









-
Species from Zinnia L. are predominantly distributed across the tropical and subtropical regions of North and Central America, with Mexico serving as the center of their diversity[1,2]. During the 18th and 19th centuries, Europe introduced several Zinnia species, such as Z. peruviana[3], Z. elegans[4], Z. haageana[3], and Z. angustifolia[3]. Numerous new cultivars have been developed through extensive hybridization and breeding efforts, laying a solid foundation for Zinnia breeding. Today, these domesticated cultivars are globally prevalent and have become staples in parks, botanical gardens, and home gardening[5].
Zinnias are highly valued for their diverse flower shapes, rich color palettes, and broad adaptability. Particularly, dwarf cultivars such as Z. elegans 'Dreamland', Z. elegans 'Double Zahara', and Z. marylandica 'Zydeco™ Fire' are favored for their compact growth habits and dense floral distribution, offering a multitude of options for landscape greening. They are easy to manage and can create a rich visual effect, making them widely used in flower beds, borders, and as potted plants. In contrast, taller cultivars like Z. elegans 'Envy', 'Benary's Giant', and 'Zinderella' dominate the cut flower market, where postharvest preservation techniques are critical for extending vase life and maintaining quality. Beyond their ornamental value, zinnias demonstrate multifunctionality: their high tolerance and accumulation of heavy metals such as cadmium, lead, and chromium position them as potent phytoremediation agents for contaminated soils[6−9], while their abundant bioactive compounds underscore significant medicinal potential[10−13]. However, escalating abiotic stresses—including salinity, drought, and extreme temperatures driven by global environmental changes—are challenging the resilience of conventional cultivars, necessitating the development of varieties with combined ornamental excellence and stress adaptability.
Against this backdrop, the growing market demand for superior Zinnia cultivars faces bottlenecks from an incomplete taxonomic framework, underutilized genetic resources, and inefficient breeding technologies. This review systematically reviews the status of Zinnia germplasm resources and genetic breeding, highlighting taxonomic controversies, genetic mechanisms of key traits, and limitations of current breeding methodologies. By exploring future directions for enhancing stress resistance and optimizing comprehensive traits, this work aims to advance theoretical and practical foundations for Zinnia improvement.
-
The genus Zinnia extends from the southern United States to Chile and Brazil[14], with the highest level of diversity found in Mexico (Fig. 1). The history of the introduction of Zinnia can be traced back to several critical time points, marking its migration from its native America to Europe and eventually worldwide. The earliest known introduction and naming record was from the early 18th century, when de Jussieu brought Peruvian Zinnia from Peru to Europe[15]. Carl Linnaeus initially described this species and classified it in the genus Chrysogonum. He later reassigned it to the newly described genus Zinnia, named in honor of Johann Gottfried Zinn, for his contribution. Linnaeus named this species Z. peruviana, officially recognizing it as the first described species of the genus, thus laying the foundation for the scientific classification of Zinnia[15].
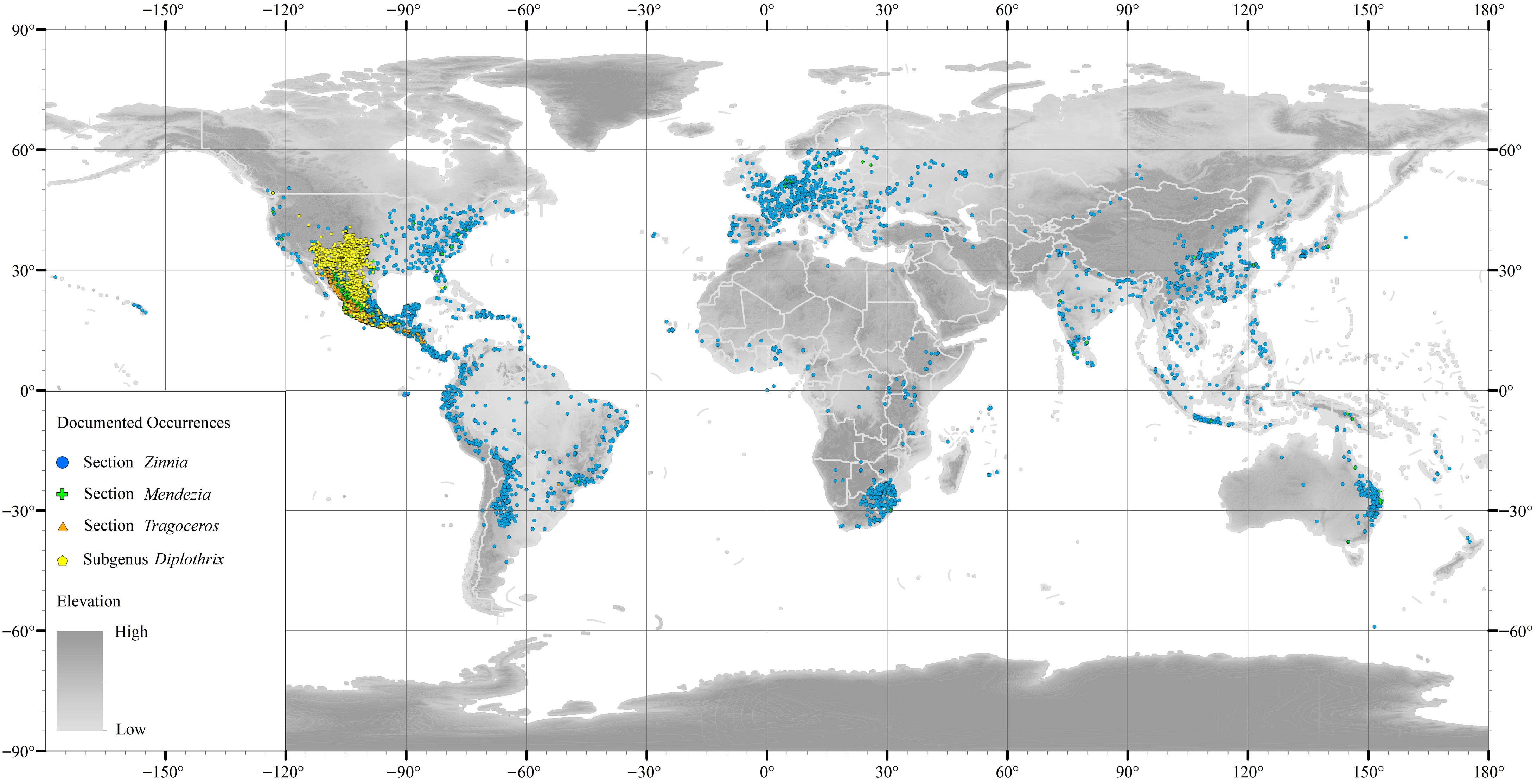
Figure 1.
