






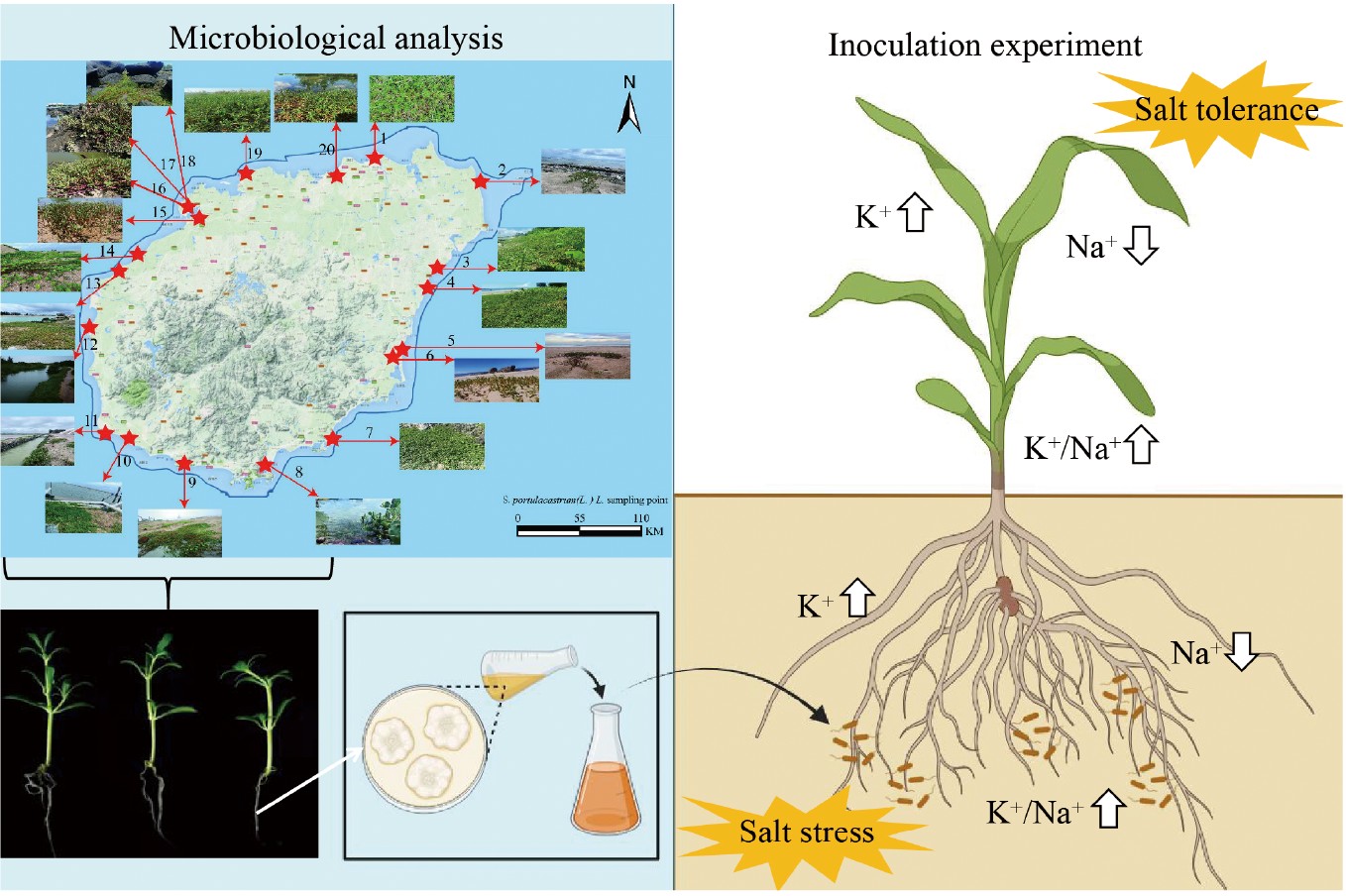



-
Soil salinization is recognized as one of the most pervasive and critical soil challenges worldwide[1]. Over 3% of the Earth's terrestrial surface is significantly affected by salinization[2], with saline soil areas expanding to 9.54 × 108 hm², a trend that continues to escalate annually[3]. Salt stress imposes a substantial threat to agricultural productivity by causing osmotic, ionic, and oxidative stress, thereby profoundly limiting crop development[4−6]. Studies have reported that crop yields can decrease by as much as 50% or more due to salt stress[7−8]. Although certain physical and chemical approaches may partially remediate saline soils, the prohibitive costs associated with such interventions hinder their widespread application[9,10]. Investigations have revealed that green additives, particularly microorganisms, can effectively alleviate the detrimental effects of salt stress on plants. Microbial inoculants have been recognized for their potential to achieve effective remediation while being environmentally sustainable, non-polluting, and capable of delivering enduring improvements[11−13].
Endophytic fungi (EF) represent a diverse group of microorganisms, extensively distributed within plants, capable of proliferating and persisting in significant quantities within cells or intercellular spaces across various plant parts, thereby showcasing remarkable species diversity[14]. These fungi are widely recognized for their substantial contributions to processes associated with plant growth, development, and reproduction[15,16], and their ability to effectively mitigate the adverse effects of salt-alkali stress on plants has been well-documented[17]. Research conducted by Moghaddam et al.[18] demonstrated that, under salt stress conditions, EF was capable of bolstering the stress resistance of tomato and cucumber, resulting in marked increases in proline levels and antioxidant enzyme activities within plant cells. Similarly, Wang et al.[19] reported that the seed-borne endophytic fungus Epichloe gansuensis enhanced the salt tolerance and increased the biomass of Achnatherum inebrians. Additionally, Falciphora oryzae, an endophytic fungus isolated from the roots of wild rice, was shown to colonize the roots of capsicum, significantly improving its salt stress tolerance[20]. The biosynthesis and signaling of indoleacetic acid and abscisic acid were notably elevated compared with non-inoculated treatments[20]. Furthermore, the endophytic fungus Piriformospora indica has been found to aid plants in adapting to saline stress by enhancing nutrient uptake and modulating plant hormone levels through polyamine-mediated changes under such conditions[21]. Collectively, these findings highlight the substantial potential of EF for improving plant salt tolerance, offering a promising biological strategy for ecological restoration and agricultural production in saline-affected lands.
EF derived from halophytes are recognized for their significant agricultural applications, including the enhancement of plant salt tolerance, promotion of plant growth, improvement of soil quality, and remediation of saline land[7,22,23]. It has been observed that endophytes residing in plants exposed to stress or adapting to extreme environments exhibit a tolerance to such conditions and are capable of transferring this tolerance to their host plants[18,24,25]. Sarocladium kiliense FS18, isolated from the halophyte Suaeda salsa, has been shown to colonize maize rhizosphere soil under high salinity conditions, enhancing soil enzyme activity, influencing microbial communities, and significantly improving the growth and physiological traits of maize seedlings, thereby alleviating salinity-induced damage to maize[10]. Pan et al.[23] revealed that the halophyte endophyte Curvularia sp. can establish a symbiotic relationship with Populus tomentosa, increasing chlorophyll and proline levels in plant leaves, thus improving plant stress resistance. Ting et al.[26] demonstrated that the endophytic fungus Fusarium oxysporum can significantly promote the growth of banana. Furthermore, Huong et al.[27] reported that EF is capable of decomposing insoluble soil substances, thereby maintaining crop health through phosphorus solubilization and nitrogen fixation. These investigations highlight the considerable potential of halophyte-derived EF for agricultural and ecological restoration applications. Such fungi not only enhance crop yields but also enable crops to adapt more effectively to diverse environmental stresses, offering innovative strategies for sustainable agricultural development.
Sesuvium portulacastrum, classified within the genus Sesuvium of the family Aizoaceae, is a succulent herbaceous halophyte commonly found in tidal flat regions, renowned for its substantial salt tolerance and environmental adaptability[28,29]. Investigations into the salt tolerance mechanisms of S. portulacastrum have predominantly focused on physiological and molecular aspects[28,30]. However, the current understanding of EF associated with S. portulacastrum and its role in plant salt tolerance remains limited, necessitating further exploration. In the present study, samples of S. portulacastrum collected from coastal regions surrounding Hainan Island were utilized as research subjects. Conventional tissue isolation culture techniques, combined with molecular biology methods, were employed to isolate and identify root EF from 20 sampling sites of S. portulacastrum. The diversity and distribution of these EF were preliminarily assessed, salt-tolerant strains were identified through plate screening, and their contributions to enhancing plant salt tolerance were evaluated. This study demonstrated that EF derived from halophyte roots can significantly improve the salt tolerance of crops and provided insights into the mechanisms by which salt-tolerant strains enhance crop resilience under salinity stress. The findings of this research highlight the potential of salt-tolerant EF to advance studies aimed at improving plant salt tolerance and optimizing the utilization of saline farmland.
