








-
Dermateaceae (Helotiales, Leotiomycetes) consists of 16 genera Coleophoma Höhn., Corniculariella P. Karst., Cryptosporiopsis Bubák & Kabát, Davidhawksworthia Crous, Dermea Fr., Gelatinoamylaria Prasher & R. Sharma, Neodermea W.J. Li, D.J. Bhat & K.D. Hyde, Neofabraea H.S. Jacks., Neogloeosporidina W.J. Li, Camporesi & K.D. Hyde, Pezicula Tul. & C. Tul., Phlyctema Desm., Pseudofabraea Chen Chen, Verkley & Crous, Pseudoxenochalara Iliushin & Kirtsideli, Rhizodermea Verkley & Zijlstra, Verkleyomyces Y. Marín & Crous, and Xenochalara M.J. Wingf. & Crous[1−6]. Species of Dermateaceae are predominantly reported from temperate regions of the world and occupy various ecological niches. They are reported as plant pathogens, endophytes, saprobes, wood destroyers, and soil inhabitants[2−4,6,7]. Dermea is the type genus of the family Dermateaceae. The sexual morph of Dermea is characterized by leathery, hard, dark brown to black apothecia. Its asci are 8-spored, cylindrical to clavate-cylindrical while the ascospores are aseptate to 3-septate, ellipsoid-fusiform to ellipsoidal and hyaline to yellowish-brown in colour[8−10]. The asexual morph exhibits diverse conidiomatal structures, which often accompany the apothecia. Two distinct types of conidia are typically observed: bacillary to filiform microconidia and elongate-fusiform to sickle-shaped macroconidia[8−10].
Phlyctema was introduced by Desmazières[11] with P. vagabunda Desm. as the type species. The sexual morph of P. vagabunda was previously classified under Neofabraea alba (E.J. Guthrie) Verkley, leading to the synonymization of Phlyctema with Neofabraea[12,13]. However, Chen et al.[2] later reinstated Phlyctema and Neofabraea as separate genera based on analyses of multi-genes and morphological traits. Subsequently, Li et al.[4] employed multi-gene phylogenetic analyses to further clarify the generic boundaries within Dermateaceae, providing support for the distinction between Phlyctema and Neofabraea. There are 61 accepted species in Phlyctema, although most have not been re-examined[14]. The sexual morph of Phlyctema is characterized by circular or irregular and merged, slightly convex, sessile, developing from acervuloid stromata apothecia, 8-spored, cylindrical-clavate asci, hyaline, elongated ellipsoid, straight or slightly curved, rounded or somewhat pointed at both ends, aseptate when young, septate at maturity ascospores, filiform, branched, slightly swollen at apical cell, with septate paraphyses[2,12,15]. However, the sexual morph is rarely been reported from the Phlyctema species. The asexual morph is characterized by having eustromatic, convulated, pulvinate to sporodochial conidiomata, branched, hyaline conidiophores, and phialidic conidiogenous cells that give rise to hyaline, aseptate, fusiform, straight to curved conidia[4,16]. Phlyctema species were reported as saprobic, parasitic, or endophytic on plant hosts in terrestrial habitats, such as Aconitium napellus L. (Ranunculaceae), Coronilla L. (Fabaceae), Erigeron L. (Asteroideae), Libertia ixioides (G. Forst.) Spreng. (Iridaceae), Malus pumila Mill., and M. sylvestris (L.) Mill. (Rosaceae)[2,4,16,17]. Although a considerable number of species have been reported in the genus Phlyctema, those associated with grassland vegetation are relatively scarce or nonexistent.
Grassland ecosystems across the world, including in China, offer numerous opportunities for fungal discovery[18−25]. For instance, bamboo ecosystems in China host more than 450 fungal species, accounting for approximately one-quarter of the global bambusicolous ascomycetes[26]. However, the investigation of fungi (such as in Dermateaceae) associated with short grass, bushes, and other grass vegetation remains underexplored. Extensive fungal sampling across a variety of grasslands in different geographical areas is urgently needed to mitigate species losses and understand their ecological significance. Therefore, we are continuously exploring the grassland-associated microfungi in Yunnan, China[25,27,28].
In this study, a taxonomic survey of foliar fungal epiphytes was conducted on herbaceous plants from grasslands in Yunnan, China. A new species, Phlyctema yunnanensis is described and illustrated based on its morphological characteristics and DNA molecular evidence. Additionally, an updated multigene phylogenetic tree for Phlyctema is presented, incorporating combined ITS, LSU, rpb2, and tub2 sequence data, including the newly identified taxon.
