









-
Zoysia grass (Zoysia japonica) is a widely used warm-season turf grass in China due to its durability, low maintenance cost, good elasticity, and resistance to environmental stresses[1]. The slow growth rate of Zoysia grass leads to a delayed recovery from injuries such as divots and traffic-induced damage. The Zoysia grass' exceptional wear tolerance makes it ideally suited for athletic fields[2,3]. However, its reputation for slow recovery from traffic-related injuries means that even on Zoysia grass athletic fields with low usage, high-traffic areas will necessitate additional time for recovery or the replacement of damaged areas with sod. The slow growth rate of Zoysia grass also caused the delayed establishment, which resulted in a slow production cycle for turf producers. Another challenge is that a slow production cycle leads to higher costs for vegetatively establishing Zoysia grass[4]. The delayed production cycle enhances expenses for sod growers, ultimately affecting the consumer. Most studies on the regeneration of Zoysia grass focus on plant tissue culture, but there are few studies on organ regeneration after mechanical damage. Z. japonica has low fertility and is difficult to modify its traits through crossbreeding[5], so it is important to study the regeneration mechanism of Z. japonica at the molecular level to expand its application scope.
Plant regeneration refers to the ability of plants to rebuild or develop new cells, tissues, or even complete organs after being injured by the external world[6]. The number of stolons is closely related to the regeneration ability of turfgrass. Studies suggested that perennial ryegrass (Lolium perenne) varieties with determinate-stolons could have superior turf quality, wear tolerance, and ability to quickly recover from traffic compared to other popular turf varieties[7]. The number of stolon and rhizome is positively related to the cold-tolerance and the establishment speed of bermudagrass (Cynodon spp.)[8,9]. Patton et al.[10] found that fast-establishing Zoysia genotypes allocated more dry matter to the stem (including stolons, and rhizomes) growth than slow-establishing genotypes. As a result of the greater stem production, the recuperative capacity of improved Zoysia grass cultivars such as El Toro and Palisades could be equivalent to 'Tifway' hybrid bermudagrass[11,12]. However, the mechanism of organ regeneration after mechanical damage was rarely reported. Studies have shown that the molecular mechanism of organ regeneration after mechanical damage in Zoysia is regulated by PIN1 and WUS grass[13]. Another study sequenced the transcriptome of tea leaves under mechanical damage stress and speculated that peroxidase (POD) may be an important antioxidant enzyme in response to mechanical damage stress[14]. However, little is known about the changes in the transcriptome during Zoysia regeneration, and the changes in hormone levels during regeneration are also unknown. In general, the molecular regulatory network during regeneration needs to be further elucidated.
Many plant hormones interact with various cytokines (regulatory proteins and genes, transcription, and epigenetic factors) to promote the expression of differential genes required for mRNA and protein synthesis, which plays an important role in the induction and development of callus[15]. Previous studies found that jasmonic acid (JA) and gibberellic acid (GA) levels increased significantly in the short term (15 and 30 min) after mowing in the creeping bentgrass (Agrostis stolonifera), and the untrimmed plants generally had higher levels of salicylic acid (SA), JA, abscisic acid (ABA), and indole-3-acetic acid (IAA) under high-temperature conditions compared with plants trimmed every 3 d[16]. Ma et al. found that cytokinin (CTK) and GA promoted rhizome formation and growth, respectively, by activating metabolic pathways that supply energy and amino acids to support cell division and expansion during rhizome initiation and elongation in tall fescue (Festuca arundinacea)[17]. Another study found that 6 phytohormones (Indole-3-acetyl-L-valine methyl ester, Indole-3-carboxylic acid, Indole-3-carboxaldehyde, Gibberellin A24, Gibberellin A4, and cis (+)-12-oxo-phytodienoic acid) increased significantly after mowing and appropriate mowing could promote the growth of Anabasis aphylla through the auxin (IAA) metabolic pathway[18]. According to previous studies on plant regeneration, when plants are subjected to some mechanical damage, some unknown signals are transmitted in the plant, which leads to the upregulation of auxin expression, and then causes different molecular mechanisms to respond. This response depends on the damage site and the content of hormones[19]. Auxin, a classical phytohormone, has a key role in many aspects of plant growth and development and also has been proposed to play an important role in tissue and organ restoration during regeneration[20]. CTKs are also involved in a number of basic developmental processes in plants. Higher concentrations of CTKs can promote bud differentiation[21]. In the tissue culture of Zoysia matrella, appropriate concentrations of CTKs were found to promote the differentiation of embryonic healing tissues[22]. Plant regeneration can be achieved by placing isolated plant explants on shoot-inducing medium (SIM) containing IAA and CTK, and inducing adventitious shoots or adventitious roots by adjusting the ratio of IAA to CTK[23].
In this study, two Z. japonica genotypes, FM1 and SM232, showed significant differences in growth and physiological changes after 0, 2, 6, 12, 24, and 48 h of mowing. The regeneration of leaf primordia and new tillers of these two genotypes was observed and transcriptome analysis was performed. The plant hormone variations and the related genes at 0, 2, 6, 12, 24, and 48 h after mowing treatment were determined. The objective of this study were to explore the potential regeneration mechanisms of two different genotypes of Z. japonica after mowing. The present results not only provide a reliable basis for Z. japonica genomic research but also lay clues for regeneration genetic breeding of turfgrass. Moreover, the identified novel candidate genes can be taken as potential genes for further regeneration mechanism elucidation for Z. japonica and other plant species.
-
The Zoysia material used in this experiment was sourced from the Zoysia Experimental Base of China Agricultural University, Shandong Haiyuan Company, Jiaozhou, Shandong Province, China. They were retrieved in the greenhouse of China Agricultural University (40°00' N, 116°35' E) in May 2018 for potting. The potting environment was 25/20 °C-day and night temperature and 50% relative humidity. The materials were planted in pots filled with a mixture of nutrient soil and vermiculite (1:1). There are 10 materials in total, numbered FM1, FM2, FM4, FM6, FM7, FM8, FM9, SM11, SM23, and SM232.
The first mowing was carried out 2 weeks after cultivation. Relevant indicators such as plant height, leaf width, number of stolon, and other related indicators were measured 1 week after the first mowing (no stolon existed at the beginning). The vertical height of each Zoysia grass was measured from base to tip with a ruler and recorded as plant height (cm). The growth rate = (plant height after one week of mowing − plant height at mowing)/growth days (cm/d). Five normal-growing leaves were taken from each plant, and their widest part was measured with a vernier caliper. The average value of each treatment was taken as leaf width (cm). Materials with large differences in regeneration rates were selected for further experiments, and the tip meristem biological tissue sections at 0, 2, 6, 12, 24, and 48 h after mowing were observed using a laser-scanning confocal microscopy (Leica SP2, Wetzlar, Germany).
