








-
Sorghum is the fifth most important cereal worldwide, after corn, wheat, rice and barley[1]. The particular agronomic characteristics of sorghum have led to an increase in the area under cultivation in recent years, since it can be included in rotations and be beneficial for the soil, occupying a fundamental role in the new Argentina agro-industrial chain and gaining more and more worldwide relevance[2]. The uses of sorghum are multiple: it is used mainly in animal feed (especially for cattle) and also in human consumption (food for celiacs). Some properties make it suitable as an input for the production of paper, adhesives, mineral refinement and sausage production, among other industrial uses[3]. These qualities have been reflected in a notable increase in its consumption worldwide. However, their productivity and economic value are threatened by fungal diseases[4] that reduce yields and alter the quality and safety of crops due to the presence of mycotoxins such as aflatoxins, fumonisins, zearalenone and deoxynivalenol[3]. However, in recent years, the focus has been on another mycotoxin, tenuazonic acid (TeA), since it was associated with the contamination of sorghum grains and derivatives[5,6]. Its presence was recently reported in 100% of the sorghum samples analyzed in our laboratory, and the isolates analyzed in this work were obtained from those same samples[7].
Tenuazonic acid [(S)-3-acetyl-5-(S)-sec-butyltetramic acid)] is a derivative of tetramic acid, a potent inhibitor of protein biosynthesis that causes various pathologies in animals and man[8,9]. Tenuazonic acid was first isolated by Rosett et al.[10] and its production is fundamentally associated with the Alternaria genus[11] and, to a lesser extent, with other fungal species such as Epicoccum sorghinum (= Phoma sorghina) and Pyricularia oryzae[12,13]. However, in recent years the scientific community has paid special attention to this toxin due to its persistent occurrence in foods and beverages, and mainly after the Bavarian Health and Food Safety established a permitted limit in sorghum/millet-based baby foods (500 μg/kg)[14].
Epicoccum sorghinum (Sacc.) Aveskamp, Gruyter & Verkley 2010, in addition to being one of the main fungal contaminants in pre and post-harvest sorghum grains[15,16], is a recognized producer of tenuazonic acid. According to Oliveira et al.[17] there are numerous reports of its presence in food and beverages in recent years, although few report its presence in sorghum and derivatives. Not enough attention has been paid to the relationship between TeA contamination and the presence of E. sorghinum, which is why more studies are needed to understand the association between TeA contamination of food and the presence of E. sorghinum in cereals. There is clear evidence of the involvement of Alternaria in contamination with TeA in food, especially in sorghum grains as a producer of that mycotoxin[18]. The presence of E. sorghinum in sorghum grains from the humid Argentine Pampa has been reported for more than a decade[19], however, there are no previous reports in Argentina that associate it with the presence of TeA.
In a previous study carried out by this research group[20], the mycotoxicological quality of 19 samples of visibly healthy sorghum grains destined for animal consumption was evaluated. These grains were harvested in March 2017 and sent post-harvest for analysis without prior storage. One hundred grains of each sample previously disinfected were placed in Dichloran-Glycerol 18% (DG18) and Dichloran Rose Bengal Chloramphenicol (DRBC) medium, incubated at 28 °C during 7 d and the infection percentage was calculated. It was observed that 100% of assayed sorghum grain samples were contaminated with several fungal genera that potentially produce mycotoxins, being Epicoccum genus the most prevalent (84%). This fact, added to the limited current information on the occurrence of this fungal species in this cereal in Argentina, is intended to characterize the isolates of Epicoccum obtained from the sorghum samples previously analyzed.
-
From a total of 180 isolates of several fungal genera obtained from sorghum grain samples (experimental hybrids) from the experimental station of the National Institute of Agricultural Technology (INTA)-Manfredi, Córdoba, 40 isolates belonging to Epicoccum genus were used in order to carry out this study.
These isolates were deposited in the Collection of the Mycotoxins Laboratory of the Center for Research and Development in Industrial Fermentations (CINDEFI-CONICET-UNLP), where they are preserved by freeze-drying and cryopreservation.
