









-
Plants are continuously subjected to unpredictable environmental conditions and encounter a multitude of stressors throughout their growth and development, posing a significant challenge to global crop production and food security[1]. Heat and drought are undoubtedly the two most important stresses that have a huge impact on crops. Both elicit a wide array of biochemical, molecular, and physiological alterations and responses, impacting diverse cellular processes and ultimately influencing crop yield and quality[2].
A primary physiological consequence of both stresses is the diminished photosynthetic capacity, partially resulting from the degradation of chlorophyll due to leaf senescence under stress conditions. Chlorophyll accumulation was diminished in numerous plants subjected to drought or heat stress conditions[3,4]. Various environmental stresses prompt excessive generation of reactive oxygen species (ROS), initiating oxidative damage that compromises lipids, and proteins, and poses a serious threat to cellular functions[2]. To mitigate oxidative stress and minimize damage, plants have developed various protective mechanisms to neutralize ROS. Several antioxidant enzymes, such as SOD, POD, and CAT, are integral to cellular antioxidative defense mechanisms. Additionally, antioxidants such as anthocyanins and proline serve as crucial ROS scavengers[5,6]. The elevation in temperature typically induces the transient synthesis of heat shock proteins (Hsps), which function as molecular chaperones in protecting proteins from denaturation and aggregation, with their activity primarily regulated at the transcriptional level by heat shock factors (Hsfs)[7]. The significance of Hsps and Hsfs in all organisms, including plants, has been assessed in various stress conditions that could disrupt cellular homeostasis and result in protein dysfunction[7]. Drought stress can also trigger the transcription of a suite of marker genes, including RD29A, RD29B, NCED3, AREB1, Rab18, etc., which assist plants in mitigating cellular damage during dehydration and bolstering their resilience to stress[8−10].
Previous research efforts focusing on the regulatory control of stress-related genes have largely centered around protein-coding genes. In recent years, non-protein-coding transcripts have emerged as important regulatory factors in gene expression. Among them, long non-coding RNAs (lncRNAs) lncRNAs have been identified as implicated in various abiotic stresses[11,12]. LncRNAs are a class of non-coding RNAs (ncRNAs) exceeding 200 nucleotides in length. They possess minimal or no protein-coding potential[13]. In plants, lncRNAs are specifically transcribed by RNA polymerases Pol IV, Pol V, Pol II, and Pol III[14,15]. LncRNAs exhibit low abundance and display strong tissue and cellular expression specificity relative to mRNAs. Moreover, sequence conservation of lncRNAs is was very poor across different plant species[13,16,17]. The widespread adoption of high-throughput RNA sequencing technology has revealed lncRNAs as potential regulators of plant development and environmental responses. In cucumber, RNA-seq analysis has predicted 2,085 lncRNAs to be heat-responsive, with some potentially acting as competitive endogenous RNAs (ceRNAs) to execute their functions[18]. In radish, a strand-specific RNA-seq (ssRNA-seq) technique identified 169 lncRNAs that were differentially expressed following heat treatment[19]. In Arabidopsis, asHSFB2a, the natural antisense transcript of HSFB2a was massively induced upon heat stress and exhibited a counteracted expression trend relative to HSFB2a. Overexpression of asHSFB2a entirely suppressed the expression of HSFB2a and impacted the plant's response to heat stress[20]. For drought stress resistance, 244 lncRNAs were predicted in tomatoes to be drought responsive probably by interacting with miRNAs and mRNAs[21]. Under drought stress and rehydration, 477 and 706 lncRNAs were differentially expressed in drought-tolerant Brassica napus Q2 compared to drought-sensitive B. napus, respectively[22]. In foxtail millet and maize, 19 and 644 lncRNAs, respectively, were identified as drought-responsive[23,24]. Despite the identification of numerous lncRNAs by high-throughput sequencing, which suggests their potential involvement in various abiotic stress processes, only a minority have been experimentally validated for function.
In our previous study, we characterized 1,229 differentially expressed (DE) lncRNAs in Chinese cabbage as heat-responsive, and subsequent bioinformatics analysis reduced this number to 81, which are more likely associated with heat resistance[25]. lnc000283 and lnc012465 were selected from among them for further functional investigation. The findings indicated that both lnc000283 and lnc012465 could be promptly induced by heat shock (HS). Overexpression of either lnc000283 or lnc012465 in Arabidopsis plants enhanced their capacity to tolerate heat stress. Additionally, both lnc000283 and lnc012465 conferred drought tolerance to transgenic Arabidopsis.
-
The lncRNA sequences examined in this study were from Chiifu-401-42 Chinese cabbage and all Arabidopsis plants were of the Col-0 background. Transgenic plants expressing lnc000283 and lnc012465 were generated using the Agrobacterium tumefaciens-mediated floral dip method[26]. Single-copy and homozygous T3 plants were identified through genetic segregation on an agar medium supplemented with kanamycin. The T3 generation plants, or their homozygous progeny, were utilized in the experiments.
For phenotypic assessment, Arabidopsis seeds were initially sown on filter paper moistened with ddH2O and placed in a 4 °C freezer for 2 d. Subsequently, they were evenly planted in nutrient-rich soil and transferred to a growth chamber operating a 16-h day/8-h night cycle, with day/night temperatures of 22 °C/18 °C and a light intensity of 250 μmol·m−2·s−1. After 10 d of growth, Arabidopsis plants with uniform growth were transferred to 50-hole plates. Arabidopsis plants grown in Petri dishes were firstly seed-sterilized and then sown on 1/2 MS medium supplemented with 10 g·L−1 sucrose. The seeds were then placed in a 4 °C refrigerator for 2 d in the dark before transferring them to a light incubator. The day/night duration was set to 16 h/8 h, the day/night temperature to 21 °C/18 °C, and the light intensity to 100 μmol·m−2·s−1.
For heat treatment, 3-week-old seedlings were subjected to 38 °C for 4 d within a light incubator, subsequently transferred to their original growth conditions under the same light/dark cycles. For drought treatment, 3-week-old Arabidopsis seedlings were deprived of water for 10 d, followed by rehydration to facilitate a 2-d recovery period. Plants were photographed and surveyed both before and after treatment.
Vector construction
-
The lncRNA sequences (lnc000283 and lnc012465) were chemically synthesized based on RNA-seq data, with restriction sites for BamH1 and Kpn1 engineered upstream and downstream. The resultant lncRNA constructs were subcloned into the pCambia2301 binary vector, incorporating a cauliflower mosaic virus (CaMV) 35S promoter. The recombinant vectors were transformed into Escherichia coli TOP10 competent cells (Clontech), incubated at 37 °C overnight, after which single clones were selected for PCR verification, and the confirmed positive colonies were submitted for sequencing. Following verification, the correct plasmids were introduced into A. tumefaciens strain GV3101 using the freeze-thaw method and subsequently transformed into Arabidopsis wild-type (Col) plants.
Chlorophyll and anthocyanin content determination
-
To quantify the chlorophyll content, the aerial portions of wild-type and transgenic Arabidopsis plants, grown in Petri dishes were weighed, minced, and then subjected to boiling in 95% ethanol until fully decolorized. Aliquots of 200 μL from the extract were transferred to a 96-well plate and the absorbance at 663 nm and 645 nm was measured via spectrophotometry by a microplate reader (Multiskan GO, Thermo Scientific, Waltham, MA, USA). Three biological replicates were analyzed for WT and each transgenic line. Chlorophyll content was determined according to the formula of the Arnon method[27]: Chlorophyll a = (12.72A663 − 2.59A645) v/w, Chlorophyll b = (22.88A645 − 4.67A663) v/w, Total chlorophyll = (20.29A645 + 8.05A663) v/w.
