









-
Transcription factors (TFs), also known as sequence-specific DNA-binding factors play pivotal roles in mediating gene expression regulation[1]. TFs generally act as key regulators of gene expression. In general, the TFs have two distinct domains, a DNA-binding domain and a transcriptional activation/repression domain[2]. With the study and report of plant TFs, an increasing number of TFs have been identified in different species, such as MADS[3], MYB[2], WRKY[4], bHLH[5], and AP2/ERF[6].
AP2/ERF (APETALA2/ethylene responsive factor) represented one of the most expansive families in the field of TFs[7]. Furthermore, in this superfamily, the sequences were characterized by the presence of one or more conserved AP2 domains, typically comprising approximately 60 to 70 amino acids within the DNA-binding region[8]. The AP2/ERF gene family was divided into five subfamilies: AP2, ERF (ethylene-responsive element binding factors), RAV (associated with ABI3/VP1), DREB, and Soloist[9]. The AP2 subfamily genes were characterized by the presence of two adjacent AP2 domains, whereas members of the RAV subfamily were distinguished by consensus sequence elements containing AP2 and B3 domains[10]. Exploration of the AP2/ERF gene family has been extensive across various plant species, e.g., 116 members in pomegranate (Punica granatum L.)[11], 158 members in kiwifruit (Actinidia eriantha)[12], 64 members in strawberry (Fragaria × ananassa Duch)[13], 97 genes in pineapple (Ananas comosus L. Merr)[14], 121 genes in lotus (Nelumbo nucifera)[15], 131 genes in peach (Prunus persica)[16], and 119 AP2/ERF genes in melon (Cucumis melo L.)[17].
The AP2/ERF gene family plays crucial roles in coordinating various aspects of plant growth, development, and associated biological processes[18]. These factors are known to be involved in the regulation of plant growth and development, response to abiotic and biotic stresses, regulation of fruit ripening, defense mechanisms, and metabolic pathways. The AP2/ERF transcription factor MdERF2 inhibits MdACS1 transcription through multiple mechanisms, thereby negatively affecting ethylene biosynthesis and fruit ripening in apples, thus acting as an antagonist of positive ripening regulators[19]. CitERF71 activates the terpene synthase gene CitTPS16, which promotes the synthesis of E-geraniol in sweet orange fruits, thereby increasing their flavor[20]. Peach transcription factor PpeERF2 regulates fruit ripening by inhibiting the expression of two ABA biosynthesis genes and one cell wall degradation gene[21]. Zhang et al.[22] found that miR172-AP2 interaction may play an important role in regulating the ripening process of blackberry fruit. Transcription factor MdAP2-34 promotes the accumulation of apple flavonoids by binding and activating MdF3'H promoter activity[23].
Loquat (Eriobotrya japonica) is a subtropical evergreen fruit tree, belonging to the Rosaceae family, native to southeastern China. Its flowering typically occurs during winter, and the fruit ripens in late spring and early summer[24]. According to historical documents, loquat cultivation has spanned over 2,000 years in China and has extended to more than 10 countries, including Italy, Turkey, Japan, Brazil, Israel, and Pakistan. Notably, China is the largest producer, with continuous growth in planting area and yield[25]. In recent years, many studies have been carried out on loquat. For instance, EjBZR1 inhibited expression and fruit cell enlargement by binding to the EjCYP90 promoter in loquat[26]. At present, a variety of transcription factor families have been identified in loquat. For example, 28 members of the SBP gene family were identified in the loquat genome[27], and 18 members of the WOX gene family[28]. However, the detection and characterization of AP2/ERF transcription factors in loquat remain unexplored. Therefore, it is necessary to investigate the AP2/ERF gene family in loquat to broaden our understanding of its regulatory mechanisms.
In the present research, the AP2/ERF gene family within the loquat genome was systematically identified and a comprehensive genome-wide analysis was conducted. The phylogenetic tree, gene structures, chromosome locations, conserved motif, syntenic, cis-elements, and gene expression patterns of the AP2/ERF genes were analyzed. The findings of this study served as a foundation for advancing our comprehension of the evolutionary trajectories and greatly facilitates further elucidation of biological functions of AP2/ERF genes in loquat.
-
Two cultivars of loquat fruits were obtained from Haining City, Zhejiang Province (China). 'YingShuang' ('YS') is an early maturing loquat cultivar with white flesh, which matures around May 10th; 'ZheHong 16' ('ZH16') is a medium ripe loquat cultivar with red flesh, which matures around May 25th. Fruits were harvested at four developmental stages, representing the expansion stage (105 days after bloom (DAB) for 'ZH16' and 90 DAB for 'YS'), the turning stage (135 DAB for 'ZH16' and 120 DAB for 'YS'), the mature yellow stage (150 DAB for 'ZH16' and 135 DAB for 'YS'), and the ripening stage (165 DAB for 'ZH16' and 150 DAB for 'YS'). Three biological replicates, each with five fruits were harvested, frozen in liquid nitrogen, and stored at −80 °C until further analysis.
Identification of AP2/ERF genes in the loquat genome
-
The genome data of loquat (Eriobotrya japonica) were obtained from the GigaScience Database (
http://gigadb.org/dataset/view/id/100711 ). The amino acid sequences of Arabidopsis AP2/ERFs were obtained from the Arabidopsis Information Resource (TAIR) (www.arabidopsis.org ). The Hidden Markov Model (HMM) profile of the AP2/ERF domain (PF00847) was retrieved through Pfam database (http://pfam-legacy.xfam.org/ ). AP2/ERF genes of loquat were screened using HMMER and BLASTP tools. Combining the NCBI Conserved Domains Database (CDD,www.ncbi.nlm.nih.gov ) to further screen genes with at least one AP2 domain. The molecular weights (MW) and isoelectric points (PI) of loquat AP2/ERF were predicted by ExPASy ProtParam (https://web.expasy.org/protparam/ ). The subcellular location of AP2/ERF was predicted using the WoLFPSORT online platform (https://wolfpsort.hgc.jp/ ).Phylogenetic analysis and chromosomal localization of AP2/ERF genes in loquat
-
Multiple sequence alignment was performed on AP2/ERF of loquat and Arabidopsis thaliana using ClustalX software. The phylogenetic tree was established using the Neighbor-joining (NJ) method in the FigTree v1.4.4 program and visualized through the Evolview (
www.evolgenius.info/evolview ) online website. The chromosomal localization information of AP2/ERF family members was obtained from the loquat genome annotation file. The results of gene chromosomal localization were visualized through the Gene Location Visualize from the GTF/GFF program of TBtools.Gene structure and conserved motif of the AP2/ERF genes
-
The exon-intron structures of the loquat AP2/ERF genes were analyzed using TBtools software. Conserved motif analysis of loquat AP2/ERF was performed using the MEME program (
https://meme-suite.org/ ) with the selected number of motifs set to 10.Syntenic and cis-regulatory elements analysis of AP2/ERF genes
-
The One Step MCScanX program was used to analyze the intraspecific syntenic of AP2/ERF genes. The collinearity relationship of AP2/ERF genes was drawn by Advanced Circos function in TBtools software. A 2,000 bp genomic sequence was extracted of each AP2/ERF gene as the putative promoter region through the GTF/GFF3 Sequences Extract program in TBtools. The cis-acting elements in the AP2/ERF promoter regions were predicted by the PlantCARE database (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) and the AP2/ERF cis-acting elements results were visualized by the Simple BioSequence Viewer program in TBtools.Expression pattern analysis of AP2/ERF genes using RNA-seq
-
The total RNA was isolated from different cultivars of loquat fruits. RNA concentration and quality were determined by NanoDrop One spectrophotometer, and the integrity of RNA was detected by 1% agarose gel electrophoresis. PrimeScript™ RT reagent Kit (Perfect Real Time) was used for the synthesis of first-strand cDNA. Gene expression analysis was performed by constructing a high-throughput Illumina strand-specific RNA-seq library. All clean reads were mapped to the loquat (Eriobotrya japonica) genome. The data was calculated by reads per kilobase per million mapped read (RPKM) values as transcript abundance. The expression patterns of loquat AP2/ERF genes were visualized using the Heatmap illustrator tool within TBtools.