Morphological characterization
-
For the morphological characterization, the Phoma Identification Manual was used[21]. All isolates were inoculated on oat agar (OA) and malt extract agar (MEA) and incubated in complete darkness at 22 °C for 7 d. The OA medium contained rolled oats (65 g·L−1) and purified agar (20 g·L−1), while the MEA medium contained the following components: malt extract (20 g·L−1), peptone (10 g·L−1), dextrose (20 g·L−1) and purified agar (20 g·L−1). Subsequently, the plates were kept for an additional 7 d at 22 °C with a day-night regime of approximately 13 h of UV light and 11 h of darkness to stimulate the pigmentation of the colonies and the formation of pycnidia[21]. The diameter and the descriptions of the colony were made after 7 d of incubation from the isolates grown in MEA whereas the micromorphological structures were studied from the isolates from OA cultures as Aveskamp et al.[22] suggest. Preparations were mounted in distilled water to study the mature ascomata/conidiomata, ascospores/conidia and conidiogenous cells. Observations were conducted with a Leica DM2500 microscope. The sizes of the structures were determined by averaging the measurements of 50 replicates of each structure.
Tenuazonic acid production
-
In order to evaluate the toxigenic potential of 40 assayed isolates, plates containing DRYES medium[23] were inoculated with each E. sorghinum isolate (at three points equidistant from each other) and incubated for 14 d at 25 °C in darkness. Basal medium consisted of glucose (10 g·L−1), peptone (5 g·L−1), MgSO4·7H2O (0.5 g·L−1); K2HPO4 (10 g·L−1), agar (15 g·L−1) and rose bengal (5% [wt/vol] aqueous). Filter-sterilized chlortetracycline was added to sterilized media to give a final concentration of 10 µg mL−1.
The extraction was carried out at micro-scale using the method described by Andersen et al.[24] for Alternaria metabolites. Three agar plugs from the centre of each colony, and therefore nine plugs from each isolate, were placed in a 4 mL vial. One mL of ethyl acetate with the addition of 1% (v/v) formic acid was added to each vial and the toxin was extracted by sonication for 30 min. The extract was transferred to a clean 2 mL vial, evaporated to dryness under N2 flow and re suspended in 200 µL of a solution of methanol : water (50/50). The extract was filtered (through filter syringe filters, 17 mm, 0.45 μm, nylon membranes, TITAN) and kept at −18 °C until analysis.
Chemical reagents
-
HPLC grade acetonitrile (ACN) was purchased from Sigma–Aldrich and all other chemicals utilized in this study were HPLC quality. Tenuazonic acid standard was dissolved in methanol by further dilution with acidified water (pH 4.0) to obtain concentrations of 30, 50, 80 and 100 mg mL−1.
Chromatographic conditions
-
The HPLC equipment used was the Waters 717 plus Autosampler, equipped with a quaternary pump and the Empower software (Chromatography Data System, CDS, Waters Corporation, Milford, MA, USA) for data analysis. The TeA was analyzed after separation on a reverse phase column using a C18 column (150 mm × 4.6 mm, 5 μm particle sizes, Waters Corporation) with an ultraviolet photodiode array detector set at 280 nm.
The mobile phase consisted of methanol : water (70:30, v/v) containing 300 mg of zinc sulphate L−1 with a constant flow rate of 1 mL·min−1. Each analysis was performed in duplicate. Photodiode array detection (DAD) was performed to control toxin identity; the injection volume was 20 μL and the retention time was around 10 ± 1 min. Identification was performed by comparing retention time and spectra monitored (280 nm) by a photodiode array detector of peak in the sample with those of the pure toxin standard, and external calibration was used for quantitation. The calibration curve for quantification purposes was constructed using the toxin standards, and the values were obtained by correlation of concentration and peak-area.
The quantification limits of the method were taken as the minimum amount of toxin detected in the samples that allowed obtaining contrary information using the diode array detector. Detection limits using the DAD detector were measured as three times the reference standard variation under the same conditions used for those samples. The limit of quantification (LOQ) for TeA was 60 μg·kg−1.
Molecular characterization
-
Of the 40 studied isolates, nine isolates were selected for the molecular characterization, including representatives of each of the morphological groups observed, as well as isolates both producers and non-producers of TeA.