-
From May 2022 to August 2023, a total of 20 sampling sites were established along the coastal areas of Hainan Island at intervals of approximately 50 to 100 km (Fig. 1, Supplementary Table S1). These sites experience an average annual temperature of approximately 25 °C and are predominantly composed of sandy soil, which is regularly inundated by seawater during high tides. The stems of S. portulacastrum grow flat on the sandy substrate, forming dense clusters that spread continuously, resulting in a carpet-like ground cover. At each location, systematical subdivision into five distinct zones was conducted, from which 5−10 S. portulacastrum plants with diverse growth characteristics were randomly sampled. The roots of S. portulacastrum were carefully excavated, and any excess soil was meticulously removed. The collected samples were then placed in sterile, sealable bags, stored in ice-packed containers, and promptly transported to the laboratory for the subsequent isolation of EF.
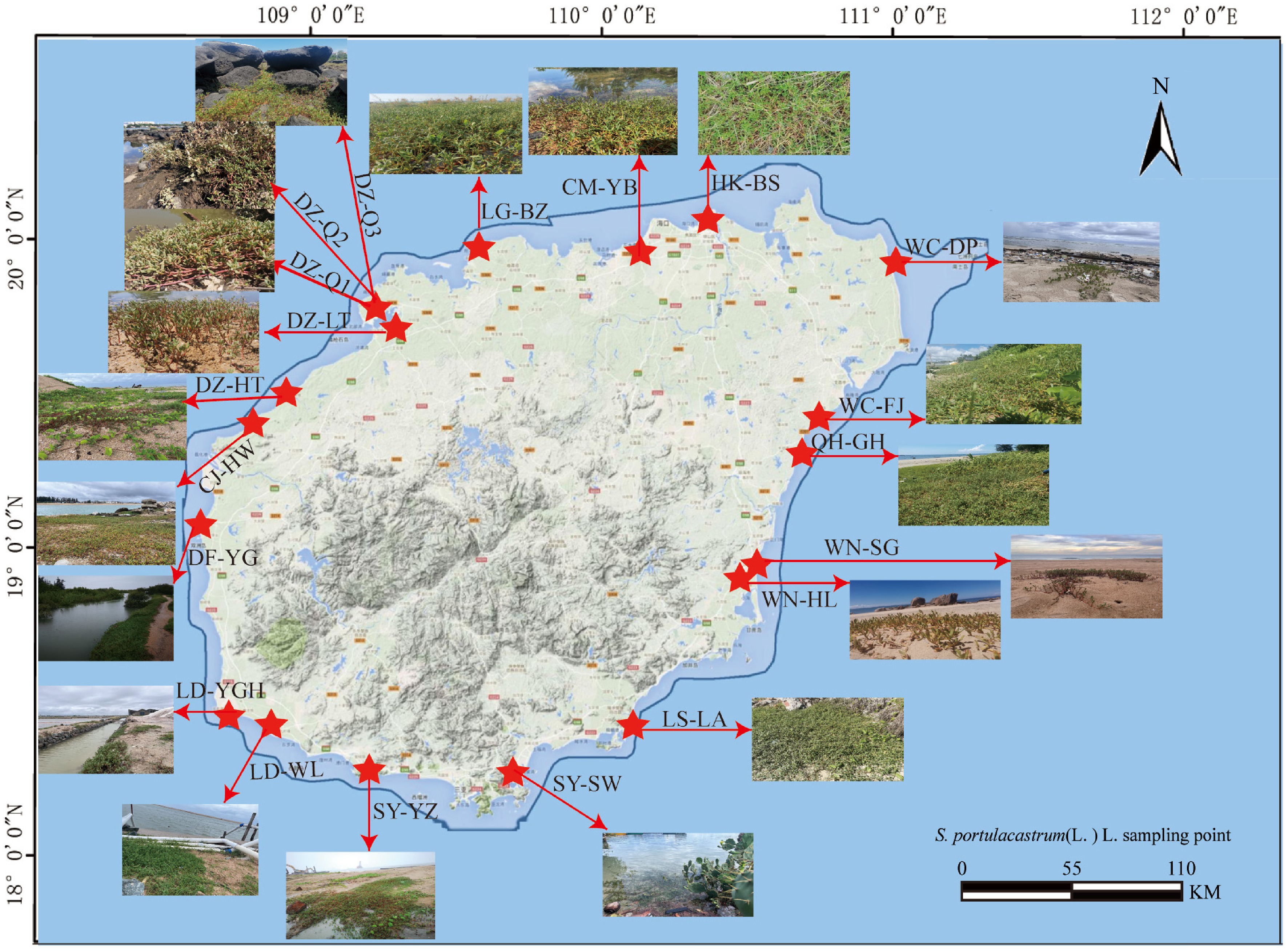
Figure 1.
Sampling points of S. portulacastrum.
Isolation and identification of root EF from S. portulacastrum
-
The isolation and purification of root EF were performed according to the method described by Wang et al.[31]. The activated EF were inoculated into centrifuge tubes containing potato dextrose broth (PDB) liquid medium and cultured in a constant-temperature shaker set at 28 °C and 200 rpm for 2 to 3 d. Subsequently, the mycelia were collected through centrifugation at 12,000 rpm for 10 min. Total DNA from the endophytic fungal strains was extracted using the Fungal Genomic DNA Extraction Kit (Product No. D2300) from Beijing Solarbio Science & Technology Co., Ltd., following the manufacturer's protocols. The extracted DNA was utilized as a template for amplification using the universal primers ITS1 (5'-TCCGTAGGTGAACCTGCGC-3') and ITS4 (5'-TCCTCCGCTTATTGATATGC-3')[32]. The polymerase chain reaction (PCR) amplification system comprised 25 μL of 2 × Taq Plus Master Mix II (Dye Plus), 2 μL each of forward and reverse primers, 2 μL of template DNA, and 19 μL of ddH2O, resulting in a total reaction volume of 50 μL. The PCR amplification conditions included an initial denaturation at 94 °C for 5 min, followed by 30 cycles of denaturation at 94 °C for 40 s, annealing at 55 °C for 40 s, and extension at 72 °C for 50 s. A final extension step was conducted at 72 °C for 10 min, after which the PCR products were sent to Sangon Biotech (Shanghai) Co., Ltd. (Shanghai, China) for sequencing. High-quality sequences obtained through sequencing were aligned using Clustal X and subsequently trimmed with BioEdit. The sequences were clustered into operational taxonomic units (OTUs) using Mothur, based on a 99% sequence identity threshold. Representative sequences from each OTU were compared to the NCBI database (
www.ncbi.nlm.nih.gov ) using BLAST to determine their taxonomic classifications[33].Analysis of root endophytic fungal diversity
-
The predominant strains were analyzed based on relative frequency (RF)[34]. To evaluate the abundance and distribution uniformity of endophytic fungal populations, the Shannon-Wiener index[35] was employed. The Simpson diversity index[36] was utilized to assess species richness and community evenness. Furthermore, the evenness index[37] was applied to examine the distribution uniformity of various EF associated with the host. Sorenson's similarity coefficients[38] were used to compare the similarity of endophytic fungal species across different locations.
Analysis of salt tolerance in root EF
-
Four mycelial discs, each measuring 5 mm in diameter, were excised from the periphery of the colony using a sterile punch. These discs were subsequently transferred onto PDA media under two conditions: without salt (control) and with NaCl concentrations of 0.25, 0.5, and 0.75 M. The cultures were incubated in an inverted position within a constant-temperature incubator maintained at 28 °C. Fungal strain growth was monitored daily, and colony diameters were measured over 7 d. Each experimental trial consisted of three biological replicates.
Evaluation of phenotypic and physiological responses of maize under salt stress
-
To investigate the effects of salt-tolerant EF on plant salt tolerance, the strain LG-BZ-9, recognized for its significant salt tolerance, was selected to evaluate its impact on maize growth under saline conditions. The EF strain LG-BZ-9 was introduced into 500 mL of PDB and cultivated under controlled conditions in a constant temperature shaker maintained at 28 °C, with an agitation speed of 200 rpm for 3−4 d. Following the incubation period, the culture underwent centrifugation at 8,000 rpm for 10 min to separate the mycelium. The obtained mycelium was subsequently processed and diluted with distilled water to achieve a concentration of 1 g/L for subsequent use. Maize seeds of a common variety were planted in a substrate mixture consisting of vermiculite and nutrient soil in a 2:1 ratio. Upon reaching the one-leaf stage, the seedlings were subjected to four distinct treatments: standard irrigation (Control), fungal root inoculation using LG-BZ-9, treatment with a 250 mM NaCl solution (NaCl), and a combined application of the fungus and salt solution (LG-BZ-9 + NaCl). Growth conditions were photographed and meticulously documented daily. On the tenth day, samples were collected to evaluate seedling fresh weight, plant height, and chlorophyll content[39]. The shoots and roots of maize plants from each treatment group were harvested separately, dried, and subjected to analysis for Na+ and K+ content[40].
Statistical analysis
-
All experiments were conducted in triplicate using three biological replicates. Statistical analysis was performed with the SPSS 20 software, and differences among groups were determined through one-way analysis of variance (ANOVA). Data are presented as means accompanied by standard errors, with p < 0.05 considered statistically significant.
