGAL9 and TF DlAGL80 in longan hairy root. (a) Left: The bright field and GFP-fluorescent DlBGAL9-OE transgenic hairy roots. Right: Molecular identification DlBGAL9-OE transgenic hairy roots. Scale bar = 2 cm. (b) The β-GAL activity in longan hairy root of DlBGAL9-OE transgenic. (c) The pectin content in longan hairy root of DlBGAL9-OE transgenic. (d) Left: The bright field and GFP-fluorescent longan hairy root of the DlAGL80-OE transgenic. Right: Molecular identification longan hairy root of the DlAGL80-OE transgenic. Scale bar = 2 cm. (e) Expression of DlBGAL9 in longan hairy roots of DlAGL80-OE transgenic. (f) The β-GAL activity in longan hairy roots of DlAGL80-OE transgenic. (g) Anatomical and morphological observations of WT, DlBGAL9-OE and DlAGL80-OE longan hairy roots. Arrows indicate the position of cell wall thickening. Scale bar = 20 μm. (* p < 0.05, ** p < 0.01).
As DlAGL80 target the DlBGAL9 promoters and activate its transcription, we tested whether the expression of DlBGAL9 was affected by DlAGL80 transgenic roots. The overexpressing DlAGL80 (DLAGL80-OE) transgenic hairy roots was confirmed by GFP-signals and PCR results, and the expression of DlAGL80 increased by 9.61-fold (Fig. 6d). In DlAGL80-OE roots, DlBGAL9 expression was dramatically increased, while β-GAL activity was increased (Fig. 6e, f). Transverse sections of DlBGAL9 and DlAGL80 transgenic hairy roots showed thickening of cell walls in both genotypes compared to WT (Fig. 6g). In summary, DlAGL80 may regulate the expression of DlBGAL9 by regulating β-GAL activity and participate in the cell wall modification processes.
DlBGAL9 and TF DlAGL80 overexpression enhances the resistance to heat stress
-
The expression of DlBGAL9 and TF DlAGL80 in roots under heat stress were assessed to investigate whether DlBGAL9 and TF DlAGL80 were involved in the resistance of longan roots to heat stress. QRT-PCR results showed that heat stress induced DlBGAL9 and TF DlAGL80 expression, which reached the peak value at 6 d (Fig. 7b). Further results showed that the hairy root cell wall of longan was thickened under heat stress (Fig. 7a). The above results indicated that DlBGAL9 and TF DlAGL80 may participate in the mechanism of longan resistance to heat stress. When plants were subjected to biotic or abiotic stress, the concentration of reactive oxygen species (ROS) in cells increases and the homeostasis of cells was destroyed. The activity of ROS scavengers (SOD, POD and CAT) will increase accordingly to maintain the homeostasis of cells. Fe-SOD (FSD) and Mn-SOD (MSD) belong to the super oxide dimutese (SOD) gene family. The expression of SOD, peroxidase (POD) and catalase (CAT) related genes in DlBGAL9-OE/DlAGL80-OE roots were significantly upregulated (Fig. 7c). In summary, DlBGAL9 and TF DlAGL80 may reduce the damage caused by heat stress to longan hairy roots by regulating the content of SOD, POD and CAT.
