









-
Potassium is often the critical nutrient for plant growth and agricultural productivity[1,2]. In the plant body, potassium functions in abundant biochemical and physiological processes, e.g., enzyme activation, nutrient transport, signal transduction, regulation of the intracellular osmotic pressures, protein synthesis, and control of membrane polarity[3−6]. Adequate potassium supply is essential for maintaining plant physiological processes including photosynthesis, enzyme activation, osmotic regulation, and assimilate translocation, which ultimately determine crop yield and quality[7,8]. In conventional agricultural practices, water-soluble potassium fertilizers, primarily potassium chloride (KCl), potassium sulfate (K2SO4), and potassium dihydrogen phosphate (KH2PO4), are extensively applied to meet crop potassium requirements[9,10]. However, the overuse of these water-soluble potassium fertilizers frequently result in the loss of potassium fertilizers, eutrophication, and soil degradation[11−13]. To alleviate these problems, it is urgent to develop alternative potassium fertilizers for agricultural production.
Potassium feldspar is an aluminosilicate mineral with the common chemical structure of K2O·Al2O3·6SiO2, which contains high potassium contents with the percentage of K2O being 16.9%[14,15]. It is a non-water-soluble having the widest distribution and the largest reserves in the earth[16,17]. With the increasing global demand of potassium fertilizers, many researchers have been devoted to transform potassium feldspar to potassium fertilizers[18−20]. A series of strategies, e.g., hydrothermal treatment with CaO[21] , thermal treatment with the assistance of sodium chloride, and calcium chloride[22], NaOH-H2O digestion[23,24] , have been developed to release potassium from the feldspar mineral. However, these strategies often entail high costs due to the use of alkaline sources and thermal energy and may frequently result in the generation of secondary pollutants. Therefore, adopting economic and environmental-friendly strategies is required for the sustainable use of potassium feldspar as a raw material in fertilizer production.
Potassium-solubilizing microbes (KSM) are microorganisms capable of converting insoluble potassium in minerals into plant accessible forms[25,26]. These beneficial microorganisms not only solubilize mineral potassium but also promote plant growth through multiple mechanisms, particularly nutrient uptake enhancement and root system development[27−30]. Numerous KSMs, including Bacillus, Enterobacter, Priestia, Cladosporium, Aminobacter, and Sphingomonas, have been previously identified[25,31−33] . Recent studies have highlighted several key mechanisms by which potassium-solubilizing microbes mobilize mineral or insoluble potassium in soils. These mechanisms include the secretion of organic acids (such as citric acid, oxalic acid, and tartaric acid), which acidify the microenvironment to enhance potassium release, and the production of extracellular enzymes that facilitate mineral degradation[25,31−33]. Additionally, biofilm formation has been recognized as an adaptive strategy that promotes close contact between microbes and mineral surfaces, enabling more efficient dissolution. Some strains also express mineral-binding proteins, such as feldspar-binding proteins, which directly interact with potassium-bearing minerals to improve nutrient availability. These mechanisms demonstrate the diversity and significance of potassium-solubilizing microbes in nutrient cycling and soil fertility enhancement[34,35]. Notably, our recent study revealed that Priestia megaterium employs specific feldspar-binding proteins to enhance potassium solubilization[36]. Transcriptomic analyses have further demonstrated that potassium limitation triggers comprehensive metabolic remodeling and upregulation of biofilm-related genes in these bacteria, suggesting sophisticated regulatory networks for mineral weathering
Rhizosphere microorganism plays a critical role in facilitating potassium release, which is essential for plant growth[37,38]. To deeply investigate the composition of rhizosphere potassium-solubilizing bacteria and the mechanisms by which they respond to insoluble mineral potassium, highly efficient potassium-solubilizing bacteria were isolated and screened from the rhizosphere soil of wild Lespedeza floribunda plants. Subsequently, mRNA sequencing and transcriptional profiling analysis were conducted on the highly efficient potassium-solubilizing strain Cupriavidus taiwanensis NKK10, cultured in a medium with potassium feldspar as the sole potassium source. This study enhances our understanding of the composition of rhizosphere potassium-solubilizing bacteria and provides valuable insights into their potassium-solubilizing mechanisms at the transcriptional level.