-
In this study, 426 cultivable EF were successfully isolated from 1,180 tissue blocks. The CM-YB site demonstrated the highest number of isolated strains, comprising a total of 48 EF, whereas the DZ-Q2 site yielded the fewest, with only ten EF obtained (Supplementary Table S2). Following sequencing, the 426 effective fungal sequences were subjected to analysis using the Basic Local Alignment Search Tool within the NCBI database (
www.ncbi.nlm.nih.gov ) to determine species and genus sequence information. The results revealed that the 426 fungal sequences were classified into 112 distinct OTUs (Fig. 2, Supplementary Table S3). Statistical evaluations indicated that these 112 EF were distributed across two phyla, eight classes, 16 orders, 37 families, and 47 genera.
Figure 2.
Morphological characteristics of root EF isolated from S. portulacastrum. The endophytic fungal isolates were cultured on a PDA medium for 7 d and photographed. The numbers correspond to the different fungal species.
Analysis of root endophytic fungal diversity in S. portulacastrum from various sampling points
-
The diversity indices of EF were observed to vary across different regions. The Shannon-Wiener index displayed its highest values in HK-BS (2.45), QH-GH (2.43), and WN-SG (2.35), whereas the lowest values were recorded in LG-BZ (1.12) and DZ-LT (1.12). Similarly, the Simpson index reached its peak in QH-GH (0.91), HK-BS (0.9), DZ-HT (0.88), WN-SG (0.88), and DZ-Q3 (0.88), while the lowest values were noted in LG-BZ (0.51) and DZ-LT (0.45). The evenness index exhibited the highest values in QH-GH (0.94), SY-SW (0.94), and CJ-HW (0.92), with the lowest indices recorded in CM-YB (0.38) and DZ-LT (0.34) (Table 1). The similarity coefficient was employed to reflect the resemblance in species composition across distinct communities. It was determined that the similarity of EF across the 20 sampling sites was generally low. The highest similarity index was identified between LG-BZ and DF-YG (0.5), whereas many sites displayed zero similarity with each other (Fig. 3). These results indicate that the root-culturable endophytic fungal populations of S. portulacastrum across the 20 sampling points were highly diverse, with minimal overlap in species composition.
Table 1. Diversity index of endophytic fungi in Sesuvium portulacastrum roots at different sampling sites
Sampling point name Shannon-Wiener index Simpson diversity index Evenness index HK-BS 2.45 0.9 0.83 WC-DP 1.87 0.78 0.72 WC-FJ 1.93 0.81 0.76 QH-GH 2.43 0.91 0.94 WN-SG 2.35 0.88 0.8 WN-HL 1.91 0.83 0.84 LS-LA 1.43 0.69 0.7 SY-SW 1.89 0.84 0.94 SY-YZ 1.84 0.83 0.9 LD-WL 1.99 0.85 0.91 LD-YGH 2.26 0.87 0.74 DF-YG 1.61 0.78 0.84 CJ-HW 1.71 0.81 0.92 DZ-HT 2.26 0.88 0.87 DZ-LT 1.12 0.45 0.34 DZ-Q1 1.48 0.69 0.63 DZ-Q2 1.97 0.84 0.9 DZ-Q3 2.28 0.88 0.82 LG-BZ 1.12 0.51 0.51 CM-YB 1.34 0.56 0.38 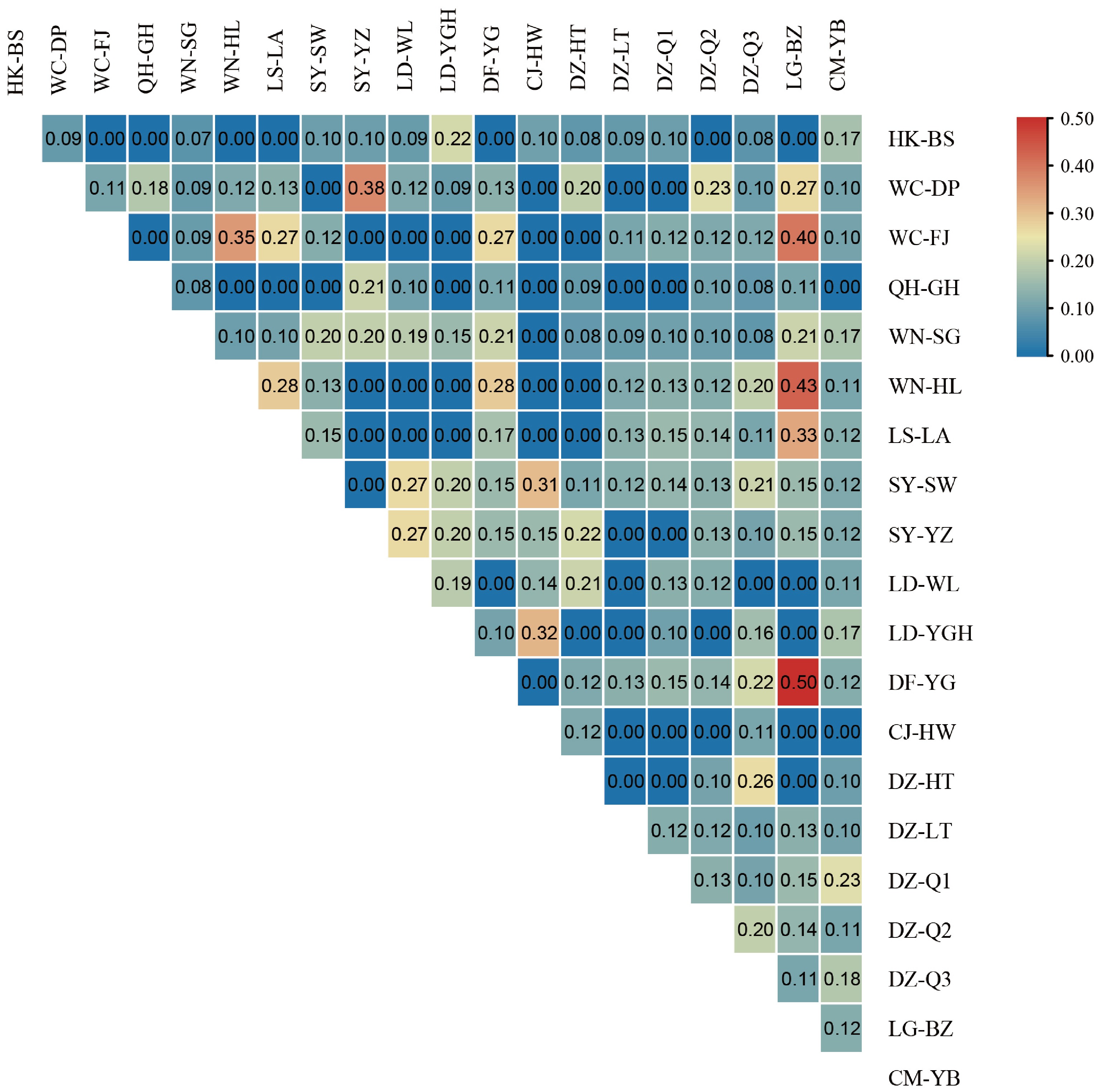
Figure 3.
Similarity index of EF from different sampling points. The values in the square box represent the similarity index of EF between two sites. Red and blue represent high and low similarity indices, respectively.
An analysis of the overall endophytic fungal community revealed that, at the family level, the most abundant families were Nectriaceae, Pleosporaceae, Xylariales incertae sedis, and Aspergillaceae, which constituted 21.4%, 19.5%, 15.5%, and 10.3% of the total, respectively (Fig. 4a). At the genus level, Fusarium emerged as the most prevalent genus, with 91 isolate sequences exhibiting the highest similarity to this genus, accounting for 21.36% of the total. Pleosporales followed, with 83 isolates identified through sequence analysis, representing 19.48%. Additionally, Monosporascus was detected in 66 isolates, comprising 15.49% of the total (Fig. 4b).