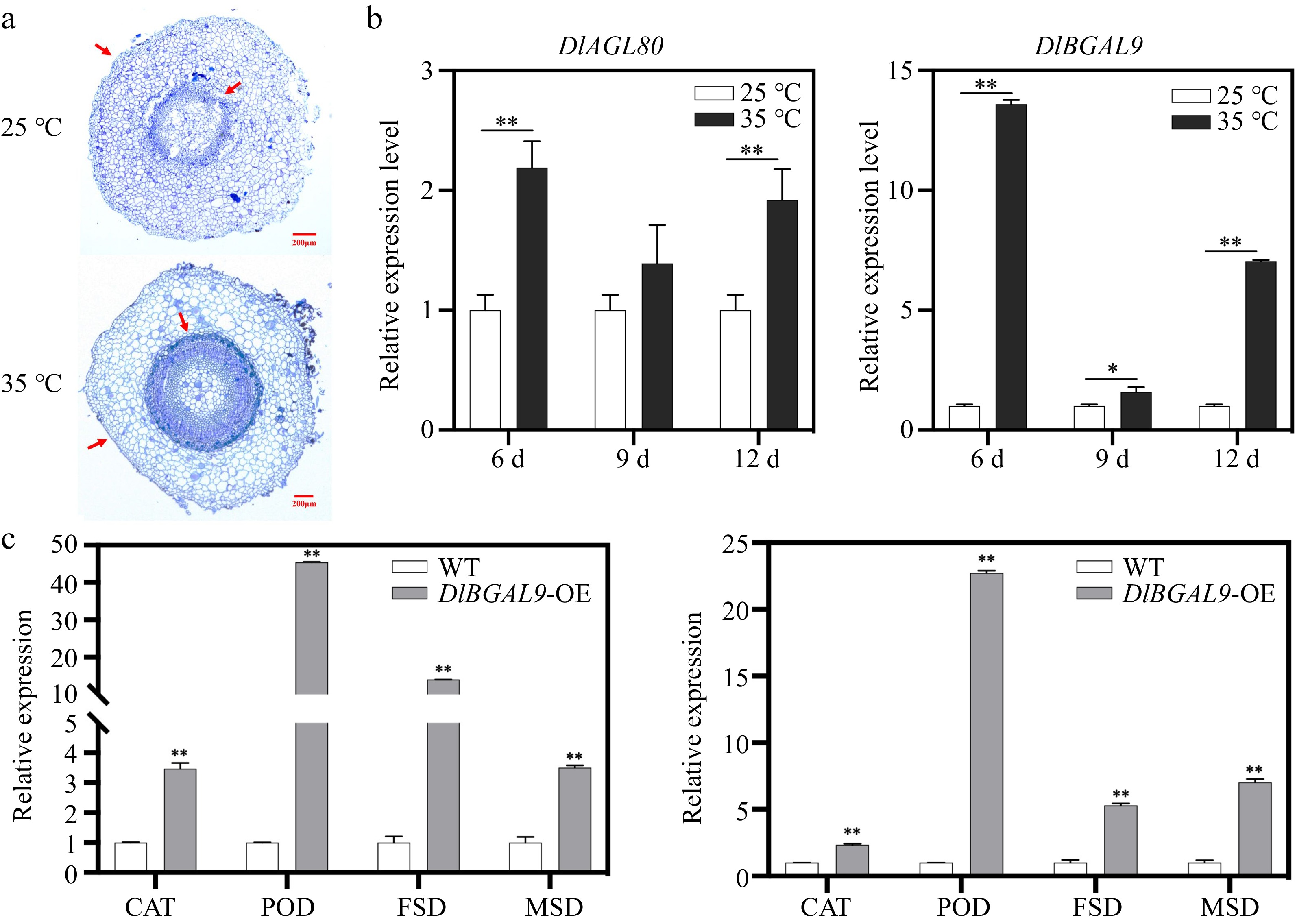
Figure 7.
Expression pattern analysis of DlBGAL9 and DlAGL80 under heat stress in longan hairy root. (a) Anatomical and morphological observations of longan hairy root under heat stress. Arrows indicate the position of cell wall discrepancy. (b) Expression pattern analysis of DlAGL80 and DlBGAL9 under different days of heat stress in longan hairy root. (c) Expression profiles of ROS pathway related genes in WT, DlBGAL9-OE and DlAGL80-OE longan hairy roots. (*p < 0.05, **p < 0.01).