-
The rhizosphere loamy soil (depth 0−15 cm, 2 mm around the plant roots) of wild floribunda plants at the vegetative growth stage was sampled from the Limutai Mountain, Jizhou District, Tianjin, China (39°45' N, 117°05' E, altitude 997 m). The physical and chemical properties of rhizosphere soil were analyzed, revealing an organic matter content of 45 g/kg, total nitrogen of 1.8 g/kg, and total phosphorus of 1.6 g/kg, and a pH of 7.5[39]. The control Pseudomonas putida strain KT2440 was sourced from the American Type Culture Collection (ATCC), USA. Feldspar, with a potassium content of 16.9%, was procured from Macklin, China. Safranine O was purchased from Dingguo, China, while other agents were purchased from Aladdin, China.
Isolation of potassium-solubilizing bacteria
-
The potassium-solubilizing bacteria were isolated from fresh rhizosphere soil collected from wild Lespedeza floribunda plants. Briefly, 1 g of soil was suspended in a liquid feldspar medium containing sucrose (10 g), Na2HPO4 (0.4 g), (NH4)2SO4 (1 g), MgSO4 (0.2 g), yeast extract (2 g), FeCl3 (0.02 g), feldspar (5 g), distilled water (1000 mL) with a pH of 7.2[36]. The soil suspension was then incubated at 30 °C with shaking for 48 h. After incubation, the cultured suspension was serially diluted with distilled water at 10-fold gradients. Aliquots of the diluted solutions were spread onto feldspar medium plates, and incubated at 30 °C for 48−72 h. Colonies were subsequently transferred to fresh plates for purification, resulting in the isolation of potassium-solubilizing strains designated as NKK1 to NKK15. To identify the phylogenetic relatedness of strain NKK10, its 16S rDNA sequence was obtained, and the sequence was submitted to the NCBI system (accession number OR53494). This sequence was then analyzed using the online BLASTN tool on the NCBI website and used to construct a phylogenetic tree via the MEGA11.0 software.
Potassium-releasing assays
-
To evaluate the ability of the strains to solubilize potassium from feldspar, they were cultivated in a liquid feldspar medium under shaking conditions at 30 °C for 7 s. After incubation, the cultures were centrifuged at 12000 rpm for 5 min to remove the feldspar and bacterial cells. The potassium concentration in the resulting supernatants was determined using an inductively coupled plasma–mass spectrometer (Elan drc-e, PerkinElmer, USA).
Bacterium staining and microscopy
-
Bacterial cells were cultured in liquid feldspar medium for 7 d. The feldspars with adsorbed bacterial cells were isolated by standing the cultures for 10 min, followed by sampling the pellets. These pellets were resuspended in distilled water, and safranine O was added to achieve a final concentration of 100 mg/L. After 10 min of staining, the feldspars were gently washed twice with distilled water and observed under a light microscope (BX43, Olympus, Japan). Additionally, the treated feldspars were sampled, fixed with 3% glutaric dialdehyde, dried using a fertilizing vacuum drier (SCIENTZ 10N, SCIENTZ, China), and estimated with a scanning electron microscope (TESCAN MIRA LMS, Czech).
To evaluate bacterial growth in the liquid feldspar medium, cultures were incubated for 0~12 d. The optical density of the cultures, measured after standing at room temperature for 10 min, was measured using a UV-Vis spectrometer (SmartSpecTM Plus, BioRad, USA). The number of viable bacterial cells on the feldspar stones at different time points was determined by the colony-forming unit (CFU) method[40]. Cultures were serially diluted by distilled water, and the diluted solutions were plated onto the K+-rich medium plates. CFUs were counted after 3 d of incubation of the plates at 30 °C.
Construction of phylogenetic tree
-
The NKK10 cells were cultured for 3 d in a K+-rich medium (containing sucrose 10 g, K2HPO4 0.3 g, NaCl 0.1 g, (NH4)2SO4 1 g, MgSO4 0.2 g, yeast extract 2 g, FeCl3 0.02 g, distilled water 1,000 mL, pH = 7.2). Genomic DNA was extracted from the cells using a soil DNA extraction kit ((Biomarker Technologies, Beijing, China). The 16S rDNA of the bacterial strains was amplified from the genomic DNA using primers 27F (5'-AGAGTTTGATCCTGGCTCAG-3') and 1492 R (5'-GGTTACCTTGTTACGACTT-3'). The PCR product was sequenced by Tsingke Biotech., China and the resulting sequence of NKK10 16S rDNA (accession number OR53494) was submitted to the NCBI BLAST system. Similar sequences were retrieved for construction of the phylogenetic tree of NKK10 via the MEGA11.0 software by using the Neighbor-Joining method[36,41] .
