









-
Due to the rapid use of fossil fuels, the drive to reduce greenhouse gas emissions, and increasing energy security concerns, there is an unprecedented opportunity arising in the development of biofuels. Elephant grass (Cenchrus purpureus) is a well-known C4 perennial forage grass utilized extensively as an energy forage crop across tropical and subtropical regions of the world[1−3]. It's a warm-season grass with an upright height of about 3−5 m and is harvested about four times in one year[1,3]. Moreover, elephant grass also has the potential to be used for the production of electric power, heat, ethanol, and bio-gas, depending on fertilization, management, and variety, making it one of the most promising energy plants[1,4−8]. Elephant grass is highly efficient at absorbing atmospheric CO2[9]. It has been found that elephant grass in the tropical areas can produce 25 to 35 t·ha−1 of dry matter annually[1], while the production of sugar cane and maize is about 21 tons (sugar and bagasse), and 13 tons (grain and straw), respectively[10]. In addition to high biomass, elephant grass is characterized by low water and fertilizer requirements on marginal land[11,12], as well as strong drought[13], and heat resistance[14]. Nevertheless, it is sensitive to cold stress, limiting its application and distribution. Cold stress was found to reduce growth parameters (e.g., leaf area, number of leaves, plant height, shoot fresh weight, shoot dry weight, root fresh weight, and root dry weight), photosynthetic rate, chlorophyll content, and chlorophyll fluorescence in elephant grass[15]. Cold stress reduces the fluidity of plant cell membranes, disrupts protein stability, inhibits enzyme activity, and impacts gene expression and protein biosynthesis[16]. Cold stress has been found to inhibit the growth and photosynthesis of elephant grass; however, the molecular mechanisms underlying its response to cold stress remain unknown. Investigating these mechanisms will aid in developing cold-resistant varieties and expanding the cultivation range of elephant grass.
The agricultural sector is facing continuous and severe challenges in food security and fuel production due to an ever-growing global human population. Cultivating bioenergy plants on marginal farmlands that are not suitable for crop production would help to achieve energy goals while minimizing competition with food production. Most marginal lands are located in temperate or frigid regions, where plants are frequently subjected to extensive abiotic stresses, including soil moisture shortage or excess, insufficient nutrient availability, salinity, and extreme temperatures (both high and low), all of which ultimately cause the yields of elephant grass to fall below their potential[17]. Uncovering the detailed molecular patterns that govern the maintenance of tolerance to cold stress is critical to achieving better bioenergy crop breeding as well as production, and will prove useful in breeding high-yield plant species for marginal lands.
Under cold stress, plants rapidly respond to cold signaling by altering cell membrane fluidity and cell wall stiffness, thereby activating downstream signaling molecules, such as Ca2+ and ROS (reactive oxygen species)[18]. Ca2+ is a crucial secondary messenger in plant response to environmental hazards[19]. A previous study has shown that cold stress-induced alterations in membrane fluidity and Ca2+ levels. Elevated Ca2+ levels elicit calcium-dependent protein kinases (CPKs) and mitogen-activated protein kinase (MAPK)-cascade-associated protein phosphorylation at cold stress[20]. CsCBL-interacting protein kinase 11 (CsCIPK11) enhances cold tolerance in tea (Camellia sinensis) by phosphorylating tau-like glutathione S-transferase 23 (CsGSTU23)[21]. It was reported that SaMKK2 and SaMAPK4/7 interacted to induce the expression of cold tolerance genes to enhance cold tolerance in potato (Solanum tuberosum)[22]. Furthermore, the plant hormone abscisic acid (ABA) also acts as an important player in mediating plant responses to abiotic stresses. Increased ABA synthesis in rice (Oryza sativa) and Arabidopsis under cold stress is accompanied by elevated ABA levels[23,24]. Exogenous ABA spraying enhanced the activities of superoxide dismutase (SOD), catalase (CAT), and ascorbate peroxidase (APX) while reducing membrane lipid peroxidation in melon (Cucumis melo) seedlings under cold stress[25]. These findings suggested that both Ca2+ and ABA were associated with cold stress. However, the mechanism of cold stress affecting Ca2+, ABA, and ABA hormone signaling pathways in elephant grass remains to be explored.