-
Specimens were collected from grasslands in Zhaotong City, Yunnan Province, China in 2021. The local environment in Zhaotong is characterized by Poaceae as the predominant plant species and features typical plateau vegetation[24,25]. This area is influenced by a three-dimensional monsoon climate and reaches a maximum elevation of approximately 4000 m[29]. Specimens were stored in plastic Ziplock bags and returned to the mycology laboratory at the Kunming Institute of Botany, Chinese Academy of Sciences (Kunming, China). Samples were examined using an Olympus SZ-61 dissecting microscope. Fungal fruiting structures were manually sectioned with a sterilized razor blade and mounted in water on a slide for microscopic examination. Micro-morphological characteristics were examined using a Nikon ECLIPSE Ni-U complex microscope with differential interference contrast (DIC) and phase contrast (PC) illumination. Photos of microscopic structures were captured using a Nikon DS-Ri2 camera. Photo plates and measurements were processed using Adobe Photoshop CS6 Extended version 13.0.1 (Adobe Systems, CA, USA). Specimens were deposited in the herbarium of the Kunming Institute of Botany Academia Sinica (HKAS) (Kunming, China). The Index Fungorum and Faces of Fungi (FoF) numbers were obtained for the new fungus[14,30].
DNA extraction, PCR amplification, and sequencing
-
Genomic DNA was extracted from fresh fruiting bodies using the E.Z.N.A.® Forensic DNA Kit-D3591 (Omega Bio-Tek, Inc, 400 Pinnacle Way, Suite 450 Norcross, Georgia 30071, USA) following the manufacturer's protocol. The DNA for polymerase chain reaction (PCR) was stored at 4 °C for regular use and at −20 °C for long-term usage. PCR amplifications were carried out for the partial 28S large subunit nuclear ribosomal DNA (LSU) and internal transcribed spacer region with intervening 5.8S nrRNA gene (ITS) were amplified with primers LR0R/LR5[31], ITS5/ ITS4[32], separately. The total volume of PCR mixtures for amplification was 25 μL containing 8.5 μL ddH2O, 12.5 μL 2 × F8FastLong PCR MasterMix (Beijing Aidlab Biotechnologies Co. Ltd, Beijing, China), 2 μL of DNA template, and 1 μL of each forward and reverse primers (stock of 10 pM). The PCR thermal cycle profiles for LSU and ITS: the thermal conditions included initial denaturation at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 10 s, annealing temperatures at 55 °C for 15 s, elongation at 72 °C for 20 s, and final extension at 72 °C for 10 min. The amplified PCR fragments were sent to the Qingke Company, Kunming City, Yunnan Province, China, and Shanghai Sangon Biological Engineering Technology and Service Co., Ltd., China, for sequencing. Sequences were deposited in the GenBank Database.
Sequence alignment, and phylogenetic analyses
-
Two phylogenetic analyses were performed based on two concatenated datasets of ITS-LSU-rpb2-tub2 and ITS-LSU. Sequences generated from this study were analyzed with other similar sequences obtained from GenBank (
www.ncbi.nlm.nih.gov ) (Table 1), and those derived from recent publications[2,4,6,17,33]. The newly generated sequences in this study were assembled by BioEdit 7.0.9.0[34]. Multiple sequence alignments with individual gene datasets were generated with the MAFFT v.7. online platform[35], and trimmed with TrimAl v.1.3[36] via the web server Phylemon2 (http://phylemon.bioinfo.cipf.es/utilities.html ; accessed on August 10, 2024), and multi-gene alignments were manually aligned wherever necessary in SequenceMatrix program (1.7.8)[37]. Phylogenetic reconstructions of individual and combined datasets were performed using maximum likelihood (ML) and Bayesian inference (BI).Table 1. GenBank accession numbers of the strains used for phylogenetic analyses in this study.
Species Strain number ITS LSU rpb 2 tub2 Coleophoma caliginosa CBS 124806T KR859090 KR858881 KR859330 KR859293 Coleophoma camelliae CBS 101376T KU728481 KU728521 NA KU728597 Coleophoma coptospermatis CPC 19864T KU728483 KU728523 NA NA Coleophoma cylindrospora CBS 591.70T KU728486 KU728526 NA NA Coleophoma ericicola CBS 301.72T KU728488 KU728528 NA NA Coleophoma eucalypticola CBS 124810T GQ303279 GQ303310 KR859331 KR859294 Coleophoma eucalyptorum CPC 19294 KF251240 KF251743 NA NA Coleophoma paracylindrospora CBS 109074T KU728491 KU728531 NA NA Coleophoma parafusiformis CBS 132692T KU728494 KU728534 NA NA Coleophoma proteae CBS 132532T JX069866 JX069850 NA NA Corniculariella rhamni MFLUCC 16-1446T MT185495 MT183457 MT432216 NA Davidhawksworthia