RNA isolation, reverse transcription, and cDNA library sequencing
-
The stems and leaves of Zoysia FM1 and SM232 were collected at 0, 2, 6, 12, 24, and 48 h after mowing, which were referred to as 'material-time', such as FM1-2h, SM232-2h, etc. The total RNA was isolated using the TianGen RNA Kit (Tiangen Biochemical Technology Beijing Co., Ltd, Beijing, China) according to the manufacturer's instructions (
www.tiangen.com ). The quality of RNA samples was analyzed using the Agilent 2100 Bioanalyzer (Agilent Technologieswww.home.agilent.com ). Library construction was performed using RNA-seq at Beijing Novogene Technologies (Beijing, China). Total RNA was fractionated using Oligo (dT) magnetic beads to yield polyA mRNA. The mRNA was randomly fragmented and converted to cDNA. After purification and adaptor ligation, fragments of different sizes were selected using AMPure XP beads. PCR amplification was performed for library construction. Finally, the qualified and quantified sample libraries were sequenced using the Illumina novaseq 6000 platform. Three biological replicates were used for each sample.Reads mapping and analysis
-
The output raw data from sequencing were initially filtered to remove adaptor tags, low-quality reads or impurities to obtain clean data. Clean reads were aligned with the reference genome sequences. The zoysia genome sequences were downloaded from the website of Hamygrass genome (
http://zoysia.kazusa.or.jp/ ). Clean reads of the sequencing samples were aligned with the designated reference genome sequence using HISAT2 (v2.0.5) to obtain the specific position information on the reference genome and the characteristics of the sample sequence itself. The correlation of gene expression levels between samples was measured using the Pearson correlation coefficientand material groups with R2 less than 0.8 were excluded. Differentially expression analysis was performed using a combination of Deseq2[24] and edgeR[25]. In this study, the raw count matrix was used as the input, and |log2 fold_change| > 1 with a false discovery rate (FDR) ≤ 0.01 were taken as the cutoff value. The two software were intersected to improve reliability. Annotation and functional classification
-
The GO gene annotation of Zoysia was obtained from NCBI (
www.ncbi.nlm.nih.gov )[26]. KEGG annotations were obtained from KEGG (www.kegg.jp )[27]. An R package 'clusterProfiler'[28] was implemented for the enrichment analysis. Specially, the 'enricher' function for GO enrichment and 'enrichKEGG' function for KEGG enrichment were used. The KEGG pathways were visualized using the R package 'path view'[29]. The gene expression patterns of each pairwise comparison (FM2-vs-FM2, SM2-vs-SM0, and FM2-vs-SM232, etc.) were analyzed. The clustering results were illustrated using heat maps by the R package 'complexHeatmap'[30].Plant hormone-targeted metabolomics analysis of Hamhaelis based on MRM (multiple reaction monitoring)
-
The standard was diluted with a methanol aqueous solution as the standard working solution for a series of concentrations. The labeling curve was established using the isotope internal standard method, and the metabolites were extracted. The samples were separated using a Water I-Class LC ultra-efficient liquid chromatography system. Mass spectrometry was performed by using a 5500 QTRAP mass spectrometer (AB SCIEX) in the positive/negative ion mode. The peak area and retention time were extracted using Multiquant software (v3.0). The phytohormone content in the samples was calculated based on the standard curve.
Statistical analysis
-
Data analysis was conducted using Statistical Package Statistix 8.1 (Tallahassee, FL, USA). The data were statistically evaluated by conducting a two-way analysis of variance. Differences between mean values for each parameter were distinguished by the least significant difference (LSD) test at the 0.5 probability level.
-
To screen out the Z. japonica materials with the greatest difference in regeneration rate, growth rate, leaf width, and number of stolons of 10 Zoysia japonica materials after mowing were measured. The results showed that FM9 had the fastest growth rate, SM23 had the widest leaf width (0.38 cm) and FM1 had the most number of stolons (2) (Fig. 1). FM1, which has the largest number of stolons and a relatively fast growth rate (0.84 cm/d), was chosen as the material with a higher regeneration rate in subsequent experiments. SM232, which has the slowest growth rate (0.53 cm/d) and the least number of stolons was chosen as the material with a slower regeneration rate.
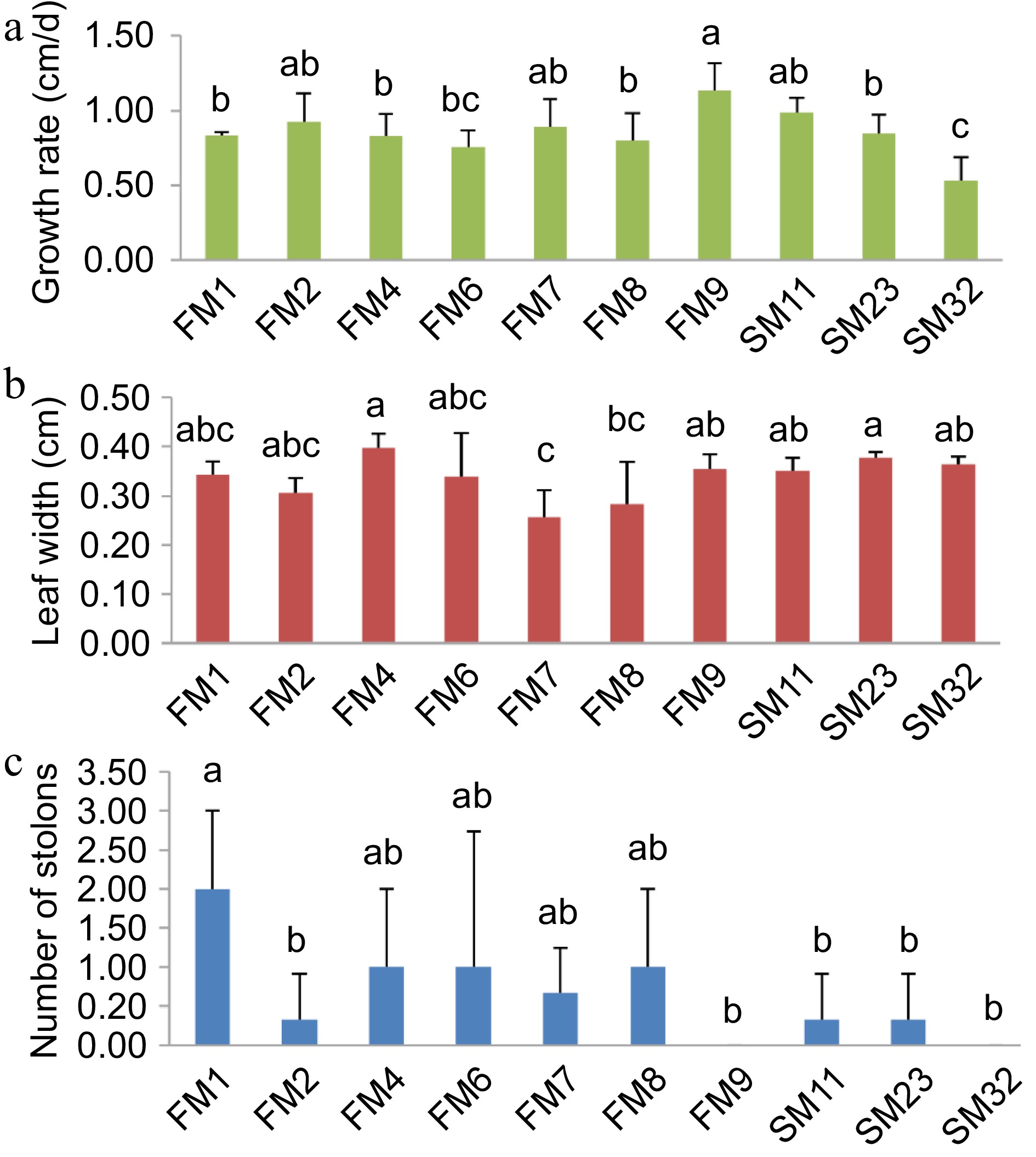
Figure 1.