DNA extraction and quantification
-
DNA extraction was done according to the protocol described by Tannous et al.[25] with some modifications. For the production of biomass, each isolate was inoculated in an Erlenmeyer flask with 2% of yeast extract and 15% of sucrose (YES broth) for 24 h, the mycelium was taken with sterile forceps and placed in a sterile Eppendorf. Then 700 µL of CTAB buffer [1 M Tris-HCl pH 8.4 (10 mL), NaCl (8.2 g), 0.5M EDTA pH 8 (5 mL), CTAB (2 g), distilled water (100 mL)] preheated to 65 °C was added and pipetted several times until a homogeneous suspension was formed. Then, it was incubated at 65 °C for half an hour, 600 µL of chloroform was added and shaken. They were centrifuged at maximum speed for 10 min. The upper aqueous phase was transferred to an Eppendorf tube, and the same volume of cold isopropanol was added to precipitate the DNA. The tubes were inverted 2−3 times and incubated for 30 min in the −20 °C freezer. They were centrifuged at maximum speed for 10 min. Supernatants were removed and the pellets washed with 600 µL of 70% ethanol. They were centrifuged at maximum speed for 10 min and the upper phase were carefully removed. The pellets were allowed to dry at room temperature and resuspended in 60 µL of sterile DNase-free water. The samples obtained were measured in NanoDrop™ 2000/2000c Spectrophotometers (Thermo Scientific) to confirm the presence of DNA in the observed precipitate and the degree of purification.
DNA amplification and sequencing
-
The extracted DNA was used to amplify two gene regions with the polymerase chain reaction (PCR). The 5.8S ribosomal RNA (rRNA) gene was amplified using primers ITS1 (TCCGTAGGTGAACCTGCGG) and ITS4 (TCCTCCGCTTATTGATATGC) and β-tubulin target gene (TUB2) was amplified with primers Bt2a (GGTAACCAAATCGGTGCTGCTTTC) and Bt2b (ACCCTCAGTGTAGTGACCCTTGGC)[26]. PCR was performed using a 50 mL reaction mixture containing the following (per reaction): 50 ng of genomic DNA; 5X GoTaq reaction buffer; 0.2 mM (concentration of each deoxynucleoside triphosphate); 2 mM concentration of each primer (ITS1-ITS4 and Bt2a-Bt2b); 1.25 units of GoTaq DNA polymerase. Amplifications with both sets of primers were performed in a GeneAmp_PCR System 9700 (Applied Biosystems) and the PCR program was as follows: 95 °C for 2 min and 30 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 45 s, and extension at 72 °C for 1 min 45 s followed by a 10 min final extension at 72 °C. Five microliters of the mixture were analysed by electrophoresis on 1% agarose gels and visualized by ethidium bromide (0.4 mg·mL−1).
Amplified and purified gene products were sent to sequence by Macrogen services (Macrogen Inc., Seul Korea). It was purified with AMPure XP beads (Beckman-Coulter), and the DNA was sequenced by capillary electrophoresis with the Genetic Analyzer 3500xl (Applied Biosystems) equipment.
Sequential analysis
-
Sequences were handled with BioEdit 7.0.5.3 software[27] which included examining the chromatograms files, assembling the forward and reverse reads and manual trimming. Combinated sequences of ITS and TUB2 were used. The similarity of nucleotide sequences separated and combined were calculated using the BLAST procedure (
http://blast.ncbi.nlm.nih.gov ) by database Nucleotide collection (nr/nt). Hits for each query sequence exceeded the threshold for coverage and sequence similarity recommended by expert mycology researchers[28].To construct the cladogram, phylogenetically close isolates to E. sorghinum previously reported by Chen et al.[29] & Hou et al.[30] were used to provide robustness to our analysis. Subsequent multiple alignment was generated with ClustalW[31]. Maximum Likelihood analyses including 1000 bootstrap replicates, which were conducted using RAxML[32]. A general time reversible (GTR) model was applied with a gamma distributed rate variation. The resulting tree was viewed using FigTree v. 1.4.4 (
http://tree.bio.ed.ac.uk/software/figtree ). Leptospaheria doliolum CBS 505.75 was selected as an outgroup. -
Significant variations in the macromorphological characteristics of the 40 analyzed E. sorghinum isolates were observed, as expected considering the high intraspecific variation presented by this group of fungi[33]. This fact allowed us to differentiate the isolates into three specifically defined groups (A, B and C) based, mainly, on their growth rate and their ability to produce pigments and exudates (Fig. 1). All the characteristics described below were observed after 7 d of incubation in MEA medium. Group A included 12 isolates with smaller diameter colonies, visibly furrowed, mycelium with a light centre that becomes darker towards the periphery and pink edges; intense pink reverse, especially in the centre, the intensity of the colour decreases towards the periphery of the colony, with the youngest mycelium being whitish. Some of these isolates presented transparent exudates. On the other hand, 21 isolates are classified within group B and the colonies, in this case, had an intermediate diameter, floccose but not scarce with mycelium from pink in the centre to greyish green towards the periphery, culminating in whitish colour; reverse of homogeneous intense brown to almost black colour, with reddish pigment that extends to the culture medium. Finally, group C was represented by seven isolates whose colonies showed the greatest development over time, presenting soft grooves, a light brown centre that turns salmon, with white edges; clear back, light salmon centre that turns whitish towards the periphery.