-
A total of 220 AP2/ERF genes were initially identified in loquat by the HMMER procedure. Subsequently, structural domain analysis was performed on all candidate genes, and genes with incomplete structural domains and duplicated sequences were excluded. Ultimately, 189 AP2/ERF family members were retained for further analysis. Biochemical and physiological characterization of these AP2/ERFs revealed that their protein lengths ranged from 138 amino acids (aa) (EVM0041952.1) to 829 aa (EVM0045558.1), with an average of 314 aa (Supplemental Table S1). The protein molecular weights (MW) ranged from 15,309.02 Da (EVM0041952.1) to 91,294.57 Da (EVM0045558.1), with an average of 34,813.01 Da. The predicted isoelectric point (PI) of the AP2/ERFs ranged from 4.47 to 10.12, with EVM0008391.1 having the lowest isoelectric point of 4.47 whereas EVM0031649.1 had the highest isoelectric point of 10.12. Prediction of subcellular location showed that 154 proteins were localized in the nucleus, 25 proteins were localized in the chloroplast, and five proteins were localized in the cytoplasm. These results provided a theoretical basis for further studies on the function of AP2/ERF genes.
Phylogenetic analysis and classification of AP2/ERF genes in loquat
-
To comprehensively determine the evolutionary relationships and functions of AP2/ERF genes in loquat, multiple sequence alignment was performed on 325 AP2/ERF protein sequences in loquat and Arabidopsis thaliana, and a phylogenetic tree was constructed using the Neighbor-joining (NJ) method. Furthermore, loquat AP2/ERF genes were classified based on the classification method of Arabidopsis AP2/ERF genes. The results showed that all of the loquat AP2/ERF genes were divided into three subfamilies, named AP2, ERF, and DREB (Fig. 1). The ERF subfamilies contained 96 genes, which was the largest among all subfamilies. The ERF subfamily was divided into B1, B2, B3, B4, B5, and B6 groups, with 15, 6, 31, 10, 8, and 26 genes, respectively. The DREB subfamily contained 51 genes, and was classified into A1, A2, A3, A4, A5, and A6, with 4, 9, 2, 17, 12, and 7 genes, respectively. The AP2 subfamily comprised 42 genes, with the fewest number of genes among all subfamilies.
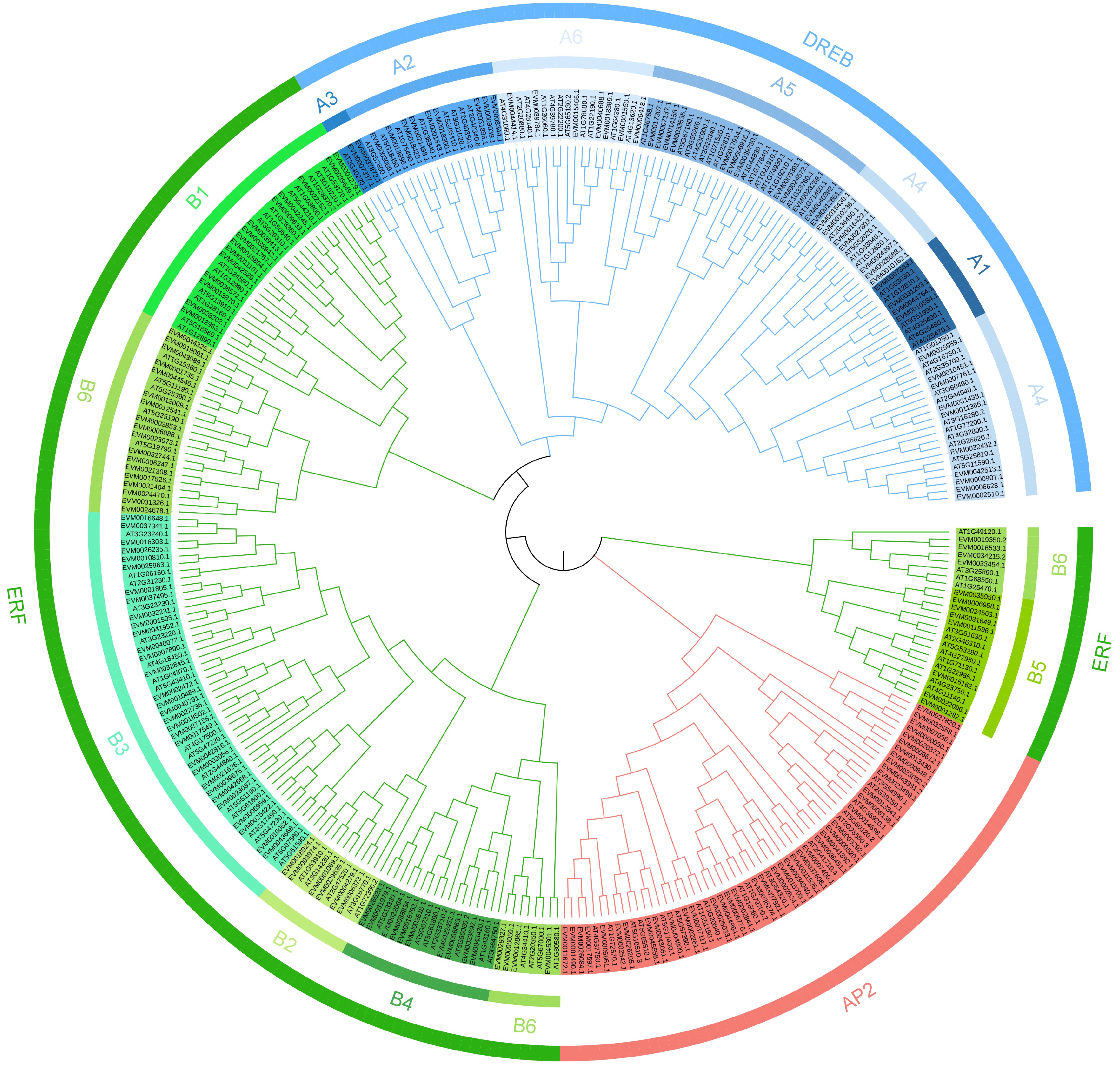
Figure 1.