Geographical distribution of Zinnia species. Dots on the map indicate the locations where Zinnia species were collected. Blue dots indicate section Zinnia, green dots indicate section Mendezia, orange dots indicate section Tragoceros, and yellow dots indicate subgenus Diplothrix. Data sourced from the Global Biodiversity Information Facility (GBIF), accessible at
www.gbif.org . This information is also presented in Supplementary Data 1.Z. elegans (formerly known as Z. violacea), was first introduced from Mexico to Spain in 1790[4]. During the subsequent 19th century, it was extensively introduced to the European mainland. Throughout this period, European breeders systematically selected and hybridized Z. elegans, developing a variety of cultivars in diverse colors, forms, and sizes, significantly enriching the genetic diversity of the genus Zinnia[16]. Consequently, Z. elegans has become the most important species resource within the genus Zinnia[17].
Additionally, with the ongoing research in the horticultural and breeding communities, other Zinnia germplasm resources such as Z. angustifolia and Z. haageana, which were introduced to Europe in the 19th century[3,17], have now been widely distributed across the globe, contributing significantly to the genetic diversity of the genus Zinnia.
The genus Zinnia was initially described as comprising ten species without any subgeneric or sectional subdivisions[18]. Subsequent taxonomic studies integrated the closely related Diplothrix and Mendezia genera into Zinnia[19,20]. This led to a morphological division of the genus into two subgenera, Zinnia and Diplothrix, with a total of 19 recognized species[1], a classification that was later supported by other researchers[4,16]. Olorode & Torres[21] observed hybridization potential between species from the genus Tragoceros and members of sect. Mendezia within the subgenus Zinnia, resulting in the incorporation of Tragoceros into Zinnia as a section within the subgenus Zinnia. The current taxonomic framework, as per Grissell[22], recognizes approximately 24 species within the genus Zinnia.
Despite the establishment of a taxonomic framework through the amalgamation of different taxonomic groups and intra-genus subdivisions, the genus Zinnia continues to evolve as new species may be discovered and existing species may require further nomenclatural clarification and taxonomic revision. It is within this context that the cytogenetic diversity of the genus Zinnia becomes particularly relevant. The chromosome counts in Zinnia species exhibit a broad range of variation, from 2n = 18 to 2n = 84 (Fig. 2), with diploid species predominating, and a minority of tetraploid and octoploid species also identified (Table 1).
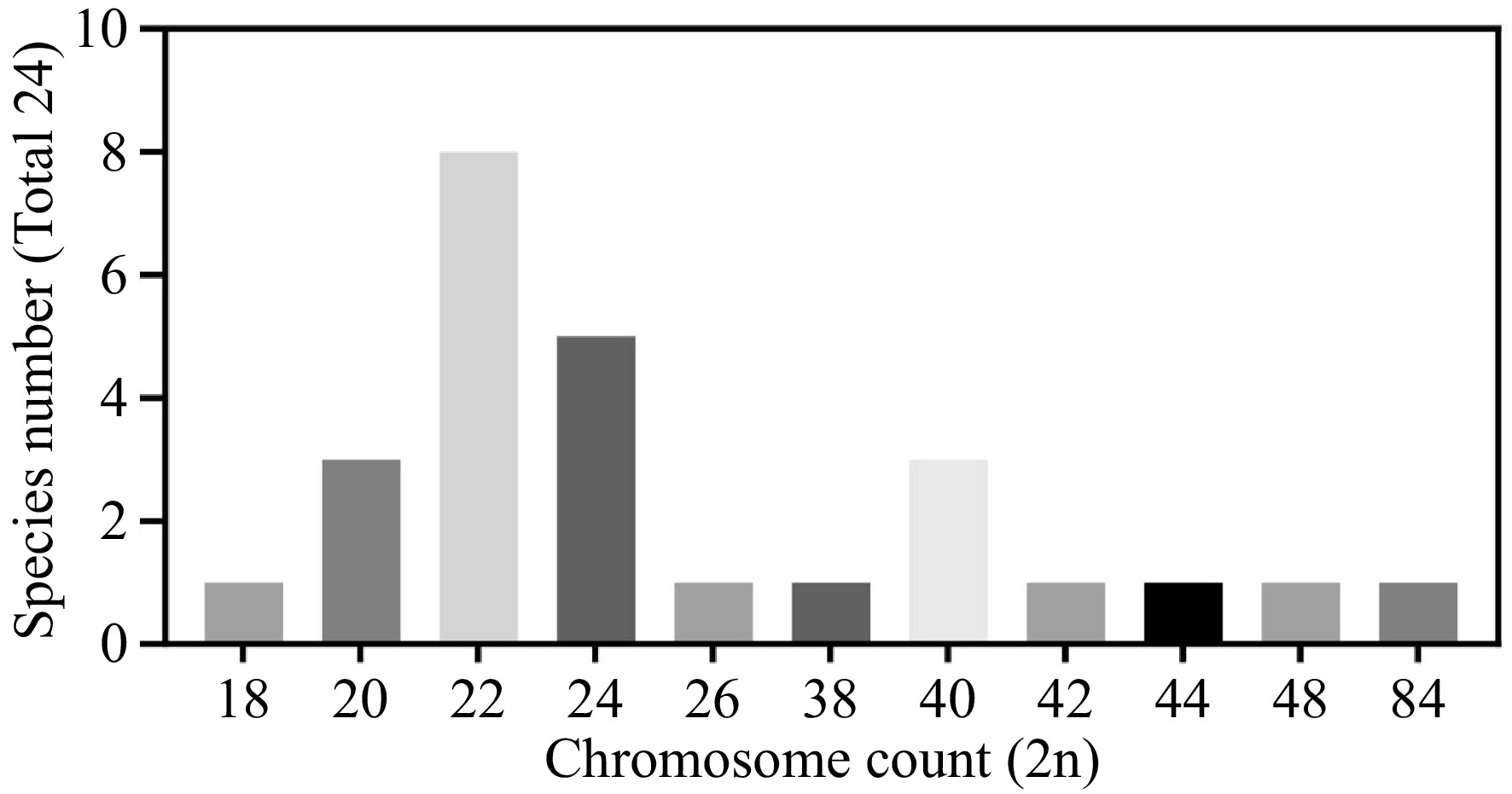
Figure 2.