The transcriptome comprises all RNA molecules transcribed in a specific cell, tissue, or organism at a given developmental stage or physiological state. It serves as an effective tool for studying the relationship between gene expression patterns and phenotypes[26]. Transcriptome analysis is an effective technique to investigate the mechanism of plant response to abiotic stresses such as water deficit stress, extreme temperature stress, and salinity stress[27−30]. Transcriptomic and physiological analyses reveal that Chinese cabbage (Brassica rapa) improves drought tolerance by modulating the expression of glucosinolate and ABA signaling pathway genes, as well as by influencing glucosinolate content and antioxidant enzyme activities[31]. In the case of cold stress, transcriptomics has been utilized to explore the response mechanisms in poplar (Populus × canadensis)[32] Chinese jujube (Ziziphus jujuba)[33], wheat[34], and maize (Zea mays)[35]. These studies revealed that the Ca2+ signaling pathway, starch-sucrose metabolism, abscisic acid signaling, DNA repair, proline biosynthesis, and raffinose accumulation play crucial roles in cold stress response.
The objective of the present study was to investigate the transcriptome response of elephant grass under cold stress to explore the association between gene expression and cold stress. The transcriptome is used to uncover signaling pathways and regulatory networks associated with cold tolerance and examine the response mechanisms of elephant grass under cold stress. Characterizing the molecular mechanisms underlying the low-temperature response in elephant grass will facilitate the development of cold-tolerant varieties of elephant grass, particularly for marginal lands.
-
Cenchrus purpureus ('Chuanxu No. 4') stems were grown hydroponically in a growth chamber at 26 °C (12 h light)/23 °C (12 h night) for 10 d with 'huaduoduo' (CL Group Ltd, Israel) nutrient solution. These materials were then divided equally into two parts, one continued to grow in the 26/23 °C (CK treatment) growth chamber and the other part was treated in a 4 °C (12 h light)/4 °C (12 h night) low-temperature incubator for cold stress treatment (CS treatment). Roots were subsequently collected from CK and CS plants at four time points (3, 6, 12, and 24 h). Collected samples were immediately frozen in liquid nitrogen for RNA preparation. Each sample was biologically triplicated for transcriptome analyses.
RNA-seq library preparation and data analyses
-
RNA libraries were prepared using typical Illumina protocols, and RNA-seq was performed using Novogene's NovaSeq 6000 PE150 platform (Beijing, China). Total RNA was prepared from elephant grass roots using Direct-zol™ RNA MiniPrep Kit (Zymo Research Co. Irvine, CA, USA), according to the manufacturer's description. Total RNA was used as input material for the RNA sample preparations. Sequencing libraries were generated using NEBNext Ultra RNA Library Prep Kit for Illumina (NEB, USA, Catalog no. E7530L) following manufacturer's recommendations, and index codes were added to attribute sequences to each sample. The Fastp (version 0.23.1)[36] was used to perform basic statistics on the quality of the raw reads. The data processing steps were as follows: (1) Pairwise reads were discarded if one of the reads contained adapter contamination. (2) Pairwise reads were discarded if more than 10% of the reads contained uncertain bases in each read. (3) Paired reads were discarded if the proportion of low-quality bases (Phred quality < 5) exceeded 50% in any of the reads. The filtered high-quality reads were mapped to the Cenchrus purpureus genome[37]. In this study, a new method for quantifying RNA sequences is established using Kallisto (v.0.46.0) software, which is performed close to the best in speed and accuracy[38,39]. The expression abundance of genes was expressed as transcripts per million (TPM). Finally, differential gene expression level was analyzed using tximport and DESeq R package. The expression levels of DEGs were compared between every two groups based on the TPM values. The software used with the kallisto software for differential gene identification is an R package called Sleuth. Sleuth allows for rapid and accurate counting of genes that are differentially expressed[40]. Genes with expression levels of |log2(CS/CK)| ≥ 1 and adjusted p value < 0.05 were considered as DEGs[27]. Gene Ontology (GO) and KEGG enrichment were performed using the free online data analysis platform OmicShare Tools (