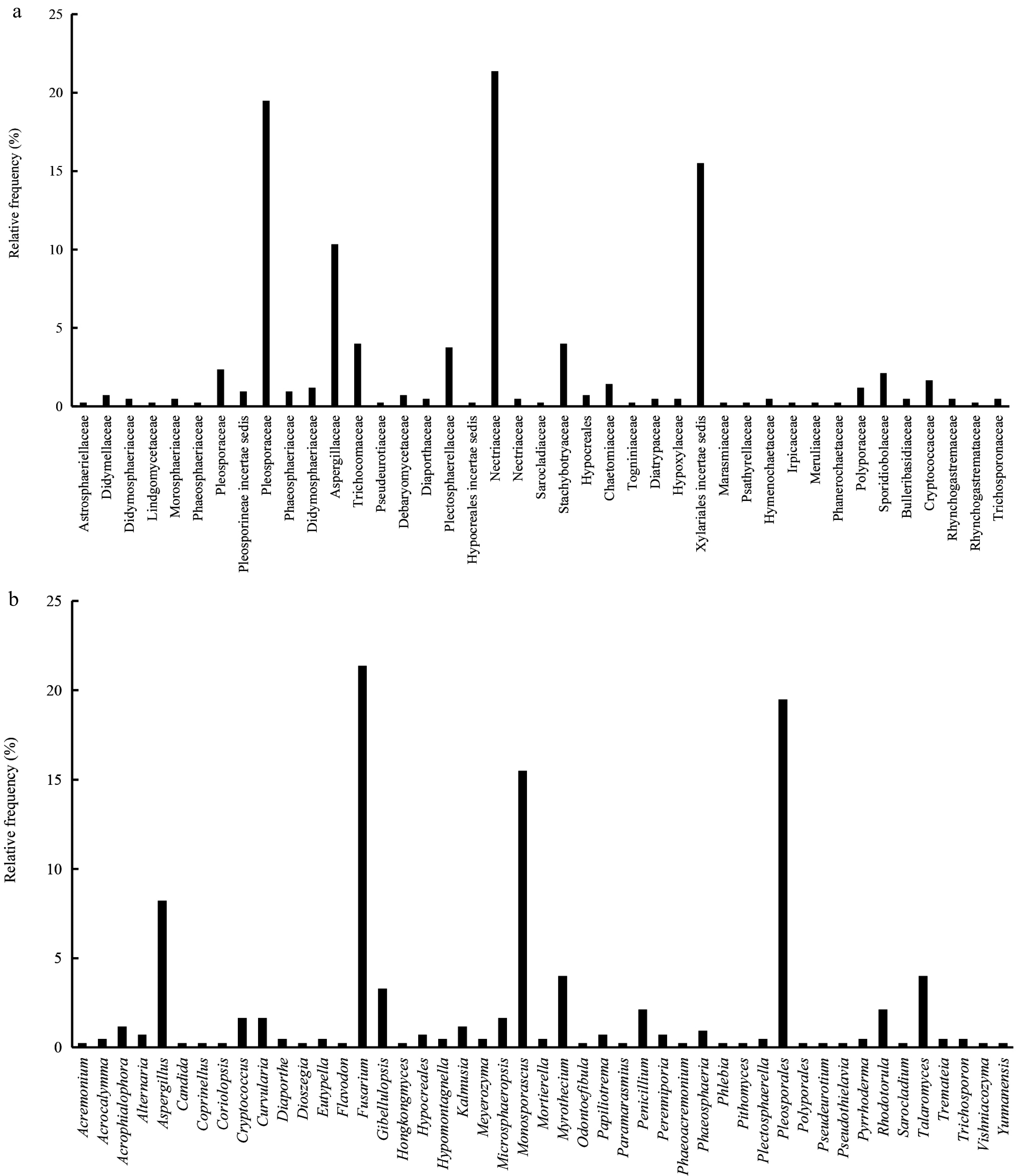
Figure 4.
Relative distribution frequency of culturable root EF. (a) RF of root EF at the family level. (b) RF of root EF at the genus level.
Using the RF of endophytic fungal isolation as the evaluation criterion, the dominant genera of EF were systematically analyzed. Fusarium was identified as the predominant genus in HK-BS (Fig. 5a), WN-HL (Fig. 5f), and CM-YB (Fig. 5t), exhibiting RF values of 40.63%, 31.26%, and 77.08%, respectively. Pleosporales emerged as the dominant genus in WC-FJ (Fig. 5c), LS-LA (Fig. 5g), DZ-LT (Fig. 5n), DZ-Q1 (Fig. 5p), and LG-BZ (Fig. 5s), with RF values of 41.18%, 46.67%, 73.53%, 50%, and 68.75%, respectively. DZ-Q2 (Fig. 5q) demonstrated co-dominance of Pleosporales and Hypocreales, both possessing RF values of 30%. The primary genus for LD-YGH (Fig. 5k), DF-YG (Fig. 5l), CJ-HW (Fig. 5m), and DZ-Q3 (Fig. 5r) was determined to be Monosporascus, with RF values of 57.58%, 50%, 33.33%, and 50%, respectively. Co-dominance of Monosporascus and Talaromyces was observed in LD-WL (Fig. 5j), each exhibiting an RF of 20%. Aspergillus served as the dominant genus in SY-YZ (Fig. 5i) and DZ-HT (Fig. 5o), with RF values of 60% and 32.14%, respectively. Certain sampling points exhibited unique dominant genera, including Myrothecium (RF = 50%) in WC-DP (Fig. 5b), Acrophialophora (RF = 22.73%) in WN-SG (Fig. 5e), and Talaromyces (RF = 38.46%) in SY-SW (Fig. 5h). In QH-GH (Fig. 5d), an even distribution of EF was observed, with Alternaria, Myrothecium, Plectosphaerella, Pleosporales, and Tremateia each demonstrating an RF of 12.5%. These findings suggest that the distribution of root EF in S. portulacastrum is characterized by both shared patterns and regional variations, with the RFs of fungal genera differing significantly across sampling points.
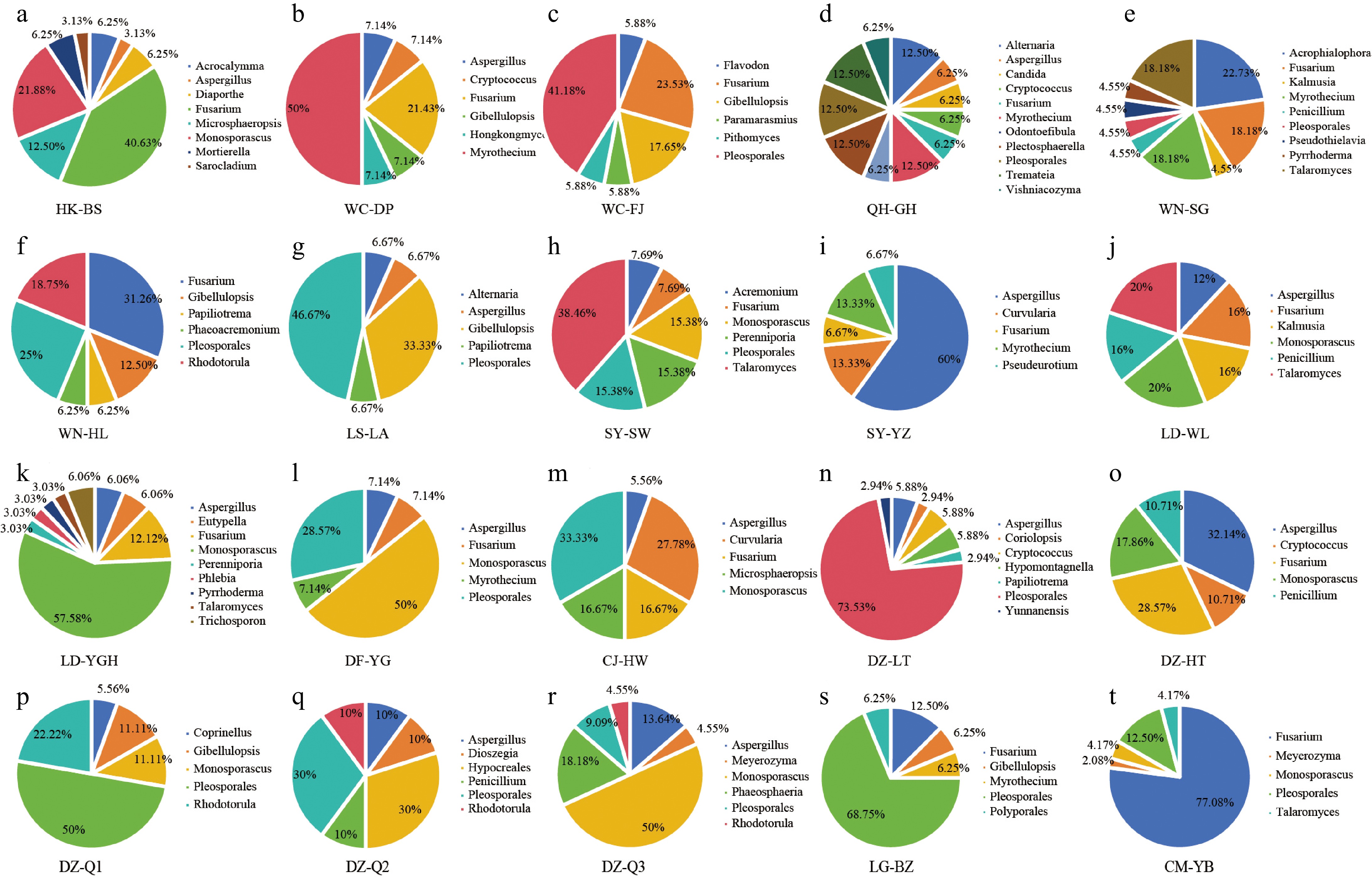
Figure 5.
RF of EF at the genus level from different sampling points. (a)−(t) Represent 20 sampling points.
