








-
Absidia was introduced by van Tieghem[1], with A. reflexa as the type species. Preliminarily, Absidia was accommodated in Absidiaceae as the type genus[2]. However, with DNA sequence techniques used in species identification, Absidia always formed a well-supported monophyletic clade with genera in Cunninghamellaceae (viz., Chlamydoabsidia, Cunninghamella, Gongronella, Hesseltinella, and Halteromyces)[3−7]. Hence, Absidia was hitherto classified in Cunninghamellaceae[8] whereas Absidiaceae was treated as a synonym of the highly polyphyletic family Mucoraceae by Kirk et al.[7]. Although, Hoffmann[9] highly recommended to resurrect the monophyletic Absidiaceae in accordance with Voigt et al.[10].
Since the genus Absidia was introduced[1], many different allied genera were subsequently established to accommodate absidia-like species based solely on morphological characteristics[11−16]. Considering the morphological distinctiveness, it may be challenging among the genera Absidia, Gongronella, Lichtheimia, Mycocladus, Phycomyces, Proabsidia, Protoabsidia, Pseudoabsidia, Rhizopus, and Tieghemella, because these genera are highly similar in morphology[17]. Nevertheless, Hesseltine & Ellis[17] accepted only Absidia and Gongronella as distinct genera. However, a morpho-molecular approach coupled with physiology significantly segregated these genera into different families. Among them, only Absidia (synonyms: Proabsidia, Pseudoabsidia, and Tieghemella), and Gongronella were classified in Cunninghamellaceae whereas Lichtheimia (synonym: Protoabsidia) was classified in Lichtheimiaceae, Mycocladus in Mycocladaceae, Phycomyces in Phycomycetaceae, and Rhizopus in Rhizopodaceae[18].
A comprehensive study based on morphology, phylogeny, and physiology provided reliable species delineation of Absidia[19]. Absidia species were segregated into three groups viz. mesophilic species (optimal growth temperature 25–34 °C), mycoparasitic species on mucoralean fungi (optimal growth temperature 14–25 °C) and thermotolerant species (optimal growth temperature 37–45 °C)[19]. Consequently, the new genus Lentamyces was established to accommodate the mycoparasitic species[20], whereas the thermotolerant species were transferred to the genus Lichtheimia[21]. To date, only the mesophilic species (optimal growth temperature 25–34 °C) were accepted as Absidia sensu stricto[9,20,21]. Currently, species identification and classification for the genus Absidia are mainly based on morphological characteristics coupled with phylogeny and physiology (e.g., growth temperature)[9,19,20,22−26]. Based on evolutionary relationships, Zhao et al.[25] updated the classification framework of the phylum Mucoromycota and calibrated the taxonomic ranks with estimated divergence times using molecular dating. Among these, the estimated divergence time of the phylum Mucoromycota was earlier than 617 Mya, the classes and orders earlier than 547 Mya, the families earlier than 199 Mya, and the genera earlier than 12 Mya. According to Zhao et al.[25], the divergence time of Absidia was estimated to be 327 Mya for the stem age and 135 Mya for the crown age, based on a time-scaled Bayesian maximum clade credibility phylogenomic tree inferred from ITS and LSU rDNA sequences[25].
Absidia is ubiquitous and distributed in a wide range of ecological niches worldwide, including air, animal dung, food, insect remains, leaf litter, and soil[24]. Notably, soil accounts for more than 50% of Absidia species[24]. China has been recognized as a region of high diversity for Absidia species, contributing ~20% to the origin of taxonomic types in this genus[24,27]. This genus is typically characterized by sporangiophores with columellae bearing one to several projections, rhizoids forming between the sporangiophores but never opposite them, and the development of pyriform, deliquescent-walled and apophysate sporangia[1,9,19,20,24,25,28−31]. Some species within Absidia such as A. biappendiculata, A. chinensis, A. cinerea, and A. jiangxiensis, produce zygospores within zygosporangia, with their opposite suspensory cells possessing appendages[9,19,25].
The secondary metabolites of Absidia have been relatively well-studied, particularly for their industrial implications[25,32−36]. For instance, A. coerulea is capable of bio-transforming saponins, and regioselectivity to 20(S)-protopanaxatriol. Also, this species has been used in bio-transformation of the (−)-α-bisabolol, and in the hydroxylation of dehydroepiandrosterone (DHEA) to 7α-hydroxy-5-androstene-17-one (7α-OH-DHEA) and 7β-hydroxy-5-androstene-17-one (7β-OH-DHEA). These processes provide efficient pathways to enhance the yield of saponins, (−)-α-Bisabolol and 7β-OH-DHEA for the cosmetic and pharmaceutical industries[34−37]. Furthermore, A. coerulea and A. glauca have been used in the bio-transforming of flavones (chrysin, apigenin, luteolin, and diosmetin), flavanones (pinocembrin, naringenin, eriodictyol, and hesperetin) and 3-Oxo-Oleanolic acid into hydroxylated metabolites. These two species are also efficient producers of chitosan, which is widely used in food processing and antimicrobial products, as well as in the bio-transforming of steroid products[33,34]. Absidia cylindrospora has demonstrated the ability to act as a biosorption for cadmium, copper, and lead under controlled experimental conditions[34,38]. Additionally, A. cylindrospora and A. fusca have been used in bioremediation processes due to their capacity to degrade polycyclic aromatic compounds, such as hydrocarbons[34,39]. Moreover, during the bio-transformation of 20(R)-panaxadiol by A. corymbifera, six oxidized and hydroxylated products reported were identified as novel compounds[32]. Other Absidia species have also been explored for enzyme engineering and metabolomics in industrial contexts. For instance, α-galactosidase from A. reflexa has been used to produce rubusoside derivatives, A. repens has been utilized to yield chitosan and laccase from A. spinosa has been applied for the biotransformation of cresol red[25,40]. Given these findings, Absidia is a promising resource for applications in food processing, antimicrobial production, and biotransformation. Besides, new secondary metabolites are frequently discovered during microbial transformation involving Absidia species.