mRNA sequencing and transcriptional profiling assays
-
To investigate the differentially expressed genes (DEGs) of NKK10 during potassium solubilization, NKK10 cells were cultured for 7 d in liquid feldspar medium or a control K+-rich medium. Following incubation, bacterial cells were harvested by centrifugation at 12,000 rpm for 3 min. Total mRNA was extracted from the obtained cells by using an RNA extraction kit (Promega, Beijing, China), and corresponding cDNA was prepared with the assistance of poly-T oligo-attached magnetic beads to constructed the cDNA libraries. The libraries were then sequenced by using the Illumina NovaSeq platform to generate 150 bp paired-end reads (BioZeron, China) system. The obtained sequences were analyzed with the C. taiwanensis reference genomes in the NCBI database (
www.ncbi.nlm.nih.gov/genome/1372?genome_assembly_id=171204 ). The differentially expressed genes were then assigned into the Gene Ontology (GO) terms and the Kyoto Encyclopedia of Genes and Genomes (KEGG) terms, by using the BioZeron analysis system and the online system of Biocloud (www.biocloud.net )[42] .Statistical analysis
-
Each experiment in this study was performed with three replicates (n = 3). The values were shown in the figures with the means ± the standard errors. Differences between the groups were compared by the t-Student tests (p < 0.05). All statistical tests were performed with the SPSS software package (Version 20, IBM).
-
After separation and purification, a total of 15 potassium solubilizing bacterial strains were obtained on the plate, coded NKK1~ NKK15, for further study. After 7 d of culturing in liquid feldspar medium, the 15 strains led to the release of K+ from potassium feldspar from 0.02 to 0.35 mg/mL (Fig. 1a). Among them, the strain NKK10 exhibited the highest potassium-releasing capacity, with the released potassium reaching 0.35 mg/mL (Fig. 1a). Time-dependent potassium release assays showed that the bacterium-free control group and the P. putida (the control P. putida strain KT2440) group only slightly released K+ to 0.05 mg/mL even after 14 d. In contrast, the NKK10 group had remarkable potassium release from day 2 to day 10 and reached 0.57 mg/mL after 14 d (Fig. 1b). Therefore, the strain NKK10 was chosen as the high-efficiency potassium-solubilizing strain for further investigations.

Figure 1.
Potassium release from potassium feldspar induced by the bacterial strains isolated from Lespedeza floribunda rhizosphere soil. (a) Potassium release induced by the strains from the isolated strains after 7 d of culturing. Control indicates the group with no addition of the bacterial strains in the potassium feldspar medium. (b) Time-dependent potassium release by the P. putida strain KT2440. The error bars indicate the standard errors of each group (n = 3). The asterisks (*) indicate significant difference between the NKK10 group and other groups (p < 0.05).
The strain NKK10 has high-growth biomass in the potassium feldspar medium
-
The growth of strain NKK10 was assessed in a feldspar medium. After 7 d of cultivation, the feldspars were isolated for microscope observation. While the control P. putida group showed sparse bacterial colonization on the feldspar, leaving its smooth surface of the feldspar, the NKK10 group exhibited extremely high levels of bacterial cells, with the cells forming dense biofilms on the feldspar (Fig. 2a, b). The NKK10 group showed much higher OD600 values and CFU than the control and the P. putida-treated group (Fig 2c, d). For instance, on day 8, the NKK10 group exhibited an OD600 value of 0.88 and the viable bacterial cells at 109.1 CFU/g stone. In contrast, both the control and the P. putida-treated group showed OD600 below 0.16, and viable cells counts below 102.8 CFU/g stone (Fig. 2c, d). These results indicated that strain NKK110 maintained stable growth in a medium using potassium feldspar as the sole potassium source. This is consistent with the observation that NKK10 possesses a high capacity to release potassium from feldspar, efficiently supplying potassium to sustain bacterial cell growth.