The quantification of anthocyanin was performed as follows: aerial parts of wild-type and transgenic Arabidopsis plants, cultivated in Petri dishes, were weighed and ground to powder in liquid nitrogen. Subsequently, the samples were incubated in 600 μL of acidified methanol (containing 1% HCl) at 70 °C for 1 h. Following this, 1 mL of chloroform was added, and the mixture was vigorously shaken to remove chlorophyll. The mixture was then centrifuged at 12,000 rpm for 5 min, after which the absorbance of the aqueous phase was determined at 535 nm using a spectrophotometer (Shimadzu, Kyoto, Japan). Three biological replicates were analyzed for WT and each transgenic line. The relative anthocyanin content was calculated according to anthocyanin concentration and extraction solution volume. One anthocyanin unit is defined as an absorption unit at a wavelength of 535 nm in 1 mL of extract solution. In the end, the quantity was normalized to the fresh weight of each sample.
Physiological analysis
-
Three-week-old transgenic and WT A. thaliana plants, subjected to normal conditions or varying durations of heat or drought stress, were utilized for subsequent physiological assessments. All assays were performed in accordance with the method described by Chen & Zhang[28]. In brief, 0.1 g of fresh leaf tissue was homogenized in 500 μL of 100 mM PBS (pH 7.8) while chilled on ice. The homogenate was then centrifuged at 4 °C, and the resultant supernatant was employed for further analysis. For the determination of MDA content, 100 μL of the supernatant was combined with 500 μL of a 0.25% thiobarbituric acid (TBA) solution (which was prepared by dissolving 0.125 g of TBA in 5 mL of 1 mol·L−1 NaOH before being added to 45 mL of 10% TCA) and boiled for 15 min. Following a 5 min cooling period on ice, the absorbance was measured at 532 nm and 600 nm. The activity of POD was determined as follows: initially, 28 μL of 0.2% guaiacol and 19 μL of 30% H2O2 were sequentially added to 50 mL of 10mM PBS (pH 7.0), after thorough heating and mixing, 1 mL was transferred into a cuvette, then 50 μL of the supernatant was added to the cuvette and the absorbance at 470 nm was monitored every 15 s for 1 min. To determine the proline content, a reaction solution was prepared by mixing 3% sulfosalicylic acid, acetic acid, and 2.5% acidic ninhydrin in a ratio of 1:1:2, then 50 μL of the supernatant was added to 1 mL of the reaction solution, which was then subjected to a boiling water bath for 15 min (the solution turned red after the boiling water bath). Following cooling on ice, the absorbance at 520 nm was recorded. For the quantification of proline, an L-proline standard curve was prepared by dissolving 0, 5, 10, 15, 20, 25, and 30 μg of L-proline in 0.5 mL of ddH2O, followed by the addition of 1 mL of the reaction solution and measuring the absorbance at 520 nm. The proline content in the samples was then determined based on the L-proline standard curve.
RNA isolation and qRT-PCR analysis
-
Total RNA was isolated from the aerial parts of Arabidopsis using the TaKaRa MiniBEST Plant RNA Extraction Kit, followed by purification and reverse transcription using the PrimeScript RT reagent Kit with gDNA Eraser (Takara). The cDNA product was diluted 10 times and real-time PCR was conducted in triplicate for each biological replicate using SYBR PCR Master Mix (Applied Biosystems) on the ABI 7500 system under the following conditions: 98 °C for 3 min, followed by 40 cycles of 98 °C for 2 s and 60 °C for 30 s. The relative expression levels of each gene were normalized against the transcript abundance of the endogenous control UBC30 (At5g56150) and calculated using the 2−ΔCᴛ method. The specific primers employed for qRT-PCR are detailed in Supplemental Table S1.
-
In our prior investigation, dozens of lncRNAs associated with the heat stress response in Chinese cabbage were identified through informatics analysis. Two lncRNAs (lnc000283 and lnc012465) were chosen for genetic transformation in Arabidopsis to elucidate their functions comprehensively. Transcriptome data analysis indicated that the expression of lnc000283 and lnc012465 in Chinese cabbage were both induced by HS. To verify the accuracy, the expression patterns of lnc000283 and lnc012465 were confirmed through quantitative real-time PCR (qRT-PCR), and the results from qRT-PCR were consistent with those obtained from RNA-seq (Fig. 1a). The corresponding homologous genes in Arabidopsis were identified as CNT2088434 and CNT2088742, exhibiting sequence similarities of 88% and 87%, respectively (Supplemental Fig. S1). Subcellular localization predictions using the lnclocator database (
www.csbio.sjtu.edu.cn/bioinf/lncLocator ) suggested that both lncRNAs are localized within the nucleus (Supplemental Table S2). Bioinformatics analysis was conducted using the CPC tool (http://cpc.cbi.pku.edu.cn/ ) indicated that lnc000283 and lnc012465 are noncoding sequences, with coding probabilities of 0.0466805 and 0.0432148, respectively comparable to the well-characterized lncRNAs COLDAIR and Xist, but significantly lower than those of the protein-coding genes UBC10 and ACT2 (Fig. 1b).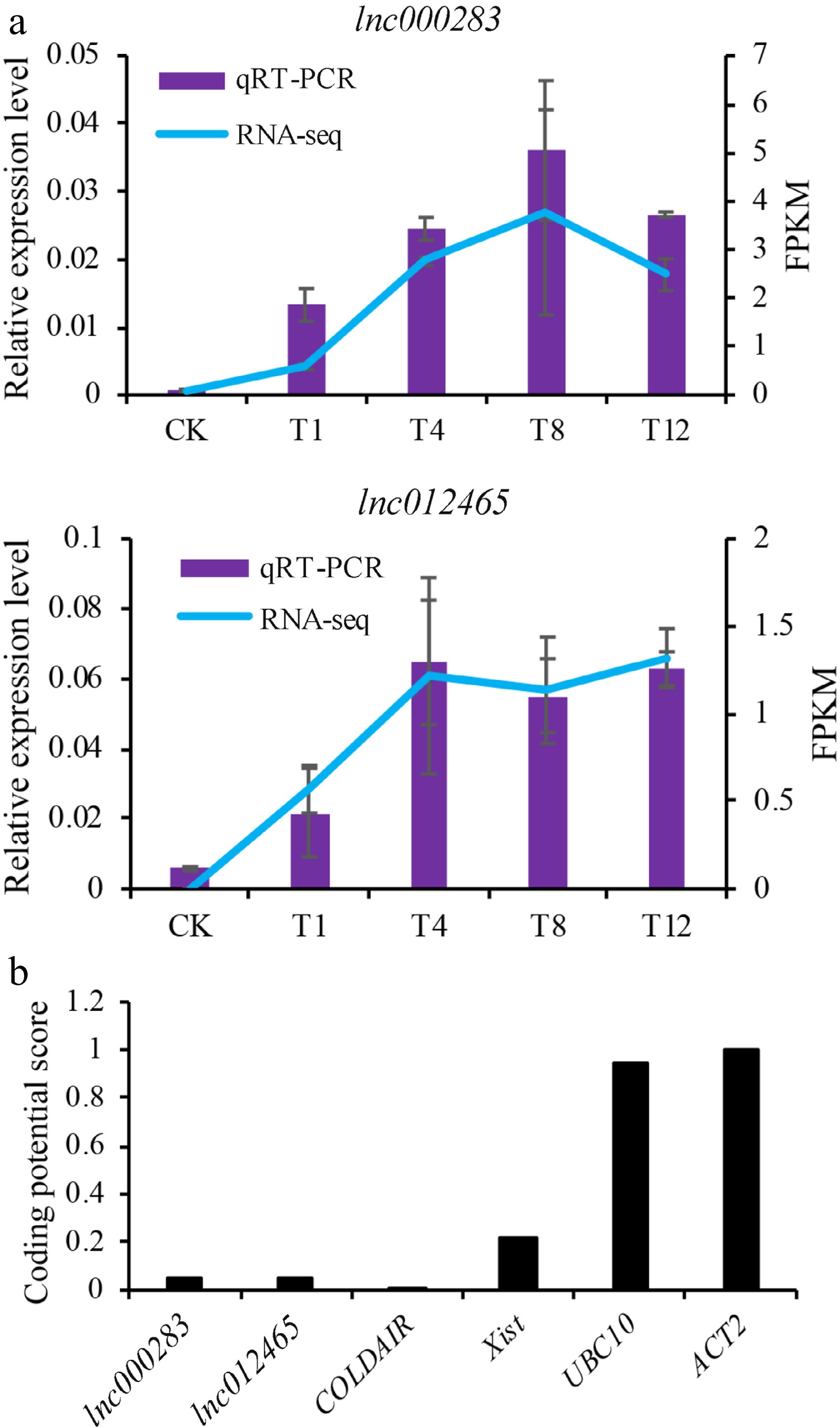
Figure 1.