ilicicola CBS 734.94T NR_154008 NG_067307 NA NA Davidhawksworthia ilicicola CBS 261.95 KU728516 KU728555 NA NA Dermea cerasi MFLU 16-0929 MT185502 MT183464 MT432221 NA Dermea cerasi MFLUCC 16-1147T MT185501 MT183463 MT432220 NA Infundichalara microchona CBS 175.74T KR859078 HQ609479 KR859318 KR859284 Neodermea rossica MFLUCC 17-2506T MT185530 MT183493 MT432236 NA Neofabraea actinidiae CBS 121403T KR859079 KR858870 KR859319 KR859285 Neofabraea brasiliensis CNPUV499T KR107002 KR107002 NA KR107012 Neofabraea brunneipila MFLU 15-0231T MK584984 MK592004 NA NA Neofabraea inaequalis CBS 326.75T KR859081 KR858872 KR859321 KR859287 Neofabraea kienholzii CBS 126461T KR859082 KR858873 KR859322 KR859288 Neofabraea krawtzewii CBS 102867 KR859084 KR858875 KR859324 AF281459 Neofabraea malicorticis CBS 122030T KR859086 KR858877 KR859326 KR859291 Neofabraea perennans CBS 275.29 KR859088 KR858879 KR859328 KR859292 Neofabraea perennans CBS 453.64 KR859089 KR858880 KR859329 AF281474 Neofabraea salicina CBS 148445T OK664708 OK663747 OK651161 NA Neogloeosporidina pruni MFLU 16-2153T NR_169721 MT183501 MT432239 NA Pezicula acericola CBS 239.97 KR859093 KR858884 KF376214 KF376283 Pezicula aurantiaca CBS 201.46 KR859102 KR858893 KF376210 KF376335 Pezicula californiae CBS 124805T KR859104 KR858895 KR859332 KR859295 Pezicula carpinea CBS 923.96T KR859108 KR858899 KF376158 KF376279 Pezicula cinnamomea CBS 239.96 KR859124 KR858915 KF376165 KF376323 Pezicula cinnamomea CBS 240.96 KR859125 KR858916 KF376163 KF376325 Pezicula cornina CBS 285.39 KR859163 KR858955 KR859333 KR859296 Pezicula eucrita CBS 259.97 KR859179 KR858971 KF376205 KF376333 Pezicula fagacearum CBS 112400T KR859201 KR858993 KR859335 KR859298 Pezicula frangulae CBS 100244 KR859204 KR858996 KF376211 KF376285 Pezicula italica MFLU 16-1284T NR_170044 NG_073839 NA NA Pezicula microspora CBS 124641T KR859212 KR859004 KR859337 KR859300 Pezicula neocinnamomea CBS 100248T KR859213 KR859005 KF376209 KF376328 Pezicula neoheterochroma CBS 127388T KR859221 KR859013 KR859338 KR859301 Pezicula neosporulosa CBS 101.96T KR859223 KR859015 KF376193 KF376305 Pezicula ocellata CBS 268.39 KR859232 KR859024 KR859339 KR859302 Pezicula pseudocinnamomea CBS 101000T KR859235 KR859027 KR859340 KR859303 Pezicula rubi CBS 253.97 KR859250 KR859042 KF376204 KF376329 Pezicula sporulosa CBS 224.96T NR_137161 NG_069840 KF376201 KF376326 Phlyctema phoenicis MFLU 15-1243 MT449717 MT449705 MT432248 NA Phlyctema phoenicis CPC 29372T KY173432 KY173522 NA NA Phlyctema phoenicis CBS 147066 ON811524 ON811581 NA NA Phlyctema vagabunda CBS 109875T KR859275 KR859069 KR859346 AY064702 Phlyctema vagabunda CBS 304.62 KR859276 KR859070 KR859347 KR859310 Phlyctema vagabunda ATCC 38338 AF281366 NA NA AF281456 Phlyctema vincetoxici CBS 102469 KR859277 KR859071 KR859348 KR859311 Phlyctema vincetoxici CBS 123727T KR859278 KR859072 KR859349 KR859312 Phlyctema yunnanensis HKAS 128745T PQ456177 PQ456179 NA NA Phlyctema yunnanensis HKAS 128746 PQ456178 PQ456180 NA NA Pseudofabraea citricarpa CBS 130297T KR859279 KR859073 KR859350 KR859313 Pseudofabraea citricarpa CBS 130532 KR859280 KR859074 KR859351 KR859314 Pseudoxenochalara grumantiana CBS 148028 OM774424 OM776920 OM782293 NA Rhizodermea veluwensis CBS 110605T KR859282 KR859076 KR859353 KR859316 Rhizodermea veluwensis CBS 110615 KR859283 KR859077 KR859354 KR859317 Xenochalara juniperi MEA-B5-SW JX869564 NA NA NA Xenochalara juniperi CBS 670.75 AF184887 NA NA NA The new sequences are indicated in bold. The ex-type strains are indicated with 'T'. 'NA' indicates that the sequence is unavailable. Maximum likelihood trees were inferred using RAxML-HPC v.8 on ACCESS (8.2.12) in CIPRES Science Gateway v. 3.3 online platform[38] under the GTRGAMMA nucleotide substitution model with 1,000 bootstrap replicates. The BI analysis was performed using MrModeltest v2.3[39] via PAUP v. 4.0b10[40]. Six simultaneous Markov chains were run for 10,000,000 generations, with trees sampled every 1000th generation. The run was configured to stop automatically when the critical value for the topological convergence diagnostic reached 0.01, and the first 25% of the trees were discarded as burn-in.
Tree topologies were visualized and exported using FigTree v. 1.4.0[41]. The phylogram was edited and annotated using Microsoft Office PowerPoint 2016 (Microsoft Inc., USA) and Adobe Photoshop CS6 Extended version 13.0.1 (Adobe Systems, CA, USA). Finally, the newly generated sequences were deposited in the GenBank database (Table 1).