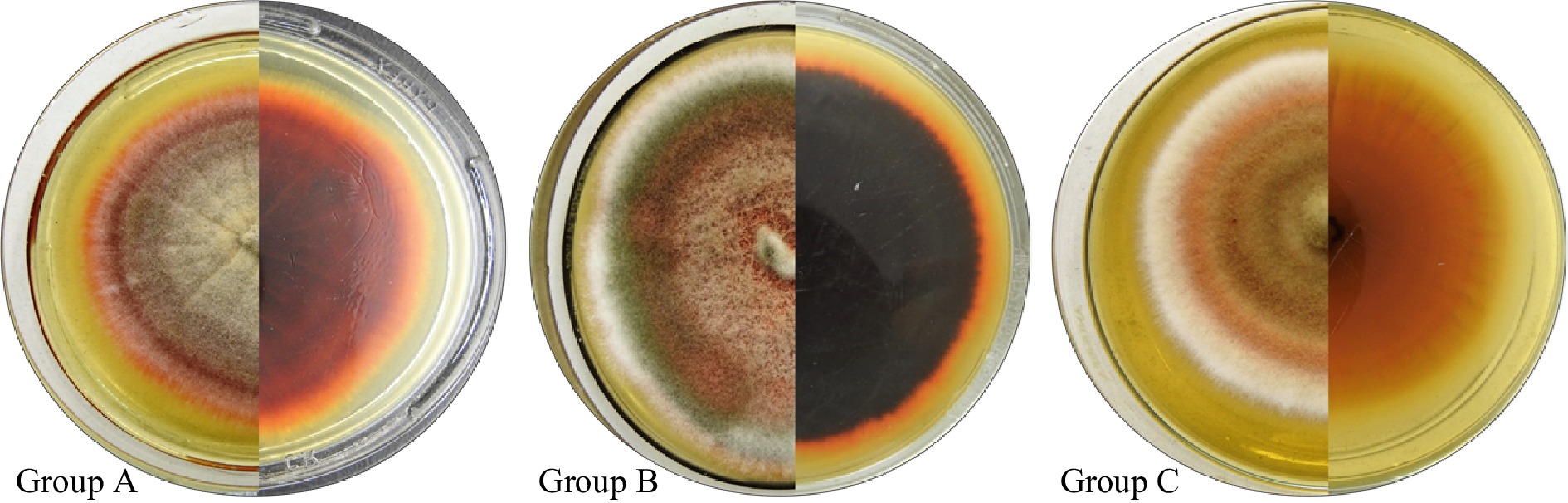
Figure 1.
Macroscopic characteristics of E. sorghinum: colony on MEA medium after 7 incubation days (front and reverse). Representatives of morphological groups A, B and C.
The members of the B group covered the whole Petri dish, in contradistinction to the members of the other two groups whose colonies reached a maximum diameter between 60−75 mm.
The macromorphology observed in the colonies grown in OA was similar for all the analyzed isolates, showing no differences between the morphological groups described. The colony reached a diameter between 50−90 mm, with regular edges, and olive-green conidia. The colonies showed a compact and felty texture, while on the reverse a brown pigmentation was observed.