Characteristics of lnc000283 and lnc012465. (a) Expression level of lnc000283 and lnc012465 in Chinese cabbage leaves treated at 38 °C at different time points, as determined by qRT-PCR and RNA-seq. CK is a representative plant before heating, and T1, T4, T8, and T12 denote plants that were subjected to 38 °C for 1, 4, 8, and 12 h, respectively. The expression levels were normalized to the expression level of Actin. (b) Analysis of coding potential for lnc000283 and lnc012465. The coding potential scores were calculated using the CPC program. UBC10 (At5g53300) and ACT2 (At3g18780) are positive controls that encode proteins. COLDAIR (HG975388) and Xist (L04961) serve as negative controls, exhibiting minimal protein-coding potential.
Generation of lncRNA overexpression lines
-
To elucidate the role of lnc000283 and lnc012465 in response to abiotic stress, overexpression vectors were constructed for these lncRNAs, driven by the CaMV 35S promoter, and they were introduced into Arabidopsis thaliana (Col-0 ecotype). Through PCR identification and generational antibiotic screening, two homozygous positive lines for lnc012465 and lnc000283 were obtained. The relative expression levels of these lncRNAs were assessed using qRT-PCR (Fig. 2a). When plants were grown in 1/2 MS medium, with the consumption of nutrients, and reduction of water, the leaves of WT began to turn yellow, but the lnc000283 and lnc012465 overexpression lines developed a deep purple color of leaf veins (Fig. 2b). Examination of chlorophyll and anthocyanin contents in the plants revealed that both overexpression lines had higher levels of chlorophyll and anthocyanin compared to the WT, suggesting that the transgenic plants might possess enhanced resistance to nutritional or water stress (Fig. 2c, d).
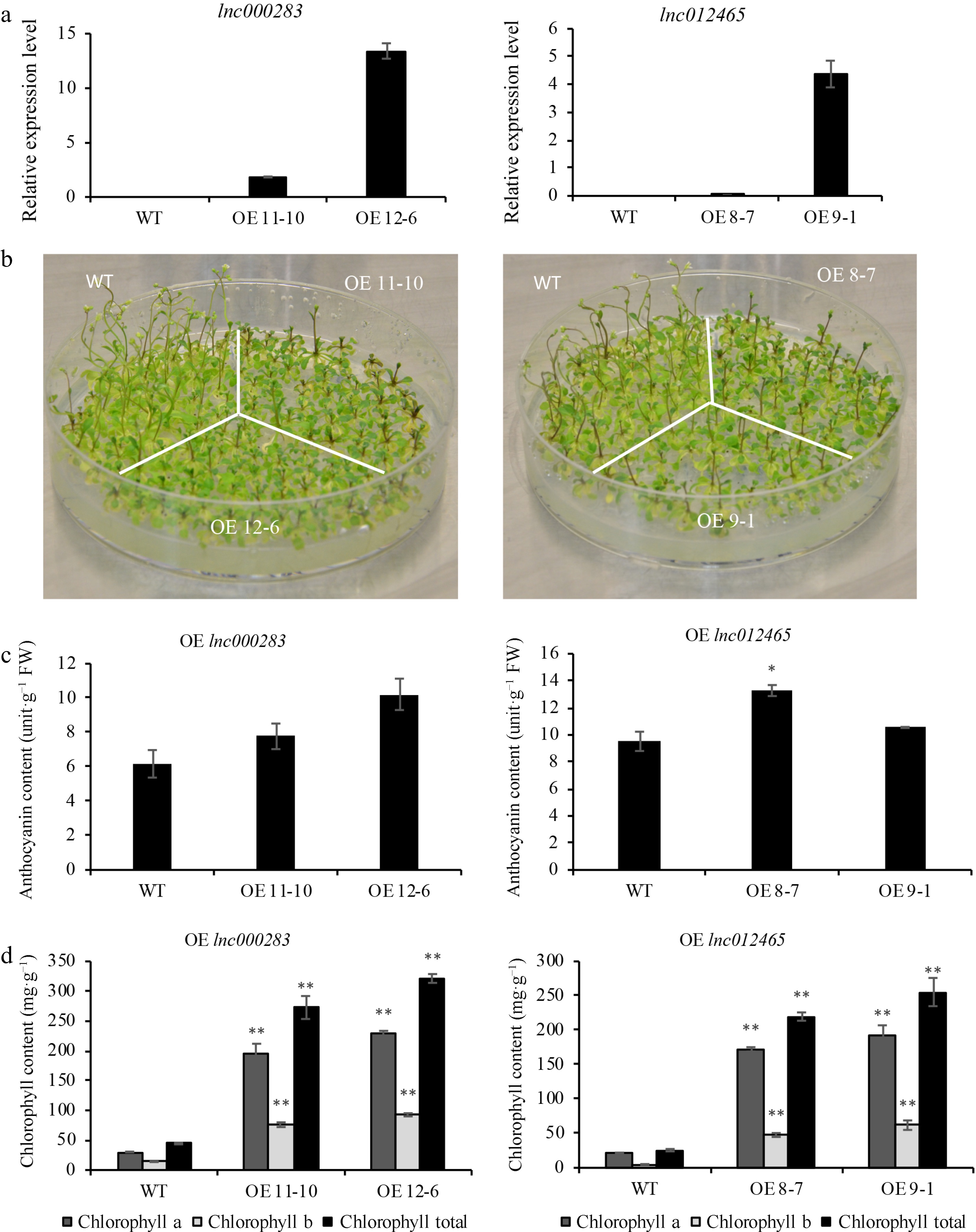
Figure 2.
Arabidopsis plants overexpressing lnc000283 and lnc012465 had higher anthocyanins and chlorophyll content. (a) The relative expression level of lnc000283 and lnc012465 in WT and different transgenic lines. UBC10 (At5g53300) was used as an internal control. Each value is mean ± sd (n = 3). (b) The phenotype of WT and Arabidopsis overexpressing lnc000283 or lnc012465 grown on 1/2 MS medium 50 d after sowing. The (c) anthocyanin and (d) chlorophyll content of WT and transgenic Arabidopsis overexpressing lnc000283 or lnc012465. The asterisks above the bars indicate statistical significance using Student's t-test (*, p < 0.05; **, p < 0.01).