Phylogenetic tree of the 355 AP2/ERF genes from loquat (189) and Arabidopsis thaliana (136). Different color blocks represent different subfamily members.
Chromosomal location analysis of AP2/ERF genes in loquat
-
Except for the three genes EVM0013696.1, EVM0022152.1, and EVM00113430.1 located on the skeleton, most genes were unevenly distributed on the loquat chromosome (Fig. 2). Specifically, 19 AP2/ERF genes (10.05%) were located on Chr 10, 18 on Chr 3 (9.52%), and 17 on Chr 6 (9.00%). However, the number of genes on Chr 4 was the smallest, with only seven AP2/ERF genes (3.70%). Consequently, the findings suggested that there was no significant correlation between chromosome length and the distribution of AP2/ERF genes.
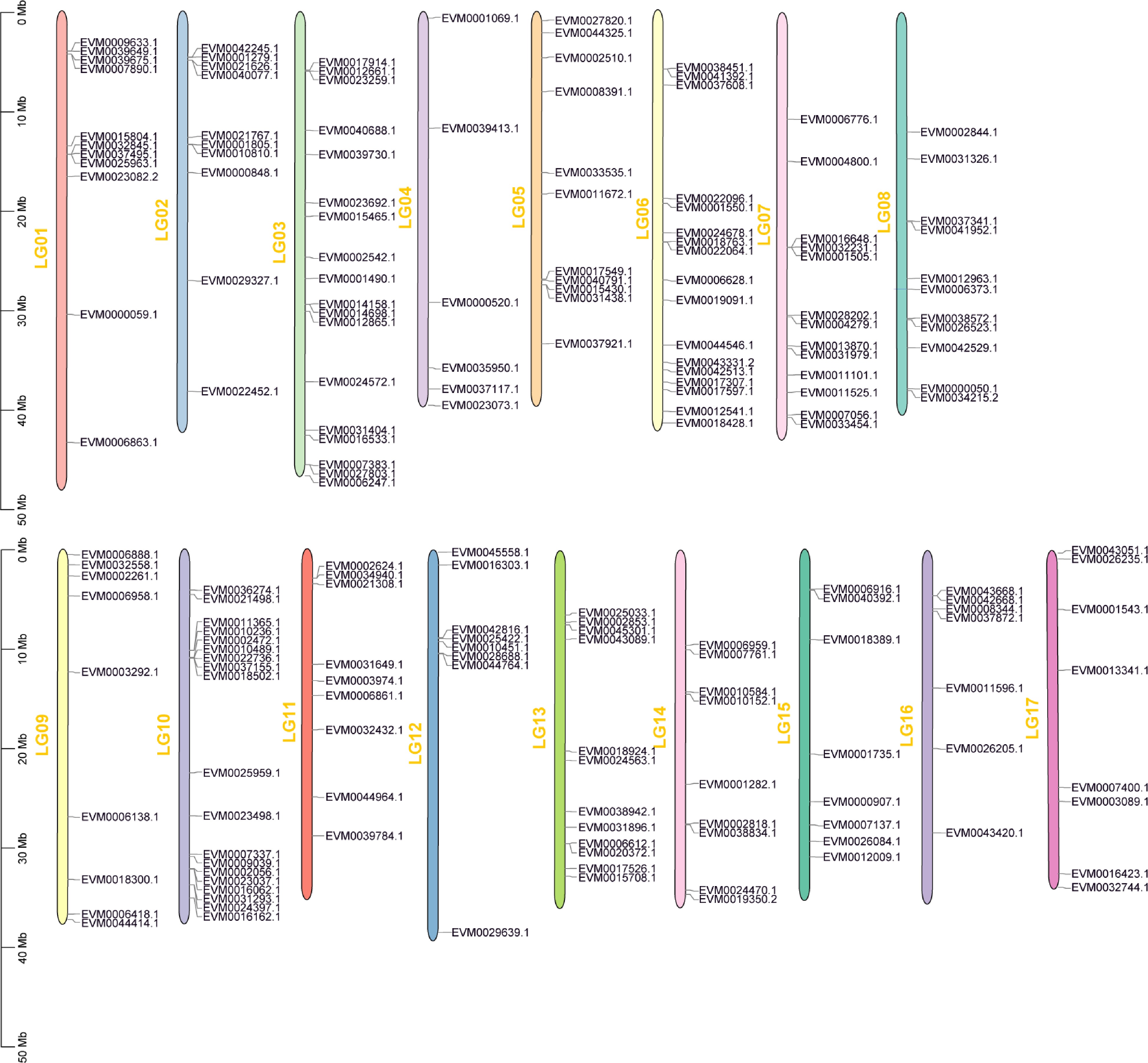
Figure 2.
Chromosomal location of the AP2/ERF genes in loquat.
Structural and motifs analysis of the loquat AP2/ERF
-
To further understand the gene structure and evolutionary relationships of the AP2/ERF gene family, the structural characteristics and conserved motifs of these genes in loquat were analyzed. The conserved motif analysis of the AP2/ERF protein was performed by MEME online sites. Ten motifs were observed in 189 AP2/ERF protein sequences of loquat (Fig. 3b, Supplemental Fig. S1). Among them, motif 6 existed in 167 genes (accounting for 88.36%), and 163 genes (86.24%) simultaneously contained motif 1, motif 2, and motif 5. However, motif 4 was only presented in 22 genes (11.64%), which belong to the AP2 subfamily. Motif 10 only existed in A1, A4, and A6 groups of the DREB subfamily. Motif 8 mainly existed in the DREB subfamily, including 31 members belonging to A1, A3, A4, A5, and A6 groups. It is worth noting that there was only one motif present in the sequences of EVM0021498.1 and EVM0002261.1, which was motif 1 and motif 4, respectively.