-
Plant BGAL family is widely involved in cell wall structure, intercellular protein modification[11,12], and plant growth and development[46,47]. In this study, 20 BGAL family members were identified in 'Honghezi' longan, which was similar to the number of Arabidopsis thaliana (17 AtBGALs)[48], Solanum lycopersicum (17 SlBGALs)[48], Prunus persica (17 PpBGALs)[46] and Oryza sativa (23 OsBGALs)[8], but less than that of Linum usitatissimum (43 LuBGALs)[49] and Brassica juncea (39 BjuBGALs)[50]. The protein sequence of the GH35 active site is the site of β-galactosidase specific binding substrate, so the absence and incompleteness of this site may be related to the loss of enzyme activity. Except for DlBGAL13-1 and DlBGAL8-3, all DlBGAls possess the typical GH35 active site sequence GGP[LIVM]xQxENE[FY]. In Arabidopsis, AtBGAL1~AtBGAL6, AtBGAL10 and AtBGAL12 contain GH35 active sites[48]. In sweetpotato, GH35 active sites were detected in all members of except Ibbgal13[10]. However, the GH35 active sites of BGAL family were missing or incomplete in plants such as Brassica junce, Chinese cabbage, flax and cotton. In the longan BGAL family, introns were identified to be numerous and dense. Similar to studies in other species, it is speculated that members of the BGAL family may be more affected by regulatory elements.
Phylogenetic trees showed that the BGAL proteins of longan and citrus were more closely related. Cis-elements are involved in the regulation of gene expression, which is an important component of the response regulatory network for plant growth and development, hormone response, and stress response[42,43]. Cis-acting elements of the DlBGAL promoter revealed that most of them contain elements related to light signaling, hormones, stress, and development. Studies have found that ethylene can inhibit the transcription of PaGAL3 and PaGAL4 in avocado fruits, as well as the relationship between auxin induced plant embryo elongation and β-galactosidase activity. In addition, the transcription of BGAL is associated with salt stress, drought stress, and pathogen infection. In summary, BGAL family members may participate in various regulatory networks such as signal transduction and stress response during the growth and development of longan.
DlAGL61/DlAGL80 activate the expression of DlBGAL9 and participated in the regulation of longan somatic embryogenesis
-
The remodeling of cell wall polysaccharide networks is an important process in plant growth and response to external signals[51]. Pectin is a polysaccharide that helps maintain cell wall porosity and cell adhesion. Pectin has the ability to bind to ions, growth factors and enzymes, and is involved in cell differentiation and elongation[52,53]. More and more evidence suggests that cell wall components like pectin and arabinogalactan-proteins play a crucial role in plant somatic and zygotic embryogenesis[54,55]. BGALs belong to the GH35 family and are widely involved in cell wall structure, modification of intercellular connections, and cell wall metabolism. In this study, six of the 11 DlBGALs whose expression were highly expressed in GE, and it is speculated that these DlBGAL genes may be conducive to promoting early SE. Similarly, transcription of all BGAL members in Arabidopsis occurs during early development[14], and CanBGal-5 in chickpea was associated with high cell division rate growth[16]. Based on the above results, it was speculated that the DlBGALs may participate in the process of SE through cell wall modification.
MADS-box genes encode a large transcription factor family in eukaryotes, widely considered as a regulatory factor involved in reproductive development, floral transformation, and organ identity[56]. We selected DlBGAL9 and two MADS-box genes (DlAGL61 and DlAGL80) of 10 TFs that based on RNA-seq. To understand the relationship between TFs DlAGL61/80 and DlBGAL9, we transiently transformed DlAGL61 and DlAGL80 into longan EC. The results confirmed that TFs DlAGL61 and DlAGL80 significantly increased the expression of DlBGAL9. We further verified that the TFs DlAGL61/80 could enhance the expression of DlBGAL9 by a LUC reporter assay performed in Nicotiana benthamiana leaves. These results showed that DlAGL61 and DlAGL80 enhance the expression of DlBGAL9 to different extents. GmAGL15 was preferentially expressed in embryos during soybean development, and overexpression of GmAGL15 promotes soybean somatic embryo development[57]. In tomato, DkGAL1 was involved in seed germination and radicle elongation[58]. The above results further confirmed that the TFs DlAGL61 and DlAGL80 could enhance the expression of DlBGAL9 and participate in the regulation of longan SE.