Growth of different Zoysia grass. (a) Zoysia grass growth rate. (b) Zoysia grass leaf width. (c) Number of Zoysia grass stolons. Error bars represent ± SD. Significance analysis was performed with 1-way ANOVA (p < 0.05).
Morphological verification of regeneration time point of Z. japonica after mowing
-
Regeneration of leaf primordia and new tillers of FM1 and SM232 biological sections were observed at different times (Fig. 2). The apical meristem of FM1 appeared and grew at 2 h after mowing (Fig. 2b) while that of SM232 occurred at 12 h after mowing (Fig. 2g). At 6 h after mowing, not only leaf primordia but also new tillers appeared in the apical meristem of FM1 (Fig. 2c). At 24 h after mowing, the leaf primordia began to appear in SM232 (Fig. 2h). New tillers also appeared at 48 h after mowing (Fig. 2i). Combined with morphological and microstructural changes in the apical meristem, it can be speculated that FM1-6h may be the stage of new organogenesis, first forming leaf primordia and then developing into leaves. SM232 may be in the stage of new organogenesis within 24 h after mowing, first forming leaf primordia and then gradually developing into visible new leaves. After 24 h, it is the stage of rapid growth and development of the plant. Both the number of leaf primordia and new tillers of FM1 were significantly higher than those of SM232.
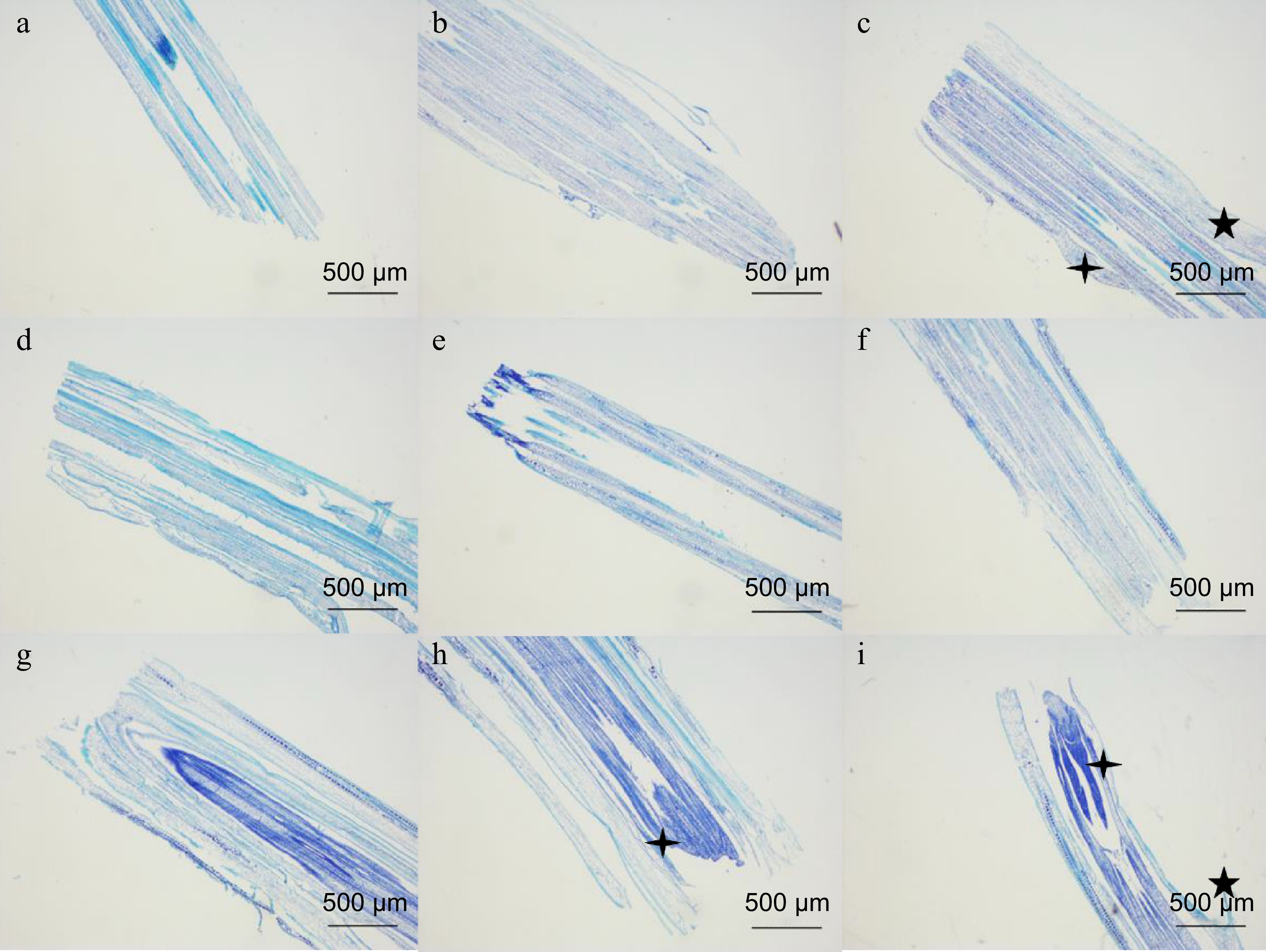
Figure 2.
Microstructural changes of tip meristems after mowing of FM1 and SM232. (a)−(c) Tip meristems at 0, 2, and 6 h after FM1 mowing. (d)−(i) Tip meristems at 0, 2, 6, 12, 24, and 48 h after SM232 mowing.


Transcriptomic analysis of the Z. japonica regeneration process
-
The insert size of cDNA library samples was detected using Agilent 2100 Bioanalyzer and the cDNA libraries were sequenced on the Illumina platform. The threshold of R2 ≥ 0.8 was used to determine whether the genes were differentially expressed (Supplemental Fig. S1). The raw data were screened to remove the low-quality results and adaptors, 62 747 913 clean reads were generated in total. The data were then mapped to the Zoysia grass genome, and more than 96% of the reads were successfully aligned with the reference genome. The entire experimental trials were repeated three times.
After removing the samples with large differences, cluster analysis of differentially expressed genes was performed (Fig. 3). In FM1, it was sharply upregulated at 2 h after mowing (FM1-2h), and eventually tended to be the same as the expression level immediately after mowing (FM1-48h). In SM232, which has a slower growth rate, the trend of gene changes gradually became obvious at 6 and 12 h after mowing (SM232-6h and SM232-12h). The change pattern of some genes was the same as that of FM1, reaching the maximum at 12 h after mowing (SM232-12h). The DEGs were filtered via DESeq with padj < 0.05. According to these criteria, numerous DEGs of FM1 and SM232 were obtained from different pairwise comparisons and displayed in the form of Venn diagrams (Fig. 4a, b). Combined with the KEGG plots of FM1 and SM232 at different mowing periods, the periods with the largest change in the number of differential genes were FM1-12h and SM232-6h, respectively (Figs 5, 6).