www.omicshare.com/tools ). The GO and KEGG annotated files of elephant grass were downloaded from Milletdb (http://milletdb.novogene.com/api/file/download/Purple/Purple.func )[41].Transcription factor identification
-
To identify transcription factors (TFs) in elephant grass, DEGs were submitted to Milletdb's transcription factor identification website (
http://milletdb.novogene.com/tools/transcriptionfactorid )[41].Weighted gene co-expression network analysis (WGCNA)
-
WGCNA analysis was performed using genes with TPM values greater than 3 in all 24 samples. A total of 11,414 screened genes were analyzed by WGCNA using the WGCNA-shinyApp (
https://github.com/ShawnWx2019/WGCNA-shinyApp ). Subsequently, the module genes were characterized through KEGG enrichment analysis using OmicShare Tools (www.omicshare.com/tools ), a free online data analysis platform.Determination of catalase (CAT) and ascorbate peroxidase (APX) activities
-
The method for measuring CAT and APX enzyme activities was adapted from Hassan et al.[42]. Briefly, approximately 0.1 g of root samples were ground at low temperatures and extracted using 2 mL of phosphate buffer (150 mM, pH 7.8). The extract was then centrifuged at 12,000 g for 30 min at 4 °C, and the supernatant was collected for subsequent analyses.
Determination of proline and ABA content
-
Proline content was measured using a commercial chemical assay kit (Pumoke Biotechnology Company Ltd, Wuhan, China) following the manufacturer's instructions.
ABA extraction was performed following the method of Li et al.[43]. Briefly, 0.1 g of root sample was homogenized and extracted twice at 4 °C for 2 h in a total volume of 4 mL of 80% methanol. The extract was then vacuum-dried at 30 °C for approximately 1 h. The dried residue was dissolved in 1 mL of extractant solution, and ABA content was measured using an ELISA kit (Mmbio, Jiangsu, China).
Construction of transgenic yeast
-
Total RNA was extracted from the roots of elephant grass ('Chuanxu No. 4') using the HiPure Plant RNA Mini Kit (Magen, China). cDNA was synthesized using the HiScript III 1st Strand cDNA Synthesis Kit (+gDNA wiper) (Vazyme, China) according to the manufacturer's instructions. The coding sequences of 9-cis-epoxycarotenoid dioxygenase 1 (NCED: CpB0401574.1), serine/threonine-protein kinase SAPK1 (SAPK1: CpA0601863.1), pyrroline-5-carboxylate synthase 2 (P5CS: CpB0204924.1), and Arginine decarboxylase 1 (ADC: CpB0403764.1) were inserted into the pYES2 vector, the primer sequences are shown in Supplementary Table S1. The pYES2-CpB0401574.1, pYES2-CpA0601863.1, pYES2-CpB0204924.1, and pYES2-CpB0403764.1 plasmids were transformed into Saccharomyces cerevisiae INVSc1, respectively.
Low-temperature tolerance assays of yeast transformants
-
The low-temperature tolerance function of transgenic yeast was verified using the method of Xiang et al.[44]. Briefly, yeast cells of INVSc1 (pYES2-CpB0401574.1, pYES2-CpA0601863.1, pYES2-CpB0204924.1, and pYES2-CpB0403764.1), and INVSc1 (pYES2) were independently cultured in SG-Ura liquid medium (0.67% yeast nitrogen base with ammonium sulfate, 0.077% DO Supplement-Ura, and 2% galactose) at 30 °C for 36 h with shaking at 200 rpm. The cultures were centrifuged at 5,000 rpm for 10 min, and all yeast cells were adjusted to the same density (OD600 = 0.8) with SG-Ura liquid medium. To assess the tolerance of yeast transformants to cold stress, samples were treated at 4 °C for 24 h, followed by serial dilution. Next, 5 μL of the diluted yeast cells was spotted onto SG-Ura plates and incubated at 30 °C to observe the yeast growth rate.
RNA isolation and real-time quantitative reverse transcription PCR (RT-qPCR) analysis
-
The methods of RNA extraction and RT-qPCR were performed according to the method described by Mao et al.[12]. Primers for RT-qPCR were designed using Primer Premier 6.0 software, and their sequences are listed in Supplementary Table S2.