DlAGL80 promote DlBGAL9 expression to thicken the cell wall and respond to heat stress
-
In plants, BGAL regulates cell wall remodeling and occurrence by regulating polysaccharide metabolism of cell wall components[59]. In Arabidopsis, all BGAL genes were expressed in the cell wall. Dean et al.[60] observed BGAL6 localization on the cell wall through subcellular localization. Gantulga et al.[61] detected the localization of BGAL2 and BGAL5 on the cell wall by dotblotting, while Hui et al.[62] detected the localization of BGAL8 on the cell wall by proteomics. Gantulga et al.[63] showed that BGAL1 and BGAL12 were present in the xylem thickened cell walls through immunogold labeling followed by transmission electron microscope of root sections. AGL regulates the development of eukaryotes by participating in cell proliferation and differentiation. AGL21 plays a crucial role in the growth process of Arabidopsis lateral roots[64]. The number of lateral roots in mutant agl21 decreases, while overexpression of AGL21 can increase the number of Arabidopsis lateral roots[64]. In this study, the longan transgenic hairy roots of DlBGAL9-OE and DlAGL80-OE promoted the accumulation of β-GAL enzyme activity and cell wall thickening. According to the above results, it is speculated that DlAGL80 regulates the β-GAL enzyme activity by regulating the expression of DlBGAL9, and then participates in the modification of cell wall. In addition, the pectin content in DlBGAL9-OE hairy roots was significantly downregulated. The many roles of pectin in plant development. Genes relevant to pectic homogalacturonan biosynthesis, modification and degradation are expressed with spatiotemporal specificity and respond to intrinsic growth and environmental cues throughout the plant life cycle. They function in many developmental processes by regulating cell wall composition, structure and mechanical properties[65]. Meanwhile, cell wall pectin, cellulose, and cytoskeleton synergistically regulate the molecular mechanisms of cell wall integrity and plant growth and development[66]. Therefore, overexpression of the AGL61/80-BGAL9 module may lead to a more complex regulatory mechanism for cell wall thickening.
Global warming has affected plant growth and development[67]. Previous studies have found that heat stress inhibits the normal somatic embryogenesis of longan[68]. In RNA-seq data of treated at different temperatures, it was found that most of the detected DlBGAL genes were upregulated at heat stress. Interestingly, the expression of DlBGAL9 and TFs DlAGL61/80 was significantly upregulated at 12 d of heat stress (35 °C) in longan EC, while β-GAL activity was significantly downregulated. This may be due to the poor thermal stability of β-GAL[69]. Some studies have found that the BGAL family is also regulated by biological and abiotic stresses[17], including salt stress[18], water stress[20] and pathogen stress[18]. It was speculated that DlBGALs may participate in stress response during SE through cell wall modification. Heat stress may lead to lower ion concentrations in cell fluid and promote the production of ROS and other toxic compounds. Excessive ROS accumulation can seriously affect plant growth and development, reduce plant cell viability, and programmed cell death[70,71]. At the same time, the activity of SOD, CAT, and POD increases to maintain intracellular homeostasis[72]. In this study, qRT-PCR was used to analyze the expression patterns of EC and hairy roots of DlBGAL9 and DlAGL80 in longan under heat stress. Under heat stress, the expression levels of DlBGAL9 and DlAGL80 were upregulated, while the β-GAL activity was decreased. In addition, SOD, CAT and POD related genes were upregulated in overexpressed DlBGAL9 and DlAGL80 longan hairy roots. In summary, DlBGAL9 and DlAGL80 may respond to the process of heat stress of longan by regulating ROS accumulation.
-
In this study, 20 BGAL genes were identified in the longan genome. The expression patterns of DlBGAL genes in early SE and stresses responses were different, which coordinately promoted the process of early SE and resistance to heat stress. We screened DlBGAL9 at the transcriptional level and verified that TFs DlAGL61/80 can promote the expression of DlBGAL9. In addition, it was verified that DlAGL80 promote DlBGAL9 expression to thickens cell wall and respond to heat stress. These findings provide a model in which DlAGL80 regulation of DlBGAL9 enhances β-GAL activity to promote cell wall modification during early longan SE and responses to heat stress (Fig. 8).