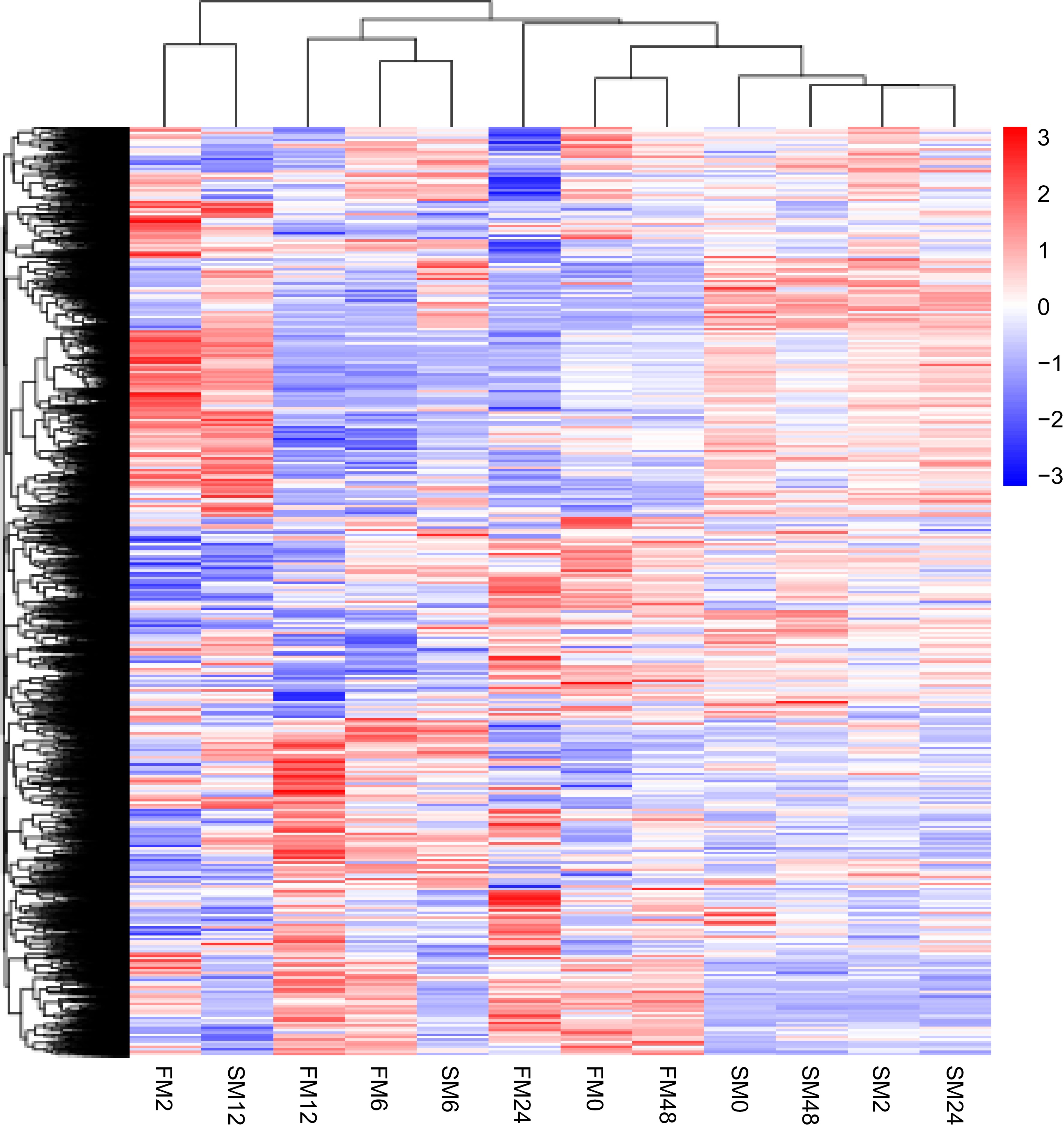
Figure 3.
Clustering of DEGs based on pairwise comparison among the 12 libraries (FM1-2h, FM232-6h, SM232-2h etc.). The fold changes of the DEGs expression levels are hierarchically clustered and shown in a heat-map. The expression level differences are represented by a color scale from blue (down-regulated expression) to red (up-regulated expression), as indicated by the scale bar. Made by novomagic (
https://magic.novogene.com ).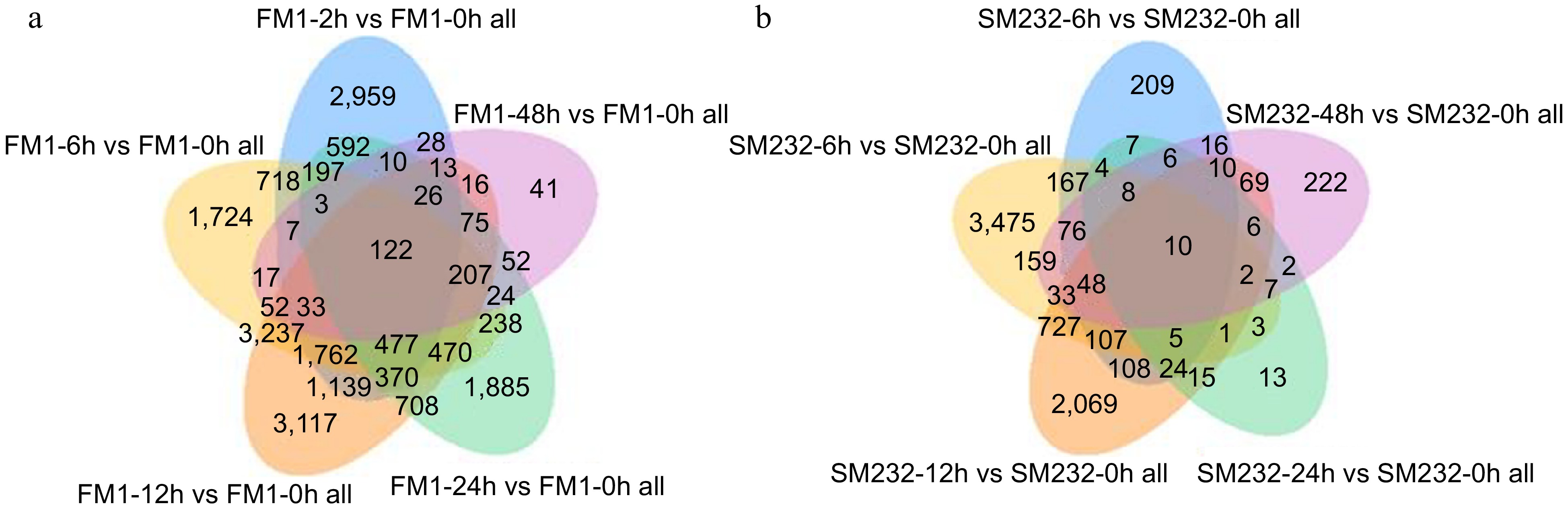
Figure 4.