Analysis of root endophytic fungal diversity in S. portulacastrum across Hainan Island
-
A total of 160 isolates of EF were obtained from nine sampling points situated in the eastern coastal regions of Hainan Island, encompassing Haikou City, Wenchang City, Qionghai City, Wanning City, Lingshui County, and Sanya City. These isolates were classified into 35 distinct genera (Fig. 6a). In addition, 266 EF isolates were collected from 11 sampling points in the western regions of Hainan Island, including Danzhou County, Lingao County, Chengmai County, Ledong County, Changjiang County, and Dongfang City, leading to their categorization into 28 genera (Fig. 6b). These results suggest that the eastern sampling points of Hainan Island exhibit greater diversity in root EF species in S. portulacastrum. Within the eastern region, Fusarium and Pleosporales were the most frequently isolated genera, accounting for 20% and 14.38%, respectively. Conversely, in the western region, Pleosporales, Fusarium, and Monosporascus emerged as the dominant genera, comprising 22.56%, 22.18%, and 21.43%, respectively. Common genera identified across both eastern and western sampling points included Aspergillus, Cryptococcus, Curvularia, Fusarium, Gibellulopsis, Kalmusia, Microsphaeropsis, Monosporascus, Myrothecium, Papiliotrema, Penicillium, Perenniporia, Pleosporales, Pyrrhoderma, Rhodotorula, and Talaromyces.
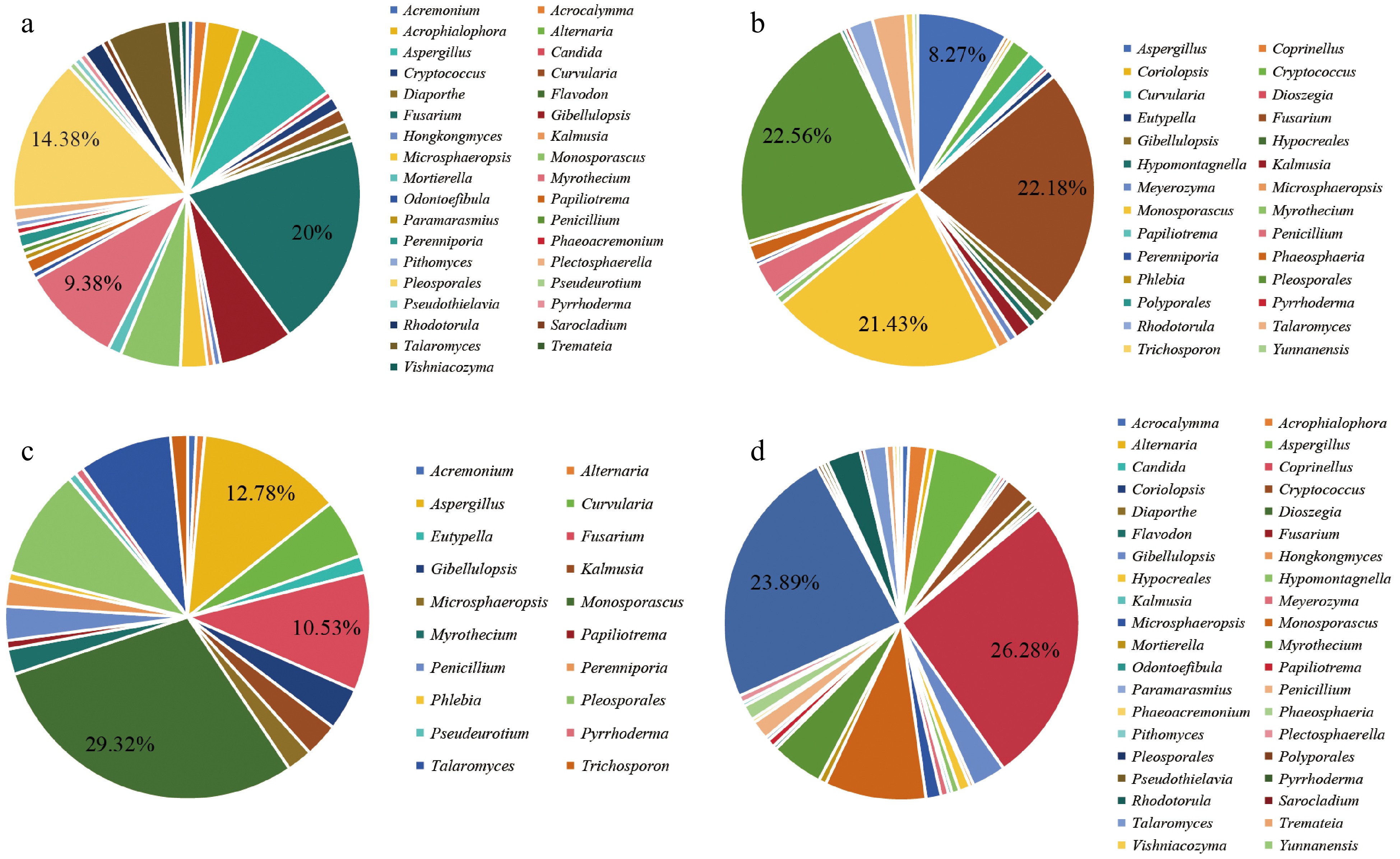
Figure 6.
Relative distribution frequency of EF in different regions. (a) Distribution frequency of S. portulacastrum root EF at the genus level in the eastern region of Hainan Island. (b) Distribution frequency of S. portulacastrum root EF at the genus level in the western region of Hainan Island. (c) Distribution frequency of S. portulacastrum root EF at the genus level in the southern region of Hainan Island. (d) Distribution frequency of S. portulacastrum root EF at the genus level in the northern region of Hainan Island.
A total of 133 isolates of EF were obtained from seven sampling sites situated in the southern region of Hainan Island, which included Changjiang County, Dongfang City, Ledong County, Lingshui County, and Sanya City. These isolates were classified into 20 distinct genera (Fig. 6c). In contrast, 293 isolates were collected from 13 sampling sites along the northern coastal areas of Hainan Island, encompassing Danzhou City, Lingao County, Chengmai County, Haikou City, Wenchang City, Qionghai City, and Wanning City. These isolates were subsequently grouped into 40 genera (Fig. 6d). In the southern region, the most abundant genera identified were Monosporascus, Aspergillus, and Fusarium, contributing 29.32%, 12.78%, and 10.53% of the total, respectively. Conversely, Fusarium and Pleosporales dominated the northern region, comprising 26.28% and 23.89% of the total, respectively. Genera commonly isolated across both regions included Alternaria, Aspergillus, Fusarium, Gibellulopsis, Kalmusia, Microsphaeropsis, Monosporascus, Myrothecium, Papiliotrema, Penicillium, Pleosporales, Pyrrhoderma, and Talaromyces. Unique to the southern region were genera such as Acremonium, Curvularia, Eutypella, Perenniporia, Phlebia, Pseudeurotium, and Trichosporon. These results indicate that the distribution of root EF within S. portulacastrum reveals both regional distinctions and overlaps, with southern coastal areas demonstrating a more diverse array of endophytic fungal species.
Screening of salt-tolerant root EF
-
To assess the salt tolerance of root EF, a plate screening method was utilized to examine the responses of 112 isolates under varying NaCl concentrations. The findings revealed that fungal growth was differentially influenced by the salt concentrations applied. Relative to the control group, eight strains (WN-HL-11, DZ-Q1-1, HK-BS-28, WC-FJ-19, WN-HL-13, LG-BZ-9, CJ-HW-15, and CJ-HW-35) demonstrated significantly enhanced growth under saline conditions (Fig. 7a). Following a 7-d exposure to salt stress, the colony diameters of these strains were notably greater than those observed in control, even at a NaCl concentration of 0.75 M (Fig. 7b), highlighting their superior salt tolerance. Specifically, the salt-tolerant strains LG-BZ-9 and WC-FJ-19 exhibited accelerated growth under salt stress, with colony diameters measuring 1.8 and 1.6 times larger than the control, respectively. Although CJ-HW-35 also outperformed the control at 0.75 M NaCl, its growth rate was comparatively slower (Fig. 7b). Furthermore, ten moderately salt-tolerant EF were identified, including WN-HL-14, HK-BS-38, QH-GH-28, DZ-Q2-24, DF-YG-36, LD-YGH-15, DZ-Q1-12, DZ-Q1-5, LD-YGH-37, and DZ-HT-7. These strains displayed enhanced growth in media containing 0.5 M NaCl relative to the control group, though their growth was significantly inhibited at 0.75 M NaCl (Supplementary Fig. S1). Additionally, certain strains exhibited satisfactory growth at lower NaCl concentrations (up to 0.25 M) but were inhibited when exposed to higher salinity levels (exceeding 0.5 M NaCl). These strains, including HK-BS-39, DZ-LT-41, WC-FJ-22, CM-YB-87, WN-SG-28, DZ-Q1-41, DF-YG-38, and CM-YB-53, were classified as average salt-tolerant EF (Supplementary Fig. S2). Lastly, several strains were markedly inhibited under salt stress, even at low salt concentrations. These strains, including WC-FJ-26, HK-BS-6, HK-BS-4, HK-BS-10, QH-GH-20, QH-GH-3, DZ-LT-29, DZ-Q1-13, CM-YB-121, and WN-SG-20, were categorized as salt-sensitive (Supplementary Fig. S3).