Chromosome count (2n) distribution of Zinnia. Chromosome count (2n): Represents the number of chromosomes observed in the genus Zinnia. The bar chart illustrates the frequency distribution of different chromosome numbers, with the horizontal axis representing chromosome numbers and the vertical axis representing the frequency of species.
Table 1. Species of Zinnia and their base chromosome count.
Subgenus Subsection Species Polyploidy Base chromosome count[33,34] I. Zinnia Zinnia Z. elegans Jacq. Diploid n = 12 Z. peruviana (L.) L. Diploid n = 12 Z. haageana (DC.)
A. Gray.Diploid n = 12 Mendezia Z. angustifolia Kunth Diploid n = 11, 12 Z. bicolor (DC) Hemsley. Diploid n = 11 Z. purpusii Brandegee. Diploid n = 12,13 Z. tenuis (S. wastson) Strother. Diploid n = 9 Z. maritima HBK. Diploid n = 12, 13 Z. leocoglossa S. F. Blake. Diploid n = 11 Tragoceros Z. americana (Mill.) Olorode. & A. M. Torres. Diploid n = 11 Z. flavicoma (DC.) Olorode. & A. M. Torres Diploid, Tetraploid n = 11, 22 Z. venusta (A. M. Torres) Olorode & A. M. Torres. ? n = ? Z. zinnioides (Kunth) Olorode & A. M. Torres. Diploid n = 11 Z. microglossa (DC.) McVaugh. Diploid n = 11 II. Diplothrix / Z. acerosa (DC.) A. Gray. Diploid, Tetraploid n = 10, 11,
19, 20Z. austrotexana B. L. Turner. Diploid n = 10 + 1 Z. guanajuatensis (Calderón. & Rzed.)
B. L. Turner.? n = ? Z. coahuilana B. L. Turner. ? n = ? Z. citrea A. M. Torres. Tetraploid n = 20 Z. anomala A. Gray. Octoploid n = ± 42 Z. grandiflora Nutt. Tetraploid n = 21, 24 Z. juniperifolia (DC.) A. Gray. Diploid, Tetraploid n = 10, 20 Z. oligantha I. M. Johnst. Diploid n = 10 Z. zamudiana Calderón. & Rzed. ? n = ? Subgenus Zinnia
-
The subgenus comprises 14 species, which are annual or perennial herbs with erect or prostrate stems, ovate leaves, and a bell-shaped to hemispherical calyx with a flat receptacle top. Notably, these species predominantly exhibit base chromosome number n = 11, 12[1], which are more frequently observed than other numbers across the genus (Table 1). The subgenus is further classified into three sections: Zinnia, Mendezia, and Tragoceros[1,4,23].
Sect. Zinnia are annual herbs with erect stems, 90−100 cm in height, flower heads with a diameter of 1 cm, and a ray floret that is enlarged at the base. The achenes are 5.8 cm long, and the ray florets exhibit a variety of colors and petal forms, making the flower heads more vibrant and broader. The plant has sparse foliage with large, ovate to lanceolate leaves, with n = 12[1]. This section includes three species, which, after long-term introduction and popularization, are now widely distributed in the eastern part of North America, Central America, the west coast of South America, as well as western Europe, South Africa, eastern Asia, and the eastern coastal regions of Australia. It is the most widely distributed species in this genus (Fig. 1).
Sect. Mendezia comprises six species with n = 10, 11, 12[1,24−26]. These species may manifest as annual or perennial plants, erect or prostrate, and possess small inflorescences[1]. They are primarily distributed in the southern region of the Mexican Plateau, covering approximately two-thirds of the area (Fig. 1). In contrast to the species within the sect. Zinnia, the species of this section are characterized by a lower latitudinal distribution and a pronounced preference for short-day photoperiods[2]. Additionally, flow cytometric analysis has revealed significant differences in genome size between these two sections[27].
Sect. Tragoceros comprises five species with n = 11[21]. Its distribution extends from Sonora in Mexico to Nicaragua in Central America (Fig. 1). The flowers of the species in this section are small; hence, they are not cultivated as ornamental plants. Some species are considered to have a close morphological association with the genus Sanvitalia[28].
Subgenus Diplothrix
-
This subgenus includes 10 perennial shrubby or subshrubby species, primarily distributed in the temperate arid regions of southwestern United States and northern Mexico (Fig. 1), with n = 10[1]. This subgenus has a higher latitude distribution than the subgenus Zinnia and exhibits flowering characteristics under long-day conditions[2]. Karyotypic studies have revealed that the chromosome morphology of species within this subgenus mainly manifests as diploid, tetraploid, and octoploid (Table 1), with significant homology between some species. For instance, the octoploid species Z. anomala (2n = 8x = ± 84) is believed to have evolved from the tetraploid Z. grandiflora (2n = 4x = 42), and the tetraploid Z. citrea (2n = 4x = 40) is considered to have originated from Z. acerosa and Z. juniperifolia under conditions of geographic isolation[16,29−31]. Additionally, certain tetraploid forms of Z. acerosa (2n = 4x = 40) may have arisen from the diploid Z. acerosa (2n = 2x = 20) through hybridization with other species such as Z. oligantha (2n = 2x = 20)[32]. These findings indicate that interspecific hybridization and genomic recombination events are widespread within this subgenus, and drive species diversity and evolution. These genetic events not only illustrate the mechanisms of species diversity formation in nature but also provide valuable genetic resources for breeding.