Venn diagram of differential genes of FM1 and SM232 at different times after mowing. (a) Number of differential genes of FM1 at 2, 6, 12, 24, and 48 h after mowing. (b) Number of differential genes of SM232 at 2, 6, 12, 24, and 48 h after mowing.
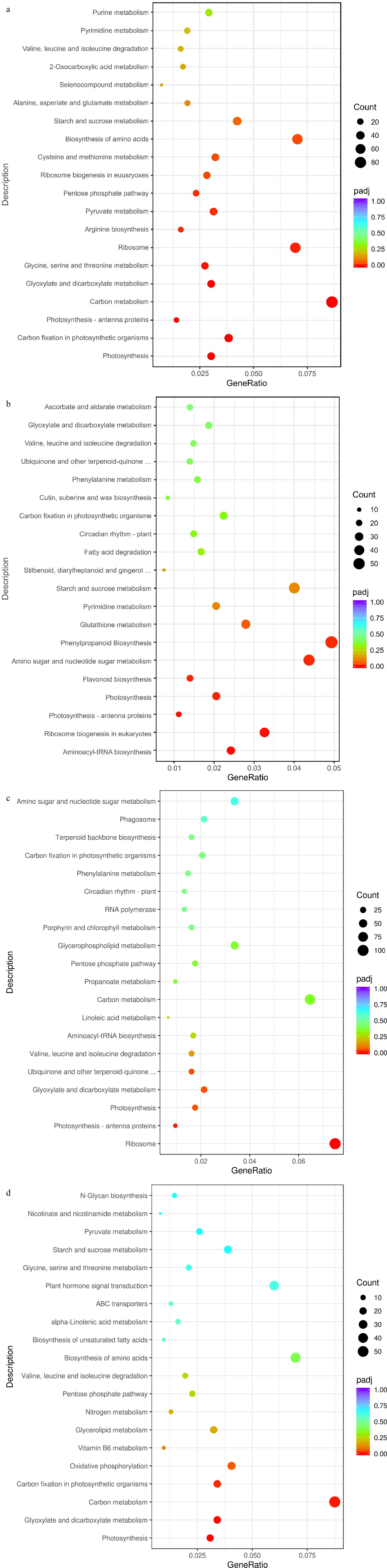
Figure 5.
KEGG enrichment bubble plots of differentially expressed genes at different times after mowing in FM1. (a) KEGG pathway analysis of DEGs at FM1-2h compared to FM1-0h. (b) KEGG pathway analysis of DEGs at FM1-6h compared to FM1-0h. (c) KEGG pathway analysis of DEGs at FM1-12h compared to FM1-0h. (d) KEGG pathway analysis of DEGs at FM1-24h compared to FM1-0h.
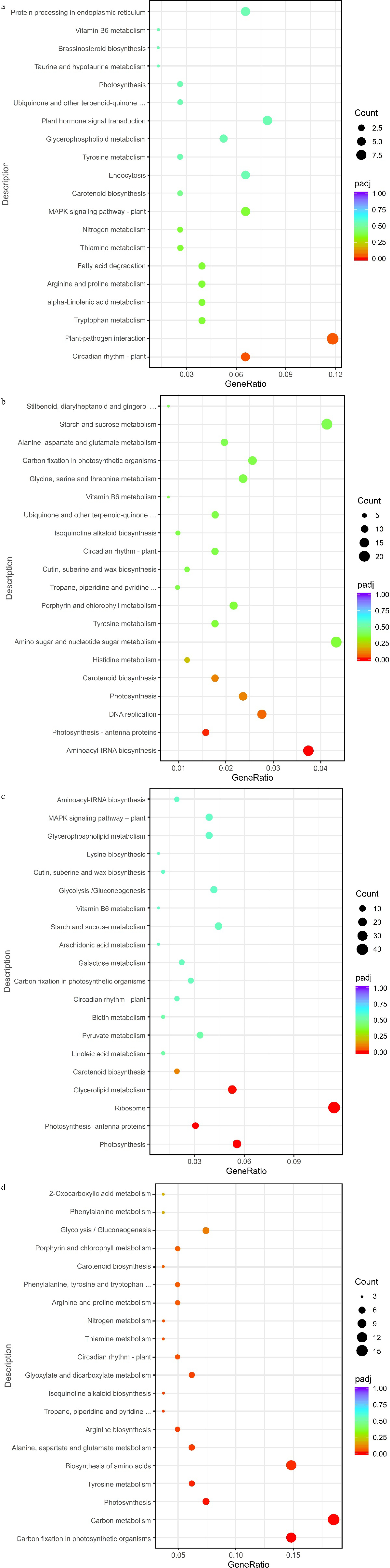
Figure 6.
KEGG enrichment bubble plots of differentially expressed genes at different times after construction in SM232. (a) KEGG pathway analysis of DEGs at SM232-2h compared to SM232-0h. (b) KEGG pathway analysis of DEGs at SM232-6h compared to SM232-0h. (c) KEGG pathway analysis of DEGs at SM232-12h compared to SM232-0h. (d) KEGG pathway analysis of DEGs at SM232-48h compared to SM232-0h.
To further identify the functions of DEGs after mowing in FM1, the annotated sequences were mapped to the reference classical pathways in the Kyoto Encyclopedia of Genes and Genomes (KEGG) (Fig. 5). The enrichment pathways of FM1-2h were mainly related to primary metabolisms, such as carbon metabolism, starch sucrose metabolism, glycolysis, etc.; related to plant hormone signal transduction; related to secondary metabolisms, such as amino acid biosynthesis, cysteine and methionine metabolism, alanine metabolism, etc.; related to photosynthesis, carbon fixation in the photosynthetic system, photosynthesis, etc.; and some related to protein synthesis, such as ribosomes, protein processing in the endoplasmic reticulum, RNA transport, etc.
A total of 3,237 annotated DEGs of SM232-12h were highly enriched in GO categories, including peptide metabolic process (GO:0006518), cellular amide metabolic process (GO:0043603), translation (GO:0006412), peptide biosynthetic process (GO:0043043), amide biosynthetic process (GO:0043604), intracellular ribonucleoprotein complex (GO:0030529), non-membrane-bounded organelle (GO:0043228), and intracellular non-membrane-bounded organelle (GO:0043232). The enrichment pathways were mainly related to ribosomes, photosynthesis, antenna proteins, and glycerol metabolism in photosynthesis (Fig. 6c). GO enrichment analysis and KEGG clustering analysis of both materials revealed that FM1 after 2 h of mowing showed the same enrichment pathway as SM232 after 12 h of mowing (Figs 5, 6). Therefore, further analysis was conducted on the two materials, FM1-2h and SM232.