-
Since we failed to obtain rpb2 and tub2 under various PCR conditions, the analysis was initially conducted using only ITS and LSU data. A total of 64 strains of taxa are included in the combined analyses. The aligned dataset contained 1,253 characters (ITS: 1–476 bp, LSU: 477–1,253 bp). Infundichalara microchona (CBS 175.74) was used as the outgroup taxon. The best-scoring RAxML tree with a final likelihood value of -6894.552551 is presented. The matrix had 337 distinct alignment patterns, with 3.47% of undetermined characters or gaps. Estimated base frequencies were as follows: A = 0.233464, C = 0.235945, G = 0.285797, T = 0.244794; substitution rates AC = 1.549458, AG = 2.488198, AT = 0.862983, CG = 0.557730, CT = 6.974513, GT = 1.000000, gamma distribution shape parameter α = 0.531475, 0.543158, Tree-Length = 1.065237 (Fig. 1).
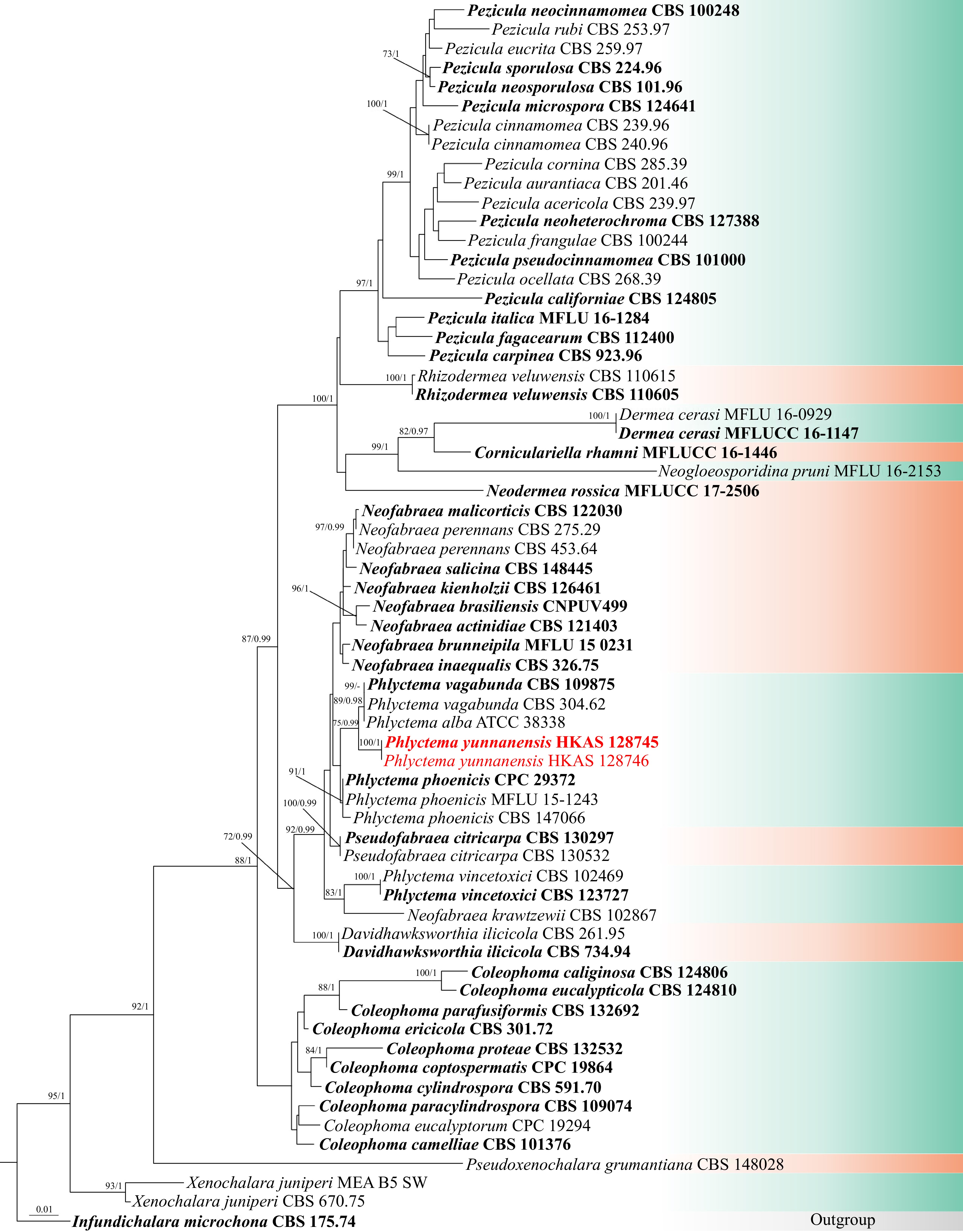
Figure 1.
Phylogram generated from ML analysis based on ITS and LSU sequence data representing Dermateaceae. The tree topology of the ML analysis is similar to the Bayesian analysis. Bootstrap values for ML equal to or greater than 70% and BYPP values greater than 0.90 (the rounding of values to two decimal proportions) are labeled at the nodes. Strains of the newly described species are in red, while type strains are in bold.