-
To explore changes in transcription in response to cold stress, we characterized RNA-seq with four time points after cold treatments at 4 °C (3, 6, 12, and 24 h). The DEGs were identified by | log2 (CS/CK) | ≥ 1 and adjusted p value < 0.05, as well as paired comparisons between the cold-treated and control groups at the same time points. After cold treatments for 3, 6, 12, and 24 h, a total of 1,480 (680 up-regulated and 800 down-regulated), 4,949 (2,050 up-regulated and 2,899 down-regulated), 8,433 (4,624 up-regulated and 3,809 down-regulated), and 9,005 (4,946 up-regulated and 4,059 down-regulated) genes were differentially expressed, respectively (Fig. 1a). Among them, 273 genes were differentially expressed at all four time points (Fig. 1b). The number of DEGs progressively increased as the duration of cold stress was extended.
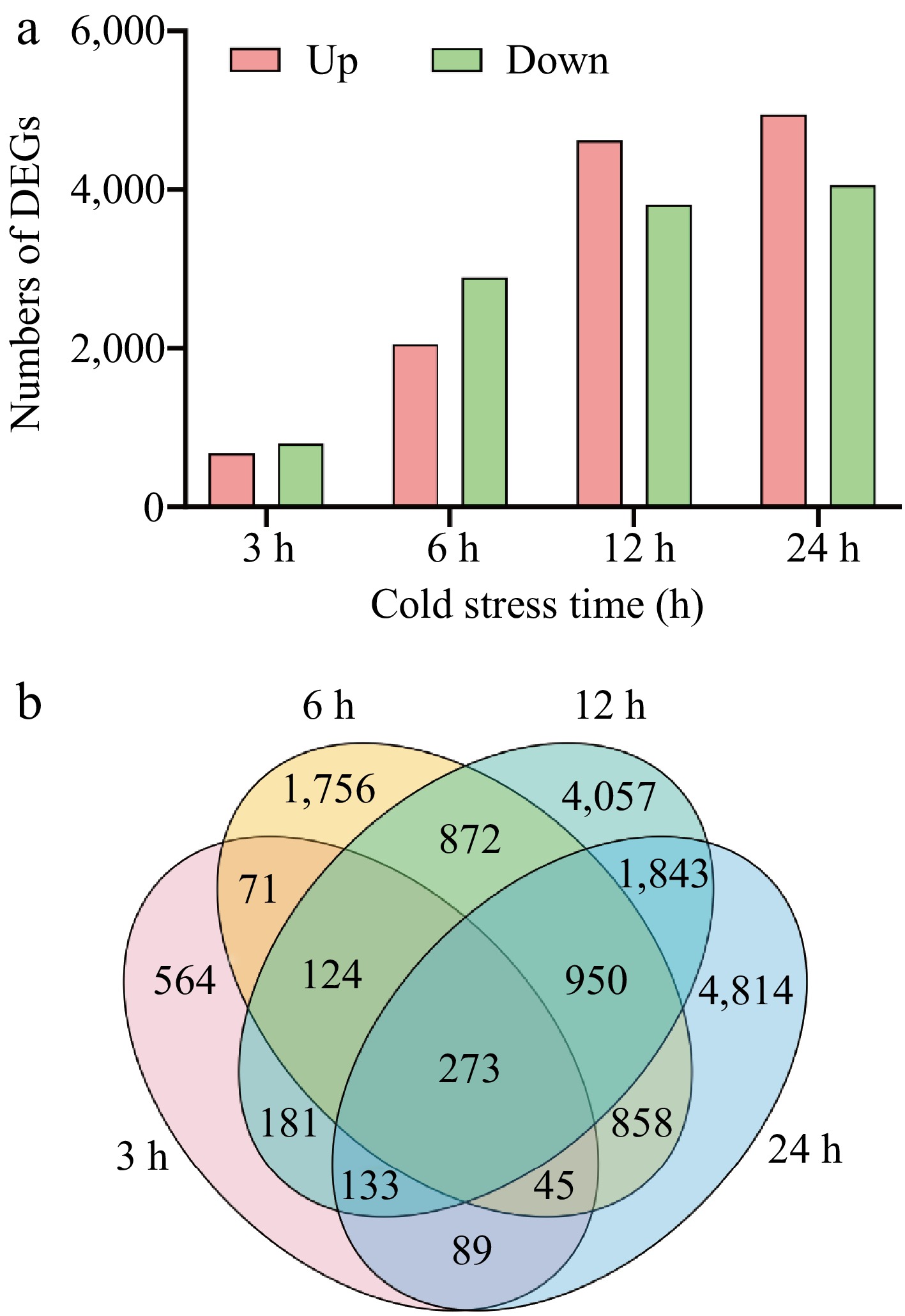
Figure 1.