All the studied isolates showed typical micromorphological characteristics of this species: brown globose pycnidia with a straight neck, of 69 to 11.5 × 44.5 to 90 µm in size (n = 50); multicellular hyaline to brown chlamydospores, massively produced with measurements ranging between 3 and 50 μm, and mostly ovoid conidia of 3.5 to 5.5 × 1.8 to 3 µm (n = 50) (Fig. 2).

Figure 2.
Micromorphological structures of E. sorghinum. (a) hyaline/brown dictyochlamydospores, and (b) Hyaline/brown glabrous pycnidia. Scale bars = 10 μm.
Tenuazonic acid production
-
A considerable variability in mycotoxin production was also observed as it happened with the morphological diversity. Regarding the toxigenic capacity of E. sorghinum, the results suggest a toxicological risk for animals exposed to tenuazonic acid (TeA) through the consumption of feed contaminated with this producer species. This assertion is based on the correlation between the sorghum samples contaminated with TeA and the origin of the isolates that turned out to be TEA-producers isolates. Sixty-five percent of the analyzed isolates (n = 40) were producers of TeA with levels that ranged from 112 to 47,237 μg·kg−1. Table 1 shows the TeA concentration by the 26 TeA-producing E. sorghinum isolates.
Table 1. Tenuazonic acid (TeA) concentration produced by assayed E. sorghinum isolates and their corresponding morphological group.
Morphological group Isolates TeA concentration (μg·kg−1) A LMCIN-2.1 13,846.39 LMCIN-5.3 7,556.21 LMCIN-5.7 112.45 LMCIN-5.11 11,948.50 LMCIN-7.1 28,000.83 LMCIN-8.1 2,001.46 LMCIN-9.6 1,679.33 LMCIN-9.11 5,129.46 LMCIN-11.1 548.78 LMCIN-12.4 3,870.20 LMCIN-18.4 721.33 B LMCIN-1.12 5,890.30 LMCIN-3.2 16,395.00 LMCIN-3.11 1,289.45 LMCIN-7.2 3,058.24 LMCIN-7.3 389.49 LMCIN-9.3 7,790.25 LMCIN-9.10 5,900.17 LMCIN-11.7 18,374.58 LMCIN-13.5 47,237.28 LMCIN-15.3 6,720.12 LMCIN-16.4 3,245.55 C LMCIN-6.9 2,190.36 LMCIN-12.8 4,069.33 LMCIN-18.7 16,280.38 LMCIN-19.2 8,130.35 Molecular characterization
-
The phylogenetic analysis of the ITS and TUB2 sequences separately and the morphological characteristics finally allowed us to identify the analyzed strains as E. sorghinum. The analysis by Blast database could identify the majority of the isolates as E. sorghinum, whose e-values and percentage of identity were of 0.0 and 100%, respectively. Except for the isolate LMCIN-18.4 (e-value 0.0 and percentage of identity 100%) that was identified as Phoma sp.
Most of the isolates showed 100% similarity in the analyzed sequences, therefore not all of them were included in the phylogenetic analysis, since the objective of the work was to verify the morphological identification and to determine the degree of similarity with those reported strains.
Table 2 shows the access number of GenBank of the nucleotide sequences of the assayed isolates in this study and the number access of the sequences used by the phylogenetic analysis.
Table 2. Reference sequences selected according to the taxonomic closeness and downloaded from GenBank to construct the phylogenetic tree.