During a survey to collect microfungi associated with avocado in Yunnan Province, China, two isolates were obtained, one from the soil in an avocado plantation and another as a root endophyte of an avocado tree. Based on morphological, phylogenetic, and physiological analyses, these isolates were determined to be conspecific and represent a novel species of Absidia.
-
The two fungal strains were isolated from soil samples from an avocado plantation and fresh healthy roots of avocado trees, both of which were collected from Menglian County, Yunnan Province, China in September, 2023. Soil samples were collected from 10 cm below ground level, near the shallow roots of avocado trees using a sterilized auger boring tool (model XDB, China, total length 110 cm, drill outer diameter 57 mm, inner diameter 51 mm, single sampling length 20 cm). A portion of the soil (1 g) was mixed with 9 mL of sterile distilled H2O. Serial dilutions of the mixtures were prepared from 10−1 to 10−4. For each dilution, a 100 μL aliquot was plated onto potato dextrose agar (PDA; Qingdao Daily Water Biotechnology Co. Ltd. Shandong, China) with antibiotics (streptomycin sulfate, 100 mg/L, and ampicillin, 100 mg/L). The plates were incubated at 25–30 °C. The PDA plate was examined daily with an Olympus stereo-microscope series MoDELSZ2-ILST. The presence of colonies was transferred to new PDA plates. The isolation and induction of root endophytes from the root tissue of avocado tree were carried out following the guidelines as described in Senanayake et al.[41] and Mattoo & Nonzom[42]. Root samples were cleaned with tap water, and each root was cut into several small pieces (5 mm long). The root pieces were surface sterilized using 2% sodium hypochlorite (NaOCl) for 1 min, followed by three repeated washings with sterilized water. Furthermore, surface sterilization was carried out via washing with 75% ethanol for 30 s and three series of washing with sterile distilled water, and finally it was allowed to dry under a laminar flow cabinet. Then, the edges of the root pieces were trimmed, transferred onto plates, and incubated at 25–30 °C for 2–5 d. When individual hyphal tips grew out from root pieces, the mycelia were picked using sterilized needles, placed onto new PDA plates, and incubated at 25–30 °C under 12 h alternating light and dark conditions for one week to one month. After one month, the sporulation was observed. Dry culture materials were deposited in the herbarium of the Cryptogams Kunming Institute of Botany Academia Sinica (KUN-HKAS). The living cultures were conserved at the Kunming Institute of Botany Culture Collection (KUNCC). Index Fungorum number is provided for the newly described taxon.
Microscopic examination and growth temperature
-
Micromorphological features (sporangia, sporangiophores, sporangiospores, and columellae, etc.) were examined and captured by a Nikon compound microscope (model ECLIPSE Ni-U, Japan) connected with a Nikon DS-Ri2 camera using DIC microscopy. Measurements of these morphological features were made in Tarosoft® Image Frame Work software version 0.9.7. Photographic plates were edited and combined in Adobe Photoshop CS6 software (Adobe Systems Inc., USA). The determination of maximum growth temperature followed the method of Zhao et al.[24]. Four fungal blocks (5 mm × 5 mm) were selected from a one-week-old colony growing on PDA and placed separately onto four new PDA plates, representing four replicates. These replicates were incubated at 30 °C for 2 d. Subsequently, the temperature was gradually increased in 2 °C increments (32, 34, and 36 °C), with incubation at each temperature for 2 d, until the colonies ceased to grow. When colony growth stopped (at 36 °C), the maximum growth temperature was confirmed by observing the mycelium growth at a reduced temperature (35 °C) at which the colonies also failed to grow.
DNA extraction, PCR amplification, and sequencing
-
The genomic DNA was extracted from fresh mycelia grown on PDA medium for two weeks using the Biospin Fungus Genomic DNA Extraction Kit (BioFlux®, Hangzhou, China) following the manufacturer's instructions. Partial DNA sequences of the internal transcribed spacers (ITS1-5.8S-ITS2) and the 28S large subunit rDNA (LSU) were amplified by primers ITS5 and ITS4[43], and LR0R and LR5[44], respectively. The component of the PCR reaction mixture was performed in a 25 μl total volume containing 2 μl of DNA template, 1 μl of each forward and reverse primers (10 μM), 12.5 μl of 2× Power Taq PCR Master Mix (mixture of EasyTaqTM DNA Polymerase, dNTPs, and optimized buffer, Beijing Bio Teke Corporation (Bio Teke), China) and 8.5 μl of ddH2O (double-distilled water). The thermal cycling of PCR amplification for ITS and LSU followed the method of Xu et al.[45] but the annealing temperature was adjusted to 52 °C. PCR products were sent to TsingKe Biological Technology (Beijing) Co., Ltd, China for purification and sequencing. Newly generated consensus sequences were deposited in GenBank under accession numbers provided in Table 1.
Table 1. Taxa names, strain numbers, and GenBank accession numbers of taxa used for the present phylogenetic analyses.