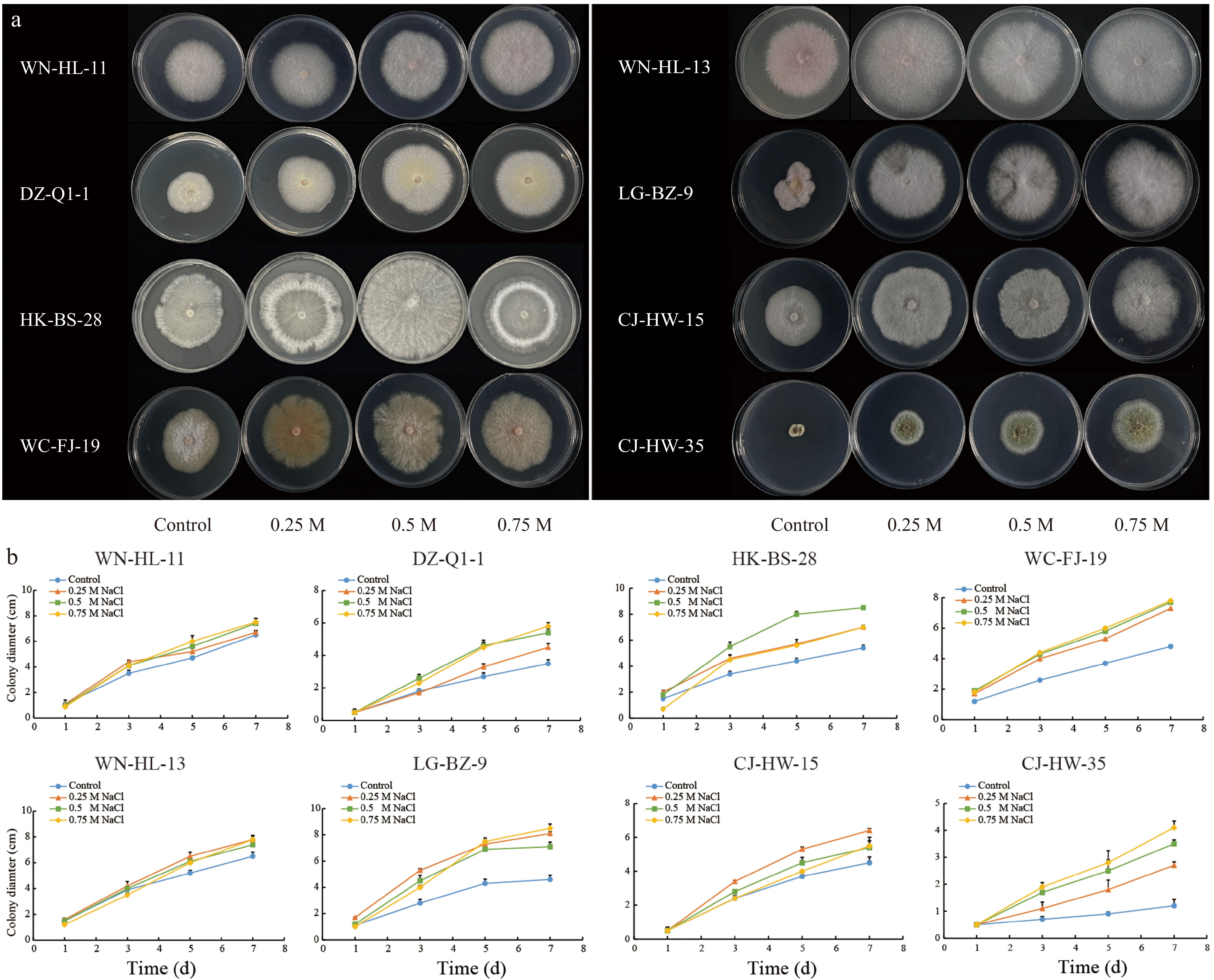
Figure 7.
Screening of salt-tolerant (capable of tolerating 0.75 M NaCl) EF. (a) Phenotypes of eight highly salt-tolerant EF on PDA plates with different salt concentrations. (b) Growth diameter of eight EF strains shown in (a) within 7 d.
LG-BZ-9 strain enhances maize growth under salt stress
-
To evaluate the influence of a salt-tolerant strain on plant growth, strain LG-BZ-9 was selected for an in-depth investigation. A fungal suspension of LG-BZ-9 was applied to irrigate maize seedling roots. Compared to the control group, maize seedlings treated with the fungal suspension displayed significantly enhanced growth, as reflected by increased fresh weight and plant height (Fig. 8a−d). These findings suggest that treatment with EF facilitates seedling growth. When maize seedlings were subjected to 250 mM NaCl, their growth was substantially inhibited, resulting in diminished fresh weight and plant height (Fig. 8c, d), leaf curling (Fig. 8b), and a pronounced reduction in chlorophyll content (Fig. 8e). Nevertheless, under conditions of salt stress, the application of the endophytic fungus LG-BZ-9 effectively mitigated the detrimental effects of salinity on maize seedlings. Specifically, fresh weight, plant height, and chlorophyll content were significantly greater compared to those of maize seedlings exposed solely to salt stress (Fig. 8).
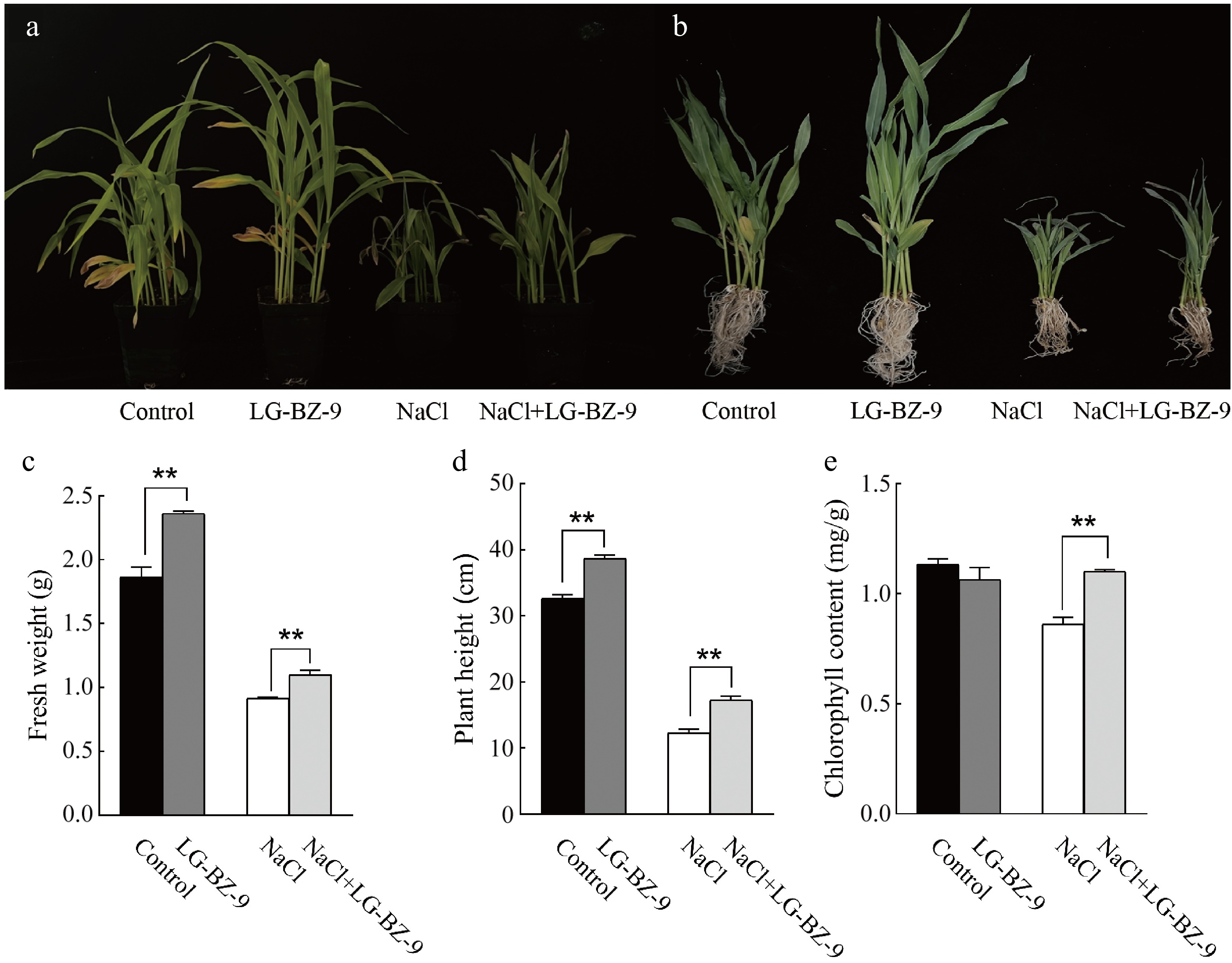
Figure 8.