Species Isolate Country GenBank accession ITS TUB2 E. sorghinum LMCIN-1.12 Argentine OQ971382 OR125009 E. sorghinum LMCIN-1.12 Argentine OQ971382 OR125010 E. sorghinum LMCIN-5.11 Argentine OQ971384 OR125011 E. sorghinum LMCIN-6.9 Argentine OQ971385 OR125012 E. sorghinum LMCIN-9.6 Argentine OQ971386 OR125013 E. sorghinum LMCIN-9.11 Argentine OQ971387 OR125014 E. sorghinum LMCIN-11.7 Argentine OQ971388 OR125015 E. sorghinum LMCIN-12.8 Argentine OQ971389 OR125016 E. sorghinum LMCIN-18.4 Argentine OQ971390 OR125017 E. sorghinum CBS 179.80 Puerto Rico FJ427067 FJ427173 E. sorghinum CBS 627.68 France FJ427072 FJ427178 E. sorghinum LC 4860 China KY742116 KY742358 E. viticis LC 5126 China KY742118 KY742360 E. camelliae LC 4858 China KY742091 KY742333 E. latusicollum LC 5158 China KY742101 KY742343 E. pimprinum CBS 246.60 India FJ427049 FJ427159 E. longiostiolatum CBS 886.95 Papua New Guinea FJ427074 FJ427180 Leptosphaeria doliolum CBS 505.75 Netherlands JF740205 JF740144 Figure 3 shows the phylogenetic dendrogram constructed starting the combined ITS + TUB2. Associations between macro morphological variation, toxicogenic capacity and the phylogenetic results were found. In this way, LMCIN-9.11, LMCIN-9.6 and LMCIN-5.11 isolates were grouped in a cluster presenting all these isolates the morphology described within group A; on the other hand, isolates representatives of morphological group B were grouped into the following cluster: LMCIN-1.12, LMCIN-3.11 and LMCIN-11.7 isolates. It is highlight that all the isolates mentioned above are mycotoxin-producing isolates while the two non-mycotoxin-producing isolates included in the molecular analysis (LMCIN-6.9 and LMCIN-12.8) are located in the same subnode, however, in this case there was no correlation with the macromorphological characteristics since they belong to different morphological groups.
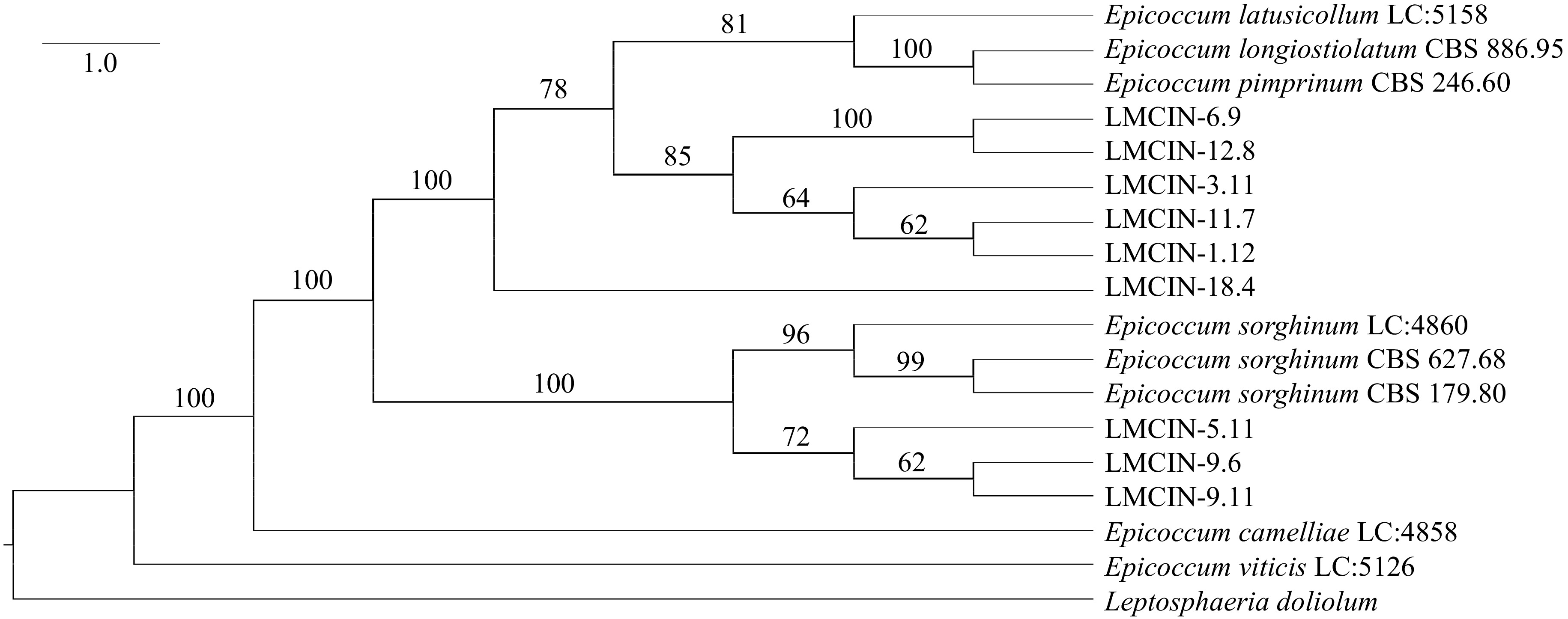
Figure 3.