Effect of lncRNA overexpression on heat tolerance in Arabidopsis
-
Given that lnc000283 and lnc012465 were highly induced by heat, the thermotolerance of the overexpressing (OE) plants were compared to that of the wild type. Arabidopsis plants were initially exposed to a an HS treatment at 38 °C for 4 d, followed by recovery at room temperature. The death caused by HS was processive. Post-severe HS challenge for 4 d, OE plants initially appeared similar to WT, but upon recovery, their leaves started to fold or curl, followed by a transition to yellow, white, and eventually drying out (Fig. 3a). OE lnc000283 and OE lnc012465 plants exhibited enhanced thermotolerance compared to WT, with lnc012465 showing particularly strong tolerance (Fig. 3a; Supplemental Fig. S2a). After 5 d of recovery, leaf coloration indicated that transgenic plants maintained a significantly higher percentage of green leaves and a lower percentage of bleached leaves compared to WT (Fig. 3b; Supplemental Fig. S2b). Under non-heat-stress conditions, WT and OE plants possessed comparable water content. However, following heat stress, the fresh-to-dry weight ratio of OE lnc000283 and lnc012465 lines was significantly greater than that of WT (Fig. 3c; Supplemental Fig. S2c). Abiotic stresses frequently trigger the production of excessive reactive oxygen species (ROS), which are believed to cause lipid peroxidation of membrane lipids, leading to damage to macromolecules. Leaf MDA content is commonly used as an indicator of lipid peroxidation under stress conditions; therefore, the MDA content in both transgenic and WT plants was assessed. Figure 3d shows that the MDA content in WT plants progressively increased after heat treatment, whereas in the two lines overexpressing lnc012465, the MDA content increased only slightly and remained significantly lower than that in WT at all time points. In plants overexpressing lnc000283, the MDA content did not significantly differ from that of WT before heat stress. However, after 4 d of heat treatment, the MDA content was significantly lower compared to WT (Supplemental Fig. S2d). The results suggested that the expression of both lnc012465 and lnc000283 can mitigate injury caused by membrane lipid peroxidation under heat-stress conditions. Peroxidase (POD) is a crucial antioxidant enzyme involved in ROS scavenging. Figure 3e and Supplemental Fig. S2e demonstrate that POD activity increased in both transgenic and WT plants after heat treatment. However, the increase in WT plants was modest, whereas OE lnc000283 and OE lnc012465 plants exhibited consistently higher POD activity. As anticipated, proline levels were induced in response to stress in all studied plants (Fig. 3f; Supplemental Fig. S2f). However, under normal conditions and 2 d post-heat stress treatment, the proline content in OE lnc000283 and OE lnc012465 plants did not exhibit significant changes compared to WT (Fig. 3f; Supplemental Fig. S2f). Moreover, after 4 d of heat stress, the proline content in OE lnc012465 lines was significantly lower than in WT, and the OE lnc000283 transgenic line 12-6 also showed a marked decrease in proline content compared to WT (Fig. 3f; Supplemental Fig. S2f). The results indicated that the thermotolerance of plants overexpressing either lnc000283 or lnc012465 was independent of proline accumulation.
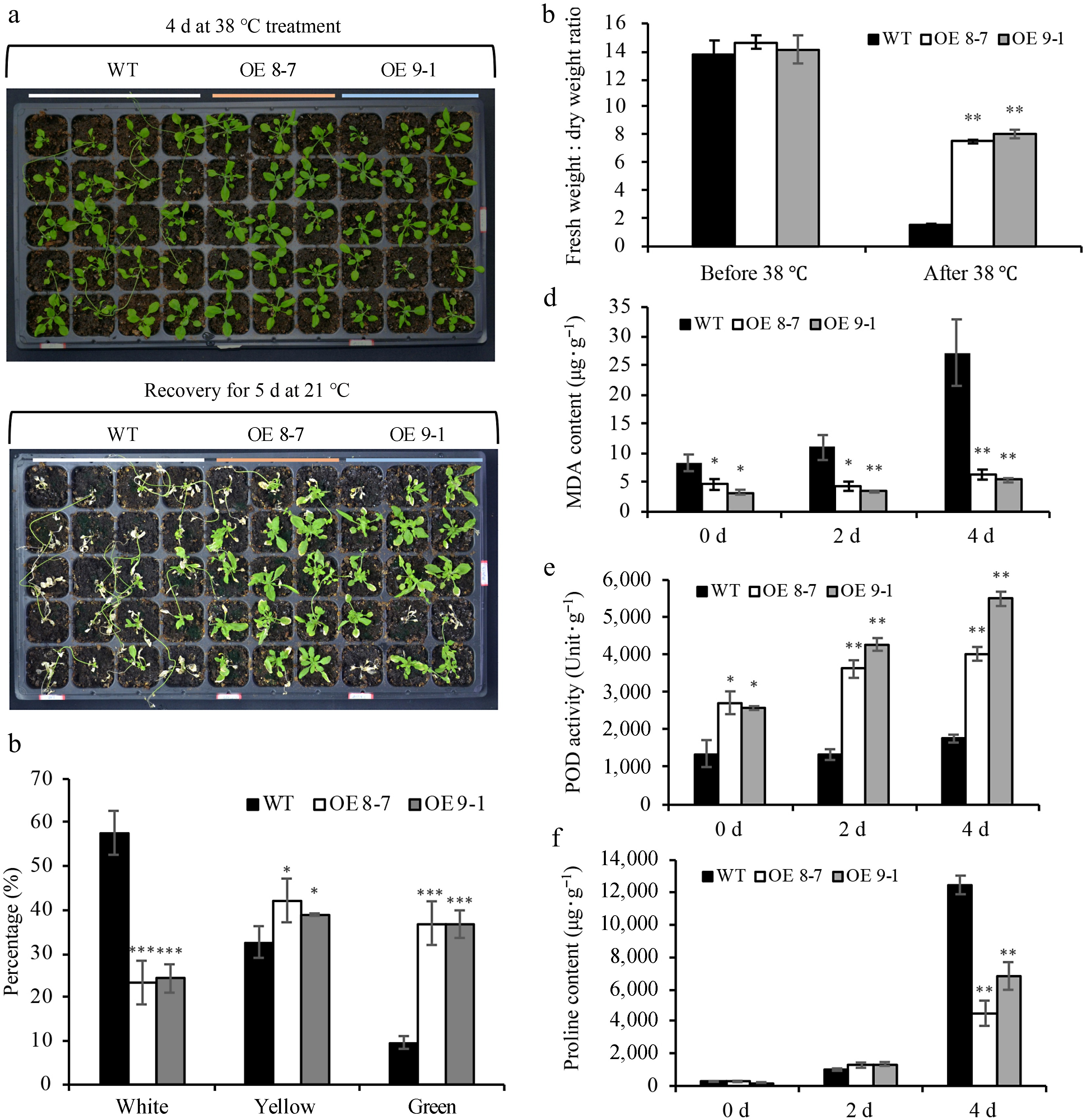
Figure 3.