To follow up with the recent phylogenetic analyses of this family by Iliushin & Kirtsideli[6] a second analysis of Dermateaceae was constructed using sequence data from ITS, LSU, rpb2, and tub2 sequence data. The total sequence dataset comprised 64 strains, Infundichalara microchona (CBS 175.74) was selected as the outgroup taxon. The aligned dataset contained 2918 characters (ITS: 1–476 bp, LSU: 477–1,253 bp, rpb2: 1,254–2,126 bp, tub2: 2,127–2,918 bp). Phylogenetic relationships were inferred by conducting analyses using both ML and BI methods. The best-scoring RAxML tree was selected to represent the relationships amongst taxa, with a final likelihood value of -20239.20371. The matrix contained 1,007 distinct alignment patterns, with a 24.36% proportion of gaps and completely undetermined characters. The estimated base frequencies of A = 0.240797, C = 0.243480, G = 0.260336, T = 0.255387; substitution rates AC = 2.098811, AG = 4.359330, AT = 1.121395, CG = 0.807926, CT = 11.035437, GT = 1.000000, gamma distribution shape parameter α = 0.756016, I = 0.553584, Tree-Length = 1.966413 (Fig. 2).
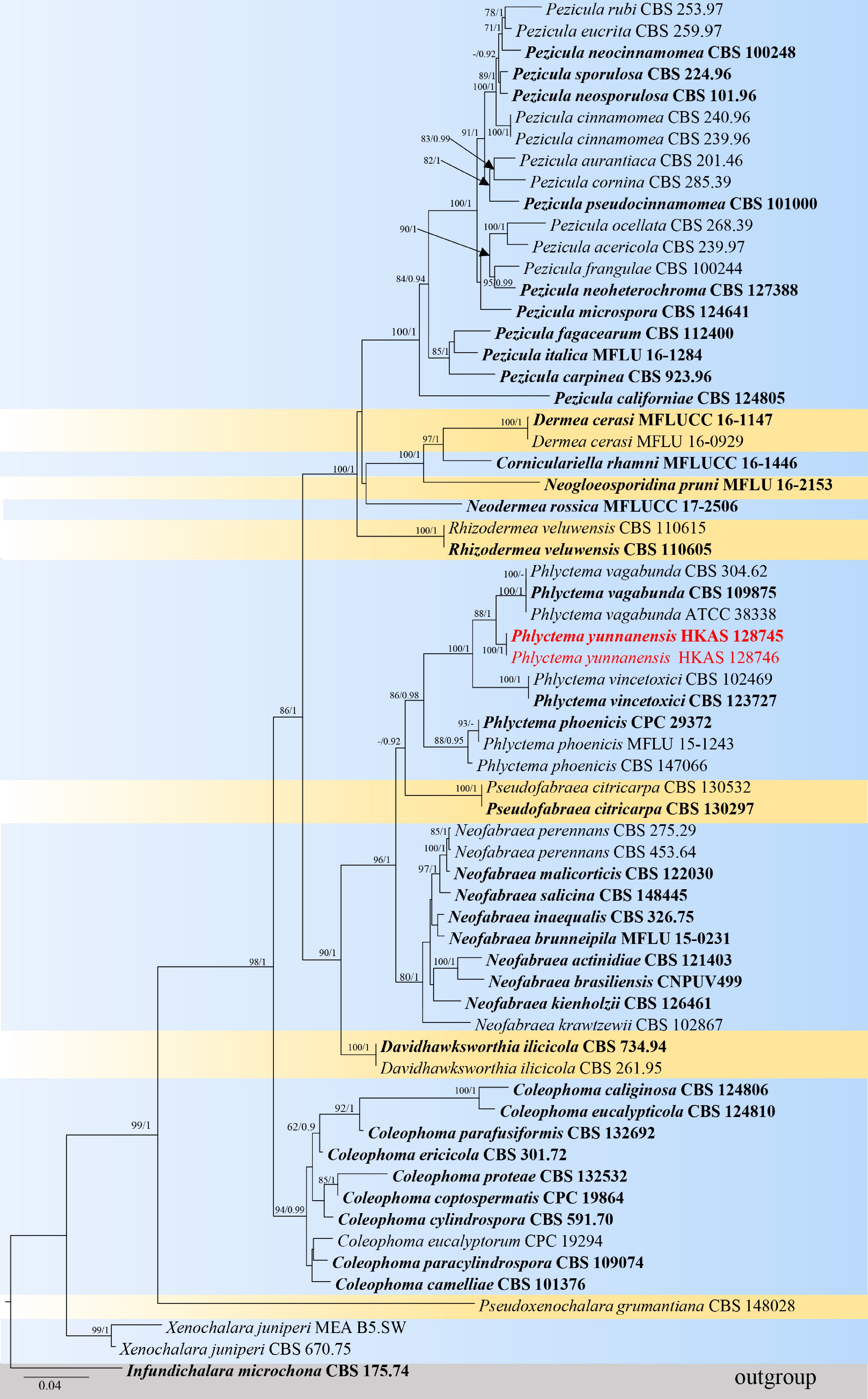
Figure 2.