The volcano plot summarized the up- and down-regulation of genes represented by red and green dots (Supplemental Fig. S2a, c). The biological functions of these DEGs during the regeneration process were annotated with Gene Ontology (GO) categories; 8,456 (FM-2h vs FM-0h) DEGs were categorized into 30 functional groups belonging to biological process, cellular component, and molecular function, respectively (Supplemental Fig. S2b, d), and all the annotated DEGs were presented in Online Resource. In FM1, these annotated DEGs were highly enriched in GO categories including transmembrane transporter activity (GO:0022857), non-membrane-bounded organelle (GO:0043228), intracellular non-membrane-bounded organelle (GO:0043232), cellular amide metabolic process (GO:0043603), amide biosynthetic process (GO:0043604), peptide metabolic process (GO:0006518), translation (GO:0006412) and peptide biosynthetic process (GO:0043043).
Hormonal changes in the regeneration process
-
The hormone (IAA and CTK) contents in the two materials were determined. The IAA content in FM1 was significantly higher than that in SM232 (Fig. 7a). For both materials, the IAA content was highest at 0 h, and the IAA content decreased 2 h after mowing and increased again at 6 h after mowing. For SM232, the IAA content was highest at 12 h after mowing and then gradually decreased. In contrast, the IAA content of FM1 was lowest at 12 h after mowing and then increased. There was no significant difference in the CTK content between the two materials. The iP and iPR contents in SM232 were higher than those in FM1 (Fig. 7b, c).
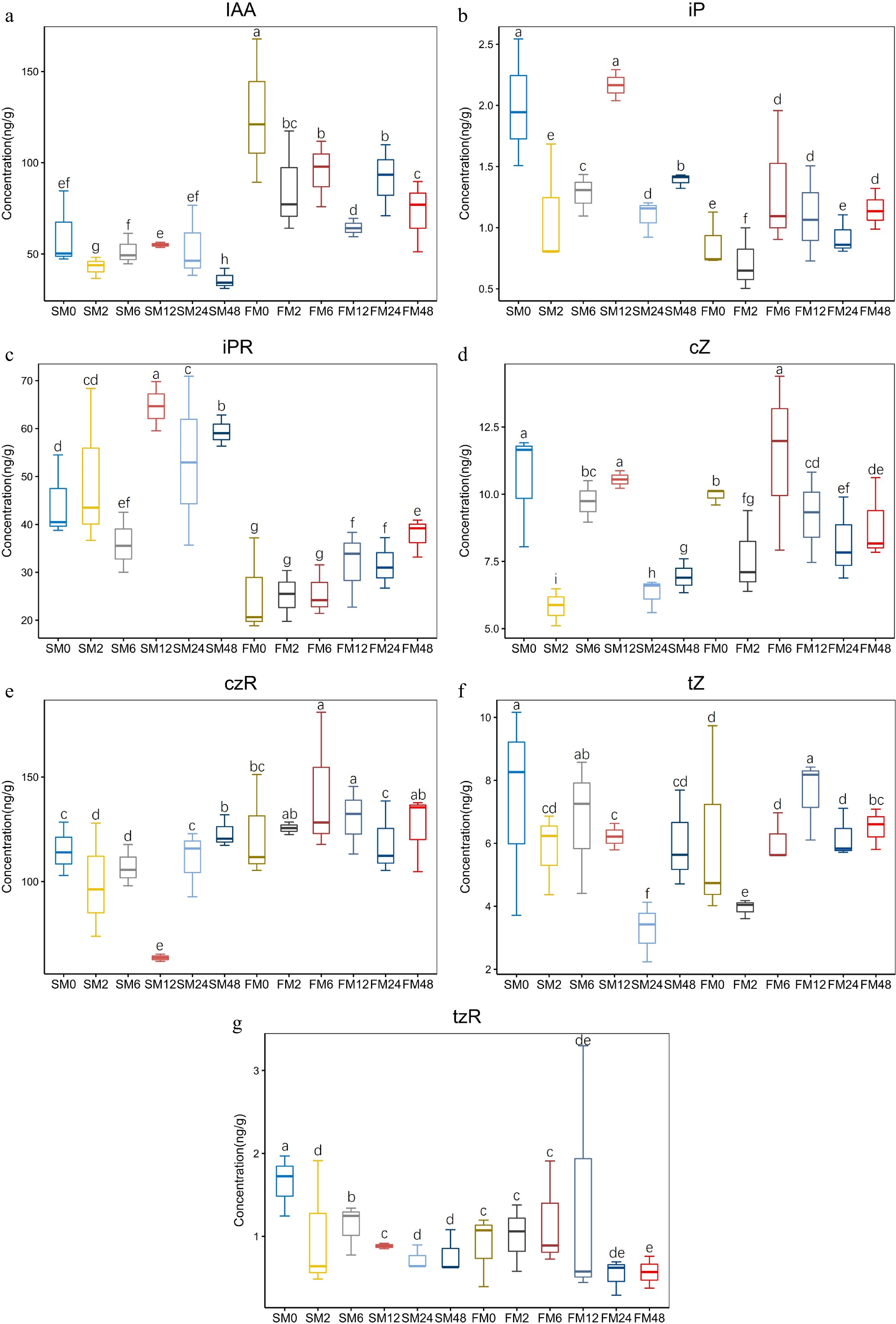
Figure 7.
Trends of IAA and CTK in FM1 and SM232 after mowing. (a) IAA content of FM1 and SM232 after mowing. CTK content of FM1 and SM232 after mowing. (b) iP, (c) iPR, (d) cZ, (e) czR, (f) tZ, (g) tzR.
The expression of hormone-related genes in the two materials also changed. In FM1, the expression of both IAA-related and CTK-related genes changed (Fig. 8a, b), while in SM232, only the expression of growth hormone-related genes was found (Fig. 8c), and no changes in the expression of CTK-related genes were found. The genes with the greatest changes in expression in both groups of material were the AUX/IAA family genes. There were 17 AUX/IAA family genes were identified in FM1 (Fig. 8a) and 10 AUX/IAA family genes were identified in SM232 (Fig. 8c). The expression of these genes did not change much in the short-term (2 h) and long-term (48 h) time after mowing. Most AUX/IAA family genes were downregulated at 6 h and 12 h after mowing, including 12 (at 6 h) and 9 (at 12 h) in FM1, 6 (at 6 h), and 2 (at 12 h) in SM232, respectively. The gene Zjn_sc00014.1.g07330.1.am.mk showed significant down-regulation in both materials. However, a few genes were up-regulated at 6 h and 12 h after mowing, including two in FM1 and one in SM232 and the gene Zjn_sc00011.1.g02910.1.sm.mkhc showed significant up-regulation in both materials. In addition, more unusual was the gene Zjn_sc00011.1.g06110.1.sm.mkhc, which was down-regulated at 6 h after mowing but up-regulated at 12 h after mowing. Gene Zjn_sc00011.1.g06110.1.sm.mkhc was also up-regulated in FM1 but not in SM232. At 24 h after mowing, the expression of most genes returned to the initial state, but the expression of a few genes was up-regulated, three in FM1 and one in SM232, including the previously mentioned gene Zjn_sc00011.1.g02910.1.sm.mkhc.