Phylogenetic tree of the Epicoccum isolates obtained from sorghum samples, derived from sequences of the ITS and β-tubulin region of the nuclear ribosomal DNA.
Other results of the taxonomic search for isolates: LMCIN-1.12, LMCIN-3.11, LMCIN-6.9, LMCIN-11.7, LMCIN-12.8, were relationated to Epicoccum latusicollum with similar values confidence to obtained above.
-
Epicoccum sorghinum is a fungal species studied worldwide in the last decade, however, in Argentina there are no current records of its incidence in sorghum grains or the occurrence of tenuazonic acid in that crop. This would be the first work in Argentina that emphasizes a new and emerging pathogen of many vegetable crops. This study also provides a molecular phylogenetic approach to E. sorghinum strains isolated from sorghum in Argentina, confirming their significant genetic and phenotypic variability. Oliveira et al.[17] have isolated this species from different cereals from tropical and subtropical areas. In Argentina as well as in other countries, its distribution has been underestimated due to the difficulty in morphological identification. Currently, molecular tools are used to identify species within the Phoma complex, achieving satisfactory results[6,34].
Species of the Didymellaceae family are cosmopolitan and distributed in a wide range of environments. Most members of this family are phytopathogens of diverse hosts, most of them showing no specificity[35−37]. The various associations with the host plant and its varied morphology make the accurate identification of species in this family challenging[22,36]. However, Chen et al.[36] published a robust main tree has been developed based on internal transcribed spacer regions and 5.8S nrDNA sequences (ITS), partial 28S large subunit (LSU) nrDNA sequences, and partial regions of the second largest subunit of RNA polymerase II (rpb2) and β-tubulin (tub2). To complement our cladogram, we included several Epicoccum type species used by these authors to investigate if the phylogenetic relationships among the isolates corresponded with morphological results and TeA production. We also included the isolate E. longiostiolatum CBS 886.95 in the phylogenetic analysis because it was originally identified as Phoma sorghina, but subsequent studies confirmed it forms a distinct clade from other strains of this species. In later phylogenetic analyses, where P. sorghina was transferred to the genus Epicoccum, the two isolates reported so far of E. longiostiolatum were excluded. More recently, in a phylogenetic study by Hou et al.[30], these two isolates formed a well-supported clade distant from E. sorghinum. Based on the group's phylogenetic history, it is not surprising that the LMCIN-18.4 isolate shows similar phylogenetic distances to both other analyzed isolates in this study and to Epicoccum strains belonging to other species.
Several researchers[6,17,34] have identified E. sorghinum by amplifying only the ITS region of rRNA. Taxonomic studies[22,38] have shown that molecular analysis of a single sequence is insufficient for resolving the group's phylogeny. In this study, molecular tools were used to complement the morphological analysis of the strains and authenticate their identity. However, a more comprehensive systematic study is needed to make sense of the phylogenetic relationships presented in the cladogram.
It is important to highlight that there are no studies of TeA production by E. sorghinum in Argentina, and there are only studies in Brasil reporting the ability of E. sorghinum isolates to produce TeA. Oliveira et al.[17] assessed the ability of a smaller number of E. sorghinum isolates also obtained from sorghum samples to produce this mycotoxin. However, the extraction and quantification methodology assayed, including the media in which the capacity was tested were different that those used in this study. They observed a percentage of producing isolates similar to that found in the present study (57%) but with much lower concentrations. One year later, the same author analyzed the capacity to produce TeA of 11 isolates and observed that 100% of them produced toxin at levels ranging from 98.6 to 148,000 μg·kg−1, concentrations that exceeded those found in our study[39]. However, it would be necessary to widen the number of analyzed isolates to be able to assert that the behaviour of this species is similar in both cases.