Overexpressing lnc012465 lines are more tolerant to heat stress. (a) Phenotypes of WT and OE lnc012465 plants were assessed before and after exposure to heat stress. The heat treatment was applied to 25-day-old Arabidopsis plants. (b) The percentage of leaves with different colors in Arabidopsis after heat treatment and recovery for 5 d. (c) The fresh-to-dry weight ratio of Arabidopsis leaves was measured before and after 38 °C heat treatment. (d)−(f) depict the MDA content, POD activity, and proline content in Arabidopsis leaves at varying durations of heat stress. The asterisks above the bars indicate statistical significance using Student's t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
Effect of lncRNA overexpression on the expression quantity of genes related to high-temperature response in Arabidopsis
-
To elucidate the molecular mechanisms by which lncRNAs enhance thermotolerance in Arabidopsis, the expression of the Hsf gene HsfA7a and three Hsps (Hsp25.3, Hsa32, and Hsp18.1-CI) in OE lnc000283, OE lnc012465, and WT Arabidopsis plants were investigated at various time points following heat treatment. As shown in Fig. 4 and Supplemental Fig. S3, both Hsf and Hsps exhibited a rapid response to heat stress with strong induction. Notably, the transcripts of HsfA7a and Hsp25.3 were significantly upregulated at 1 h after heat exposure, then experienced a sharp decrease. Hsa32 and Hsp18.1-CI were highly induced at 1 h and, unlike the other proteins, sustained high expression levels at 3 h (Fig. 4; Supplemental Fig. S3). At 1 h post-heat treatment, the transcript levels of Hsa32 and HsfA7a in OE lnc000283 did not significantly differ from those in WT. However, by 3 h, Hsa32 expression was roughly 50% of the WT level, while HsfA7a expression was approximately double that of WT (Supplemental Fig. S3). The overexpression of lnc000283 did not significantly affect the transcript level of Hsp25.3 at any of the tested time points. Notably, Hsp18.1-CI expression in both lines overexpressing lnc000283 was significantly induced at all three detection points post-heat treatment, reaching approximately 4-9-fold higher levels than in the WT (Supplemental Fig. S3). In Arabidopsis plants with elevated expression of lnc012465, the expression patterns of all Hsp and Hsf genes were similar to those in plants overexpressing lnc000283, with the notable exception of Hsa32. Unlike the WT, Hsa32 did not show a trend of down-regulation at 3 h post-heat treatment (Fig. 4). The findings suggest that the substantial induction of Hsp18.1-CI may play a role in enhancing the thermotolerance of Arabidopsis plants overexpressing lnc000283 and lnc012465.
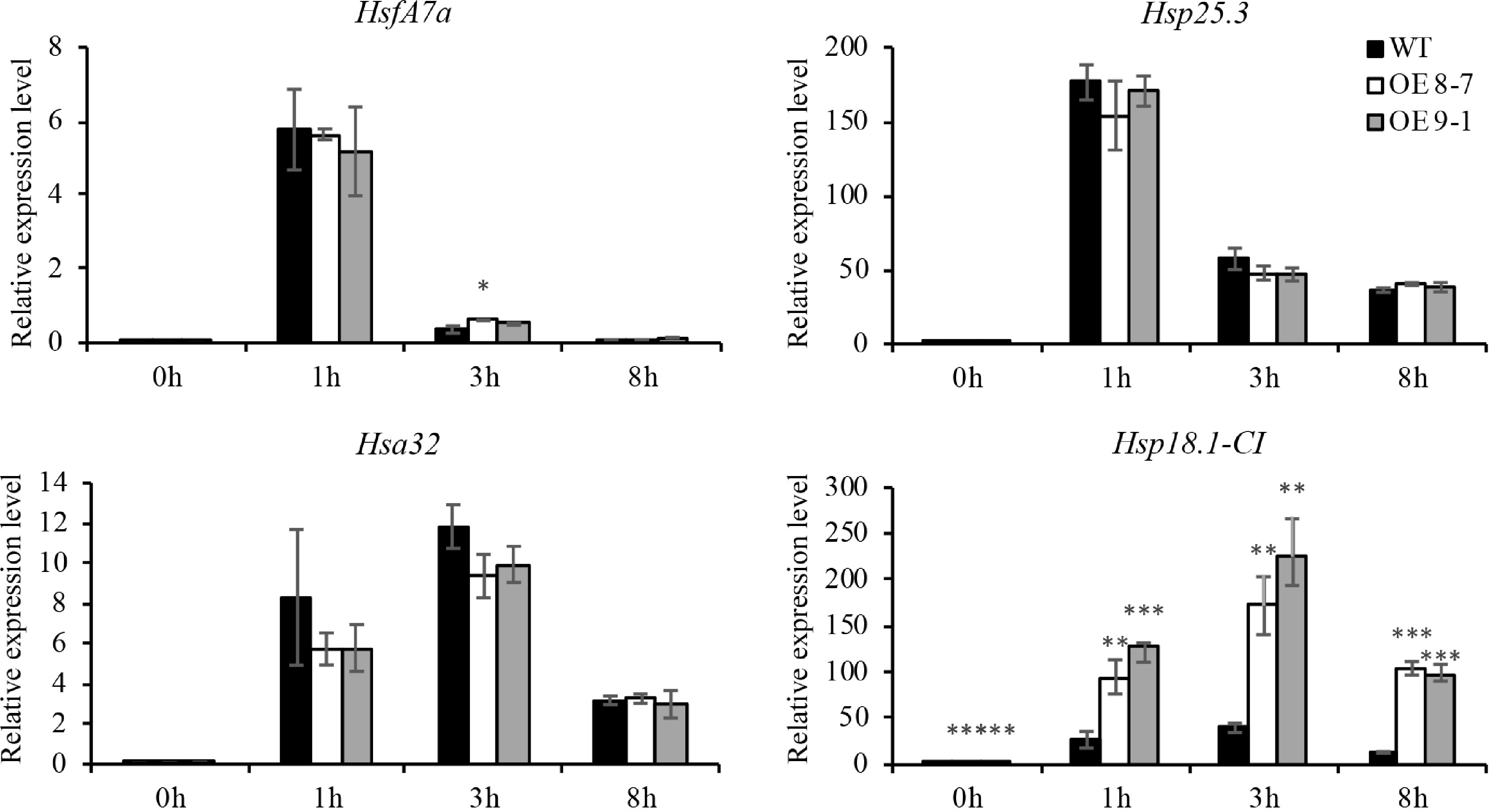
Figure 4.
The expression of HSF and HSP genes in lnc012465 overexpressing lines before and after different heat treatment times. Gene expression levels were quantified using RT-qPCR and normalized to UBC10 (At5g53300). Each value represents the mean ± standard deviation (n = 3). The asterisks above the bars indicate statistical significance using Student's t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
Effect of lncRNA overexpression on drought tolerance in Arabidopsis
-
Prior research has implicated a significant proportion of genes in conferring resistance to various abiotic stresses. To elucidate the functions of lnc000283 and lnc012465 more thoroughly, WT and transgenic plants were subjected to drought stress by depriving them of water for 9 d. It was noted that the majority of leaves in WT plants withered and dried, whereas the OE lnc000283 and OE lnc012465 plants exhibited reduced withering, with only a minority displaying dryness (Fig. 5a; Supplemental Fig. S4a). Eight days post-rewatering, a negligible fraction of WT seedlings exhibited recovery, whereas the overwhelming majority of transgenic plants regained vigorous growth (Fig. 5a; Supplemental Fig. S4a). The transgenic plants demonstrated a significantly higher survival rate compared to the WT plants. Following 9 d of water deficit treatment, less than 40% of the WT plants survived, whereas the OE 012465 lines 8-7 and 9-1 exhibited survival rates of 100% and 95%, respectively, and the OE 000283 lines 11-10 and 12-6 had survival rates of 87% each. (Fig. 5b; Supplemental Fig. S4b). Water loss serves as a critical metric for assessing plant drought tolerance, hence the fresh-to-dry weight ratio of excised leaves was assessed via desiccation analysis. Following 4 d of drought treatment, the fresh-to-dry weight ratio for WT plants was reduced to 43%, whereas for OE lnc000283 lines 11-10 and 12-6, it was reduced to 73% and 75%, respectively. For OE 012465 lines 8-7 and 9-1, the ratios were reduced to 67% and 62%, respectively (Fig. 5c; Supplemental Fig. S4c). The findings indicated that lnc000283 and lnc012465 endow the transgenic plants with drought tolerance.