Selecting individuals or populations with rich genetic diversity as parents can significantly increase the genetic variability of the hybrid offspring, thereby effectively enhancing breeding efficiency. However, the current assessment of genetic diversity in wild Zinnia remains insufficient.
Cultivated germplasm resources
-
Zinnia species were introduced to Europe in the 18th and 19th centuries[3,4]. Subsequently, in the early 20th century, European countries conducted planned selective breeding for this species, developing a multitude of cultivar resources for the genus Zinnia. To date, over 100 Zinnia cultivars have been successfully developed, featuring a rich array of flower colors including white, cream, green, yellow, apricot, orange, red, bronze, deep red, and purple, along with a selection of bi-color and variegated cultivars (Fig. 3). Additionally, these cultivars exhibit a variety of flower types, spanning from the single type to the more complex dahlia, scabious, and cactus types. In terms of plant height, the cultivars ranged from dwarf cultivars under 6 inches (approximately 15 cm) to tall cut-flower cultivars, reaching up to 3 feet (approximately 0.9 m). From 1932 to 2024, the All-America Selections (AAS) has granted its renowned awards to a total of 891 cultivars, spanning across the categories of flowers and vegetables. Among these awardees, Zinnia has claimed 55 spots, representing a significant 6.9% of the total, with cultivar details provided in Supplementary Data 2. The total number positions Zinnia as the second most recognized genus in the AAS awards, just behind Petunia. The substantial proportion of Zinnia cultivars among the AAS awards not only highlights the remarkable diversity of the genus but also underscores its central role in the horticultural industry.

Figure 3.
Variation of flower color and flower shape of Zinnia. (a)−(h) Flower color: solid type; (i)−(u) bi-color type. (a)−(e), (i)−(u) Flower shape: single to double type; (f) scabious type; (g) cactus type.
Species within sect. Zinnia are valued as important breeding resources owing to their vibrant and eye-catching flowers. The most widely cultivated species, Z. elegans, has been naturalized in parts of several continents since its introduction to Europe in 1796 and subsequent focused selection[16]. Current breeding research primarily focuses on this species, and through long-term selection, numerous cultivars with diverse flower types and colors have been developed. Z. haageana, known for its bi-colored flower cultivars, is an important horticultural resource. Its natural hybridization with Z. elegans may have introduced the bi-color trait into modern cultivars of the latter species. Within sect. Mendezia, Z. angustifolia is valued for its exceptional resistance to diseases. Through artificial hybridization with Z. elegans and subsequent polyploid induction, a new variety, Z. marylandica, with large, brightly colored flowers, and strong disease resistance, was successfully cultivated, further enriching genetic resources for breeding. In contrast, most species of the subgenus Diplothrix have not yet been widely used in horticulture because of their growth habits. Currently, breeding efforts for Zinnia are primarily focused on a few selected species, whereas the potential value of many wild resources remains underexplored.
-
Flower color is one of the most important traits in ornamental plants, and in Zinnia, the color of the flower is primarily determined by pigments located in the upper epidermis of the ray florets[35]. The coloration mechanism involves coordinated regulation of three major pigment systems - anthocyanins, carotenoids, and chlorophyll - along with vacuolar pH[2]. Anthocyanins in vacuoles, through their accumulation levels and pH-dependent interactions, govern pink-to-purple color variations; carotenoids form orange-red hues through specific deposition in chromoplasts; while chlorophyll exhibits cultivar-specific spatial distribution - its enrichment in upper epidermal cells directly determines green phenotypes, whereas in non-green cultivars it is predominantly localized in the spongy tissue of the lower epidermis, forming a pale green base. This stratified distribution pattern may correlate with spatiotemporal regulation of chlorophyll biosynthesis genes such as CHLI (a magnesium chelatase subunit). When both anthocyanins and carotenoids are absent, the presence of chlorophyll becomes the decisive factor in coloration: retained chlorophyll reveals the lower epidermal green base, while simultaneous deficiency of all three pigments results in white ray florets. Genetic studies have revealed the complexity of this system, with the dominant anthocyanin synthesis gene An1 and recessive carotenoid inhibition gene ca serving as major regulatory factors[36], interacting with multiple alleles exhibiting additive effects (e.g., complete dominance of rose-color alleles versus incomplete dominance of vermilion alleles[37]) to form genetic networks.
In Z. angustifolia, the content of flavonoids primarily affects flower color, with pH having a lesser impact[2]. A dominant gene (An2) regulates the presence of orange and yellow pigments[36]. Unlike Z. elegans, carotenoids do not contribute to flower color in Z. angustifolia or tetraploid Z. marylandica[38,39]. In Z. juniperifolia and Z. acerosa, pigment production varies, demonstrating a simple dominant inheritance pattern, with equivalent color phenotypes resulting from different gene combinations[2].
At the molecular level, the metabolic mechanisms of carotenoids and flavonoids influencing the formation of flower color in Z. elegans have received extensive attention. In this species, a total of nine carotenoid components have been detected, the ray florets contain a total of nine carotenoid components. Among these, β-carotene stands out as the predominant compound, representing more than 65% of the total carotenoid content[40]. The lycopene cyclase genes ZeLCYBs and ZeLCYEs, associated with the synthesis of this key carotenoid, facilitate its accumulation by promoting the cyclization reaction[40]. Concurrently, the carotenoid cleavage dioxygenase ZeCCD4-2 is involved in catalyzing the degradation of carotenoids, which in turn regulates pigmentation in the petals[41]. Meanwhile, in Z. elegans, there are four types of acetylated anthocyanins, including two types of cyanidin and two types of pelargonidin[42]. Research on anthocyanin metabolism genes has shown that a 5 bp insertion in the F3H sequence of ivory-white and yellow cultivars leads to a premature stop codon, which is the reason for the lack of anthocyanin accumulation in these cultivars. In rose and red cultivars, the high expression of ZeF3'H is associated with a higher proportion of cyanidin accumulation[43]. At the transcriptional regulation level, the ZeMYB9 transcription factor positively regulates anthocyanin biosynthesis by interacting with ZeF3'H and forms a complex with the bHLH protein GL3 to positively regulate anthocyanin accumulation[43]. In contrast, ZeMYB32 acts as a negative regulator of anthocyanin biosynthesis; its overexpression in tobacco inhibits the accumulation of anthocyanins and alters flower pigmentation. ZeMYB32 competes with ZeMYB9 for binding to ZeGL3, thereby weakening the promotion of anthocyanin synthesis by ZeMYB9 and ZeGL3[44]. These findings preliminarily reveal the molecular mechanisms of flower color formation in Z. elegans and provide potential genetic resources for the improvement of flower color in ornamental plants.