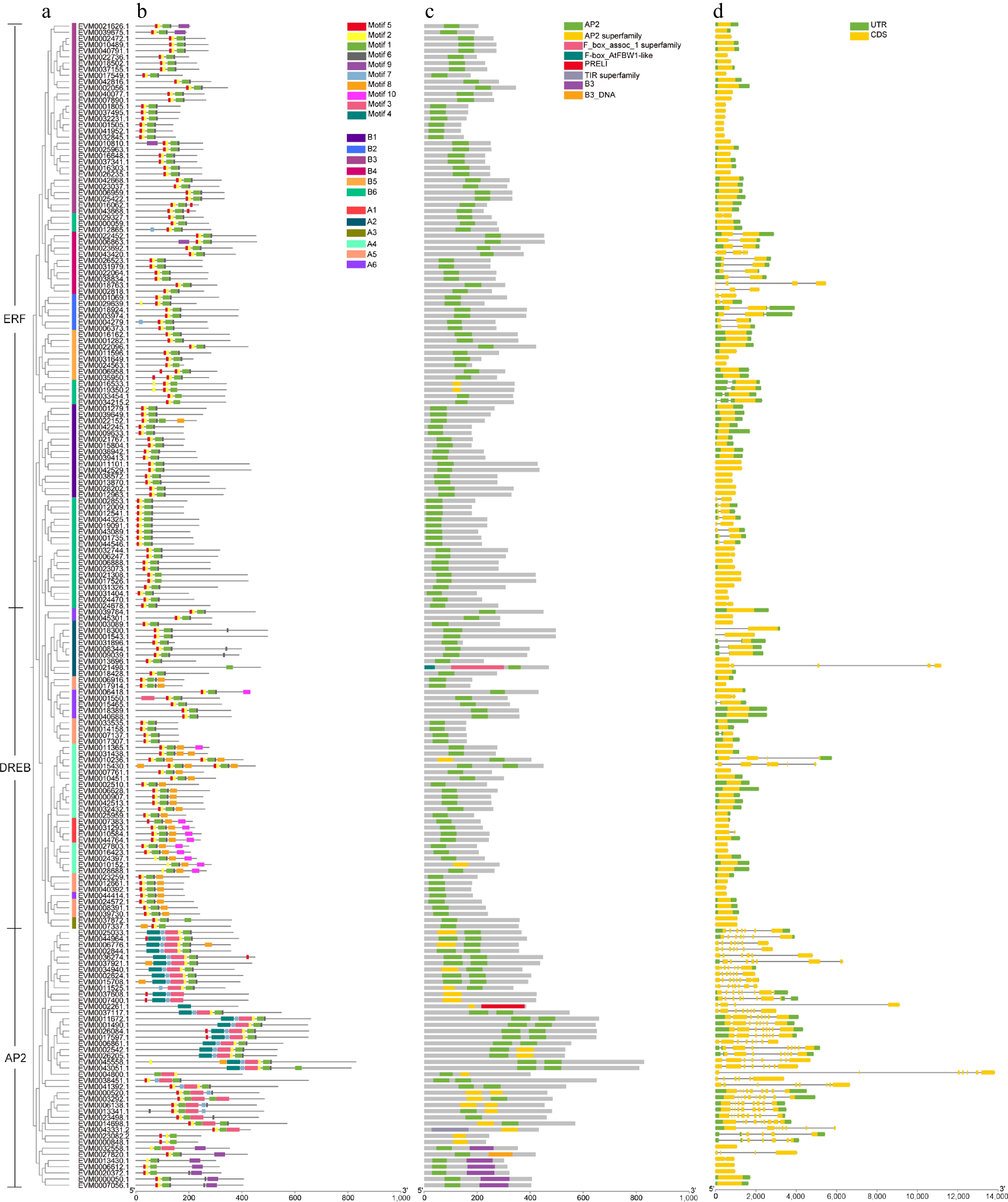
Figure 3.
Conserved motif and gene structure of loquat AP2/ERF gene family. (a) The phylogenetic tree of AP2/ERF genes. (b) Conserved motif distributions of AP2/ERF proteins. (c) Domains of loquat AP2/ERF genes. (d) Exon-intron distributions of AP2/ERF genes. The green and yellow boxes represent UTR and CDS, respectively.
The results of the conserved domain revealed that all loquat AP2/ERF genes had an AP2 domain (Fig. 3c). As shown in Fig. 3d, gene structural analysis showed that not all AP2/ERF genes had at least one intron, of which 109 out of 189 genes (57.67%) exhibiting no intron presence, including all genes in the A3 group of the DREB subfamily and the B1, B3, and B5 groups of the ERF subfamily. The AP2 subfamily had the highest number of introns, with 1~9 introns presented in all 37 gene members except for five members without introns. The number of exons ranged from 1 to 10. Notably, 24.87% of AP2/ERF gene family members had one or two exons. The genes with the highest number of exons were EVM0014698.1 and EVM0003292.1, each containing 10 exons. These findings emphasized the conservation of the gene structure of AP2/ERFs during the evolutionary process.
Synteny analysis of AP2/ERF genes in loquat
-
To explore the evolution of AP2/ERF genes in loquat, the tandem and segmental duplication events of these genes were performed by MCScanX. As shown in Fig. 4, a total of 3,068 collinear gene pairs in the loquat genome, among which 175 duplicated gene pairs were found in the AP2/ERF gene family of loquat (Supplemental Table S2). Subsequently, the Ka/Ks values of these 175 AP2/ERF gene pairs were calculated as indicators of selection pressure during gene evolution. A Ka/Ks ratio less than 1.0 indicated that the gene was subject to purification selection, while Ka/Ks ratio greater than 1.0 indicated a positive selection of the gene. A total of 157 (83.10%) homologous gene pairs exhibited ratios below 1.0 (Supplemental Table S3), indicating that these genes had undergone purification selection pressure.