Species names Strain numbers GenBank accession numbers ITS LSU Absidia abundans XY09265 ON074697 ON074681 Absidia abundans CGMCC 3.16255 NR_182590 ON074683 Absidia abundans XY09274 ON074696 ON074682 Absidia aguabelensis URM 8213 NR_189383 NG_241934 Absidia alpina CGMCC 3.16104 OL678133 / Absidia ampullacea CGMCC 3.16054 NR_191130 NG_242502 Absidia anomala CBS 125.68 NR_103626 NG_058562 Absidia biappendiculata CBS 187.64 NR_190243 MZ350147 Absidia bonitoensis URM 7889 MN977786 MN977805 Absidia brunnea CGMCC 3.16055 NR_191131 NG_242503 Absidia caatinguensis URM 7156 NR_154704 NG_058582 Absidia caerulea CBS 101.36 MH855718 MH867230 Absidia caerulea XY00608 OL620081 / Absidia californica CBS 126.68 NR_077169 NG_056998 Absidia californica FSU4748 AY944873 EU736301 Absidia cheongyangensis CNUFC CY2203 PP844904 PP852788 Absidia chinensis CGMCC 3.16057 MZ354141 MZ350135 Absidia chinensis CGMCC 3.16056 NR_191132 NG_242504 Absidia cinerea CGMCC 3.16062 NR_191134 NG_242506 Absidia coerulea FSU767 AY944870 / Absidia coerulea XY00729 OL620082 / Absidia cornuta URM 6100 NR_172976 MN625255 Absidia crystalloides CGMCC3.27496 PP377803 PP373736 Absidia crystalloides SAUCC693201 PP377804 PP373737 Absidia cuneospora CBS 101.59 MH857828 NG_058559 Absidia cylindrospora CBS 153.63 / NG_058563 Absidia cylindrospora XY00313 ON123744 ON123765 Absidia cylindrospora var. cylindrospora CBS 100.08 JN205822 JN206588 Absidia digitula CGMCC 3.16058 MZ354142 MZ350136 Absidia edaphica MFLU 20-0416 NR_172305 NG_075367 Absidia fluvii CNUFC CY2240 PP844891 PP852703 Absidia frigida CGMCC 3.16201 NR_182565 NG_149032 Absidia fusca CBS 102.35 NR_103625 NG_058552 Absidia gemella CGMCC 3.16202 OM108488 NG_149033 Absidia glauca CBS 129233 MH865253 MH876693 Absidia glauca CBS 101.08 NR_111658 NG_058550 Absidia glauca FSU660 AY944879 EU736302 Absidia globospora CGMCC 3.16031 NR_189829 NG_241930 Absidia globospora CGMCC 3.16035 MW671538 MW671545 Absidia globospora CGMCC 3.16036 MW671539 MW671546 Absidia healeyae UoMAU1 MT436028 MT436027 Absidia heterospora SHTH021 JN942683 JN982936 Absidia jiangxiensis CGMCC 3.16105 OL678134 / Absidia jiangxiensis [as jindoensis] XY000810 ON123748 ON123770 Absidia jindoensis CNUFC-PTI1-1 MF926622 MF926616 Absidia koreana EML-IFS45-1 KR030062 KR030056 Absidia kunryangriensis CNUFC CY2230 PP844905 PP956882 Absidia lobata CGMCC 3.16256 ON074690 ON074679 Absidia longissima CGMCC 3.16203 NR_182566 OM030225 Absidia macrospora FSU4746 AY944882 EU736303 Absidia medulla CGMCC 3.16034 NR_189832 NG_241932 Absidia menglianensis KUNCC 24-18541 PQ594927 PQ594929 Absidia menglianensis KUNCC 24-18542 PQ594928 PQ594930 Absidia microsporangia KUNF-22-121A OQ868364 OQ868366 Absidia microsporangia KUNF-22-126A OQ868365 OQ868367 Absidia microsporangia KUNF-22-316 PP264375 PP264469 Absidia montepascoalis URM 8218 MW473494 MW561560 Absidia multispora URM 8210 MN953780 MN953782 Absidia nigra CGMCC 3.16060 MZ354144 MZ350138 Absidia nigra CGMCC 3.16059 MZ354143 MZ350137 Absidia nigra CBS 127.68 NR_173068 NG_058560 Absidia oblongispora CGMCC 3.16061 NR_191133 NG_242505 Absidia ovalispora CGMCC 3.16019 NR_176748 MW264131 Absidia pacifica CGMCC3.27497 PP377802 PP373735 Absidia pacifica SAUCC413601 PP377801 PP373734 Absidia panacisoli SYPF 7183 NR_159563 NG_063948 Absidia paracylindrospora CNUFC L2207 PP844907 PP956883 Absidia pararepens CCF 6352 MT193669 NG_242483 Absidia pateriformis CGMCC3.27495 PP377805 PP373738 Absidia pateriformis SAUCC634702 PP377806 PP373739 Absidia pernambucoensis URM 7219 MN635568 MN635569 Absidia pseudocylindrospora CBS 100.62 NR_145276 NG_058561 Absidia psychrophilia FSU4745 AY944874 EU736306 Absidia psychrophilia CBS 128.68 NR_154667 NG_058544 Absidia purpurea CGMCC 3.16106 NR_191153 / Absidia radiata CGMCC 3.16257 ON074698 ON074684 Absidia radiata XY09330 ON074699 ON074685 Absidia repens CBS 115583 NR_103624 NG_058551 Absidia saloaensis URM 8209 MN953781 MN953783 Absidia sichuanensis CGMCC 3.16258 NR_182589 ON074688 Absidia soli MFLU 20-0414 NR_172306 NG_075368 Absidia sp. [as pseudocylindrospora] EML-FSDY6-2 KU923817 KU923814 Absidia spinosa FSU551 AY944887 EU736307 Absidia stercoraria EML-DG8-1 KU168828 KT921998 Absidia sympodialis CGMCC 3.16063 MZ354147 MZ350141 Absidia sympodialis CGMCC 3.16064 NR_191135 NG_242507 Absidia tarda URM 8412 PP844911 PP956884 Absidia terrestris FMR 14989 LT795003 LT795005 Absidia thailandica MFLUCC 23-0073 OR606547 OR606546 Absidia turgida CGMCC 3.16032 NR_189830 NG_241931 Absidia varians CGMCC 3.16065 NR_191136 NG_242508 Absidia variiprojecta URM 8620 PP844913 PP956885 Absidia variispora URM 8720 PP844915 PP956886 Absidia virescens CGMCC 3.16066 MZ354150 MZ350144 Absidia virescens CGMCC 3.16067 NR_191137 NG_242509 Absidia xinjiangensis CGMCC 3.16107 OL678136 / Absidia yunnanensis XY09528 ON074701 ON074686 Absidia yunnanensis CGMCC 3.16259 NR_182591 NG_149054 Absidia zonata CGMCC 3.16033 NR_189831 NG_242490 Absidia zygospora MFLUCC 23-0061 OR104965 OR104992 Cunninghamella blakesleeana CBS 