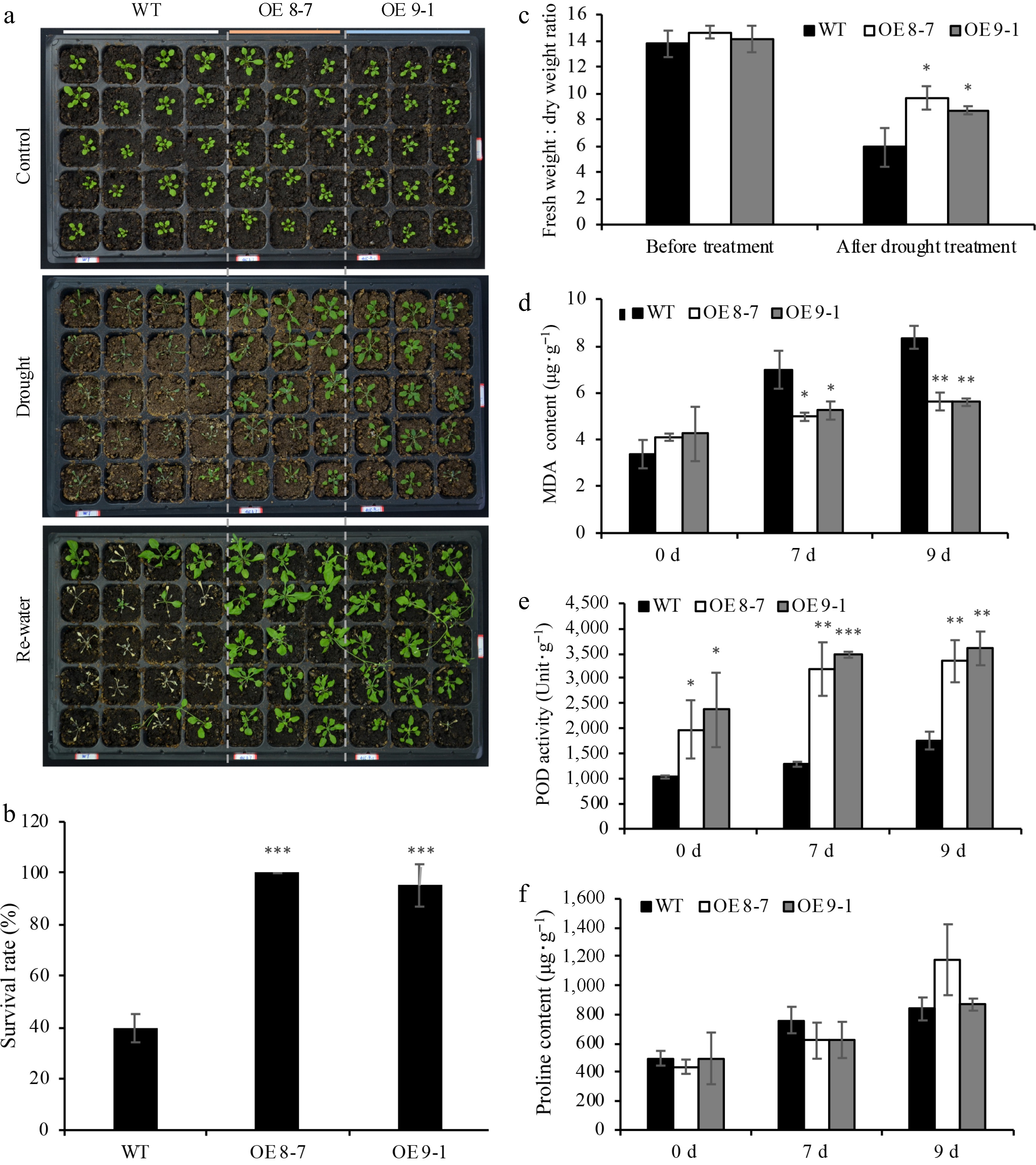
Figure 5.
Overexpressing lnc012465 lines are more tolerant to drought stress. (a) Phenotype of WT and OE lnc012465 plants before and after subjecting to drought stress. Drought treatment was carried out on 20-day-old Arabidopsis plants. (b) The percentage of leaves with different colors in Arabidopsis after heat treatment and recovery for 5 d. (c) The fresh weight to dry weight ratio of Arabidopsis leaves before and after undergoing 38 °C heat treatment. (d)−(f) MDA content, POD activity, and proline content in Arabidopsis leaves under different times of heat stress. The asterisks above the bars indicate statistical significance using Student's t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001)
MDA content in leaves is a standard biomarker for assessing the extent of drought stress-induced damage. Prior to drought stress exposure, MDA levels in WT and transgenic plants were comparable. However, following 7 and 9 d of water deficit, the MDA content in the transgenic plants was markedly reduced compared to the WT, suggesting a less severe degree of membrane lipid peroxidation in the transgenic plants (Fig. 5d; Supplemental Fig. S4d). Oxidative stress frequently coincides with drought stress, hence the activity of POD was assessed to evaluate the ROS scavenging ability. The findings indicated that as the duration of drought treatment increased, POD activity progressively rose. Before drought exposure, the POD activity in lines 11-10 and 12-6 of OE 000283 was 2.4-fold and 2.2-fold higher than that of the WT, respectively (refer to Supplemental Fig. S4e). Following drought treatment, the POD activity in the transgenic lines remained significantly elevated compared to the wild type, although the enhancement was less pronounced than before the treatment (Supplemental Fig. S4e). In the OE 012465 plants, the POD activity in lines 8-7 and 9-1 significantly surpassed that of the wild type, with the discrepancy being more pronounced during drought stress (Fig. 5e). The proline content in WT and OE 000283 plants exhibited no significant differences before and after 7 d of treatment. However, after 9 d of drought, the proline content in OE 000283 plants was significantly lower compared to that in the WT (Supplemental Fig. S4f). OE 000465 plants showed no significant difference from the wild type before and after drought treatment (Fig. 5f). The findings were consistent with those under heat stress, indicating that the enhanced stress resistance due to the overexpression of lnc000283 and lnc012465 in Arabidopsis is not reliant on proline accumulation.
Following drought stress treatment, the expression levels of drought-related genes such as RD29A, RD29B, NCED3, AREB1, and Rab18 were significantly elevated in plants overexpressing lnc000283 and lnc012465 compared to WT plants. These findings suggest that lnc000283 and lnc012465 modulate Arabidopsis drought tolerance by regulating the expression of genes associated with the drought stress response (Fig. 6; Supplemental Fig. S5).