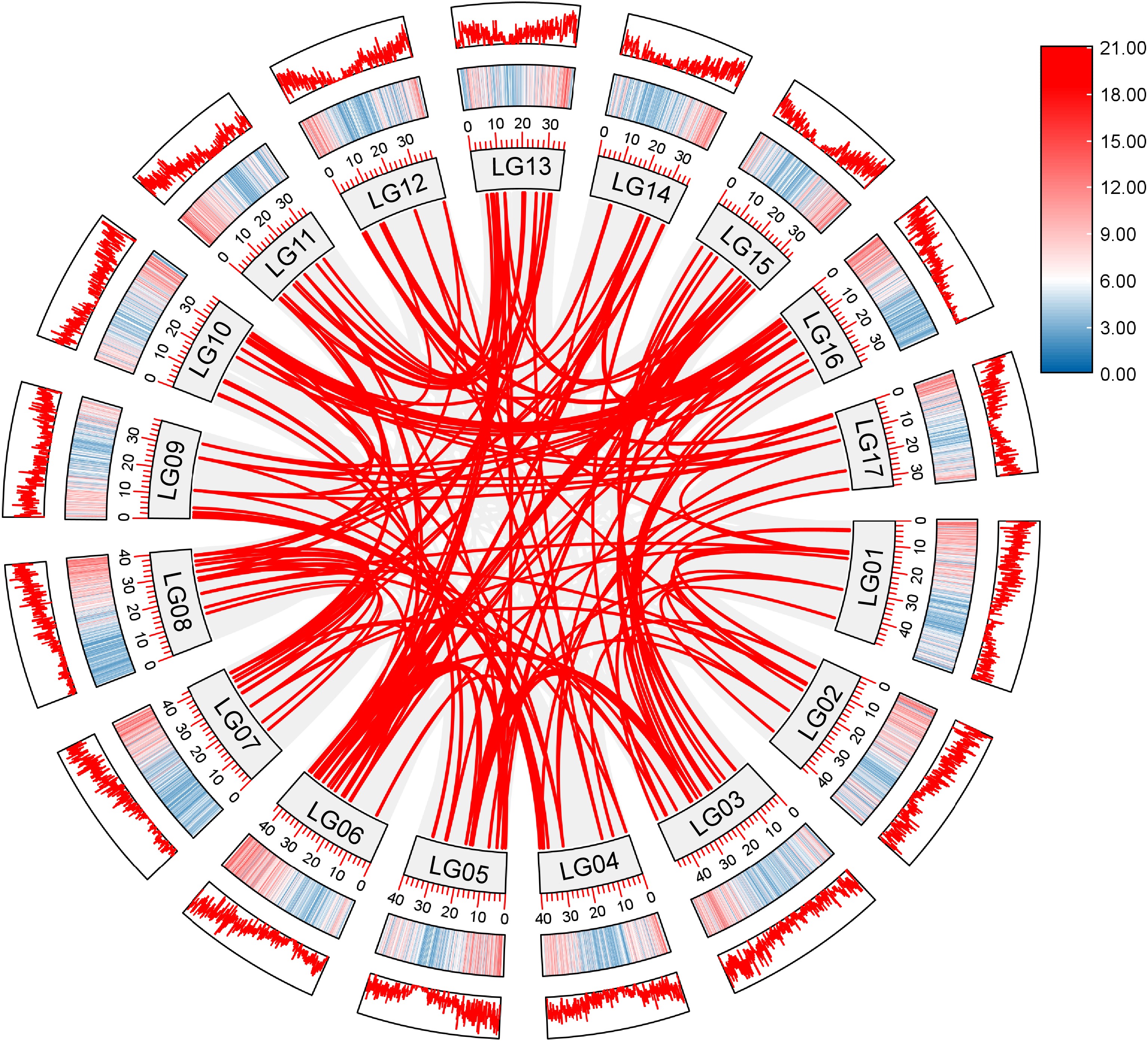
Figure 4.
Synteny analysis of AP2/ERF genes in loquat. Grey lines indicate all syntenic gene pairs in the loquat genome, red lines indicate collinearity relationships among loquat AP2/ERF genes. LG represents the different chromosome. Red and blue represent the level of gene distribution density on the chromosome. The deeper color means the higher density of genes.
To further elucidate the phylogenetic mechanism of the AP2/ERF family, collinear analysis was performed on the AP2/ERF genes of Arabidopsis, pomegranate (Punica granatum L.), and peach (Prunus persica L.), as well as the AP2/ERF genes of loquat. As shown in Fig. 5, the results suggested that there were 208 syntenic AP2/ERF gene pairs between loquat and Arabidopsis, 261 between loquat and pomegranate, and 236 between loquat and peach, respectively (Supplemental Table S4). Overall, these findings provided new insights into the evolution of fruit AP2/ERF genes.
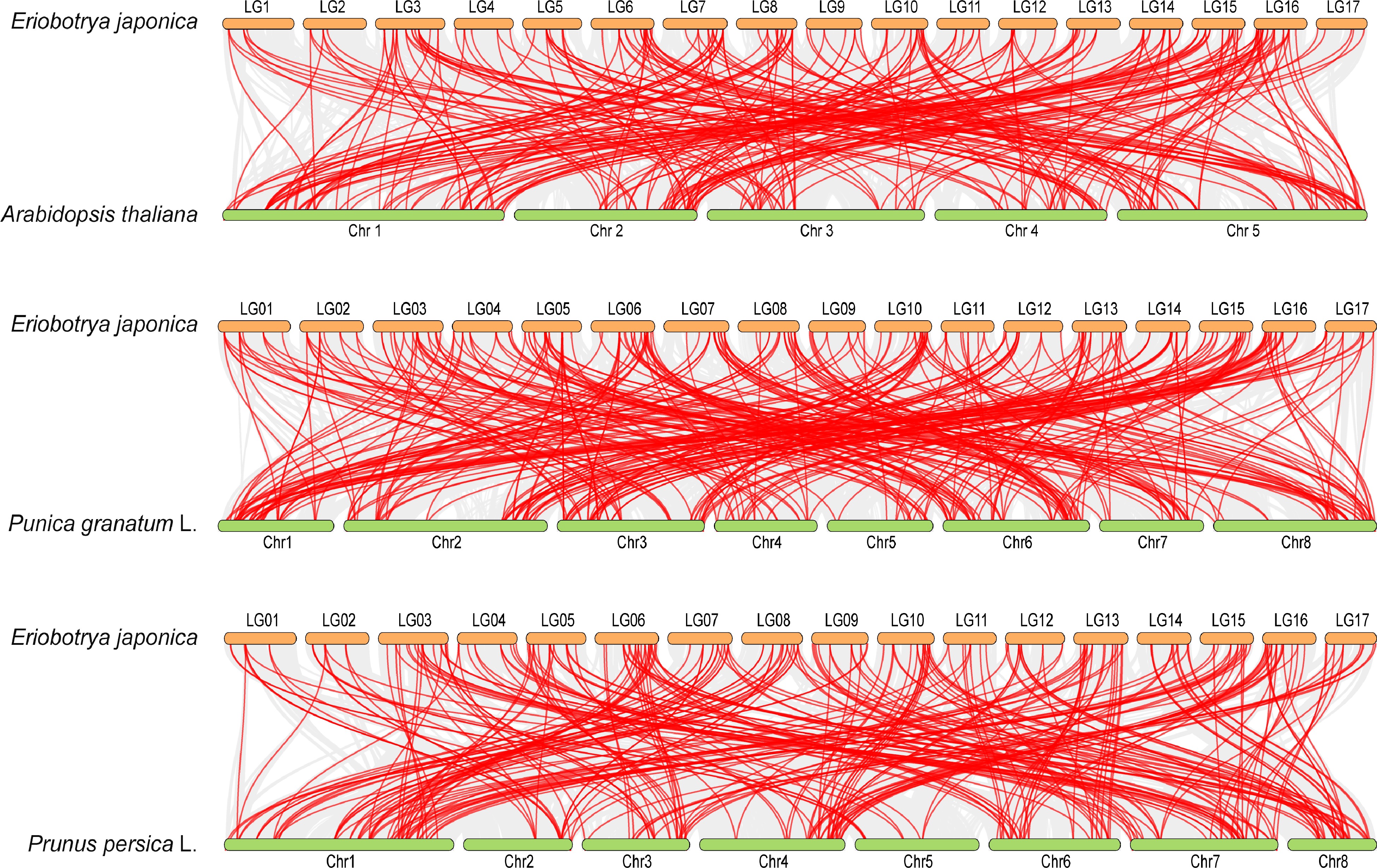
Figure 5.