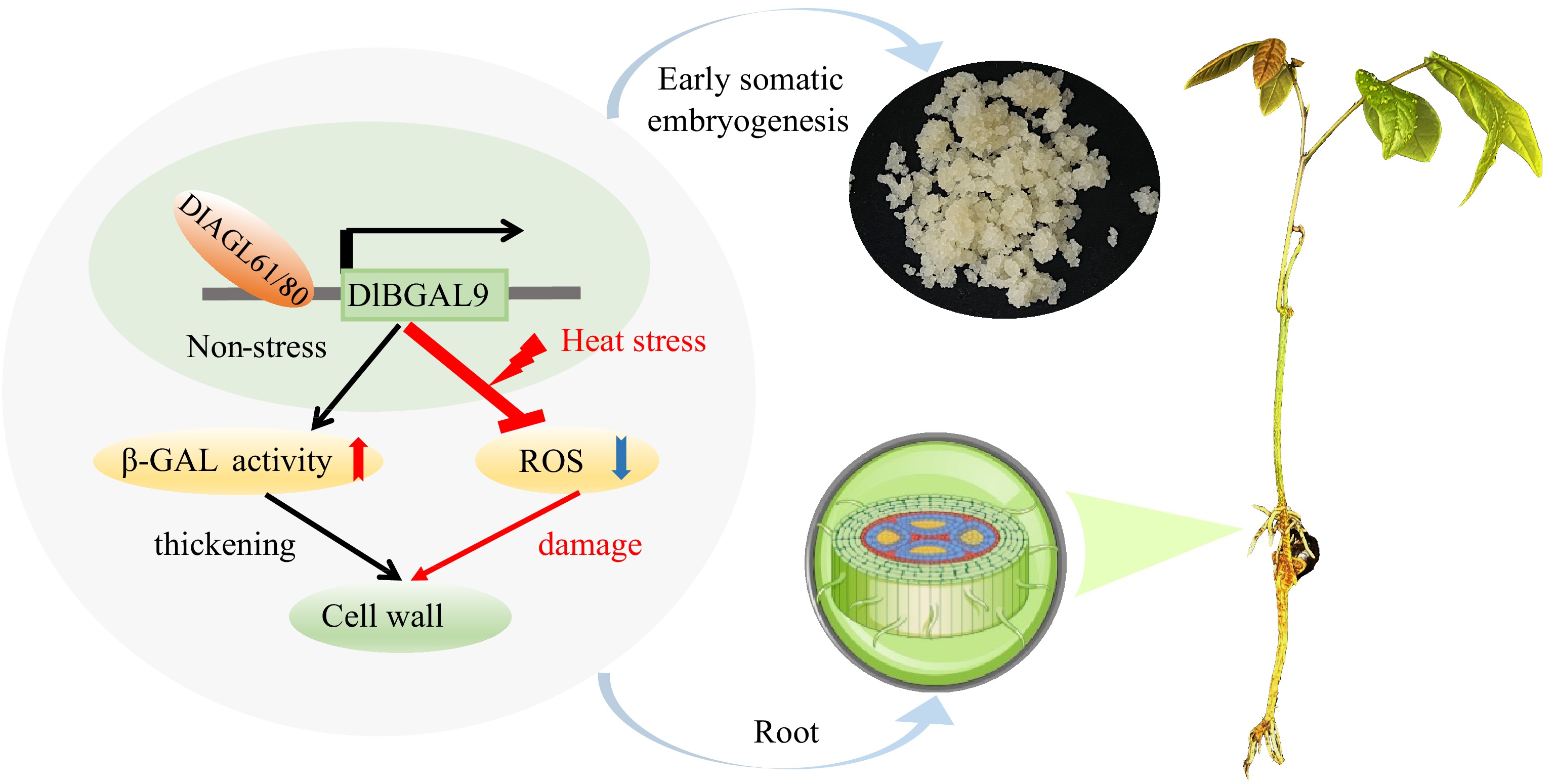
Figure 8.