Phylogenetic tree based on analysis of a combined ITS, LSU, rpb2, and tub2 sequence dataset. Bootstrap support values for ML equal to or greater than 70% and BYPP equal to or greater than 0.90 are shown as ML/BI above the nodes. The ex-type strains are in bold and the new isolate in this study is in red. The tree is rooted with Infundichalara microchona (CBS 175.74). The scale bar represents the expected number of nucleotide substitutions per site.
Based on the phylogenetic analyses of the combined LSU and ITS sequence data, the novel species Phlyctema yunnanensis (HKAS 128745 and HKAS 128746) is closely related to P. vagabunda (CBS 109875, CBS 304.62, and ATCC 38338) with 75% ML and 0.99 PP statistical support (Fig. 1). The multigene phylogenetic analyses of the concatenated ITS, LSU, rpb2, and tub2 sequence data showed that all representative species formed well-resolved clades in the present study. Phlyctema yunnanensis (HKAS 128745 and HKAS 128746) is a sister to P. vagabunda (CBS 109875, CBS 304.62, and ATCC 38338) with 88% ML and 1.00 PP support values (Fig. 2).
Taxonomy
-
Phlyctema yunnanensis Y. Gao, H. Gui & K.D. Hyde, sp. nov.
Index Fungorum number: IF 902420; Facesoffungi numbers: FoF 16782; Fig. 3.
Etymology: The specific epithet 'yunnanensis' refers to Yunnan Province (China), where the holotype was collected.
Holotype: HKAS 128745.
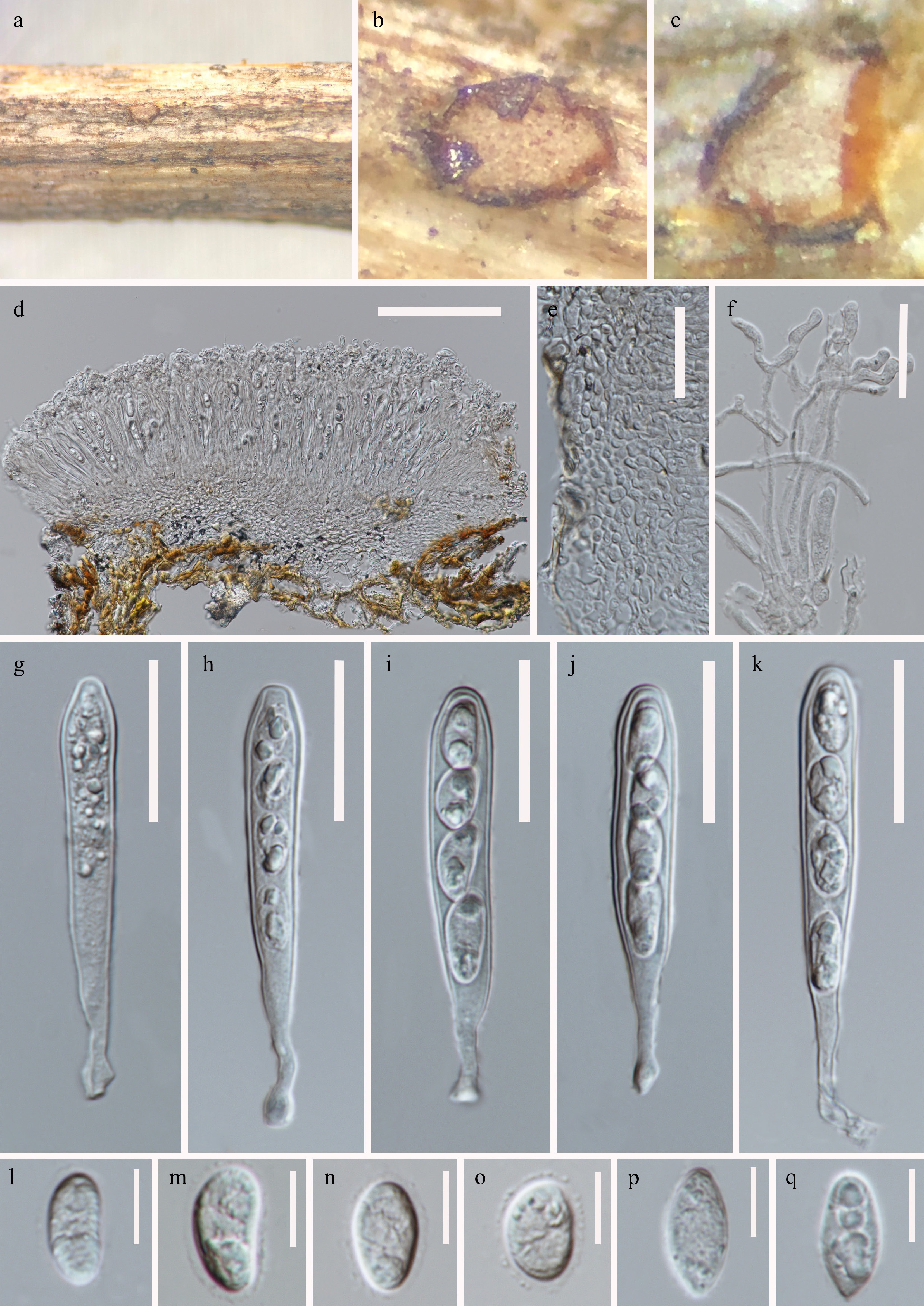
Figure 3.