Synteny analysis of AP2/ERF genes between loquat and other plants. Grey lines in the background represent the collinear blocks between different plant genomes, and the red lines highlight the syntenic AP2/ERF gene pairs. The three species respectively indicate Arabidopsis, pomegranate (Punica granatum L.), and peach (Prunus persica L.).
Cis-elements in the promoters of AP2/ERF genes in loquat
-
To further understand the potential functions and regulatory patterns of the loquat AP2/ERF genes, the cis-elements in the promoter sequences were analyzed (Fig. 6). It was found that there were four AP2/ERF promoters without cis-acting elements, including two ERF subfamily members EVM0022152.1 and EVM0025422.1, one AP2 subfamily member EVM0013430.1, and one DREB subfamily member EVM13696.1. Various cis-regulatory elements were detected in 185 AP2/ERF promoters, which were classified according to their function, including hormone-responsive elements, light-responsive elements, and abiotic stress response elements. The light-responsive elements were widely present in the AP2/ERF gene family, with a total of 487 light responsive elements in the AP2 subfamily, accounting for 44.68% of the total cis-acting elements in the AP2 subfamily. In the ERF subfamily, there were 1,204 light responsive elements (42.73%), and 641 in the DREB subfamily (43.49%). Hormone responsive elements were the most abundant in the ERF subfamily, with 340 (12.07%) ABA-responsive elements, 358 (12.70%) MeJA responsive elements, and 70 (2.48%) auxin responsive elements. Most AP2/ERF promoters had multiple cis-acting elements, among which EVM0009633.1 had the most cis-acting elements with 60.
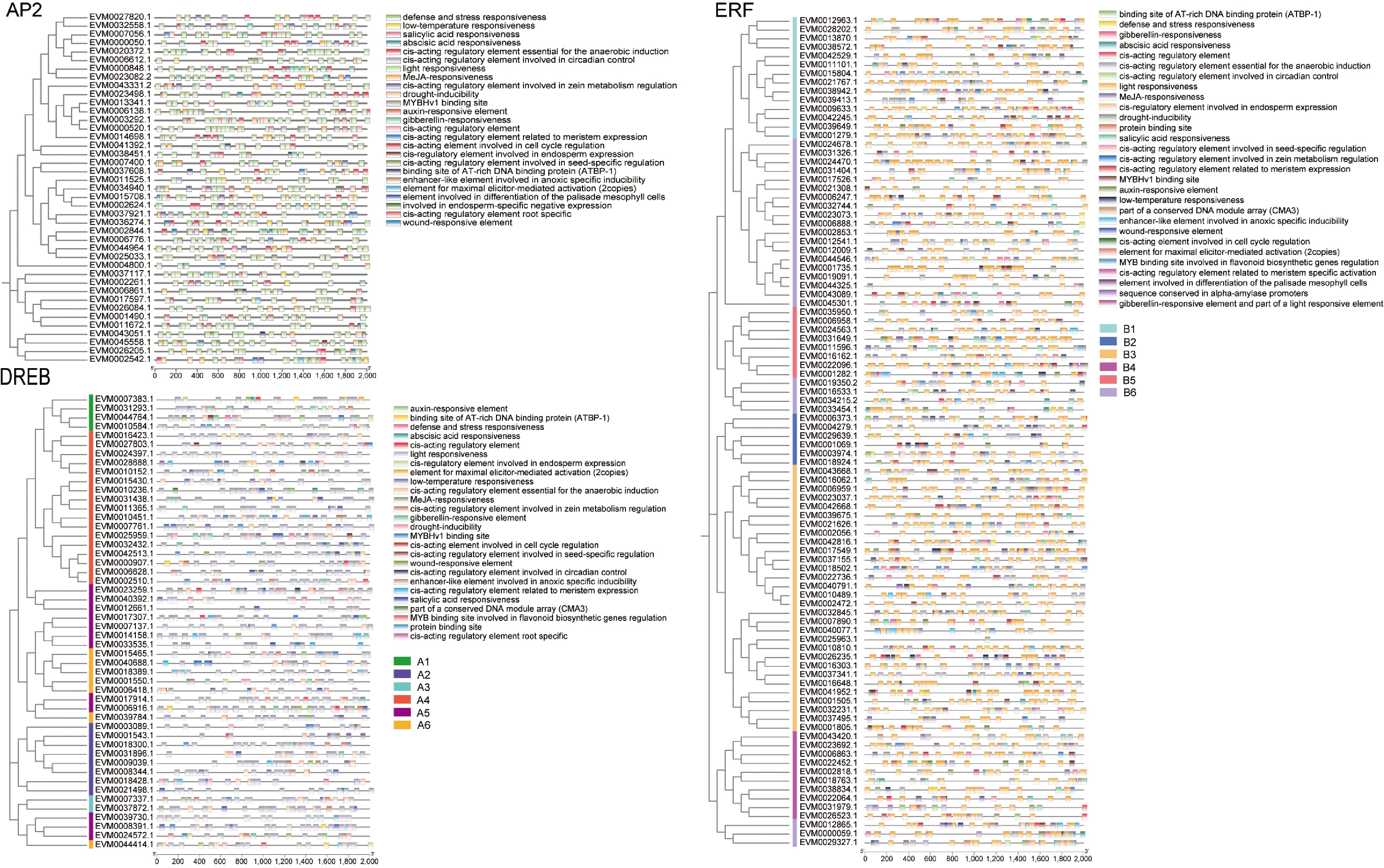
Figure 6.
Cis-acting elements in the promoter region of AP2/ERF genes in loquat. The 2,000 bp promoter region of the gene was analyzed. Different colors boxes represent different types of cis-acting elements and different groups of gene subfamilies.
In summary, cis-element analysis suggested that AP2/ERF genes may respond to different hormone responses and environmental stress, and be regulated differently during plant growth and development.
Expression pattern analysis of AP2/ERF genes in loquat fruit
-
To elucidate the specific spatiotemporal expression patterns of AP2/ERF genes, the expression profiles of AP2/ERF family members in different cultivars and developmental stages of loquat fruits were analyzed (Fig. 7). A total of 151 AP2/ERF genes were expressed in the two cultivars of loquat fruits. It is worth noting that the expression levels of 49 genes exhibited variety specificity. Among them, 29 members were mainly expressed in 'YS' fruits, including 14 members belonging to the ERF subfamily, six members belonging to the AP2 subfamily, and nine members belonging to the DREB subfamily. In addition, 20 genes were highly expressed in 'ZH 16' fruits, including nine ERF genes, four AP2 genes and seven DREB genes. Moreover, the AP2/ERF gene family exhibited diverse expression patterns during fruit development and maturation, with a total of 33 genes highly expressed in mature fruits, including 16 ERF genes, 10 DREB genes, and seven AP2 genes, and 40 genes were mainly expressed in early fruit development, including 20 ERF genes, 10 DREB genes, and 10 AP2 genes. However, the AP2 subfamily member EVM0006861.1 was significantly expressed in mature fruits of both varieties. These results suggested that the AP2/ERF gene family was closely associated with the physiological processes of fruit ripening.