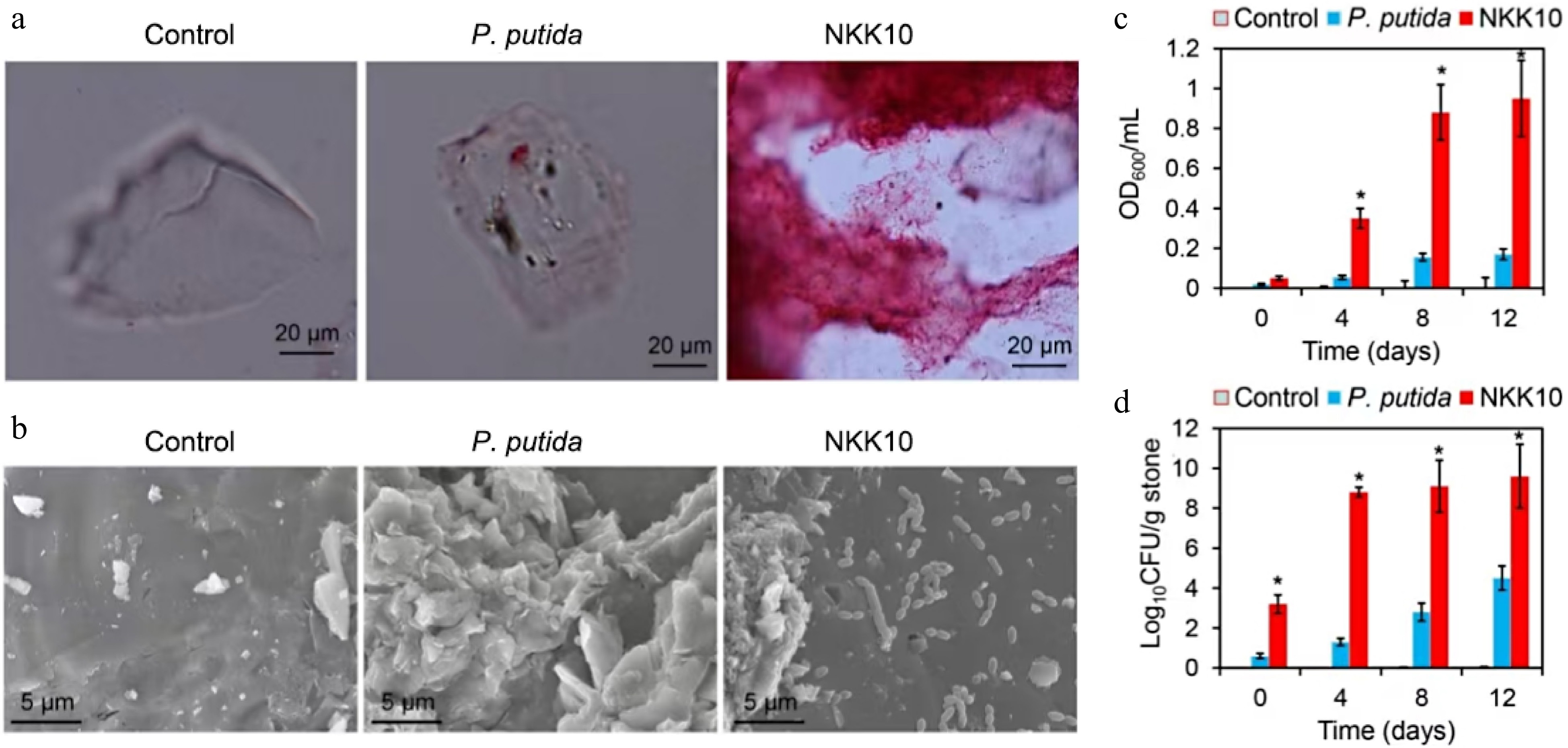
Figure 2.
Effect of potassium feldspar on growth of the P. putida KT2440 and NKK10. (a) Microscope images of feldspars after co-incubation of the strains for 7 d. The strains were incubated in the potassium feldspar medium, the feldspars were then sampled, and stained by safranine O for microscopy. (b) Scanning Electron Microscope (SEM) images of the feldspar after 7 d of treatment by the bacterial cells. The blue arrows indicate the distribution of bacterial cells on the feldspar surface. (c) Optical density of the cultures after 10 min of standing at the room temperature. (d) The number of viable bacterial cells on the feldspar stones at different time points. The error bars indicate the standard errors of each group (n = 3). (*) indicate significant difference between the NKK10 group and other groups (p < 0.05).