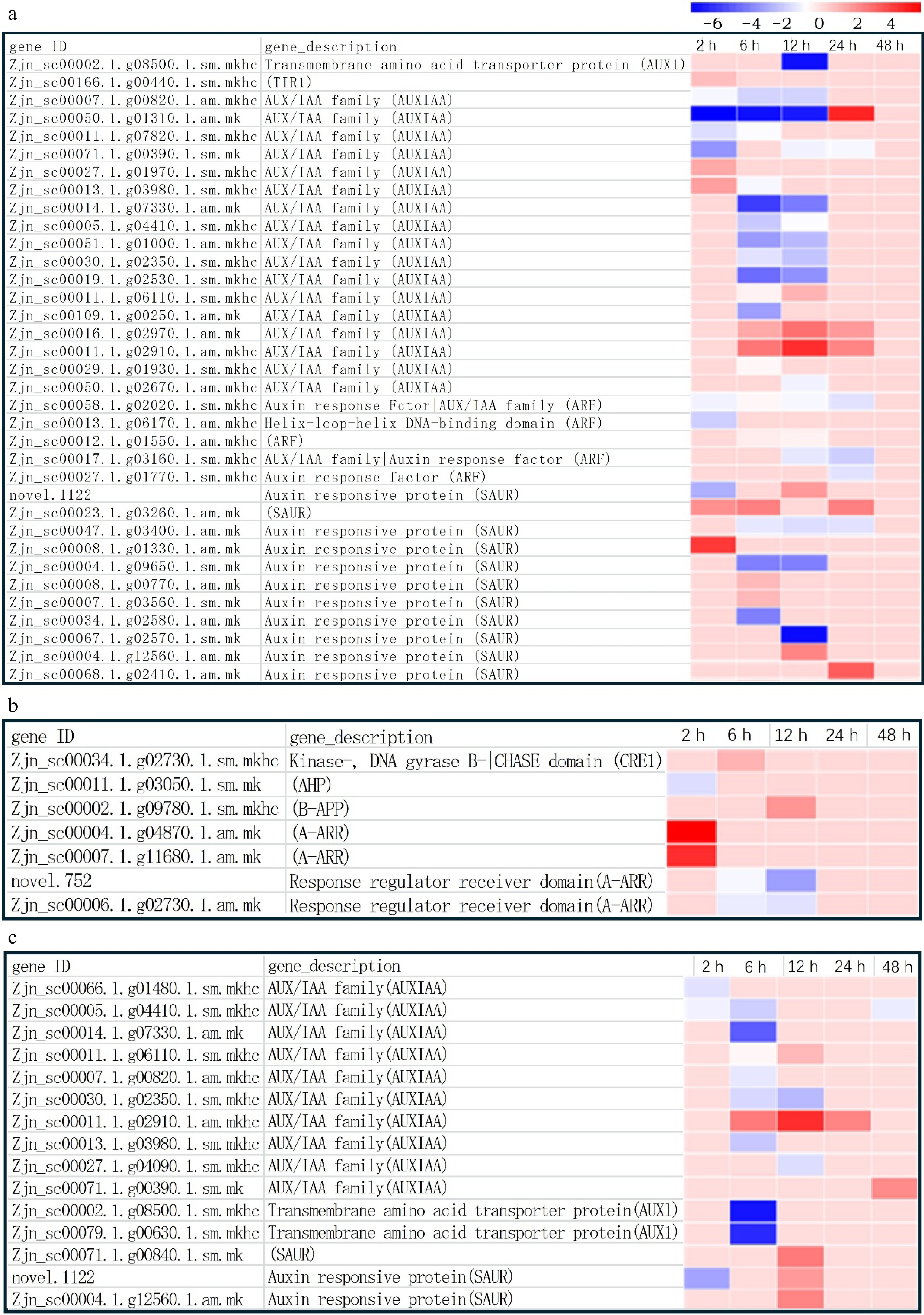
Figure 8.
Gene expression of IAA- and CTK-related hormones in FM1 and SM232. (a) Changes in IAA-related gene expression at different times after FM1 mowing. (b) Changes in CTK-related gene expression at different times after FM1 mowing. (c) Changes in IAA-related gene expression at different times after SM232 mowing.
In addition, some SAUR family genes were identified in both materials, 11 SAUR family genes were identified in FM1 (Fig. 8a), and three SAUR family genes were identified in SM232 (Fig. 8c). The changes in the expression of the SAUR family genes were also mainly concentrated in the period of 6 h or 12 h, while there were no changes in the short term (2 h) and long term (48 h). AUX1 showed a significant down-regulation trend in both materials, but in FM1 the down-regulation time was at 12 h after mowing, while in SM232 the down-regulation time was at 6 h after mowing (Fig. 8a, c). In addition, five ARF family genes were identified in FM1, belonging to the growth factor response factors, growth factor response proteins, AUX/IAA family, and helical-loop DNA-binding domains, and the ARF family genes were in a down-regulated state at different stages after mowing, respectively.
Changes in the expression of CTK-related genes were only detected in FM1 (Fig. 8b). Unlike IAA-related genes, CTK-related genes showed more obvious changes in short term (2 h), with two genes significantly upregulated and one gene downregulated. In addition to these three genes that changed at 2 h after mowing, another four genes were upregulated or downregulated at 6 h and 12 h after mowing. The expression of these genes was upregulated at 6 h and 12 h, respectively. However, all these genes did not change in expression in the long term (24 h and 48 h).
-
Z. japonica is widely used in the establishment of neighborhood and park landscaping, sports field lawns, leisure lawns and soil preservation lawns, etc[1]. However, the relatively slow growth rate of Zoysia grass translates to poor recuperative potential, which often limits its use for sports fields where the traffic injury is quite frequent. In this study, FM1 and SM232 were selected from nine kinds of Zoysia materials with the largest differences in growth rate after mowing. Transcriptomes sequencing was performed on them at 0, 2, 6, 12, 24, and 48 h after mowing, and the data were compared. The contents of auxin and CTK were determined, and biological sections were observed. The GO and KEGG analysis results showed that Zoysia is frequently involved in metabolism, amino acid transport, protein translation, ribosome transport generation, cell wall, cell membrane, and capsule formation in the biological process of responding to mowing processing, and the timing and metabolic emphasis of responding to stress after mowing treatment varies.
Proteins defined by the conserved lateral organ boundaries (LOB) domain are key regulators of plant organ development and are expressed in a group of cells at the proximal axis and lateral roots of all lateral organs[31]. Previous studies have shown that LOB is involved in the formation and development of lateral organ boundaries, and the number of lateral organs in plants overexpressing LOB 7 increased significantly[32]. The successive differentiation and development of lateral organs directly affects the regeneration and establishment rate of turfgrass. In this study, FM1 had a large number of LOBs family protein-related genes upregulated after mowing, especially at 6, 12, and 24 h after mowing. However, the expression of LOBs-related genes in SM232 material showed a continuous downward trend, suggesting that the regeneration of Zoysia grass after mowing might be closely related to the expression of LOB family proteins. Combined with the results of biological sections, it is speculated that the initiation of lateral primordia and the morphogenesis of lateral organs might have occurred in FM1 after 2 h of mowing.