Flower type
-
The double-flowering trait of Z. elegans enhances its ornamental value and is a key characteristic regulated by multiple genes, and the genetic capability of the double-flowering trait is greater than that of the single-flowering trait[45]. Phenotypic expression of this trait primarily depends on the number and arrangement of ray florets. Single-flowering heads consist of only one layer of ray florets, whereas semi-double-flowering heads contain two or more layers of ray florets. In contrast, fully double-flowering heads exhibit multiple layers of ray florets that cover the entire floral disk; however, this usually results in a significant reduction or even complete absence of disc florets. Miyajima & Nakayama[46] confirmed an inverse correlation between the number of ray and disc florets. Additionally, there is a significant correlation between seed morphology and double-flowering traits of plants[47]. Specifically, plants grown from thin-edged ray floret seeds had the highest proportion of fully double-flowering heads and the greatest number of ray florets per head, whereas plants grown from disc-floret seeds had the lowest proportion. This finding suggests that seed selection could increase the proportion of double-flowering offspring by removing some or all the central floret seeds, thereby increasing the proportion of double-flowering heads in the seed batch.
In a study of petal morphology in Z. elegans, Li[48] crossed a tubular petal mutant with a flat wild-type petal, creating backcross (BC1) and F2 populations, revealing that the tubular petal trait in Z. elegans is controlled by a pair of single recessive nuclear genes, with the tubular petal trait being recessive and the flat petal trait being dominant. This is similar to the findings on the tubular petal mutants in sunflowers[49,50]. These findings provide important genetic information for the breeding of petal morphology in Zinnia, aiding the development of new cultivars with specific petal characteristics.
Resistance to disease
-
Z. elegans is particularly susceptible to a variety of pathogens, including Alternaria Zinniae Pape (which causes Alternaria blight), Erysiphe cichoracearum DC. ex Merat (responsible for powdery mildew) and Xanthomonas campestris pv. zinniae (bacterial leaf and flower spot pathogens)[51]. These pathogens can infect plants under both greenhouse and open-field cultivation conditions, leading to decreased ornamental value and plant death. Notably, both Alternaria blight and bacterial leaf spots can be transmitted through seeds, causing significant economic losses to seed producers and commercial growers. Gombert et al.[52] tested the disease resistance of 57 cultivars of Z. elegans over a 17-week period. Only a few cultivars showed good resistance in the first 10 weeks. For example, the 'Ruffles' series exhibited the best resistance to Alternaria blight at week 10, while 'Peter Pan White' and 'Short Stuff Orange' demonstrated notable tolerance to powdery mildew. Researchers have sought to develop new species with enhanced disease resistance,In 1983, Terry-Lewandowski[53] successfully developed a new species, Z. marylandica, through the hybridization of Z. angustifolia and Z. elegans, which exhibits strong resistance to three diseases. The 'Profusion' and 'Zahara' series of this species have gained widespread popularity due to their exceptional disease resistance, increasingly becoming the preferred choice for low-maintenance landscaping over traditional Z. elegans cultivars. In particular, this resistance to powdery mildew was inherited from Z. angustifolia. The genetic mechanisms underlying this resistance are complex and not cytoplasmically inherited. Genes that control resistance in petals and leaves act independently. Subsequent studies clarified this resistance mechanism. Boyle & Wick[54] screened the resistance of the BC1 progeny derived from crosses between Z. marylandica and either Z. angustifolia or Z. elegans to these three pathogens. All BC1 families showed high levels of resistance to Alternaria blight and powdery mildew, with BC1 families from the cross between Z. marylandica and Z. angustifolia also demonstrated high resistance to bacterial leaf spot, while those from the cross with Z. elegans were sensitive to this pathogen. The observed differences in disease resistance among the BC1 progeny underscore the positive impact of Z. angustifolia genomes on their overall resistance. The higher the proportion of Z. angustifolia genomes in the BC1 progeny, the better their resistance to the pathogens tested. However, the absence of such resistance genes in Z. elegans has made it challenging to achieve significant improvements in disease resistance through traditional breeding methods, particularly against powdery mildew. Therefore, exploring new breeding strategies and genetic improvement methods is of great significance to enhance disease resistance in Zinnia.
Adapting to environmental stresses
-
Under the context of climate change, the synergistic threats posed by soil salinization, high-temperature stress, and drought to the growth and development of Zinnia are intensifying, necessitating stress-resistant breeding strategies to balance its ornamental value and economic benefits. Research indicates significant variability in salt tolerance among Zinnia cultivars. For instance, Z. elegans 'Salmon Rose' can produce marketable cut flowers under electrical conductivity (EC) levels as high as 10 dS/m, whereas most cultivars exhibit suppressed dry matter accumulation at EC ≥ 4.2 dS/m[55,56]. This genetic diversity provides a foundation for targeted selection of salt-tolerant varieties and parental hybridization. Furthermore, exogenous polyamines and melatonin have been shown to enhance salt tolerance by regulating ion homeostasis and antioxidant systems, highlighting the potential of hormonal pathway modulation in stress-resistant breeding[57]. Notably, while high-temperature stress induces adaptive responses in Zinnia—such as increased stomatal density, reduced stomatal size, and proline accumulation—the concomitant decline in photosynthetic efficiency underscores the limitations of existing heat tolerance mechanisms[58]. This suggests that future breeding efforts could integrate stomatal regulation with heat-stable photosynthetic systems through hybridization or marker-assisted selection to develop climate-resilient germplasm. Additionally, drought-resistant cultivars like Z. elegans 'Lilliput' which maintain ornamental phenotypes under mild-to-moderate drought conditions due to low transpiration rates[59], offer phenotypic screening criteria for water-efficient variety development. Integrating genomics and metabolomics to identify key genetic components—such as dehydrins and stomatal development regulators—in stress-tolerant cultivars will accelerate the precision improvement of adaptive traits, thereby systematically enhancing the ecological adaptability and agricultural resilience of Zinnia under compound stress environments.