The potassium-solubilizing strain NKK10 belongs to C. taiwanensis
-
To determine the phylogenetic relatedness of strain NKK10, its 16S rDNA sequence was analyzed. As shown in Fig. 3, strain NKK10 is closely related to members of the Cupriavidus species, including C. taiwanensis, C. nantongensis, C. malaysiensis, and C. pampae. Especially, NKK10 shared the highest similarity (99%) with C. taiwanensis CD3, confirming its closest phylogenetic relatedness to C. taiwanensis.

Figure 3.
Phylogenetic tree of NKK10 constructed by the MEGA11.0 software with the Neighbor-Joining method.
Strain NKK10 drastically alters transcriptional profiling analysis in feldspar medium
-
To elucidate the mechanism of potassium release by NKK10, transcriptional profiling analysis was performed on NKK10 cells cultured in feldspar medium, compared to those grown in a K+-rich medium control. The heat map revealed remarkable changes in gene expression in the feldspar medium (FD1-FD3) compared to the control group (CK1-CK3, Fig. 4a). Especially, in samples FD1 and FD3, the majority of genes were upregulated (Fig. 4a). A volcano plot further highlighted that 523 genes were upregulated, whereas only 81 genes were downregulated in the feldspar medium (Fig. 4b). These findings demonstrated that culturing NKK10 cells in feldspar medium strongly altered the transcriptional profiling analysis of this potassium-solubilizing bacterium.

Figure 4.
Altered transcriptional profiling of the NKK10 cells cultured in feldspar medium vs in the control K+-rich medium. (a) Heat map of the transcription profiling. The samples CK1-CK3 indicate the control samples in which the NKK10 cells were cultured in the K+-rich medium, while the samples FD1-FD3 indicate the treated samples in which the cells were cultured in the feldspar medium. (b) Volcano mapping of the expressed genes.
The strain NKK10 exhibits changed GO map in feldspar medium
-
GO analysis was performed to illustrate the change in the GO map f NKK10 cells cultured in the feldspar medium. Regarding biological processes, culturing bacterial cells in the felspar medium led to the upregulation of genes involved in biological regulation, interspecies interaction between organisms, localization, multi-organism process, reproduction, reproductive process, response to stimulus, and signaling (Fig. 5a). For molecular functions, the feldspar-treated group exhibited increased expression of genes related to transcriptional regulator activity (Fig. 5a). These results indicated that NKK10 cells robustly activated a series of signaling pathways to strengthen intercellular interaction, thereby maintaining high metabolic activity and growth rate. In addition, the feldspar-treated group showed a comparable number of genes related to cellular components (CC, Fig. 5a), suggesting that the alteration of the potassium sources had minimal impact on cellular components. However, the dot map analysis revealed slight changes in the expression of the genes associated with transferase activity (transferring hexosyl groups), cytokinesis, acid-amino acid ligase activity, localization, and protein binding (Fig. 5b). In contrast, the expression of the genes related to cell movement, including bacterial-type flagellum basal body, bacterial-type flagellum-dependent swarming motility, chemotaxis, bacterial-type flagellum, and locomotion, was significantly changed, with an R-factor > 0.50, and P value < 0.01 (Fig. 5b). These results indicated that bacterial motility was significantly affected in the feldspar medium.