The numbers of DEGs of elephant grass under cold stress. (a) The number of up- and down-regulated genes at 3, 6, 12, and 24 h; (b) Venn diagram of DEGs.
To estimate whether DEGs are associated with any critical biological relevance, we performed GO enrichment analysis of DEGs. At all cold stress time points, DEGs were predominantly enriched in cellular processes, metabolic processes, single-organism processes, response to stimulus, and biological regulation of biological processes. As for cellular components, DEGs showed enrichment to cell, cell part, organelle, membrane, and organelle parts. Within the molecular function the enriched GO term at cold treatments 3, 6, 12, and 24 h were catalytic activity, binding, nucleic acid binding transcription factor activity, transporter activity, and signal transducer activity, respectively (Supplementary Fig. S1).
Enrichment analysis of the KEGG pathway to determine the biological pathway of cold stress. KEGG profiling revealed that DEGs in the elephant grass root were mainly engaged in plant hormone signal transduction, MAPK signaling pathway, arginine, and proline metabolism (Fig. 2a−d). These pathways may be involved in the response of elephant grass to cold stress.
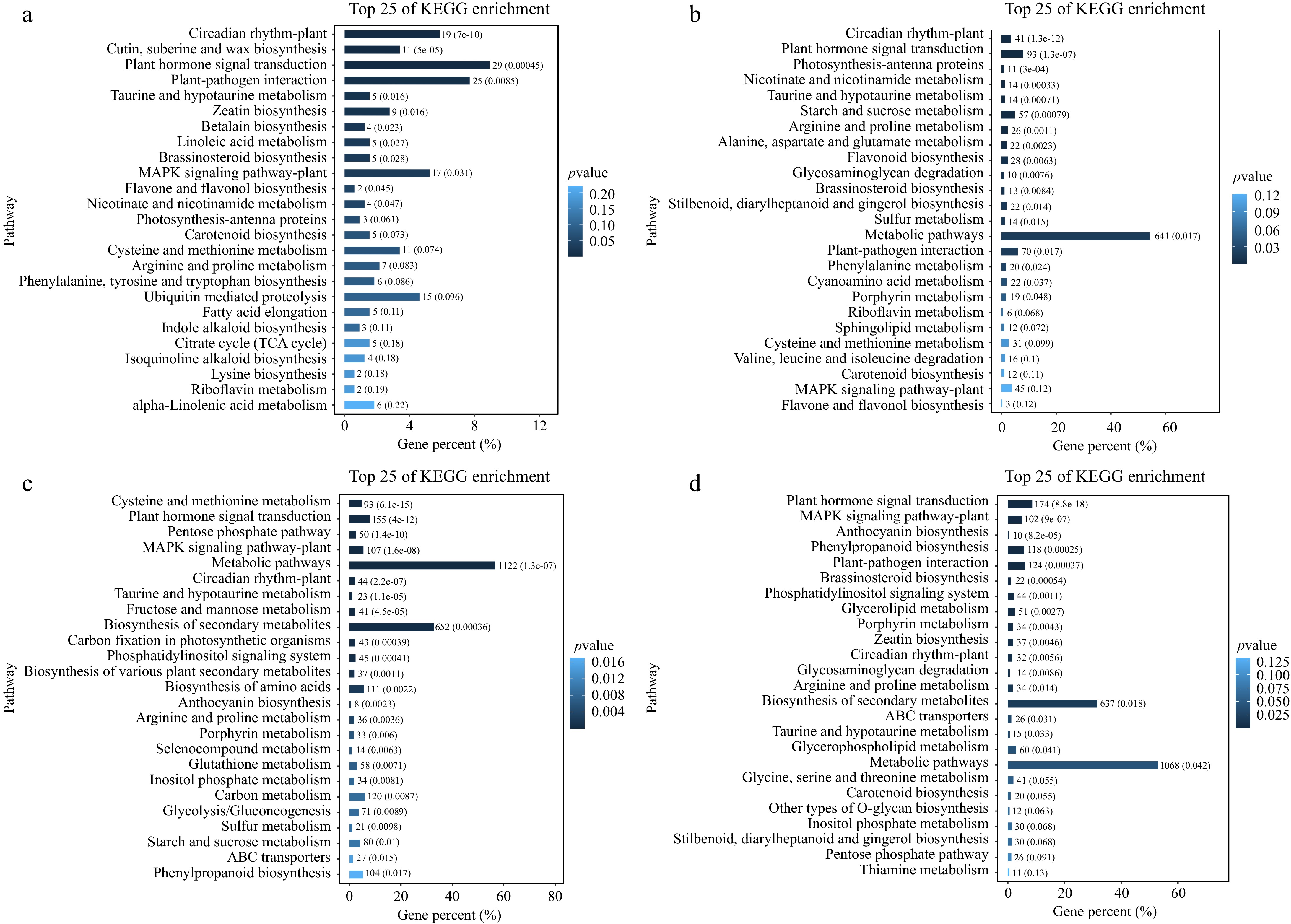
Figure 2.