782.68 JN205869 MH870950 Cunninghamella elegans CBS 167.53 MH857146 HM849700 The ex-type strains are in bold and newly sequences in the present study are indicated in blue. Abbreviations: CBS: Westerdijk Fungal Biodiversity Institute, Utrecht, Netherlands; CCF: Culture Collection of Fungi, Charles University in Prague; CGMCC: China General Microbiological Culture Collection Center, Beijing, China; CNUFC: Chonnam National University Fungarium, Gwangju, South Korea; FMR: Facultad de Medicina, Reus, Tarragona, Spain; KUNCC: Kunming Institute of Botany Culture Collection, Kunming, China; MFLU: Herbarium of Mae Fah Luang University, Chiang Rai, Thailand; MFLUCC: Mae Fah Luang University Culture Collection, Chiang Rai, Thailand; NRRL: USDA-Agricultural Research Service Culture Collection, US; SAUCC: Shandong Agricultural University Culture Collection (SAUCC), China; URM: University Recife Mycology Culture Collection of the Universidade Federal de Pernambuco, Brazil. Sequence alignment and phylogenetic analyses
-
The nucleotide BLAST search (
https://blast.ncbi.nlm.nih.gov/Blast.cgi , accessed on 10 August 2024) was applied to discover taxa closely related to the new isolates (KUNCC 24-18541 and KUNCC 24-18542). Similarity indices from the BLAST search indicated that KUNCC 24-18541 and KUNCC 24-18542 are conspecific and belong to Absidia (Cunninghamellaceae). Therefore, to reveal accurate phylogenetic placements of the new isolates, a concatenated ITS-LSU phylogeny of Absidia were analyzed based on maximum-likelihood and Bayesian inference criteria. Sequence datasets of representative taxa in Absidia and Cunninghamella (Table 1) were retrieved from recent publications[22−26,29−31], downloaded from GenBank (www.ncbi.nlm.nih.gov/nucleotide ; accessed on 29 November 2024). Preliminarily individual DNA sequence matrixes were aligned via the online platform, MAFFT v. 7.511[46]. The ambiguous sites of the aligned sequence datasets were further trimmed and improved where necessary using BioEdit v. 6.0.7[47]. The complementation of the missing nucleotide bases at the start and end of the consensus sequence was trimmed. Individual gene datasets were prior analyzed by maximum likelihood to compare the congruence of tree topologies and further concatenated ITS-LSU sequence matrix was analyzed.Maximum-likelihood (ML) analysis was performed via the online portal CIPRES Science Gateway v. 3.3[48], with RAxML-HPC v.8 on XSEDE (8.2.12) tool, using default settings but following the adjustments: the GAMMA nucleotide substitution model and 1000 rapid bootstrap replicates. The evolutionary model of nucleotide substitution for Bayesian inference (BI) analysis was selected independently for each locus using MrModeltest 2.3[49]. GTR + I + G was the best fit for ITS and LSU loci under the Akaike Information Criterion (AIC). BI analysis was performed via MrBayes v. 3.2.6[50]. Markov chain Monte Carlo sampling (MCMC) was used to determine posterior probabilities (PP)[51,52]. Six simultaneous Markov chains were run for 1,000,000 generations and trees were sampled every 100th generation. The 0.15 'temperature' value was set in the MCMC heated chain. All sampled topologies beneath the asymptote (20%) were discarded as part of a burn-in procedure and the remaining 8,000 trees were used for calculating posterior probabilities (PP) in the 50% majority rule consensus tree (when the standard deviation of split frequency is lower than 0.01).
The tree topologies generated in this study were visualized on FigTree v. 1.4.0[53]. The phylogram was edited and laid out using Microsoft Office PowerPoint 2016 (Microsoft Inc., Redmond, WA, USA) and converted to a tiff file in Adobe Photoshop CS6 software (Adobe Systems Inc., San Jose, CA, USA). The final alignment and phylogram were submitted to TreeBASE (submission ID: 31814,
www.treebase.org , accessed on 10 December 2024). -
The concatenated ITS-LSU sequence matrix comprised 102 strains of representative species in Absidia, with Cunninghamella blakesleeana (CBS 782.68) and C. elegans (CBS 167.53) as the outgroup taxa. The dataset consisted of 2,175 total characters, including gaps (ITS: 1–1,175 bp, LSU: 1,176–2,175 bp). The best-scoring ML tree was selected to represent the phylogenetic relationships of two new isolates with other representative taxa in Absidia (Fig. 1), with the final ML optimization likelihood value of –31,904.047878 (ln). All free model parameters were estimated by the RAxML GAMMA model of rate heterogeneity, with 1,431 distinct alignment patterns. Estimated base frequencies were as follows: A = 0.266550, C = 0.195987, G = 0.247413, T = 0.290050, with substitution rates AC = 1.040788, AG = 2.710746, AT = 1.633236, CG = 0.778485, CT = 5.127044, and GT = 1.000000. The gamma distribution shape parameter alpha = 0.336716 and the tree-length = 11.191212. The final average standard deviation of split frequencies at the end of the total MCMC generations was calculated as 0.009041 in BI analysis.