Postharvest quality
-
Postharvest preservation plays a critical role in maintaining the commercial value of cut zinnia flowers. Research on preservative solutions has revealed that acidic conditions (pH 3.0–4.0) enhance vase life by inhibiting bacterial growth, improving water uptake, and reducing air embolisms[60]. However, the interaction between solution electrical conductivity (EC) and cultivar-specific responses requires further exploration. Notably, preservatives combining 8-hydroxyquinoline citrate (8-HQC) and sucrose have shown efficacy, though sucrose concentration must be carefully optimized to avoid floral necrosis caused by extremes[61]. Emerging studies also highlight the potential of biological controls, such as specific bacterial strains, to extend longevity, offering eco-friendly alternatives for future preservative development. Environmental factors significantly influence preservation outcomes. Short-term low-temperature storage (1 °C for 1–3 d) effectively prolongs vase life, yet excessive drying irreversibly damages stem rehydration capacity[62]. Harvest timing is equally critical, with early-harvested flowers exhibiting extended longevity, likely due to preharvest environmental adaptations. Despite progress, systematic insights into the molecular mechanisms regulating Zinnia senescence remain limited. Elucidating key genes and signaling pathways could drive innovations in preservation technologies and quality improvement.
Breeding methods
Cross-breeding
Intraspecies
-
Most species within the genus Zinnia, except Z. peruviana, are naturally outcrossing plants with a low rate of self-pollination in their capitulum inflorescences[21,63−65]. This self-incompatibility trait is beneficial for Zinnia to avoid inbreeding depression and to maintain high genetic diversity[66]; however, it also poses challenges to artificial pollination, increasing production costs. In particular, cultivars with a high degree of double flowering have multiple petal layers covering the stigmas, resulting in inefficient pollination. Moreover, cultivars with fewer double flowers require labor-intensive emasculation[63]. Thus, the floral structural characteristics of Zinnia dictate that their hybrid vigor utilization involves male sterility.
In Zinnia, male sterility is manifested through the degeneration of the ray petal and the transformation of stamens into filamentous structures without pollen, while the pistil develops normally, enabling fertilization and fruiting. This condition represents a complete structural form of male sterilization and is considered an ideal model for hybrid breeding[63]. Male sterile lines can be developed by self-pollination and then repeatedly backcrossed with the parent to transfer traits[37]. In 1964, W. Atlee Burpee Co. utilized male sterility techniques to produce the F1 hybrid Zinnia, developing the cactus-flower variety Z. elegans 'Zenith'[16].
The combining ability between male sterile lines and selfing lines of Zinnia is high. Moreover, F1 plants produced using male sterile biparental lines are more robust than their parents, exhibit greater stress resistance, and demonstrate heterosis in several quantitative traits. Lou et al.[67] found that hybrid vigor is particularly beneficial in Zinnia hybrid breeding, increasing the number of layers and branches in the capitulum while reducing plant height, crown width, pedicel length, and internodal length. Subsequently, Lou et al.[68] conducted studies on the combining ability and heritability of major ornamental traits in selected parents and F1 hybrids and found that plant height, crown width, and internodal length exhibited significant additive genetic effects.
While Zinnia male sterile lines facilitate hybrid seed production and are easily identified through closely linked petal-less traits, the segregation of sterility traits in biparental male sterile lines results in only 50% of the progeny exhibiting male sterility under ideal crosses, which increases production costs[69]. Furthermore, the appearance of bisexual flowers in female lines before flowering can lead to the contamination of inbred and hybrid seeds[70]. Therefore, in Zinnia breeding, research on the genetic control of male sterility, along with the stable production of traits and early selection, is crucial for the effective production of uniform and fertile F1 hybrid seeds.
Currently, there are two hypotheses regarding the genetic regulation of male sterility in Zinnia. Cowen & Ewart[69] suggested that sterility is controlled by three independent recessive genes, whereas Ye et al.[71] proposed that this trait is governed by a single recessive gene causing cytoplasmic male sterility. This discrepancy may result from the observations of different genetic populations and environmental conditions, necessitating further in-depth research to elucidate the genetic mechanisms of fertility. To enhance the propagation of male sterile plants, Rogers et al.[72] explored an axillary bud tissue culture technique to produce clonal lines of sterile plants. This method has been shown to consistently maintain sterility, increase reproductive efficiency, and result in clonal lines that are more compact and flower earlier than their sexual counterparts. Given the segregation of sterility traits in biparental male sterile lines, designing hybrid experiments require strategies to either fix or separate these genes through a combination of different parents to produce the desired male sterile or fertile progeny. The development of molecular markers closely linked to sterility genes in Zinnia could facilitate the early identification of plant fertility during the seedling stage, thereby reducing production costs.
Interspecies
-
Interspecific hybridization plays a crucial role in plant breeding by combining superior genes from different species, thereby enhancing the genetic diversity and resilience of plants[73,74]. To date, 24 hybrid combinations have been attempted within the genus Zinnia, aimed at uncovering phylogenetic relationships and evidence of polyploidy and partly at developing new cultivars. To verify phylogenetic relationships within the genus, Torres[65] conducted hybridization experiments among six species of the subgenus Diplothrix, revealing genomic homology and exploring the complexity of polyploid Zinnia species origins. Olorode[24] artificially hybridized four species within sect. Mendezia, successfully yielding fertile offspring. The segregation of flower color in the F1 hybrids further confirmed the high genomic homology among the species. Ramalingam et al.[75] conducted cytological analyses on the progeny of artificially hybridized Z. angustifolia (2n = 2x = 22) and Z. elegans (2n = 2x = 24), suggesting that Z. elegans likely originated from a hybridization event involving two species, each with 12 chromosomes. This study offers a new perspective on the genetic diversity and adaptability of this species. To develop new cultivars and novel mutations, researchers have attempted hybridization among three species within sect. Zinnia and between certain species of sect. Mendezia. These attempts successfully produced hybrid offspring[76−78]; however, some combinations resulted in sterile progeny and were unable to produce viable seeds using conventional methods.