KEGG pathway enrichment analysis of DEGs during cold treatments. Enriched pathways of cold treatments at (a) 3 h, (b) 6 h, (c) 12 h, and (d) 24 h.
Abscisic acid synthesis and signaling transduction analysis of elephant grass under cold stress
-
Numerous researchers have documented that plant hormones take significant roles in the plant stress response, especially ABA[32,34,35]. NCED (9-cis-epoxycarotenoid dioxygenase gene) is an essential enzyme in ABA synthesis, six NCED genes were up-regulated under cold stress (Fig. 3a, b). Carotenoid biosynthesis is related to the biosynthesis of ABA. It was found that most carotenoid biosynthesis genes crtZ (beta-carotene 3-hydroxylase), ZEP (zeaxanthin epoxidase), NCED, and AAO3 (abscisic-aldehyde oxidase) were up-regulated after cold stress (Fig. 3a, b). This may account for the accumulation of ABA in the root of elephant grass after cold stress. In this research, eight ABA receptors of PYR/PYL (in addition to CpB0105421.1, which was up-regulated at 24 h of cold-stress) were down-regulated (Fig 3c, d). In addition, 13 PP2C (12 up-regulated and one down-regulated) genes were detected by cold stress. Moreover, 15 SnRK2 and 16 ABF genes were differently expressed under cold stress, and most of them (9 of 15 SnRK2 and 10 of 16 ABF) were up-regulated in cold stress (Fig 3c, d).