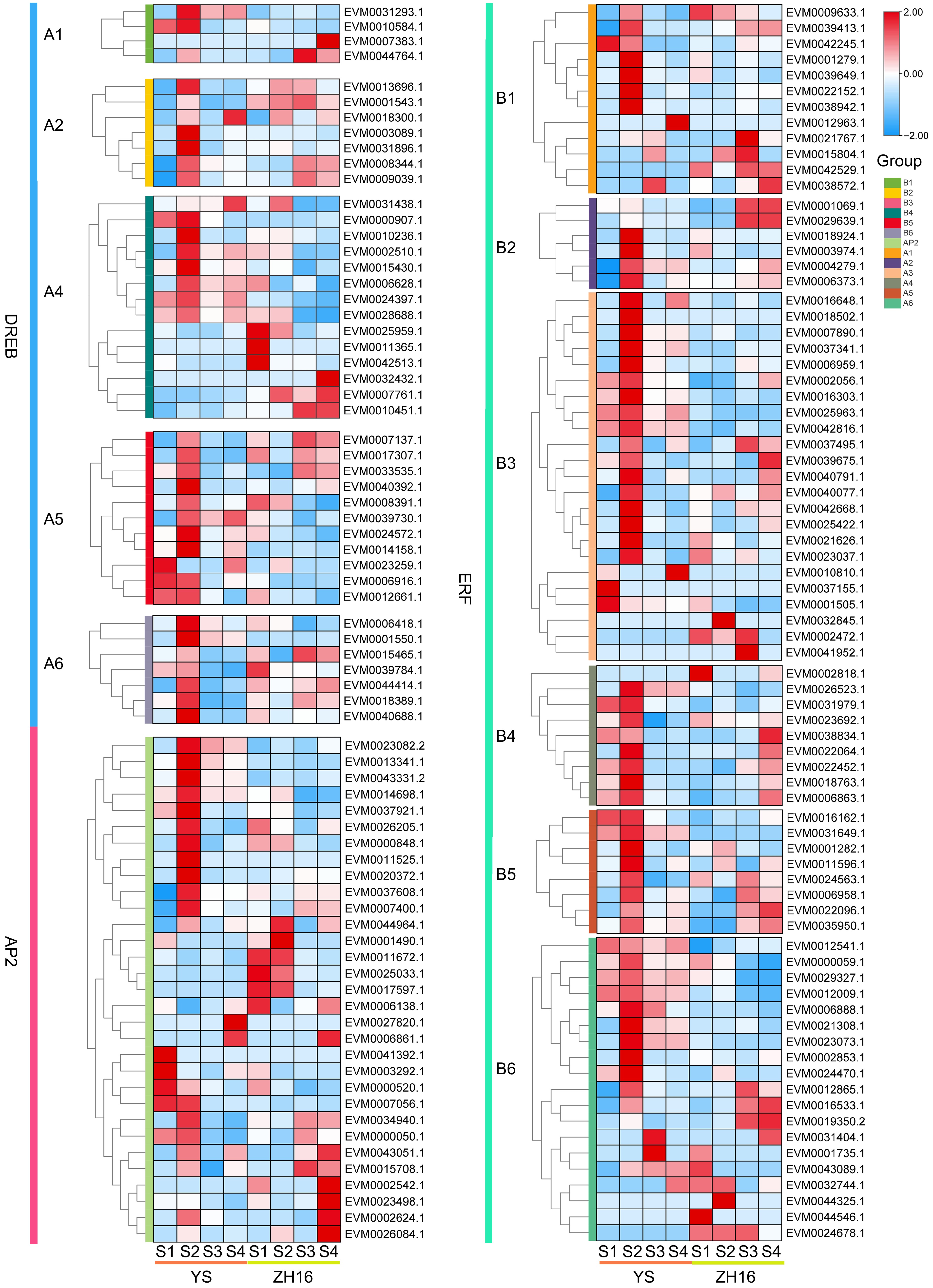
Figure 7.
The expression pattern analysis of the AP2/ERF gene family in different cultivars and developmental stages of loquat fruits. Red color represents high expression, while blue represents low expression. Fruits were harvested at four developmental stages (S1−S4), representing the expansion stage, the turning stage, the mature yellow stage, and the ripening stage.
-
The AP2/ERF gene family is one of the largest transcription factor families, and serve as crucial regulators in numerous biological processes[10,29,30]. However, the genome-wide analysis of the AP2/ERF gene family has not been reported in loquat (Eriobotrya japonica), a fruit tree with important economic and nutritional value.
In this study, a total of 189 AP2/ERF genes were observed in the loquat genome. The number of AP2/ERF gene family members was higher than that of cultivated persimmon (Diospyros oleifera) (157)[31], pearl millet (Pennisetum glaucum) (167)[32], and Morus notabilis (106)[33], but lower than that of sand pear (Pyrus pyrifolia) (234)[34]. There was no significant difference in AP2/ERF gene numbers between loquat and mung bean (Vigna radiata L.)[35]. In addition, the AP2/ERF gene family of loquat is divided into three subfamilies, while citrus is divided into four subfamilies[36]. The AP2/ERF genes of melon[17], and peach[16] were also divided into three subfamilies, but different from loquat. The three subfamilies of loquat were AP2, ERF, and DREB, while the three subfamilies of melon and peach were AP2, ERF, and RAV. Thus, the large variation in the number of AP2/ERF members among different plant species was likely due to gene duplication or loss during gene evolution[37]. The AP2/ERF gene family were unevenly distributed on all chromosomes, with the most on Chr 6 and Chr 10, while Chr 4 had the lowest number. A similar distribution pattern has been observed for AP2/ERF genes in other plant species, such as cotton[38], and sweet potato[39].
Previous studies have highlighted the close relationship between gene function and conserved domains within amino acid sequences[40]. To better understand the structural attributes of AP2/ERF in loquat, an analysis of their conservation patterns and intron-exon distributions was conducted. Gene structure analysis showed that 57.67% of AP2/ERF genes had no introns. Among the three AP2/ERF subfamilies, the AP2 subfamily had the largest number of exons, up to 4−10. The number of exons in the ERF and DREB subfamilies was more conservative. These results were similar to those of AP2/ERF family members in tartary buckwheat[41]. Intron/exon location patterns provide clues to the evolution of AP2/ERF family members[42].
Gene duplication, as a pivotal driver for the swift expansion and evolutionary diversification of gene families, has facilitated the emergence of novel functions while aiding plants in adapting to diverse environmental stimuli throughout their life cycles[43]. Extensive investigations have elucidated gene duplication events within the AP2/ERF gene family across various plant species. Nonetheless, a comprehensive examination of gene duplication events within the AP2/ERF gene family in loquat remains absent. Therefore, to further understand the amplification mechanisms of the AP2/ERF gene family, this study analyzed gene duplication events in loquat, revealing a total of 175 gene pairs among AP2/ERF family members. In previous studies, 299 and 237 gene pairs were found in willow[7] and kiwi[12], respectively. The gene pairs of AP2/ERF gene family in loquat were less than those in willow and kiwi. In addition, AP2/ERF family members in loquat and Arabidopsis thaliana, pomegranate, and peach shared 208, 261, and 236 pairs of homologous genes, respectively. These results indicated that the genetic relationship between loquat and pomegranate was the closest among these species. Ka/Ks values were computed for the 177 AP2/ERF gene pairs to measure selection pressure during gene evolution. Remarkably, 157 (83.10%) homologous gene pairs displayed ratios below 1.0, implying that these genes have been subjected to purifying selection pressure.