Proposed model of the underlying mechanism via which TFs DlAGL61/80 and DlBGAL9 regulate early somatic embryogenesis and responding to heat stress in longan. DlAGL61/80 activate the expression of DlBGAL9, leading to upregulation of β-GAL activity and thickening of cell walls. Reduce the content of ROS and reduce damage under heat stress. Arrow for promotion and blunt ends indicate inhibition, respectively. Red arrow represents upregulation of β-GAL activity and blue arrow represents downregulation of ROS content.
-
The authors confirm contribution to the paper as follows: study conception and design: Lin Y, Lai Z; data collection: Xu L, Lai C, Zhang X, Guan Q, Zhang Z, Chen Y; analysis and interpretation of results: Chen Y, Ma X, Ma W; draft manuscript preparation: Chen Y, Ma X. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by the National Natural Science Foundation of China (31672127), the Natural Science Foundation of Fujian Province (2020J01543), the Constructions of Plateau Discipline of Fujian Province (102/71201801101), and the Technology Innovation Fund of Fujian Agriculture and Forestry University (KFb22022XA and CXZX2019033S).
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yan Chen, Xiangwei Ma
- Supplemental Table S1 Summary of primers used in this study.
- Supplemental Table S2 The synonymous and non-synonymous substitution rates of DlBGAL famliy.
- Supplemental Fig. S1 Schematics of the phylogenetic tree, gene structures and conserved motifs of the DlBGAL gene family. (a) The subgroup names were labeled according phylogenetic tree. (b) Exon-intron structure of DlBGAL genes. Yellow boxes indicate exons, purple boxes indicate UTR and gray lines indicate introns. (c) Conserved motif analysis. The motifs, numbers 1–10, were displayed in different colored boxes.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Chen Y, Ma X, Ma W, Xu L, Lai C, et al. 2024. AGL61/80 regulates BGAL9, which participates in early somatic embryogenesis and increases longan resistance to heat stress. Fruit Research 4: e013 doi: 10.48130/frures-0024-0005 

AGL61/80 regulates BGAL9, which participates in early somatic embryogenesis and increases longan resistance to heat stress
- Received: 17 November 2023
- Revised: 01 January 2024
- Accepted: 04 January 2024
- Published online: 02 April 2024
Abstract: β-galactosidase (BGAL) is a glycosyl hydrolase that participates in cell wall modification, playing an essential role in plant growth, development and environmental stress adaptation. However, the function of BGALs in longan remains unclear. In this study, a total of 20 BGAL genes were identified from the longan (Dimocarpus longan Lour.) genome, which were distributed to eight chromosomes. Transcript profiles revealed that the majority of 11 detected DlBGAL genes were highly expressed in the embryogenic callus (EC) (four) and globular embryos (GE) (six), and responsive to heat stress. Combined with RNA-seq, ATAC-seq, and ChIP-seq results, most DlBGAL genes differentially expressed during early somatic embryogenesis (SE) were related with chromatin accessibility and high levels of H3K4me1. DlBGAL9 was selected for further analysis. Dual-luciferase assays and transient transformation indicated that the transcription factor (TF) DlAGL61 and DlAGL80 might bind to the DlBGAL9 promoters to activate DlBGAL9 transcription. Overexpression of DlBGAL9 and TF DlAGL80 induces longan hairy roots β-GAL activity and thickening of cell walls. The expression levels of DlBGAL9 and TF DlAGL80 in longan hairy roots were significantly increased under heat stress, and ROS scavengers related genes were significantly upregulated in overexpressing DlBGAL9 and TF DlAGL80 hairy roots. This study proposes the significance of the regulatory network composed of DlBGAL9 and TF DlAGL80 in regulating the early longan SE and heat stress response.
-
Key words:
- D. longan /
- β-galactosidase (BGAL) /
- Transcriptional factor /
- Somatic embryogenesis /
- Heat stress