Based on the changes in gene expression and related metabolic activities, it is speculated that the regeneration of FM1 occurred at 6 h after mowing and reached a climax at 12 h, whereas the regeneration of SM232 plants might occur after 24 or 48 h. The plant's own defense and repair responses were also slow, so it was speculated that the regeneration process of SM232 might be slow, and the main reason for the slow regeneration process might be closely related to the expression of CTK-related genes. The main reason for the slow regeneration process of SM232 may be related to the expression of cytokinin-related genes. Forty-eight hours after FM1 mowing, related genes such as isocitratelyase, stress proteins, and ketoreduction family proteins were all upregulated. It was speculated that the plant began to store energy, metabolism gradually returned to normal, and mowing stress was gradually restored[33]. Within 48 h after mowing, SM232 was always in the self-repair period, and the photosynthetic system and energy metabolism were recovered to a certain extent.
IAA and CTK regulate cell elongation, cell division, and the establishment and maintenance of their synthesis, transport, and signal transduction pathways[34,35]. Studies have shown that regeneration is initiated by the rapid accumulation of auxin near the injury site[36]. A study in Triticum aestivum found that the decreased auxin content after mowing relieved the inhibition of cytokinin synthesis, which controls the transition from cell division to cell expansion and stimulates cell expansion and differentiation during the cell expansion phase, and eventually accelerates post-mowing regeneration of seedlings[37]. The decrease in auxin concentration and the increase in cytokinin concentration may accelerate the regeneration of Triticum aestivum seedlings after harvest. In the present study, according to the morphological observation, leaf primordia and new tillers appeared at 6 h after mowing in FM1, while the IAA content and CTK content increased at 6 h after mowing. Leaf primordia began to appear at 24 h after mowing in SM232, while the IAA content and CTK content remained at a high level at 24 h after mowing (Fig. 7). It was speculated that the changes of IAA and CTK content may have a positive effect on the formation of leaf primordia. Higher concentrations of CTK can promote bud differentiation[38].
The genes with the largest variation in gene expression levels in both materials were AUX/IAA family genes. AUX/IAA is a transcriptional repressor that provides a transduction pathway for auxin signaling, which is widely involved in the regulation of plant growth and development[39]. In total, 17 AUX/IAA family genes were identified in FM1, and 10 AUX/IAA family genes were identified in SM232. The expression levels of these genes in FM1 were higher than those in SM232. It is speculated that the regeneration rate of Zoysia grass may be closely related to the AUX/IAA family genes.
SAUR is an early response gene, the most rapid and intense auxin gene, which is involved in the regulation of a wide range of cellular, physiological, and developmental processes[40]. SAURs are key effector outputs of hormonal and environmental signals that regulate plant growth and development[41]. SAURs activate the plasma membrane (PM) H+-ATPases and promote cell expansion by inhibiting PP2C.D phosphatase[41]. Arabidopsis SAUR63 can stimulate the elongation of different organs by genes related to auxin[42]. In maize, ZmSAUR2 is involved in auxin mediation, while SAUR41 regulates cellular of auxin[43], and the rice SAUR39 gene is involved in the synthesis and transport of auxin[44]. In this study, 12 SAUR family genes were identified in FM1, but only three SAUR family genes were identified in SM232. For SM232, all three SAURs genes were upregulated and appeared at 12 h after mowing. ARF (auxin response factors) are important transcription factors that transmit auxin signals, express functional genes, and are auxin response functional centers in plants[45].
The ARF family genes annotated in this study belong to the auxin response factor, auxin-responsive protein, AUX/IAA family, and spiral circular DNA binding domain, respectively, and the gene expression was downregulated at each stage. ARF generally expressed at different stages of plant growth and development[46], and the temporal change in ARF expression in FM1 is consistent with this rule. It is possible that the downregulation of ARF family genes and the reduced binding of AUX/IAA to ARF, which leads to a gradual decrease in auxin levels in plants under lower auxin levels.
-
This study conducted a time-course transcriptomic analysis of two Z. japonica genotypes at 0, 6, 12, and 24 h after mowing to obtain the overall molecular regulation mechanism. The physiological performance of the FM1 genotype within 24 h after mowing was faster than that of the SM232 genotype. In terms of the transcriptome results, dynamic DEGs analysis showed that the genotypes FM1 and SM232 were significantly different in cellular components, molecular functions, and biological processes. In addition, some candidate genes related to the regeneration process were emphasized, and their roles in the regeneration process needs to be further elucidated. Changes in the expression of growth hormone and CTK-related genes were identified in the FM1 mowing treatment, while only changes in the expression of growth hormone-related genes were only identified in SM232. In summary, the results of this study enriched the transcriptomic information, improved the understanding of the regeneration regulation mechanism of Z. japonica, and also provided important clues for turfgrass species selection and lawn maintenance.
-
The authors confirm contribution to the paper as follows: study conception and design: Sun Y, Hu Q; data collection: Xiao K; data curation: Li Y; analysis and interpretation of results: Xiao K, Wang L, Li Y; draft manuscript preparation: Wang L, Hu Q, Zi Z; funding acquisition: Sun Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This research was funded by the National Center of Technology Innovation for Comprehensive Utilization of Saline-Alkali Land.
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Correlation between the transcriptome samples.
- Supplemental Fig. S2 Differential genes at 2 h after FM1 pruning and 12 h after SM232 pruning.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang L, Xiao K, Li Y, Zi Z, Sun Y, et al. 2024. Transcriptional regulation and hormone action analysis in the regeneration process of Zoysia japonica after mowing. Grass Research 4: e018 doi: 10.48130/grares-0024-0016 

Transcriptional regulation and hormone action analysis in the regeneration process of Zoysia japonica after mowing
- Received: 26 May 2024
- Revised: 25 July 2024
- Accepted: 05 August 2024
- Published online: 28 August 2024
Abstract: Zoysia grass (Zoysia japonica) is an important native grass species in China, while the slow regeneration speed limits its use. Two Z. japonica materials FM1 (fast regeneration rate) and SM232 (slow regeneration rate) were chosen from 10 different Zoysia grass by phenotypic index in this study. The genetic differences associated with zoysia regeneration and metabolism pathway are elucidated based on the transcriptome analysis, determination of hormone, and molecular biological section. Transcriptome sequencing results showed that both FM1 and SM232 had the maximum number of DEGs at 2 h and 12 h after mowing, indicating it may be the key stage of Zoysia grass response to mowing. After mowing treatment, FM1 was accompanied by up-regulation of regeneration-related genes, such as the LOB protein family and amide synthesis family. However, SM232 showed a continuous down-regulation of protein-related genes of the LOB family. Transcriptome analysis showed that the expression of IAA-related genes in both groups showed a downward trend, which was consistent with the results of IAA content determination. The IAA content in FM1 was significantly higher than that in SM232. Changes in CTK-related genes were only detected in FM1. Mowing induces plant defense systems, and plant tissue regeneration is closely related to the plant defense system, which may be the focus of further research. These results provide a basis for further research on the molecular mechanism of grass regeneration.
-
Key words:
- Zoysia japonica /
- Transcriptome analysis /
- Auxin /
- Cytokinin /
- Regeneration