This prompts us to continue studying these isolates in depth and to evaluate the transcription profiles of the TeA (TAS1) biosynthetic gene to show whether its expression is consistent with the production of TeA under the conditions in which all the isolates were tested in this study. Knowing this could serve for the formulation of some type of bioinput that blocks or considerably reduces the production of mycotoxins from the varieties of E. sorghinum present in Argentina. In order to propose strategies to sow and postharvest storage of this sorghum and avoid their TEA-contaminated, future in situ ecophysiology studies could be conducted.
This preliminary study reveals that E. sorghinum isolates obtained from sorghum showed wide phenotypic variability, confirming high intraspecific diversity. In addition, it was determined that most of them were TeA producers, representing an economic and sanitary problem for the producers of that crop in Argentina. This finding is an important advance to focus on agroecological alternatives such as the search for bio-inputs (microorganisms isolated from the sorghum ecosystem) based on microorganisms with a fungistatic/fungicidal effect against the growth of these this species and consequently inhibit the TeA production. However, these results transcend the mere replacement of technologies, and challenge us to broaden the look and study of the productive aspects that enhance the contamination of sorghum grains by various fungal species with the expectation of achieving extensive agro-ecological based production that ensures the quality and safety of the sorghum grains that are harvested in Argentina.
Genus Epicoccum exhibits significant genetic and phenotypic variability, making its accurate identification challenging. This has sparked considerable debate and taxonomic changes in recent years. Nonetheless, its environmental, economic, and health significance warrants further ecological and taxonomic research.
-
The authors confirm contribution to the paper as follows: Marcela Hipperdinger: Writing– original draft, molecular characterization and investigation. Déborah Colman: Methodology, formal analysis phylogenetic and investigation. Cecilia Gortari: Morphological characterization and investigation. Carina Pereyra: Physiological characterization, writing – review and editing and supervision. Andrea Astoreca: Conceptualization, resources, writing – review and editing, supervision, project administration and funding acquisition. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
This work was financed by the Consejo Nacional de Investigaciones Científicas y Tecnológicas and Universidad Nacional de La Plata.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hipperdinger ML, Colman DI, Gortari MC, Pereyra CM, Astoreca AL. 2024. Characterization of Epicoccum isolates obtained from Argentinean sorghum grain samples. Studies in Fungi 9: e003 doi: 10.48130/sif-0024-0004 

Characterization of Epicoccum isolates obtained from Argentinean sorghum grain samples
- Received: 04 November 2023
- Revised: 13 March 2024
- Accepted: 21 March 2024
- Published online: 16 April 2024
Abstract: Sorghum has numerous agronomic advantages, a great economic importance in food production and various industrial applications. Its consumption has increased in the last ten years and probably its importance may even increase in the future, considering its relationship with global warming since this plant is less demanding with water. However, its productivity is affected by various fungal diseases with the production of mycotoxins that cause great economic losses. Alternaria, Epicoccum and Pyricularia genera are the main fungal contaminants in sorghum grains, and recognized producers of tenuazonic acid, a mycotoxin previously found in assayed sorghum samples in the Mycology and Mycotoxicology laboratory belonging to the Center for Research and Development in Industrial Fermentations. Fungal isolates obtained from these sorghum grains from the National Institute of Agricultural Technology (INTA, Manfredi, Córdoba, Argentina) were characterized using a polyphasic approach based on morphological and genetic characteristics and in the ability to produce mycotoxins. Morphological analysis suggested the identity of Epicoccum sorghinum, which was later confirmed by molecular analysis. The ability of these isolates to produce tenuazonic acid was evaluated and it was determined that 65% of the studied isolates produced tenuazonic acid at variable levels. This is the first study that provides a molecular approach to E. sorghinum isolates in Argentina and clearly confirms the wide genetic and phenotypic variability previously reported for this species in other countries. The presence of these tenuazonic acid-producing isolates in sorghum grains represent an economic and health problem for Argentina that it is considered one of the main exporters worldwide.
-
Key words:
- Cereals /
- Food safety /
- Fungi /
- Phenotypic and molecular analysis /
- Tenuazonic acid