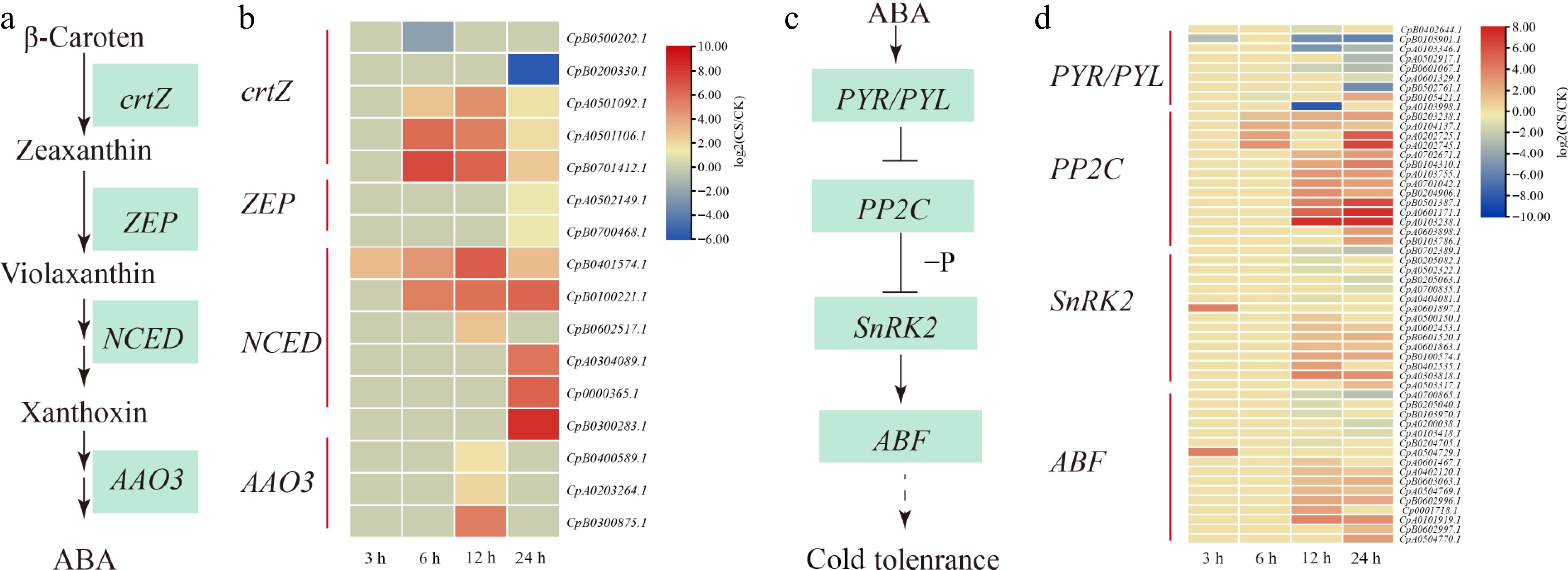
Figure 3.
ABA biosynthesis and ABA signaling pathway in response to cold stress. (a) The ABA biosynthetic pathway. (b) Heat map clustering analysis of ABA biosynthesis under cold stress. (c) The ABA signaling pathway. (d) Heat map clustering analysis of ABA signal pathway under cold stress. '−P' denotes dephosphorylation, arrows indicate promotion, and horizontal lines represent inhibition. Expression data are presented as log2(CS/CK) values, yellow indicates gene upregulation, while blue represents downregulation.
Calcium signaling and MAPK signaling pathway analysis of elephant grass under cold stress
-
Ca2+ is one of the most important secondary messengers responsible for sensing unfavorable environmental conditions[32,33,45]. Many transcripts associated with calcium signaling-related genes exhibited altered expression in response to cold stress, including, five CNGCs (Cyclic nucleotide gated channel), 15 CPKs (Calcium-dependent protein kinases), four CBLs (Calmodulin-like protein), 15 CaMs (Calmodulin), and two CRPK1 (Fig. 4b). It was identified that under cold stress, the DEGs were significantly enriched in MAPK-signaling pathways (Fig. 2). In MAPK-signaling pathways, 11 MAPKKK17/18 (Mitogen-activated protein kinase kinase kinase 17/18), one MKK3 (Mitogen-activated protein kinase kinase 3), one MPK1/2 (Mitogen-activated protein kinase 1/2), and three MEKK1 (Mitogen-activated protein kinase kinase 1) were detected by cold stress (Fig. 4a, b).

Figure 4.
Calcium signaling and MAPK signaling pathway in response to cold stress. (a) The calcium signaling and MAPK signaling pathway. (b) Heat map clustering analysis of calcium signaling and MAPK signaling pathway for cold stress. '+P' denotes phosphorylation, arrow indicates promotion. Expression data are presented as log2(CS/CK) values, yellow indicates gene upregulation, while blue represents downregulation.