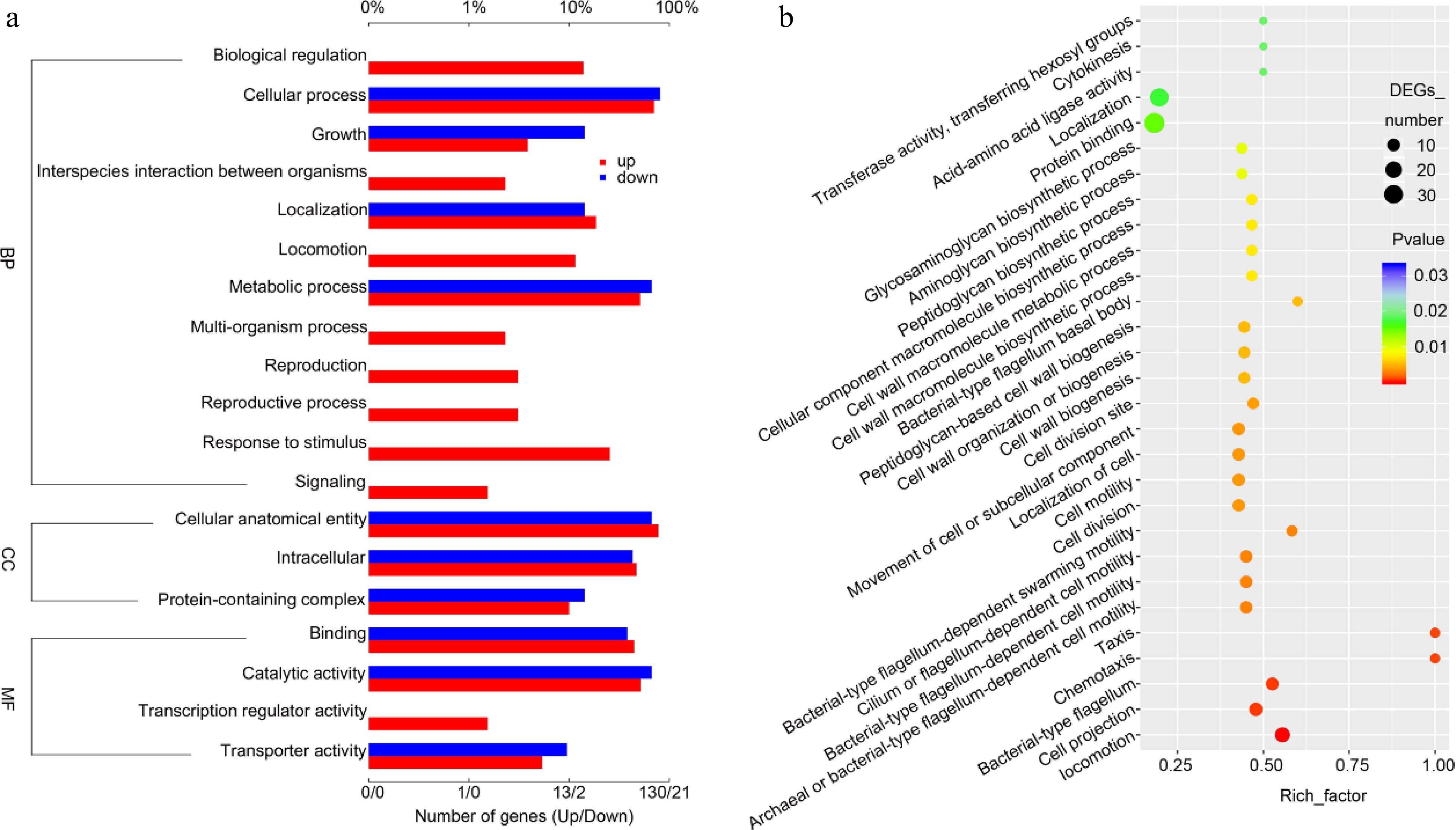
Figure 5.
Gene Ontology analysis (GO analysis) of the NKK10 transcriptional profiling in feldspar medium versus in the control K+-rich medium. (a) Bar chart of GO analysis showing the gene groups with changed expression levels. (b) Dot map of GO analysis showing the gene groups with changed expression levels.
The strain NKK10 remodels metabolic pathways in feldspar medium
-
KEGG pathway analysis of transcriptional profiling was conducted in NKK10 cells. While pathways related to lysine/aminobenzoate/sugar metabolism were only slightly changed (p > 0.12, R factor < 0.25), several pathways were strongly impacted in feldspar medium (p < 0.04, Fig. 6a). These included ABC transporters, Pseudomonas aeruginosa-type biofilm formation, flagellar assembly, peptidoglycan biosynthesis, starch sucrose metabolism, and bacterial chemotaxis, For example, the pathways of peptidoglycan biosynthesis and Pseudomonas aeruginosa-type biofilm formation had R factors exceeding 0.38 and 0.25, respectively (Fig. 6a).

Figure 6.
KEGG analysis of the NKK10 transcriptional profiling in feldspar medium vs in the control K+-rich medium. (a) Dot map of KEGG analysis showing the gene groups with changed expression levels. (b) The proteins with up-regulated expression (indicated by red color) in the pathway of peptidoglycan biosynthesis. The numbers in the brackets indicate the Log2(fold change) values of the corresponding genes. (c) The proteins with up-regulated expression (indicated by red color) in the pathway of biofilm formation.