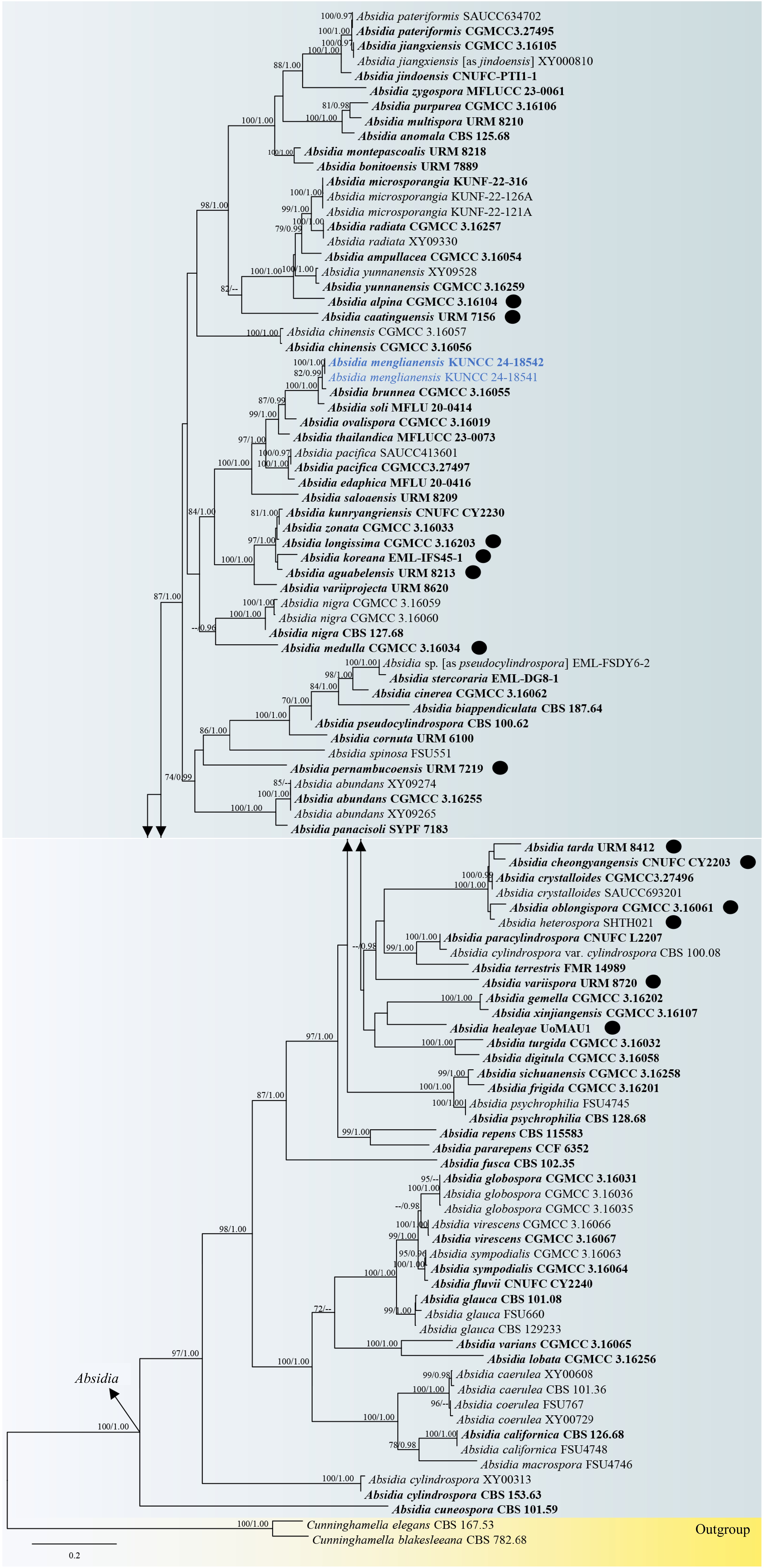
Figure 1.
Maximum likelihood phylogenetic tree generated by RAxML based on a concatenated ITS and LSU sequence data. The tree is rooted to Cunninghamella blakesleeana (CBS 782.68) and C. elegans (CBS 167.53). Maximum likelihood bootstrap values (MLBS) greater than 70% and Bayesian posterior probabilities (BYPP) greater than 0.95 are noted above the nodes as MLBS/BYPP. Ex-type strains are in bold and newly generated sequences are in blue.
Tree topologies generated based on ML and BI analyses were similar in the present study and the ML phylogenetic backbone tree is shown in Fig. 1. In the present study, phylogenetic affinities of most Absidia species were well-resolved with significant support in ML and BI analyses (up to 70% MLBS and 0.95 BYPP). However, A. aguabelensis, A. alpina, A. caatinguensis, A. cheongyangensis, A. healeyae, A. heterospora, A. longissimi, A. medulla, A. koreana, A. oblongispora, A. pernambucoensis, A. tarda, and A. variispora are not phylogenetically well-resolved in the present study (remarkable as black circle on the tree). This lack of resolution may be attributed to limited taxon sampling and ambiguous sites in the ITS alignments. Absidia caatinguensis formed a well-resolved branch with significant support in the ML analysis (82% MLBS) but lacked significant support in BI analysis. This species formed an independent branch basal to A. microsporangia, A. radiata, A. ampullacea, A. yunnanensis, and A. alpina, respectively. Absidia medulla and A. variispora formed well-supported branches in the BI analysis (0.96 BYPP and 0.98 BYPP, respectively), though both species showed weak support in the ML analysis. Absidia medulla formed an independent branch positioned as a sister to A. nigra. Conversely, A. variispora was basal to a subclade comprising A. tarda, A. cheongyangensis, A. crystalloides, A. oblongispora, A. heterospora, A. paracylindrospora, A. cylindrospora var. cylindrospora, and A. terrestris. Absidia tarda was identified as a sister to A. cheongyangensis, while A. oblongispora was sister to A. heterospora. These four species formed a weakly supported monophyletic subclade with A. crystalloides. Absidia aguabelensis formed a separated, weakly supported branch with A. koreana and clustered with A. longissimi, A. zonata, and A. kunryangriensis, supported by significant values (97% MLBS and 1.00 BYPP). Absidia healeyae was closely related to A. gemella and A. xinjiangensis, forming a basal branch clustered with these two species. Absidia pernambucoensis clustered as basal to a group comprising Absidia sp. [as pseudocylindrospora] strain EML-FSDY6-2, A. stercoraria, A. cinerea, A. biappendiculata, A. pseudocylindrospora, A. cornuta, and A. spinosa. The two new isolates (strains KUNCC 24-18541 and KUNCC 24-18542) formed a robust subclade (100% MLBS and 1.00 BYPP), and is sister to A. brunnea (82% MLBS and 0.99 BYPP), and A. soli (100% MLBS and 1.00 BYPP) within Absidia. Hence, Absidia menglianensis (strains KUNCC 24-18541 and KUNCC 24-18542) is introduced as a novel species.