The most successful interspecific hybrids were those of Z. angustifolia and Z. elegans. Boyle & Stimart[76] hybridized these two species and induced polyploidy in their embryos, creating a new variety that combines traits from both species, named Z. marylandica. Follow-up studies on this hybrid revealed that various traits, such as germination rate, plant height, fresh weight, and flowering days in the offspring, were predominantly controlled by polygenes from Z. angustifolia[38,79]. This indicates that Z. angustifolia contributes significantly to the traits of its offspring, thereby demonstrating considerable potential for breeding improvements. Cultivating and selecting Z. angustifolia with superior traits could lead to even more advantageous progeny in this hybrid combination.
Mutagenesis breeding
-
Mutation breeding, an innovative method for plant improvement, rapidly expands genetic diversity by inducing mutations through physical or chemical means. Recently, researchers have successfully employed radiation sources such as X-rays, gamma rays, and fast neutrons to induce mutations in plants such as Zinnia, study variant traits, and create novel cultivars. For instance, Johnson[80] demonstrated Zinnia's sensitivity to radiation through X-ray treatment, whereas Swarup & Raghava[81] developed a leaf curl-resistant variety using X-ray radiation. A series of studies further revealed the potential of gamma rays to induce variations in Zinnia protein expression, chlorophyll mutations, and changes in morphology and flower color[82−84]. Li et al.[85] research on fast neutron radiation not only stabilized floral traits but also reduced disease susceptibility in Zinnia. Experiments by Pallavi et al.[86] with gamma radiation provided dosage guidelines for achieving specific flower color mutations in Zinnia. Although mutation breeding of Zinnia still has untapped potential, its inherent randomness and the extensive screening required are mitigated by integrating modern biotechnologies such as molecular marker-assisted selection, enhancing screening efficiency, and optimizing the breeding process. The development of more precise mutagenesis techniques and the integration of gene editing with mutation breeding promises to pave new pathways for the genetic improvement of Zinnia.
Heritable variation induced by in vitro culture
-
In the field of horticultural crop breeding, in vitro culture techniques are crucial for inducing genetic variation and enhancing production efficiency.
Tissue culture forms the foundation of in vitro induction, involving the study and optimization of medium components and environmental conditions to ensure the normal development of plant tissues under in vitro conditions. Research on Zinnia species has demonstrated the various advantages of this technology. Stieve et al.[87] investigated the induction of genetic variation in Z. marylandica through tissue culture and its heritability and found that different concentrations of thidiazuron can induce morphological variation, which can be passed on to offspring through sexual reproduction. Cao et al.[88] identified optimal medium formulations for axillary bud induction, proliferation culture, and rooting, providing key technical support for the rapid propagation and breeding of Zinnia. Liu & Wang[89] successfully conducted tissue culture and clone establishment of Z. angustifolia, preserving the genetic characteristics of beneficial variants and providing an effective method for the rapid propagation and genetic stability of the species. Additionally, Stieve & Stimart[90] explored somaclonal variations in Z. elegans, particularly focusing on developing new cultivars resistant to powdery mildew, leaf blight, and bacterial leaf spot through tissue culture and developed stable lines resistant to leaf curl disease through backcrossing and selection.
Polyploid induction is an important strategy for inducing genetic variation in in vitro culture conditions. Chromosome doubling not only enhances plant traits but also increases genetic diversity, thereby enabling the development of cultivars with unique characteristics. In breeding research of Zinnia species, colchicine treatment has proven to be an effective method for inducing polyploidy and has been successfully applied to various species such as Z. elegans, Z. angustifolia, and Z. marylandica[53,91,92]. These polyploid plants exhibit a range of significant phenotypic variations, including notably increased flower size, reduced growth rate, increased leaf thickness, shortened internode length, thicker basal diameter, and significantly increased branching. In addition, polyploid plants displayed changes in leaf morphology, reduced stomatal density, increased stomatal size, larger pollen grains, and higher rates of pollen sterility. Chen[93] further confirmed the existence of triploids, pentaploids, and decaploids in Zinnia species through polyploidy induction research, providing new perspectives and materials for the study of genetic diversity in Zinnia. In practical breeding, tetraploid and diploid plants can be distinguished by observing pollen grain size or guard cell length, thereby providing practical morphological markers for breeding selection. The Z. elegeans 'State Fair' series developed by Ferry-Morse Company represents the first Zinnia variety successfully bred using polyploid induction technology[16], featuring a rich selection of flower colors and resistance to mildew and Alternaria leaf spot, demonstrating the potential and applicability of polyploid induction technology in Zinnia breeding.
Polyploid induction technology also plays a crucial role in restoring the fertility of the interspecific hybrid offspring. For example, colchicine treatment to double the chromosomes of infertile offspring produced from crosses between Z. elegans and Z. angustifolia successfully restores fertility. The application of this technology not only broadens the genetic background of Zinnia cultivars but also provides an effective means for developing innovative germplasm resources.
Furthermore, the performance of Zinnia polyploidy plants in terms of stress adaptation is noteworthy. Studies have shown that polyploid plants generally exhibit enhanced resistance to various stress conditions, including drought, cold, and diseases[94]. Therefore, polyploid induction technology has great potential for breeding new Zinnia cultivars with superior stress resistance.