Salt-tolerant strain LG-BZ-9 enhances salt tolerance in maize seedlings. (a) Phenotypes of maize seedlings in pots under different treatments. (b) Photographs of maize seedlings under different treatments. (c) Fresh weight of maize seedlings under different treatments. (d) Plant height of maize seedlings under different treatments. (e) Leaf chlorophyll content of maize seedlings under different treatments. Control: untreated; LG-BZ-9: maize seedlings treated with LG-BZ-9 suspension by root irrigation; NaCl: maize seedlings treated with 250 mM NaCl; NaCl + LG-BZ-9: maize seedlings treated with salt solution containing LG-BZ-9.
LG-BZ-9 strain enhances maize salt tolerance through the regulation of K+/Na+ balance
-
In comparison to the control group, the application of the fungal suspension resulted in elevated K+ levels and reduced Na+ levels in both the roots and shoots of maize (Fig. 9a, d and Fig. 9b, e, respectively), although the observed differences were not statistically significant (p > 0.05). Under salt stress conditions, K+ concentrations in plant tissues were significantly reduced, whereas Na+ concentrations increased substantially, leading to a markedly lower K+/Na+ ratio compared to the control. Conversely, the introduction of EF under salt stress conditions led to an increase in K+ content and a reduction in Na+ content in both roots and shoots relative to salt stress alone. As a result, a significantly higher K+/Na+ ratio was achieved compared to salt stress conditions without fungal treatment (p < 0.05) (Fig. 9c, f).
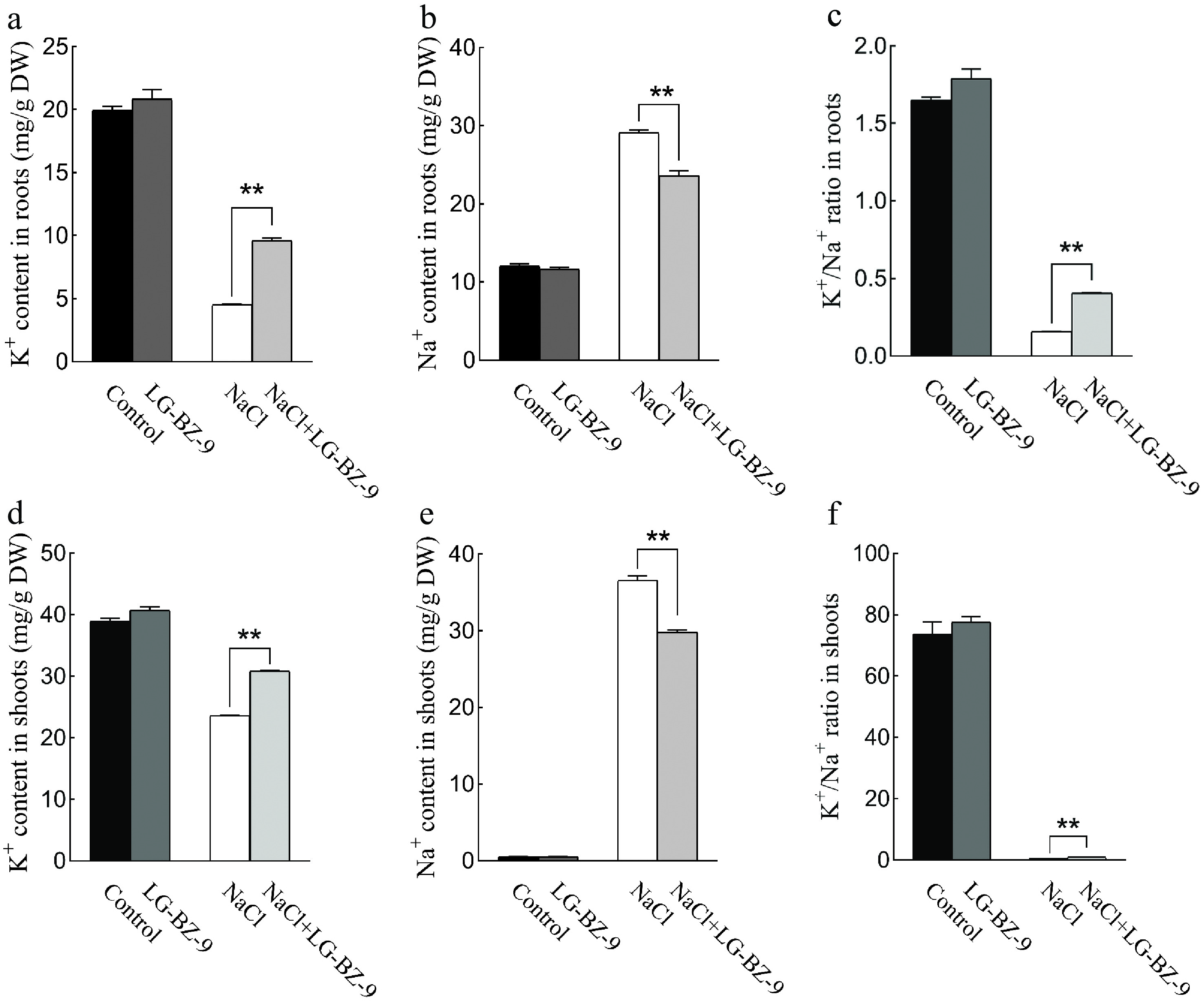
Figure 9.
Ion concentrations in maize seedlings under different treatments. (a) K+ content in the roots of maize. (b) Na+ content in the roots of maize. (c) K+/Na+ ratio in the roots of maize. (d) K+ content in maize shoots. (e) Na+ content in maize shoots. (f) K+/Na+ ratio in maize shoots.
-
In this study, the exceptionally rich diversity of EF is demonstrated. A total of 426 cultivable endophytic fungal isolates were obtained from 1,180 root tissue segments of the halophyte S. portulacastrum. These isolates were categorized into 112 OTUs based on morphological characteristics and sequence alignment analyses, which are distributed across two phyla, eight classes, 16 orders, 37 families, and 47 genera. Among these, 79.5% of the isolates were attributed to Ascomycota, while 20.5% were identified as Basidiomycota, findings that align with earlier studies[41,42]. Yao et al.[41] observed that the majority of identifiable endophytic fungal taxa in the xylem and phloem of Sophorae tonkinensis were Ascomycota (76.6%), followed by Basidiomycota (8.5%) and Zygomycota (4.3%). Similarly, Ikram et al.[42] identified 22 species of EF from 92 samples of three halophytes, with Ascomycota being predominant. Within the 112 OTUs identified here, Fusarium represented the largest proportion at 21.36%, followed by Pleosporales (19.48%) and Monosporascus (15.49%) (Fig. 4b). It has been consistently observed that Fusarium and Pleosporales are dominant fungal species frequently associated with plant species[43,44]. Furthermore, Fusarium oxysporum and Fusarium equiseti have exhibited the potential to mitigate plant salt stress and enhance growth. Bitas et al.[45] and Zhu et al.[46] reported that volatile organic compounds produced by Fusarium oxysporum promote plant growth by modulating auxin transport and signaling pathways, and upregulating enzymes critical to secondary metabolism. Additionally, Feng et al.[47] revealed that Fusarium equiseti produces indole-3-acetic acid and siderophores, which facilitate the growth of perennial ryegrass and enhance its tolerance to salt.
The Shannon-Wiener index, a metric representing community diversity influenced by both evenness and richness[48], was analyzed for culturable EF in S. portulacastrum from 20 sampling points. Significant variations in the diversity and evenness of EF within S. portulacastrum were identified across distinct locations, likely attributed to the saline soil environments. It has been demonstrated in previous studies that environmental conditions can profoundly influence species diversity[49]. Adverse environments, such as saline soils, are known to diminish species diversity, whereas more favorable conditions tend to enhance it[50]. An examination of root EF at varying sampling points identified unique genera at each site: Phlebia at LD-YGH, Polyporales at LG-BZ, Pseudeurotium at SY-YZ, Pseudothielavia at WN-SG, and Sarocladium at HK-BS (Fig. 5). These findings highlight notable differences in endophytic fungal populations across different locations, potentially explained by specific host preferences of certain fungal species[51]. Through comparative analysis of the diversity of root cultivable EF in S. portulacastrum from various coastal regions of Hainan Island, both differences and similarities in fungal distribution were observed. Notably, sampling points in the southern and eastern regions exhibited a higher diversity of endophytic fungal species (Fig. 6). This variation is plausibly influenced by environmental factors. For instance, the southern region experiences a higher average annual temperature compared to the northern areas, while eastern coastal regions, shaped by oceanic conditions and topography, exhibit higher temperatures and increased precipitation relative to the western region[52]. Considering these temperature variations, S. portulacastrum has been shown to possess heat tolerance but limited cold resistance, with more vigorous growth observed in warmer regions[53]. This adaptability likely contributes to the enhanced diversity of endophytic fungal species identified in the southern and eastern regions.