Phlyctema yunnanensis (HKAS 128745, holotype). (a), (b) Apothecia on natural substrate. (c) Sections through apothecium. (d) Longitudinal section of apothecium. (e) Excipular structure of margin. (f) Paraphyses. (g)–(k) Asci. (l)–(q) Ascospores. Scale bars: (d) = 100 μm, (e)–(k) = 30 μm, (l)–(q) = 10 μm.
Saprobic on a decaying stem of herbaceous plants. Sexual morph: Apothecia 360–440 μm diam × 130–180 μm high (
$\bar {\rm x} $ $\bar {\rm x} $ $\bar {\rm x} $ $\bar {\rm x} $ $\bar {\rm x} $ $\bar {\rm x} $ $\bar {\rm x} $ Material examined: China, Yunnan Province, Zhaotong City, grassland (26°55'3''N, 103°8'36''E), on a decaying stem of herbaceous plants, 21 August 2021, Ying Gao, ZG16A (HKAS 128745, holotype); ibid., ZG16B (HKAS 128746).
Notes: Phlyctema yunnanensis is introduced as a new species based on its distinct morphology and phylogenetic analysis of combined ITS, LSU, rpb2, and tub2 sequence data. The new isolate clustered with P. vagabunda (strains CBS 304.62, CBS 304.62, and ATCC 38338) with support in ML and BI analyses (88% ML, 100 PP; Fig. 2). A pairwise nucleotide comparison showed that P. yunnanensis differs from P. vagabunda (CBS 109875, holotype) in 3/836 bp of LSU (0.4%, without gaps) and 8/510 bp of ITS (1.6%, without gaps). The sexual morph of P. vagabunda (a synonym of Pezicula alba E.J. Guthrie) appears to have been collected only once[12,13,15]. Phlyctema yunnanensis differs from P. vagabunda in apothecia size (360–440 μm diam × 130–180 μm high vs. 1 mm diam), asci (65–90 × 8.5–13 μm, 4-spored, clavate, with a rounded apex vs. 125–150 × 13–24, 8-spored, cylindrical-clavate), and ascospores (13–19 × 6–9.5 μm, subglobose to ellipsoid or oblong to obovoide, aseptate vs. 20–30 × 7–10 μm, elongated ellipsoid, 3–5-septate)[4,12]. Therefore, based on the guideline for a polyphasic approach recommended for species boundary delimitation[42,43], P. yunnanensis is introduced as a novel taxon.
-
The phylogenetic analysis presented in this study confirms the placement of Phlyctema within the Dermateaceae, providing robust molecular evidence for the classification of P. yunnanensis (Figs 1, 2). The data indicate that P. yunnanensis can be differentiated from other Phlyctema species based on both phylogenetic and morphological analyses. Similar to the concatenated ITS-LSU-rpb2-tub2 dataset phylogeny, the phylogenetic analysis of the ITS-LSU data also placed the sequences of P. yunnanensis into a supported subclade in both methods. To date, the only known sexual morph within Phlyctema is that of the type species, P. vagabunda[2,12,15]. Notably, P. yunnanensis exhibits morphological differences from P. vagabunda, particularly in its asci and ascospores. However, it is important to highlight that the type material of P. vagabunda is in relatively poor condition, which may limit the accuracy of morphological comparisons[12]. The asexual form of P. yunnanensis remains unidentified and warrants further investigation. In contrast, the asexual morph of P. vagabunda is well-documented as a pathogen responsible for bull's eye rot in apples and pears, with a prevalence reported in Australia, the USA, and Chile[44,45]. It is also implicated in causing coin canker on ash trees[46,47], and fruit spots on olives[48]. Additionally, P. vagabunda has been identified as a saprobe on various woody and herbaceous plants[12].
The phylogenetic analyses reveals that Phlyctema coronillae clusters within Phlyctema phoenicis (Figs 1, 2). Phlyctema phoenicis was introduced by Crous et al.[49] from Phoenix canariensis (Arecaceae) in New Zealand whereas P. coronillae was described by Li et al.[4] from dead aerial branches of Coronilla emerus (Fabaceae) in Italy. However, Li et al.[4] did not include the holotype of Phlyctema phoenicis in their phylogenetic analyses. Later, Crous et al.[17] re-analyzed the phylogeny of Phlyctema and included both Phlyctema phoenicis and P. coronillae. Despite their co-occurrence in the same clade, no explanation was provided to distinguish between these two species. To clarify their relationship, a comparative study was conducted based on molecular and morphological data. Pairwise nucleotide comparison shows that Phlyctema phoenicis (CPC 29372) and P. coronillae (MFLU 15-1243) differ by only 2/548 bp (0.36%, with 1 gap) in ITS and show no variation 0/827 bp in LSU. These minimal differences indicate no significant molecular distinction between the two taxa. Morphologically, Phlyctema phoenicis and P. coronillae are also similar[4,49]. Based on the combined molecular and morphological evidence, we propose that Phlyctema coronillae is a synonym of P. phoenicis. The present analyses also confirm that Neofabraea, Phlyctema, and Pseudofabraea form distinct, well-supported clades in the phylogenetic trees (Figs 1, 2). Previously, species within these genera were grouped under Neofabraea. However, Chen et al.[2] demonstrated significant morphological variation among species, leading to the recognition of these three as separate genera. The present phylogenetic results support this distinction, with Neofabraea, Phlyctema, and Pseudofabraea forming independent lineages.