In the pathway of DAP-type peptidogulcan synthesis, the proteins MurC and MurD, which catalyze the conversion of UDP-GlcNAc to UDP-MurNAc-L-Ala-γ-D-Glu, as well as MurF, MraY, MurG, and ClassA PBP, which drives the subsequent transformation of UDP-MurNAc-L-Ala-γ-D-Glu to DAP-type peptidoglucan, were upregulated (Fig. 6b). Similarly, in the Pseudomonas aeruginosa-type biofilm formation pathway, upregulation was observed in proteins related to the Type 3 secretion system (PilJ, PilH, and PilG), the Type 6 secretion system (HSI-I), and polysacchride synthesis (WspA, WspB/D & WspE, WspR, PelF) in the cells cultured in the feldspar medium were upregulated (Fig. 6c). These results indicate that NKK10 cells remarkably remodeled their metabolic pathways, particularly those related to cell wall synthesis and biofilm formation, to optimize metabolism and growth with feldspar as the sole potassium source.
-
Potassium-solubilizing bacteria hold great promise in crop production due to their ability to reduce reliance on synthetic fertilizers and promote plant growth[43,44] . Rhizosphere soil serves as a rich reservoir for the potassium-solubilizing bacteria[45,46] . Numerous species, such as Priestia megaterium, Klebsiella variicola, Agrobacterium tumefaciens, Enterobacter cloacae, Mesorhizobium sp., Paenibacillus sp., Arthrobacter sp., Burkholderia sp., Pseudomonas rhodesiae, and Ralstonia mannitolilytica, have been isolated from various plant rhizosphere soils[36,47,48]. However, the diversity and functions of potassium-solubilizing bacteria in the rhizosphere of Leguminosae plants remain largely unexplored. In this study, rhizosphere soil from Lespedeza floribunda, a wild Leguminosae plant species, was sampled, leading to the isolation of potassium-solubilizing bacteria, including C. taiwanensis, a novel potassium-solubilizing bacterium. Traditionally, C. taiwanensis has been recognized for its high tolerance to heavy metal, having been isolated from heavy metal-contaminated soils[48,49]. However, its additional role in potassium solubilization and nutrient cycling in the rhizosphere is demonstrated. Therefore, C. taiwanensis is not only a heavy metal-tolerant bacterium, but also a potential PGPM, which may be used for development of inocula to optimize the soil quality and promote the plant growth.
Role of biofilms during potassium solubilization
-
Bacterial biofilms are specialized aggregates of bacterial cells that integrate and communicate with one another through extracellular polymeric substrates (EPS)[50,51]. These EPS, comprising polysaccharides, proteins, and nucleic acids envelop the cells, facilitating enhanced intercellular interaction and improving their tolerance to environmental stress[52,53]. Recently, it has been discovered that biofilms formed by heavy metal-capturing bacteria play a crucial role in strengthening bacterium-plant host interaction and enhancing heavy metal removal efficiency[54,55]. In this study, it was further found that the C. taiwanensis cells developed dense biofilms on the surface of feldspars for the solubilization of potassium, which is associated with the strong potassium-releasing capacity of this bacterium. Together, these findings implied that the regulation of bacterial biofilm may promote the activity of functional bacteria during agricultural production and environmental treatment.
Transcriptional regulation of potassium-solubilizing bacteria
-
Transcriptional regulation is a fundamental mechanism of living cells to intelligently respond to extracellular nutrition, stress, and other signals[56,57]. When the cells encounter nutrients with low water solubility, they may remodel the transcriptional profiling, leading to production and secretion of extracellular enzymes, organic acids, and other extracellular components to dissolve the nutrients[51,57,58] Transcription of functional genes dominates bacterial transformation and utilization of extracellular substances[59]. Herein, it was found that the C. taiwanensis cells, when they were cultured in feldspar medium, drastically remodeled transcriptional profiling for adapting to the potassium-depleted condition. As revealed by GO analysis, a series of gene groups, e.g., biological regulation, interspecies interaction between organisms, localization, multi-organism process, reproduction, reproductive process, response to stimulus, and signaling were upregulated in the feldspar medium as compared to the K+-rich medium. This suggested that the signal pathways involved in stress response and intracellular interaction were activated, resulting in a sensitive response to the feldspar sources. Further insights from the KEGG pathway analysis highlighted the activation of critical systems such as the Type III and Type VI secretion systems and polysaccharide synthesis pathways. These molecular mechanisms likely play a pivotal role in facilitating mineral attachment, biofilm stabilization, and the secretion of enzymes or organic acids that contribute to potassium release. For example, the upregulation of peptidoglycan biosynthesis-related proteins (e.g., MurC, MurD, MurF) and biofilm-associated proteins (e.g., WspA, WspB/D, PelF) underscores the bacterium's ability to remodel its cellular machinery for effective mineral weathering. Such adaptive responses demonstrate that C. taiwanensis employs sophisticated transcriptional strategies not only to survive under potassium-limited conditions but also to actively modify its local environment to enhance nutrient availability. These findings provide a deeper understanding of the transcriptional regulation underlying potassium solubilization and highlight the strain’s potential application in sustainable agriculture.