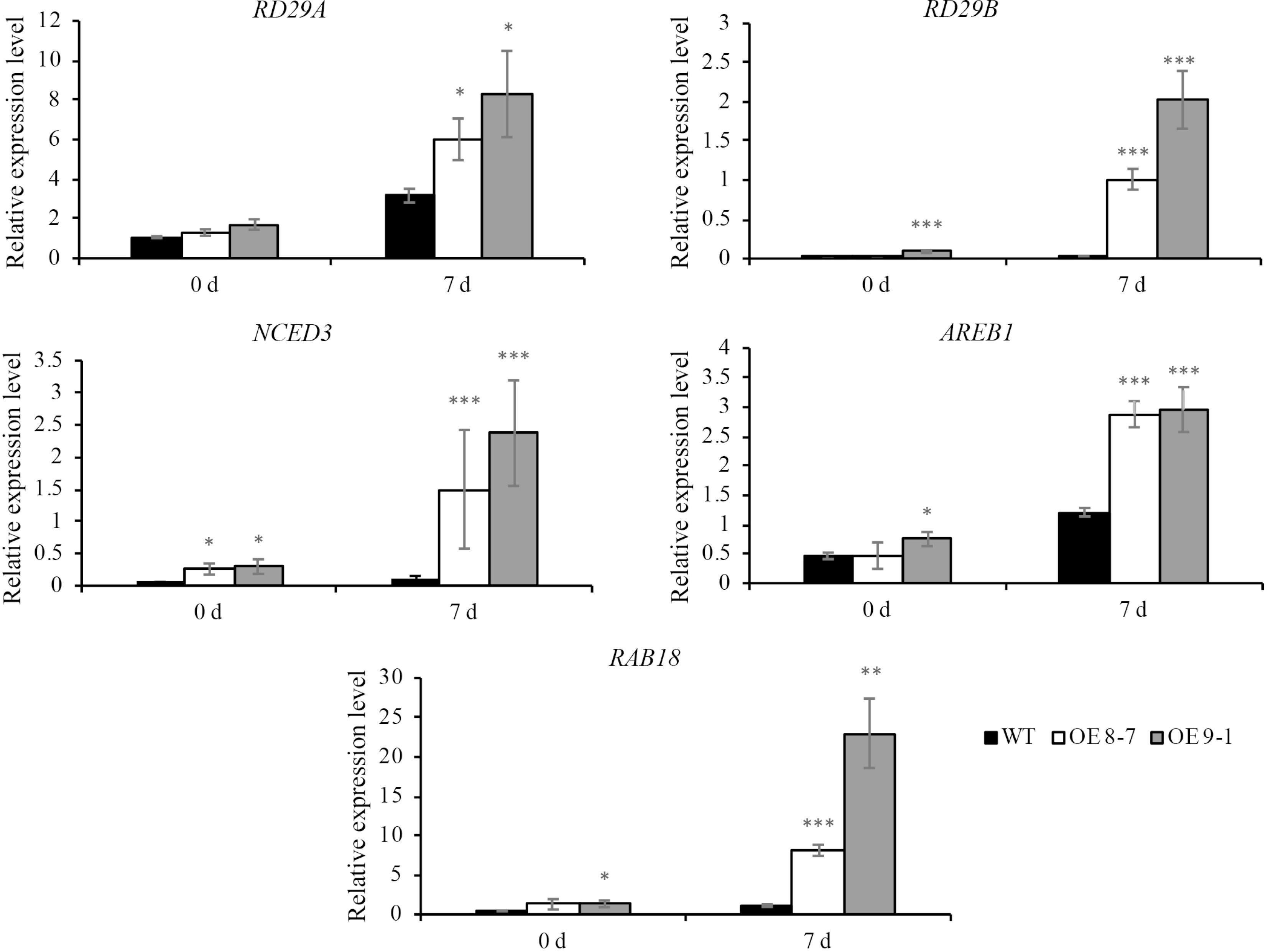
Figure 6.
The expression of drought-responsive genes in lnc012465 overexpressing lines before and after different drought treatment time. Gene expression levels were determined by qRT-PCR normalized against UBC10 (At5g53300). Each value is mean ± sd (n = 3). The asterisks above the bars indicate statistical significance using Student's t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
-
The integrity of global food security is under threat due to the confluence of rapid population expansion and profound climatic shifts[29]. Amidst the shifting climatic landscape, heat and drought stress have emerged as primary limitations to crop yield and global food security. Understanding how plants detect stress cues and acclimate to challenging conditions is a pivotal biological inquiry. Moreover, enhancing plant resilience to stress is essential for maintaining agricultural productivity and fostering environmental sustainability[2]. Concurrently, the advancement of next-generation sequencing (NGS) technology has led to the identification of a substantial number of lncRNAs that participate in diverse stress responses, with functional analyses having been conducted on several of these molecules.[30] For instance, in the case of potatoes, the lncRNA StFLORE has been identified to modulate water loss through its interaction with the homologous gene StCDF1[31]. LncRNA TCONS_00021861 can activate the IAA biosynthetic pathway, thereby endowing rice with resistance to drought stress[32]. In wheat, the expression of TalnRNA27 and TalnRNA5 was upregulated in response to heat stress[33]. Our prior investigation identified a total of 81 lncRNAs in Chinese cabbage that engage in intricate interactions with their respective mRNA targets across various phases of heat treatment[25]. Two lncRNAs, lnc000283 and lnc012465, were chosen for subsequent functional analysis. Findings confirmed that these lncRNAs endow transgenic Arabidopsis plants with enhanced tolerance to both heat and drought, thereby offering novel resources for enhancing stress resistance through genetic engineering.
Abiotic stresses frequently trigger the synthesis of anthocyanins, serving as natural antioxidants that mitigate oxidative damage by neutralizing surplus reactive oxygen species (ROS), thereby protecting plants from growth inhibition and cell death, allowing plants to adapt to abiotic stresses[34,35]. For instance, during chilling stress, the accumulation of anthocyanins within leaves can mitigate oxidative damage, thereby enhancing the photosynthetic rate[36]. Consequently, the level of abiotic stress tolerance can be inferred from the concentration of anthocyanins. The reduction of photosynthetic ability is one of the key physiological phenomena of stresses, which is partly due to the degradation of chlorophyll caused by leaf senescence during stress. The reduced accumulation of chlorophyll in the plants was seen in many plants when exposed to drought or heat stress conditions. The current investigation revealed that lncRNA-overexpressing plants cultivated in Petri dishes exhibited increased accumulation of both chlorophyll and anthocyanins in advanced growth phases, indicating that these transgenic plants, overexpressing lnc000283 and lnc012465, demonstrated enhanced stress tolerance and superior growth performance relative to WT (Fig. 2c, d).
Upon exposure to heat stress, there is a marked induction of transcription for numerous genes that encode molecular chaperones in plants, with the vast majority of these genes contributing to the prevention of protein denaturation-related damage and the augmentation of thermotolerance[37−39]. The present investigation identified multiple heat-inducible genes in plants overexpressing lnc000283 and lnc012465, as well as in WT (Fig. 4; Supplemental Fig. S3). The findings indicated that of the four HSP or HSF genes examined, Hsp18.1-CI exhibited a significantly greater abundance in both OE lnc000283 and OE lnc012465 plants compared to the WT following heat treatment for several days. Hsp18.1-CI, formerly referred to as Hsp18.2 has been the subject of investigation since 1989.[40] Following the fusion of the 5' region of Hsp18.2 in frame with the uidA gene of Escherichia coli, the activity of GUS, serving as the driver gene was observed to increase upon exposure to HS[40]. The Arabidopsis hsfA2 mutant exhibited diminished thermotolerance after heat acclimation, with the transcript levels of Hsp18.1-CI being substantially reduced compared to those in wild-type plants following a 4-h recovery period[41]. The findings revealed that the upregulation of Hsp18.1-CI protein is a critical mechanism by which plants achieve enhanced protection against heat stress in adverse environmental conditions, thereby bolstering their thermotolerance.