Taxonomy
-
Absidia menglianensis H.B. Jiang, C.Y. Luo & Phookamsak, sp. nov.
Index Fungorum number: IF 902073; Fig. 2.
Etymology: The specific epithet 'menglianensis' refers to the location, Menglian, Yunnan Province of China, where the new species was collected.
Holotype: KUN-HKAS 144524.

Figure 2.
Absidia menglianensis (KUN-HKAS 144524, holotype). (a), (b) Culture from above and reverse; (c) Sporangiospores; (d) Sporangium; (e), (f) Sporangia with columellae (arrow); (g) Rhizoids; (h) Swelling on sporangiophores; (i), (j) Sporangiophores. Scale bars: (c), (f) = 10 μm, (d) = 15 μm, (e) = 7 μm, (g) = 20 μm, (h), (j) = 50 μm, (i) = 30 μm.
Endophytic on roots of avocado trees (Persea americana) and living in the soil of an avocado plantation. Hyphae hyaline at first, becoming brown when mature, 5–14.5 μm wide. Stolons branched, hyaline or light brown, smooth, with few septa near the base of sporangiophores, 3–8.5 μm wide. Rhizoids brown to dark brown, rarely branched. Sporangiophores 110–185 × 2.5–4.5 μm (
$\overline x $ $\overline x $ $\overline x $ $\overline x $ Maximum growth temperature: 36 °C.
Culture characteristics: Colony grows fast on PDA, reaching 30 mm in 3 d at room temperature (25–30 °C) under normal light conditions, colonies cottony to floccose, irregular, raised, pale brown to brown in the center, white at the margin. Mycelium superficial or immersed in agar medium, with brown to dark brown, branched, septate, smooth hyphae. The fungus can grow at room temperature, 32, 34, and 35 °C and cannot be grown at 36 °C (Fig. 3).
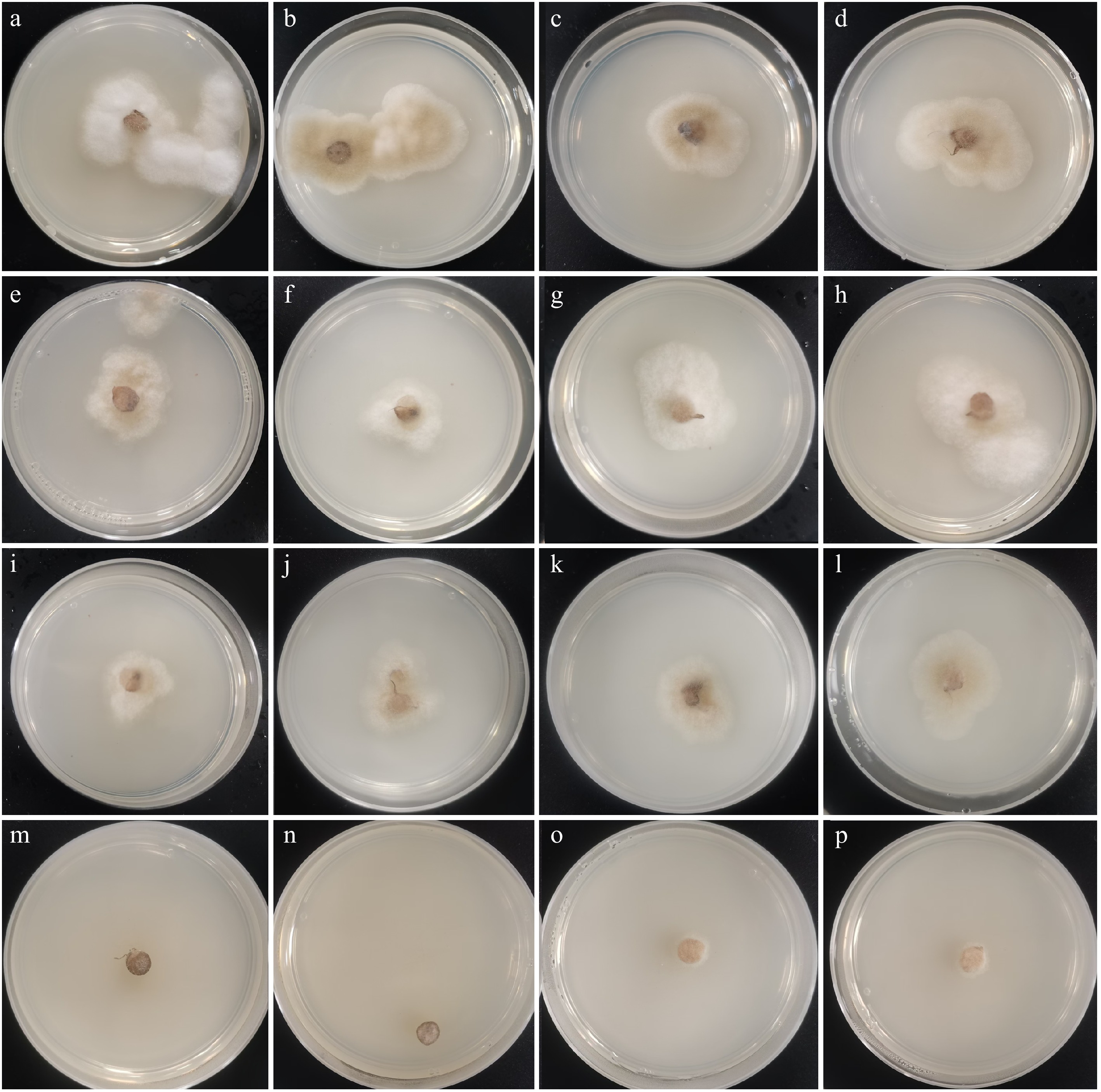
Figure 3.
Mycelial growth of Absidia menglianensis in PDA at different temperatures (32, 34, 35, 36 °C) after 3 d. (a)–(d) Colonies at 32 °C; (e)–(h) Colonies at 34 °C; (i)–(l) Colonies at 35 °C; (m)–(p) Colonies at 36 °C.