-
This study investigated the presence of potassium-solubilizing bacteria in the rhizosphere soil of wild Lespedeza floribunda and identified a novel high-efficiency potassium-solubilizing bacterium, C. taiwanensis, expanding the known diversity of functional soil microorganisms. It revealed the critical role of biofilm formation in bacterial potassium solubilization, demonstrating that C. taiwanensis forms specialized surface structures to enhance mineral weathering efficiency. Through transcriptional profiling analysis, this study uncovered the complex molecular networks regulating bacterial adaptation to mineral potassium, particularly the coordinated regulation of pathways related to cell wall synthesis, secretion systems, and biofilm formation. These molecular insights provide new strategies for improving microbial potassium-solubilization efficiency. Overall, the present research deepens the fundamental understanding of microbe-mineral interactions and offers valuable guidance for developing effective microbial agents to enhance soil quality and support sustainable agriculture.
This work was supported by Tianjin Science and Technology Research Project (22YFYSHZ00310, 24YFXTHZ00170).
-
The authors confirm contribution to the paper as follows: conceptualization, writing – original draft preparation: Zhang Y, Li P, Zhao L; data curation: Zhang Y, Li P, Zhang R; investigation: Zhang Y; validation: Zhang Y, Zhang R, Zhang X, Cong H, Zhao L; writing – review & editing: Zhang R, Zhang X, Cong H, Zhao L; formal analysis: Li P, Zhang R; resources: Li P, Zhao L; software: Li P, Cong H; methodology: Zhang R; visualization: Zhang R, Zhang X; project administration: Zhao L. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang Y, Li P, Zhang R, Zhang X, Cong H, et al. 2024. Characterization of the rhizosphere potassium-solubilizing Cupriavidus taiwanensis strain NKK10 by transcriptional profiling analysis. Progress in Reaction Kinetics and Mechanism 49: e004 doi: 10.48130/prkm-0024-0003 

Characterization of the rhizosphere potassium-solubilizing Cupriavidus taiwanensis strain NKK10 by transcriptional profiling analysis
- Received: 29 October 2024
- Revised: 24 November 2024
- Accepted: 26 November 2024
- Published online: 31 December 2024
Abstract: Potassium-solubilizing bacteria are microorganisms capable of dissolving potassium from potassium-containing minerals, with some species carrying the ability to promote plant growth. This study aims to explore the composition of potassium-solubilizing bacteria from the rhizosphere of wild Lespedeza floribunda, a representative Leguminosae plant, and the mechanisms underlying potassium release. Among the isolated bacteria, the Cupriavidus taiwanensis strain NKK10 exhibited highly efficient potassium solubilizing capacity from potassium feldspar, leading to potassium release of 0.35 and 0.57 mg/mL after 7 d and 14 d, respectively. Microscopic observation showed that the NKK10 cells were able to bind strongly to feldspar, resulting in the formation of a dense biofilm on the mineral surface. As revealed by mRNA sequencing and transcriptional profiling analysis, C. taiwanensis NKK10 drastically remodeled transcriptional profiling analysis in the feldspar medium, with upregulation of critical signaling pathways involved in interspecies interaction between organisms and biofilm formation in response to the stimulus of peptidoglycan synthesis. The results of this study extend our knowledge of the functional microorganisms in Leguminosae rhizospherie soil, and provide insights into the transcriptional regulation of potassium-solubilizing bacteria during the solubilization process.