Plants cultivated in natural settings are often subjected to concurrent multiple abiotic stresses, which can exacerbate threats to their routine physiological functions, growth, and developmental processes[42,43]. Elucidating the molecular mechanisms underlying plant responses to abiotic stress is crucial for the development of new crop varieties with enhanced tolerance to multiple abiotic stresses. Previous research has indicated that the overexpression of certain protein-coding genes can endow plants with resistance to a variety of abiotic stresses. For instance, tomatoes with robust expression of ShCML44 demonstrated significantly enhanced tolerance to drought, cold, and salinity stresses[44]. Overexpression of PeCBF4a in poplar plants confers enhanced tolerance to a range of abiotic stresses[45]. With respect to lncRNAs, transgenic Arabidopsis plants that overexpress lncRNA-DRIR displayed marked increased tolerance to salt and drought stresses compared to the wild-type[46]. In the present study, both overexpression lines of lnc000283 and lnc012465 exhibited resistance to heat and drought stresses, thereby contributing to the enhancement of plant resilience against multiple stresses (Figs 3, 5; Supplemental Figs S2, S4).
The number of genes implicated in plant drought resistance is regulated by both ABA-dependent and ABA-independent pathways[47,48]. It is well established that the expression of RD29A exhibits a high level of responsiveness to drought stress, operating through both ABA-dependent and ABA-independent mechanisms[49]. RD29B, AREB1, and RAB18 are governed by an ABA-dependent regulatory pathway[10,49,50]. NCED3 is involved in ABA biosynthesis[51]. In the present study, the transcript levels of RD29A, RD29B, NCED3, AREB1, and RAB18 were significantly elevated in OE lnc000283 and OE lnc012465 plants compared to those in the WT plants (Fig. 6; Supplemental Fig. S5). The findings indicated that the drought tolerance imparted by OE lnc000283 and OE lnc012465 plants is contingent upon an ABA-dependent mechanism.
Prior research has indicated that certain long non-coding RNAs (lncRNAs) can assume analogous roles across diverse biological contexts. For example, the lncRNA bra-miR156HG has been shown to modulate leaf morphology and flowering time in both B. campestris and Arabidopsis[52]. Heterogeneous expression of MSL-lncRNAs in Arabidopsis has been associated with the promotion of maleness, and similarly, it is implicated in the sexual lability observed in female poplars[53]. In the present study, lnc000283 and lnc012465 were induced by heat in Chinese cabbage, and their heterologous expression was found to confer heat tolerance in Arabidopsis. Additionally, sequences homologous to lnc000283 and lnc012465 were identified in Arabidopsis (Supplemental Fig. S1). The data suggest that these sequences may share a comparable function to that of heat-inducible sequences, potentially accounting for the conservation of lnc000283 and lnc012465'os functionality across various species.
-
In conclusion, the functions of two heat-inducible lncRNAs, lnc000283 and lnc012465 have been elucidated. Transgenic Arabidopsis lines overexpressing these lncRNAs accumulated higher levels of anthocyanins and chlorophyll at a later stage of growth compared to the WT when grown on Petri dishes. Furthermore, under heat and drought stress conditions, these OE plants exhibited enhanced stress tolerance, with several genes related to the stress resistance pathway being significantly upregulated. Collectively, these findings offer novel insights for the development of new varieties with tolerance to multiple stresses.
-
The authors confirm contribution to the paper as follows: study conception and supervision: Li N, Song X; experiment performing: Wang Y, Sun S; manuscript preparation and revision: Wang Y, Feng X, Li N. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
This work was supported by the National Natural Science Foundation of China (32172583), the Natural Science Foundation of Hebei (C2021209019), the Natural Science Foundation for Distinguished Young Scholars of Hebei (C2022209010), and the Basic Research Program of Tangshan (22130231H).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 The primers sequences used in this study for qRT-PCR.
- Supplemental Table S2 Subcellular localization of lnc000283 and lnc012465 predicted by database lnclocator.
- Supplemental Fig. S1 Sequence alignment of lnc000283 and lnc012465 in Chinese cabbage with their homologous lncRNAs in Arabidopsis, respectively. (a) Sequence alignment of lnc000283 and its homologous lncRNA CNT2088434. (b) Sequence alignment of lnc012465 and its homologous lncRNA CNT2088742.
- Supplemental Fig. S2 Overexpressing lnc000283 lines are more tolerant to heat stress. (a) Phenotype of WT and OE lnc000283 plants before and after subjecting to heat stress. Heat treatment was carried out on 25-day-old Arabidopsis plants. (b) The percentage of different colors of leaves after subjecting to heat treatment and recovered for 5 days. (c) The fresh weight versus dry weight ratio of Arabidopsis leaves before and after undergoing 38 ℃ heat treatment. (d)-(f) MDA content, POD activity, and proline content in Arabidopsis leaves under different times of heat stress. The asterisks above the bars indicate statistical significance using Student’s t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
- Supplemental Fig. S3 The expression of HSF and HSP genes in lnc000283 overexpressing lines before and after different heat treatment times. Gene expression levels were determined by RT-qPCR normalized against UBC10 (At5g53300). Each value is mean ± sd (n = 3). The asterisks above the bars indicate statistical significance using Student’s t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
- Supplemental Fig. S4 Overexpressing lnc000283 lines are more tolerant to drought stress. (a) Phenotype of WT and OE lnc000283 plants before and after subjecting to heat stress. Drought treatment was carried out on 20-day-old Arabidopsis plants. (b) The percentage of leaves with different colors recovered 5 days after heat treatment in Arabidopsis. (c) The fresh weight to dry weight ratio of Arabidopsis leaves before and after undergoing 38 ℃ heat treatment. (d)-(f) MDA content, POD activity, and proline content in Arabidopsis leaves under different times of heat stress. The asterisks above the bars indicate statistical significance using Student’s t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
- Supplemental Fig. S5 The expression of drought-responsive genes in lnc000283 overexpressing lines before and after different drought treatment times. Gene expression levels were determined by RT-qPCR normalized against UBC10 (At5g53300). Each value is mean ± sd (n = 3). The asterisks above the bars indicate statistical significance using Student’s t-test (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Sun S, Feng X, Li N, Song X. 2024. Two lncRNAs of Chinese cabbage confer Arabidopsis with heat and drought tolerance. Vegetable Research 4: e029 doi: 10.48130/vegres-0024-0029 

Two lncRNAs of Chinese cabbage confer Arabidopsis with heat and drought tolerance
- Received: 26 April 2024
- Revised: 23 June 2024
- Accepted: 22 July 2024
- Published online: 19 September 2024
Abstract: Currently, frequent occurrences of abnormal weather events, including drought and extreme temperatures, are impacting plant growth and crop yields globally. Numerous lncRNAs in plants are known to be associated with stress responses, yet only a select few have been validated experimentally. Our prior research identified a set of lncRNAs in Chinese cabbage that were closely related to heat stress response through RNA-seq and bioinformatics analysis. In this study, two lncRNAs (lnc000283 and lnc012465) were selected for genetic transformation in Arabidopsis to elucidate their functions. Arabidopsis plants over-expressing (OE) lnc000283 and lnc012465 accumulated higher levels of anthocyanins in the later growth stage when cultivated in a culture dish. When subjected to heat stress, both OE lnc000283 and OE lnc012465 plants exhibited enhanced thermotolerance, with lnc012465 showing particularly pronounced effects. Moreover, the transgenic Arabidopsis plants demonstrated greater resistance to drought stress compared to the wild type. Under heat or drought treatments, the transgenic plants exhibited significantly higher fresh-to-dry weight ratios, POD activities, green leaf percentages, and survival rates compared to the wild type. Additionally, the transcript levels of Hsp18.1-CI were substantially elevated in the transgenic plants, both before and after heat treatment, and the expression levels of several drought-responsive genes were significantly higher than those in the wild type following drought stress. Altogether, the present results provide new ideas for the breeding of new multi-stress-tolerant varieties.
-
Key words:
- Chinese cabbage /
- Arabidopsis /
- lncRNA /
- Thermotolerance /
- Drought tolerance /
- Physiological analysis /
- Gene expression