In this study, the phylogenetic tree of Dermateaceae was updated based on the most recent literature and the available sequence data, including Iliushin & Kirtsideli[6], who introduced a new genus, Pseudoxenochalara. Specifically, the phylogenetic analysis of Phlyctema was expanded by incorporating findings from Li et al.[4] and Crous et al.[17]. The updated tree includes Neofabraea, Phlyctema, and Pseudofabraea, which formed three distinct clades (Figs 1, 2). This separation reflects the morphological and genetic diversity among these genera, previously grouped under Neofabraea[2]. The present study highlights some differences in the results derived from analyses of the ITS and LSU dataset compared to the multigene sequence dataset (ITS-LSU-rpb2-tub2). The multigene phylogenetic analysis provided better resolution, successfully delineating Neofabraea, Phlyctema, and Pseudofabraea as separate genera. In contrast, the ITS and LSU datasets alone resulted in unresolved clades for some species, including Phlyctema vincetoxici, which formed a distinct clade with Neofabraea krawtzewii and Pseudofabraea citricarpa, which clustered within the Phlyctema clade (Fig. 1). These results emphasize that ITS and LSU markers alone are insufficient for accurately distinguishing these genera due to limited phylogenetic resolution, necessitating the use of multigene datasets for robust analysis. The updated phylogenetic tree also provides new insights into the relationships among genera within the Dermateaceae. Many genera included in this study show unresolved clades in ITS and LSU-based analyses. This pattern reflects inherent limitations in single-locus analyses and reinforces the importance of multigene phylogenies. By presenting ITS and LSU phylogenetic trees alongside multigene trees, we validate that our species identification results are consistent across methods while demonstrating the added resolution provided by the latter approach.
The present study marks the first report of the sexual morph of Phlyctema yunnanensis on herbaceous plants from grasslands in China. The present findings enhance the taxonomic diversity within the Phlyctema genus and contribute to the broader understanding of fungal diversity in this group. Furthermore, several recent studies[24,25,50] have documented new species from Yunnan grasslands, suggesting a wealth of undiscovered species remains in this region, highlighting the need for continued exploration of fungal diversity.
The authors thank the Yunnan Department of Sciences and Technology of China (Grant Nos 202302AE090023 and 202303AP140001). D.N. Wanasinghe is funded by the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Kingdom of Saudi Arabia.
-
The authors confirm their contribution to the paper as follows: study conception and design, data collection, draft manuscript preparation: Gao Y; analysis and interpretation of results: Gao Y, Zhong T, Wanasinghe DN; review and editing: Wanasinghe DN, Eungwanichayapant PD, Jayawardena RS, Hyde KD, Gui H. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated for this study are available in NCBI, Index Fungorum, and Facesoffungi databases.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Gao Y, Zhong T, Wanasinghe DN, Eungwanichayapant PD, Jayawardena RS, et al. 2024. Phlyctema yunnanensis (Dermateaceae, Helotiales), a novel species from herbaceous plants in grassland ecosystems of Yunnan, China. Studies in Fungi 9: e019 doi: 10.48130/sif-0024-0020 

Phlyctema yunnanensis (Dermateaceae, Helotiales), a novel species from herbaceous plants in grassland ecosystems of Yunnan, China
- Received: 25 November 2024
- Revised: 23 December 2024
- Accepted: 24 December 2024
- Published online: 30 December 2024
Abstract: During a survey of grassland microfungi in Yunnan Province, China, a fungus with distinctive apothecia was collected. Initial BLAST analyses of its sequence data indicated a close affiliation with taxa in the Dermateaceae. Phylogenetic analyses based on a combined dataset of the internal transcribed spacer (ITS) and large subunit (LSU) of ribosomal DNA, RNA polymerase II second largest subunit (rpb2), and β-tubulin (tub2) genes confirmed its placement within the genus Phlyctema (Dermateaceae). The species is morphologically characterized by discoid, sessile apothecia, bitunicate, fissitunicate, 4-spored clavate asci, and subglobose to ellipsoid or oblong to obovoid, slightly inequilateral, hyaline, aseptate, and thin-walled ascospores. Based on both morphological features and multigene analyses, this fungus is introduced as a new species, Phlyctema yunnanensis.
-
Key words:
- Leotiomycetes /
- molecular phylogeny /
- new taxon /
- sexual morph /
- taxonomy