Li et al.[54] isolated and identified a dark-septate endophytic fungus, Laburnicola rhizohalophila, from saline-alkali tidal flats, which was found to significantly enhance root development and salt tolerance in seedlings. This discovery highlights the pivotal role of EF derived from saline-alkali soils in augmenting plant salt tolerance. In the present study, 112 EF were isolated from the roots of the halophyte S. portulacastrum, with Fusarium identified as the predominant genus (Fig. 4b). While Fusarium is widely recognized as one of the primary pathogenic fungi affecting crops, it is important to note that not all species within this genus act as plant pathogens. For example, a strain of Fusarium culmorum isolated from the monocotyledonous plant Leymus mollis was shown to significantly enhance biomass accumulation and improve heat tolerance in rice and various dicotyledonous crops[55]. Moreover, Redman et al.[56] reported that the endophytic F. culmorum enhanced plant salt tolerance, drought resistance, and cold tolerance. Similarly, another study demonstrated that inoculating F. equiseti into barley roots reduced damage caused by Gaeumannomyces graminis to the barley root system while promoting growth, thereby indicating that F. equiseti possesses protective attributes against pathogenic fungi and confers benefits to host plants[57]. The inoculation of endophytic F. verticillioides onto maize seedlings resulted in a marked reduction in the severity of maize smut disease and simultaneously accelerated seedling growth. This finding suggests that endophytic F. verticillioides supports plant growth by interfering with the infection process of the maize smut pathogen (Ustilago maydis DC) and mitigating disease progression[58]. Additionally, the endophytic F. solani strain K was observed to colonize tomato plant roots, providing protection against fungal pathogens while enhancing tolerance to water deficit stress[59].
Jalili et al.[60] demonstrated that lower salt concentrations facilitate fungal growth, whereas elevated salt concentrations inhibit it. The appropriate addition of salt ions has been shown to enhance cellular metabolism in plant tissues and stimulate cell division. However, excessive salt concentrations disrupt the intracellular balance of Na+ and K+, resulting in cell dehydration and eventual death[61]. In the present study, 112 EF isolated from the roots of S. portulacastrum were subjected to varying levels of salt stress. Plate salt tolerance analysis revealed that the growth of the majority of EF was significantly suppressed as the salt concentration increased. This investigation identified eight strains of EF exhibiting superior growth compared to the control under low salt concentration (0.25 M NaCl) (Supplementary Fig. S2), ten strains performing better than the control under moderately high salt concentration (0.5 M NaCl) (Supplementary Fig. S1), and eight strains surpassing the control at high salt concentration (0.75 M NaCl) (Fig. 7). A salt-tolerant strain of Fusarium incarnatum, LG-BZ-9, was selected for further examination. Co-cultivation of this strain with maize seedlings revealed a marked enhancement in both the fresh weight (Fig. 8c) and plant height (Fig. 8d) of maize. These findings indicate that inoculation with EF can influence the biomass of host plants, thereby mitigating the adverse effects of salt stress on maize, consistent with the findings of Bakhshi et al.[62]. K+ is recognized as an essential macronutrient for plant growth and development, playing a critical role in regulating enzymatic reactions and osmotic adjustment[63,64]. Salt stress leads to the excessive accumulation of Na+ in plant cells, hindering K+ absorption and disrupting the Na+/K+ balance, ultimately causing metabolic disorders[63,64]. Maintaining a high K+/Na+ ratio within cells is one of the strategies employed by plants to combat salt stress[65]. When compared to salt stress alone, co-cultivation with the LG-BZ-9 strain resulted in a significant reduction in Na+ content in both the leaves and roots of maize seedlings (Fig. 9b, e), while simultaneously increasing K+ content (Fig. 9a, d), thereby maintaining a higher K+/Na+ ratio (Fig. 9c, f). These results suggest that the salt-tolerant strain LG-BZ-9 enhances salt tolerance in maize seedlings by regulating the intracellular K+/Na+ balance.
-
A total of 112 EF were isolated from the root systems of the halophyte S. portulacastrum. The salt-tolerant strain Fusarium incarnatum LG-BZ-9 was found to not only promote maize growth but also enhance salt tolerance by regulating K+/Na+ homeostasis. The identification of salt-tolerant, growth-promoting EF in this study offers a theoretical foundation for the development of microbial fertilizers aimed at improving crop productivity in saline environments, thereby contributing to the sustainable advancement of agriculture in these regions.
This study was supported by Hainan Provincial Natural Science Foundation of China (318QN189), Jiangsu Provincial Natural Science Foundation of China (BK20241849), the Open Project of Ministry of Education Key Laboratory for Ecology of Tropical Islands, Hainan Normal University, China (HNSF-OP-202303), and the Education Department of Hainan Province (Hnky2021-19, Qhys2022-100).
-
The authors confirm contribution to the paper as follows: study conception and design: Zhou Y, Hu Y; data collection: Wang P, Liu W, Zhang T, Xia H, Li Y, Wang Y; analysis and interpretation of results: Zhou Y, Hu Y, Wang P, Liu W; draft manuscript preparation: Zhou Y, Wang P. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0005)
-
Received 18 October 2024; Accepted 17 February 2025; Published online 19 March 2025
-
A total of 426 cultivable root endophytic fungi were isolated from the halophyte Sesuvium portulacastrum.
These endophytic fungi could be classified into 112 OTU and exhibited a higher diversity across different regions.
Eight highly salt-tolerant strains were screened, among which the Fusarium incarnatum strain LG-BZ-9 not only exhibited a growth-promoting effect on maize seedlings but also demonstrated an enhanced capacity for salt tolerance.
-
# Authors contributed equally: Peng Wang, Wen Liu
- Supplementary Table S1 Information of the sampling point.
- Supplementary Table S2 Isolation of EF from S. portulacastrum at 20 sampling points.
- Supplementary Table S3 Comparison results of 112 OTUs.
- Supplementary Fig. S1 Screening of moderately salt-tolerant EF (capable of tolerating 0.5 M NaCl). (a) Phenotypes of 10 moderately salt-tolerant EF on PDA plates with different salt concentrations. (b) Growth diameters of the ten EF shown in (a) over 7 days.
- Supplementary Fig. S2 Screening of average salt-tolerant EF (capable of tolerating 0.25 M NaCl). (a) Phenotypes of 8 average salt-tolerant EF on PDA plates with different salt concentrations. (b) Growth diameters of the eight EF shown in (a) over 7 days.
- Supplementary Fig. S3 Screening of non-salt-tolerant EF. (a) Phenotypes of 10 non-salt-tolerant EF on PDA plates with different salt concentrations. (b) Growth diameters of the eight EF shown in (a) over 7 days.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang P, Liu W, Zhang T, Xia H, Li Y, et al. 2025. Endophytic fungi isolated from the roots of the coastal halophyte Sesuvium portulacastrum around Hainan Island enhance salt tolerance in maize through regulating K+/Na+ homeostasis. Tropical Plants 4: e009 doi: 10.48130/tp-0025-0005 

Endophytic fungi isolated from the roots of the coastal halophyte Sesuvium portulacastrum around Hainan Island enhance salt tolerance in maize through regulating K+/Na+ homeostasis
- Received: 18 October 2024
- Revised: 27 January 2025
- Accepted: 17 February 2025
- Published online: 19 March 2025
Abstract: Plant endophytic fungi (EF) play crucial roles in enhancing plant resilience to saline conditions. In this study, the halophyte Sesuvium portulacastrum was collected from coastal saline zones adjacent to Hainan Island. Root EF was isolated employing the tissue block method, followed by comprehensive identification and diversity analysis. Salt-tolerant strains were subsequently identified, and pot experiments were conducted to examine their effects on maize growth under saline conditions. A total of 426 cultivable EF isolates were obtained from the roots of S. portulacastrum sampled across 20 distinct locations, categorized into 112 operational taxonomic units (OTUs). Notable differences in EF species distribution were observed across the sampled regions. Screening on potato dextrose agar (PDA) plates supplemented with 0.75 M NaCl revealed that eight EF isolates exhibited significant salt tolerance, with enhanced growth compared to controls. Among these, Fusarium incarnatum strain LG-BZ-9 was shown to not only promote maize growth but also bolster its salt tolerance. Under salt stress conditions, the application of strain LG-BZ-9 led to increased fresh weight, plant height, and leaf chlorophyll content in maize seedlings. Furthermore, a substantial reduction in Na+ concentration within maize roots and shoots was observed, accompanied by an increase in K+ concentration, resulting in a higher K+/Na+ ratio. The EF strain isolated in this research effectively enhanced salt tolerance and stimulated maize growth by modulating ion homeostasis. These findings offer a theoretical basis for leveraging beneficial microorganisms to improve crop salt tolerance and augment yields in saline soils.
-
Key words:
- Salt stress /
- Salt-tolerant endophytic fungi /
- Ion homeostasis /
- Sesuvium portulacastrum