AP2/ERF has been shown to play an important role in plant growth, abiotic stress, and hormonal responses[44]. To further understand the potential functions and regulatory patterns of AP2/ERF gene family. The cis-elements of loquat AP2/ERF promoters were predicted. The results showed that 97.35% of AP2/ERF genes contained light responsiveness. 80.95% of AP2/ERF genes had MeJA-responsive elements in their promoter region. 49.74% of AP2/ERF genes contained gibberellin response elements. 42.86% of AP2/ERF genes had auxin-reactive elements. Similar results have been reported for AP2/ERF genes in other plants, such as all PgAP2/ERF promoters in pomegranate had light-responsive elements[11]. In conclusion, cis-element analysis suggested that AP2/ERF genes were responsive to different environmental stresses, and different subfamilies were regulated differently during plant growth and development.
Gene expression patterns were essential for the prediction of gene function. In previous studies, a total of 140 pear AP2/ERF gene family members (73.29% of the total) were identified to be expressed in at least one stage of pear fruit development, and the expression patterns of these genes in pear fruit were varied[8]. One of the pear AP2/ERF members, PuERF2, was upregulated through ethephon treatment, promoting the expression of PuGH3.1 to regulate IAA metabolism during pear fruit ripening[45]. In other species, only 83 FcAP2/ERF genes (accounting for 69.75%) were expressed at different developmental stages in fig (Ficus carica)[46], while 115 RsAP2/ERF genes (accounting for 95.83%) were expressed at different developmental stages of Rhododendron[47]. In this study, a total of 151 AP2/ERF genes (accounting for 79.89%) expressed in loquat fruits. Among them, 49 AP2/ERF genes were specifically expressed in different loquat cultivars, including 29 genes that were highly expressed in 'YS' fruits, and 20 genes that were expressed in 'ZH 16' fruits. The AP2/ERF gene family also exhibited diverse expression patterns during loquat fruit development and ripening, indicating that they play an important role in the physiological processes of fruit growth and maturation.
-
In the present study, a systematic genome-wide identification and analysis of the AP2/ERF genes in loquat was carried out. A total of 189 AP2/ERF members were identified in the loquat genome and further divided into three subfamilies (AP2, ERF, and DREB). The gene structures, conserved motif, syntenic, cis-elements, and gene expression patterns of the AP2/ERF genes were analyzed. Cis-element analysis suggested that AP2/ERF genes may respond to different hormone responses and environmental stress, including light responsiveness, low-temperature responsiveness, and various hormones. The differential expression of AP2/ERF genes in different cultivars and their developmental stages reflected gene diversity and variety specificity. In summary, the results of this study displayed the first comprehensive analysis of AP2/ERF genes in loquat, and this study provides a foundation for future research on the function of AP2/ERF genes in loquat.
-
The authors confirm contribution to the paper as follows: study conception and design: Wu B, Chen J, Xu K; data collection: Lin Y, Chen M, Zhang H, Fan X; technical assistance: Chen J, Xu K; analysis and interpretation of results: Liu Y, Cai L; draft manuscript preparation: Liu Y, Wu B. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed in this study are included in this published article and supplementary information files. The datasets analyzed during the current study are available in the NCBI Sequence Read Archive database (project number: PRJNA1109454). Further enquiries can be directed to the corresponding author.
This work was funded by the Key Scientific and Technological Grant of Zhejiang for Breeding New Agricultural Varieties (2021C02066-3), the Natural Science Foundation of Zhejiang province (LQ21C150001) and the Scientific Research and Development Foundation of Zhejiang A&F University (2019FR046). We thank Dr. Shaojia Li (Zhejiang University, China) for the critical revisions to the manuscript.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yumeng Liu, Linqi Cai, Xiurun Fan
- Supplemental Table S1 The AP2/ERF gene family in loquat.
- Supplemental Table S2 Gene duplication pairs of the AP2/ERF genes within the loquat genome.
- Supplemental Table S3 Ka/Ks ratios of the duplicated AP2/ERF genes in loquat.
- Supplemental Table S4 Syntenic gene pairs between AP2/ERF genes in loquat and those species.
- Supplemental Fig. S1 Sequence information for each motif (Motif 1–Motif 10).
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Cai L, Fan X, Zhang H, Chen M, et al. 2024. Genome-wide identification, evolutionary expansion and expression divergence of the AP2/ERF gene family in loquat (Eriobotrya japonica). Fruit Research 4: e034 doi: 10.48130/frures-0024-0028 

Genome-wide identification, evolutionary expansion and expression divergence of the AP2/ERF gene family in loquat (Eriobotrya japonica)
- Received: 16 May 2024
- Revised: 22 August 2024
- Accepted: 26 August 2024
- Published online: 08 October 2024
An Author Correction to this article was published on 9 December 2024, https://doi/10.48130/frures-0024-0038.
Abstract: Loquat (Eriobotrya japonica) is a subtropical evergreen fruit tree with high economic benefits and nutritional value. AP2/ERF (APETALA2/ethylene responsive factor) are a wide range of transcription factors found in plants, which play an important role in regulating plant development and biological processes. However, the systematic identification and characterization of AP2/ERF genes in loquat remains unclear. In the present research, a total of 189 loquat AP2/ERF members were identified. According to the phylogenetic tree, these members were divided into three subfamilies, including AP2 (42), ERF (96), and DREB (51). Except for three genes located on the skeleton, most of the loquat AP2/ERF genes were unevenly distributed across all chromosomes. The collinearity results showed that 175 duplicate gene pairs were found in the AP2/ERF gene family of loquat. Ka/Ks results indicated that these genes underwent purification selection pressure. The syntenic analysis of AP2/ERF genes between Arabidopsis, pomegranate, peach, and loquat provided valuable clues for the potential evolution of AP2/ERF family in loquat. Cis-element analysis showed that most of the AP2/ERF genes in loquat had multiple cis-elements related to stress response, plant hormone signal transduction, and plant growth and development. Transcriptomic data indicated that the expression patterns of AP2/ERF genes exhibited diversity in different loquat cultivars during fruit development and ripening. The results of this study displayed the first comprehensive analysis of AP2/ERF genes in loquat, which provides a solid foundation for future research on the function of AP2/ERF genes in loquat.
-
Key words:
- Loquat /
- AP2/ERF /
- Phylogenetic analysis /
- Expression patterns