Material examined: China, Yunnan Province, Pu'er Prefecture, Menglian Dai, Lahu, and Va Autonomous County, Mangxin Town, isolated from soil in the avocado plantation (22°10′34″ N, 99°33′42″ E, altitude 1,078.6 m), 15 September 2023, H.B. Jiang, dried herbarium culture, MLAS011 (KUN-HKAS 144524, holotype), ex-holotype living culture, KUNCC 24-18542.
Other materials: China, Yunnan Province, Pu'er Prefecture, Menglian Dai, Lahu, and Va Autonomous County, Mangxin Town, associated with the root of avocado trees (22°10′34″ N, 99°33′42″ E, altitude 1,078.6 m), 15 September 2023, H.B. Jiang, root 2-17 (KUN-HKAS 144523); living culture, KUNCC 24-18541.
Notes: Based on NCBI nucleotide blast search of ITS region, Absidia menglianensis (strains KUNCC 24-18541 and KUNCC 24-18542) showed the closest similarity with A. brunnea strain CGMCC 3.16055 with 98.17% similarity (Identities = 536/546 bp, with two gaps), and A. soli strain MFLU 20-0413 (96.45% similarity, Identities = 380/394 bp, with five gaps) and the type strain of A. soli (MFLUCC 20-0089) with 95.50% similarity (Identities = 467/489 bp, with six gaps). Absidia menglianensis has a close relationship with A. brunnea and A. soli based on morphology and phylogeny. These three species are morphological resemblance in forming monopodial, erect sporangiophores with a septum below apophyses, brown, globose sporangia at the maturity, sometimes with a swelling beneath sporangia, funnel-shaped apophyses, hyaline columellae bearing collarette and apical projection, and hyaline sporangiospores (this study)[25,54]. However, A. menglianensis can be distinguished from A. brunnea and A. soli in size of sporangia, range of the septum below the apophyses, sporangiospore shape, and maximum growth temperature. Absidia brunnea, A. menglianensis, and A. soli presented sister relationships in the concatenated ITS-LSU phylogeny and can be distinct with high statistically supported values (Fig. 1). Compared to the size of sporangia, A. brunnea (17.5–38 × 19–34.5 μm)[25] and A. soli (16–51 × 15–45.5 μm)[54] have larger sporangia than A. menglianensis (13–25 × 10–22 μm). The septum below the apophysis presents different measurement ranges among A. brunnea (11–17 μm)[25], A. menglianensis (16–21.5 μm), and A. soli (21.5–37.5 μm)[54]. Besides, A. menglianensis and A. soli formed only cylindrical to ovoid sporangiospores[54], while A. brunnea formed ovoid sporangiospores with vacuoles[25]. Moreover, from physiology, the maximum growth temperature of A. menglianensis (36 °C on PDA) differs from A. brunnea (35 °C on MEA)[25] and A. soli (37 °C on YMA and PDA)[54]. Although they are slight differences in maximum growth temperature (1–2 °C), the limitation of maximum growth temperature range is a significant feature in delineating species in this fungal group. Furthermore, the nucleotide pairwise comparison of the ITS region demonstrated that A. menglianensis (strains KUNCC 24-18541 and KUNCC 24-18542) differs from A. brunnea (CGMCC 3.16055, ex-type strain) in 9/523 bp (1.72%) and differs from A. soli (MFLUCC 20-0089, ex-type strain) in 22/489 bp (4.5%). Hence, the species is justified as a new species.
-
Avocado (Persea americana) is one of the important economic crops in the Yunnan Province of China. However, there is relatively little knowledge about fungi associated with this plant as well as soil fungi in its plantation. Therefore, the present study attempted to explore the fungal diversity on avocado as well as soil fungi in the land-use for avocado plantations. In the present study, the new species Absidia menglianensis was found as a root endophyte as well as a soil fungus in an avocado plantation, which confirmed that Absidia is ubiquitous in a wide range of hosts and habitats in various ecological niches. In accordance with soil fungi that are usually recognized as saprobes in nature; however, many soil fungi are also known as soil borne pathogens, taking part in regulating soil health and managing interactions between soil and economic crops[54−56]. Interestingly, A. menglianensis can act either as a root endophyte or a soil fungus. This demonstrates the possibility of horizontal propagation of A. menglianensis. Unfortunately, a neighboring avocado plantation is facing root rot disease, where the causal agent was identified as Phytophthora sp. Whether A. menglianensis as a basal fungus joins the pathogenic process of root rot disease on avocado or as an endophyte helps the resistance to Phytophthora sp., the correlation between the two basal fungi and how they influence the pathogenic process of root rot disease in avocado plantations requires further study.
Studies on fungi associated with avocado plants are underrepresented in China, with only 15 fungal species reported from avocados in this region[27]. In the present study, a new species Absidia menglianensis is introduced, thereby enriching the number of fungi associated with the avocado. This is also the first report of an Absidia species associated with the avocado in China, with implications for expanding this knowledge globally. This novel discovery contributes significantly to fungal research on economic plants, such as the avocado, by providing insights into the diversity and polyphasic taxonomy of fungi associated with this host plant. Furthermore, it offers potential strategies for crop management aimed at reducing avocado diseases caused by fungi. It is likely that more fungal species will be discovered in avocado plantations in China as further sampling and investigations are conducted.
The present study enriches the diversity of Absidia species that improves an accurate species number of the genus within the country and globally. Currently, about 33 Absidia species have been reported in China including the novel species introduced herein[25,57]. Consequently, China could be considered as a hotspot for species richness of Absidia, which currently accounts for more than 1/3 of the species number of total Absidia globally[24,25,29−31]. In a large-scale geography of China, species of Absidia were mostly studied in western China, particularly Yunnan Province, which has been reported to contain 14 species (including A. menglianensis)[57].