Molecular Marker-Assisted Selection
-
Molecular Marker-Assisted Selection (MAS) is an efficient selection strategy that integrates molecular biological techniques with traditional plant breeding methods. It leverages genetic markers that are closely linked to target traits to predict and select individuals with desirable genetic variations[95]. This approach allows breeders to rapidly and accurately screen plants carrying target genes or Quantitative Trait Loci (QTLs) during the early stages of growth, thereby accelerating the development of new cultivars and enhancing the precision and efficiency of breeding. In Zinnia breeding, male sterility and flower color characteristics are two crucial breeding targets. Traditionally, assessment of these traits often requires waiting until the plants reach the bud or flowering stage, which extends the breeding cycle. However, with the aid of MAS, breeders can preemptively select individuals with the desired traits at the seedling stage based on molecular marker data. Hu et al.[96] developed five pairs of polymorphic EST-SSR primers that could be used to identify male sterility in Zinnia. Gultom et al.[97] studied the association between Random Amplified Polymorphic DNA (RAPD) markers and Zinnia flower color genes and successfully identified two molecular markers closely linked to the genes controlling flower color. Ye et al.[98] evaluated the genetic diversity of Zinnia inbred lines using RAPD and ISSR markers, and their results indicated that molecular marker technology is more effective and accurate than traditional morphological evaluations for revealing genetic differences and relationships among inbred lines. These studies not only affirm the potential of molecular marker technology in heterosis breeding but also make it feasible to select parents with substantial genetic distances. Despite progress made in the application of MAS in Zinnia breeding, a system for its application has not yet been fully established. The stability and general applicability of existing markers require further validation to ensure their effectiveness in practical breeding. Moreover, the specific positions of genes controlling many complex traits in Zinnia and their associated molecular markers have not been fully elucidated. Therefore, constructing a comprehensive genetic map of Zinnia or conducting Genome-Wide Association Studies (GWAS) on specific genetic populations to identify the locations of critical trait-controlling genes and to develop corresponding molecular markers are key areas of future research. This will not only facilitate the transition from single-trait to multi-trait selection but also pave new paths for the genetic improvement and breeding efficiency of Zinnia.
-
The taxonomic research of the genus Zinnia requires further refinement. To elucidate the evolutionary relationships and genetic diversity among species within the genus, conducting more comprehensive taxonomic and phylogenetic studies is crucial, which is essential for understanding the evolutionary dynamics among species. Despite nearly two centuries of breeding, which has resulted in over 100 cultivars with diverse flower colors and morphologies, the development of genetic resources in Zinnia has primarily focused on a few species, such as Z. elegans, Z. angustifolia, and Z. haageana. However, the research and application of many wild species and cultivars within the genus remain to be further explored. There is a need for improvement in various aspects of Zinnia cultivars. For instance, the susceptibility of Z. elegans to powdery mildew has not been adequately controlled, and there is a market gap between winter-blooming cultivars and those with blue-purple flowers. Therefore, future research in the breeding of Zinnia will focus on several key areas: developing new varieties with unique flower colors to meet the market's demand for diverse ornamental plants; extending the flowering period to enhance aesthetic value; improving the vase life of flowers to increase their commercial potential as cut flowers; enhancing the resistance of plants to mold and bacterial leaf spot diseases to reduce disease-related losses in production; increasing the adaptability of plants to high temperature and humidity environments to expand their cultivation range and enhancing the plant's ability to accumulate heavy metals to promote its application potential in environmental remediation.
The cultivation and production of new Zinnia cultivars primarily rely on hybrid vigor, especially by utilizing male sterility systems to enhance production efficiency and yield. However, improvements in many traits cannot be achieved solely through traditional breeding methods, which emphasizes the necessity of strengthening molecular breeding research. Currently, molecular breeding research on Zinnia, particularly focusing on the exploration of trait formation mechanisms, still has a gap to bridge before actual variety development can be achieved. The use of viral vector-based homologous genetic transformation methods has been successfully tested in Z. elegans and Z. haageana, resulting in the acquisition of transgenic plants[99]. In the future, we can continuously refine the genetic transformation methods for Zinnia and reveal the specific roles of candidate genes in the growth, development, and trait formation of Zinnia by combining overexpression and RNA interference techniques. With an understanding of the mechanisms of trait regulation, we can use gene editing technologies such as CRISPR/Cas9 to achieve precise genetic modifications, thereby creating new Zinnia cultivars with novel traits. Developing genomic data for Zinnia and advancing omics research to annotate and functionally characterize key trait genes will deepen our understanding of the genetic basis of traits such as flower color and disease resistance in Zinnia. Multi-omics analysis and trait data can be used to identify genes and linked markers associated with target traits, thereby facilitating QTL mapping or GWAS in genetic populations. Validated QTL markers can be applied to marker-assisted selection (MAS) or transformed into markers more suitable for breeders to accelerate the breeding process.
This research was supported by the Natural Science Foundation of Zhejiang Province (LY24C150001) and the Scientific Research Fund of Zhejiang Provincial Education Department (Y202456419).
-
The authors confirm contribution to the paper as follows: literature collation: Liu X; study conception and design: Song C, Fu J, Zhang C; data analysis and visualization: Song C, Liu X, Xu M, Ying M; writing and polishing: Song C, Liu X; resources and project administration: Fu J, Zhang C; review and editing: Fu J, Zhang C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Chunlin Song, Xinyue Liu
- Supplementary Data 1 Zinnia GBIF Distribution Data EN.
- Supplementary Data 2 Zinnia AAS Winners data en.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Song C, Liu X, Xu M, Ying M, Fu J, et al. 2025. Germplasm resource and genetic breeding of Zinnia: a review. Ornamental Plant Research 5: e015 doi: 10.48130/opr-0025-0014 

Germplasm resource and genetic breeding of Zinnia: a review
- Received: 15 November 2024
- Revised: 12 February 2025
- Accepted: 03 March 2025
- Published online: 15 April 2025
Abstract: Zinnia species are among the most popular flowering plants worldwide owing to their diverse ornamental characteristics and economically valuable biological traits. Since the 18th century, the identification and selective breeding of wild species has resulted in the development of over 100 cultivars. However, information regarding Zinnia breeding remains highly fragmented, hindering the effective formulation of breeding strategies. This paper provides a comprehensive overview of the germplasm resources of Zinnia, explores the genetic mechanisms underlying key traits such as flower color, flower type, stress resistance, and postharvest longevity, and thoroughly evaluates the current breeding methods and their practical applications. The objective is to establish a robust scientific foundation and propose innovative approaches for future breeding strategies, thereby facilitating the sustained development and advancement of Zinnia in the horticultural sector.
-
Key words:
- Breeding /
- Zinnia /
- Germplasm resource