Through the phylogenetic analyses in the present study, some strains of Absidia species that are available in GenBank, were misidentified. For instance, the unpublished strain XY000810 was identified as A. jindoensis, of which sequence data are currently only known on ITS and LSU in the GenBank database without other relevant information such as collection site, host information, morphological illustration, etc. However, the nucleotide blast search of the ITS region indicated that the strain is identical to A. jiangxiensis (CGMCC 3.16105), occurring with the phylogenetic results that strain XY000810 shared the same branch length with A. jiangxiensis (CGMCC 3.16105). Therefore, the strain XY000810 was identified as A. jiangxiensis herein temporarily based on the present phylogeny. Furthermore, A. pseudocylindrospora strain EML-FSDY6-2 formed a robust subclade (100% MLBS and 1.00 BYPP; Fig. 1) with A. stercoraria (EML-DG8-1) and was distant from type strain of A. pseudocylindrospora (CBS 100.62) (Fig. 1), of which the results are similar to those of Zhao et al.[25]. The nucleotide blast search of the ITS region indicated A. pseudocylindrospora strain EML-FSDY6-2 is closest in similarity to A. stercoraria (EML-DG8-1) with 98.97% similarity. Besides, a comparison between A. pseudocylindrospora strain EML-FSDY6-2 and ex-type strain CBS 100.62 demonstrated their morphological resemblances. However, they are different in size of columellae (9.5–19.5 μm vs 9–26 μm[58]). Hence, the A. pseudocylindrospora strain EML-FSDY6-2 was treated as Absidia sp. in the present study and the conspecific status of Absidia sp. strain EML-FSDY6-2 and A. stercoraria (EML-DG8-1) require further clarification and study. These findings should be taken into account for further resolving taxonomic ambiguities occurring in the genus Absidia, providing a better taxonomic resolution in the genus and extending to the higher taxonomic rank.
In the current state of species number in Absidia, there is confusion on the taxonomic classification and a variable of species number in different fungal repositories. Index Fungorum[57] lists 88 species of Absidia, nine varieties, and one form under A. ramosa var. ramosa, whereas Species Fungorum[59] accepted only 78 species of Absidia without any varieties or forms. In contrast, there are 83 species of Absidia and seven varieties accommodated in MycoBank[60]. Therefore, the accurate species number of Absidia requires further refinement and a solution to this problem needs to be found. Otherwise, there will be continuous confusion on the extant species in this fungal group for subsequent research. Among these, 73 Absidia species were clarified based on their molecular phylogeny.
We acknowledge the Biology Experimental Center, Germplasm Bank of Wild Species, Kunming Institute of Botany, Chinese Academy of Sciences for providing the facilities of their molecular laboratory. Hongbo Jiang appreciates the Postdoctoral Directional Training Foundation of Yunnan Province (Grant No. E33O38E261) under Yunnan Provincial Department of Human Resources and Social Security, Yunnan, China and the 1st batch of national (Chinese) postdoctoral overseas attraction program in 2023 from Ministry of Education of China (MOE). The authors thank Prof. Shaun Pennycook for his help in the nomenclature of the species. Rungtiwa Phookamsak sincerely acknowledges Introducing Talents Start-up Fund of Kunming Institute of Botany, Chinese Academy of Sciences, Yunnan Revitalization Talent Support Program 'Young Talent' Project (Grant No. YNWR-QNBJ-2020-120), Yunnan Revitalization Talent Support Program 'High-end Foreign Expert' Project, and Independent research of Department of Economic Plants and Biotechnology, Yunnan Key Laboratory for Wild Plant Resources, Kunming Institute of Botany, Chinese Academy of Sciences (Grant No. Y537731261), Jianchu Xu thanks Yunnan Department of Sciences and Technology of China (Grant No. 202302AE090023, 202303AP140001).
-
The authors confirm contribution to the paper as follows: conceptualization, data curation, and formal analysis: Luo C, Chen F, Jiang H, Phookamsak R; funding acquisition: Xu J, Jiang H; investigation, methodology, and writing − original draft: Luo C, Chen F, Jiang H; project administration: Jiang H, Phookamsak R; supervision: Xu J, Jiang H, Phookamsak R; writing − review and editing: Jiang H, Sun F, Phookamsak R. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated and analyzed during this study are available in this article. DNA sequence data are available in the GenBank database, and the accession numbers are provided in Table 1. Specimens and living cultures of new species have been deposited in a herbarium. The final alignment and phylogram were deposited in TreeBASE (submission ID: 31814). The introduced novel species, Absidia menglianensis, has been registered in Index Fungorum (Index Fungorum number: IF 902073).
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Luo C, Chen F, Phookamsak R, Sun F, Xu J, et al. 2024. Polyphasic taxonomic study of Absidia menglianensis sp. nov. (Cunninghamellaceae, Mucorales) isolated from an avocado plantation in Yunnan, China. Studies in Fungi 9: e018 doi: 10.48130/sif-0024-0018 

Polyphasic taxonomic study of Absidia menglianensis sp. nov. (Cunninghamellaceae, Mucorales) isolated from an avocado plantation in Yunnan, China
- Received: 15 November 2024
- Revised: 19 December 2024
- Accepted: 19 December 2024
- Published online: 27 December 2024
Abstract: During our ongoing studies of microfungi associated with avocado trees in Yunnan Province of China, the samples were collected from Menglian Dai, Lahu, and Va Autonomous County, Pu'er Prefecture. Two new strains were isolated from soil samples in the avocado plantation and roots of avocado trees (Persea americana), respectively. Through morphological observation and multigene phylogenetic analyses of concatenated ITS and LSU sequence data, these two new isolates are conspecific and introduced as a new species of Absidia, namely A. menglianensis. Absidia menglianensis is closely related to A. brunnea and A. soli based on morphology and phylogeny. However, the new species differs significantly from A. brunnea and A. soli in sizes of sporangia, range of the septum below the apophysis, and maximum growth temperature. They presented sister relationships in the concatenated ITS-LSU phylogeny and can be distinct with high statistical supported values. Detailed descriptions, illustrations, updated phylogeny, as well as determination of maximum growth temperature are provided.
-
Key words:
- Mucoromycetes /
- New species /
- Root endophyte /
- Soil fungi /
- Taxonomy