Analysis of cold-responsive transcription factors (TF) of elephant grass under cold stress
-
Transcription factors have an influential regulatory role in plant growth and development as well as in response to abiotic and biotic stresses[46]. TFs were involved in the response to cold stress. The top six differentially expressed TFs were MYB, bHLH, AP2/ERF, NAC, bZIP, and WRKY (Fig. 5a). There were 42 TFs (including six AP2/ERF, five MYB, five Dof, three LBD, and so on) differentially expressed under cold treatments 3, 6, 12, and 24 h (Fig. 5b, c). The differential expression of TFs possibly regulates the activation of cold stress-responsive genes in elephant grass.

Figure 5.
Number and classification of TF-encoded genes of elephant grass under cold stress. (a) Number of differentially expressed TF. (b) Venn diagram of the number of differentially expressed TFs at various cold treatments 3, 6, 12, and 24 h. (c) Heatmap of 42 TFs at various cold treatments 3, 6, 12, and 24 h. Expression data are presented as log2(CS/CK) values, yellow indicates gene upregulation, while blue represents downregulation.
Antioxidant oxidase analysis of elephant grass under cold stress
-
Cold stress results in massive accumulation of ROS responsible for oxidative damage to cells, which is attenuated by antioxidant enzymes. Our results revealed that two SOD (Superoxide dismutase), one CAT (Catalase), five GPX (Glutathione peroxidase), and two GR (Glutathione Reductase) genes were significantly up-regulated under cold stress (Fig. 6a, b). Meanwhile, most MDHAR (Monodehydroascorbate reductase, two up-regulated and one down-regulated), APX (Ascorbate peroxidase eight up-regulated and one down-regulated), and GST (Glutathione S-transferase, 21 up-regulated and 12 down-regulated) were also significantly up-regulated in response to cold stress (Fig. 6a, b). However, the transcript level of the DHAR (Dehydroascorbate reductase, one transcript) was repressed after cold stress (Fig. 6a, b).

Figure 6.
The response of antioxidant enzymes in elephant grass under cold stress. (a) The antioxidant enzymes metabolic pathway. (b) Heat map clustering analysis of antioxidant oxidase cold stress. SOD: Superoxide dismutase; POD: peroxidase; APX: Ascorbate peroxidase; CAT: catalase; MDHAR: Monodehydroascorbate reductase; DHAR: Dehydroascorbate reductase: GPX: Glutathione peroxidase; GST: Glutathione S-transferase; GR: Glutathione Reductase; Expression data are presented as log2(CS/CK) values, yellow indicates gene upregulation, while blue represents downregulation.
Arginine and proline metabolism analysis of elephant grass under cold stress
-
Under cold stress, the arginine and proline metabolism were significantly enriched at various cold intervals i.e., 3, 6, 12, and 24 h (Fig. 2). Results showed that six ADC (Arginine decarboxylase), one CPA (N-carbamoylputrescine amidase), and three ODC (Ornithine decarboxylase) genes were significantly induced by cold stress, except for CpB0602089.1 (ADC) and CpB0302029.1 (ODC), which were down-regulated at 6 and 24 h after cold treatments respectively (Fig. 7a, b). PAO (Polyamine oxidase) catalyzes the conversion of spermidine to putrescine. Under cold stress at 6 h, five PAO genes were repressed, whereas at 12 and 24 h, most PAO genes (5 out of 7) were upregulated (Fig. 7a, b). Pyrroline-5-carboxylate synthase (P5CS) is a key enzyme involved in proline synthesis. Results showed that the expression of two P5CS genes were dramatically induced by cold stress (Fig. 7c, d). Moreover, 11 PHA (Prolyl 4-hydroxylase) genes involved in proline metabolism were identified under cold stress, and 10 of these genes were inhibited, which might increase proline accumulation (Fig. 7c, d).

Figure 7.
The response of arginine and proline in elephant grass under cold stress. (a) The putrescine biosynthesis pathway. (b) Heat map clustering analysis of putrescine biosynthesis under cold stress. (c) The proline biosynthesis and metabolism pathway. (d) Heat map clustering analysis of proline biosynthesis and metabolism under cold stress. Arrows indicate promotion. Expression data are presented as log2(CS/CK) values, yellow indicates gene upregulation, while blue represents downregulation.
The activity of CAT and APX, and the content of proline and ABA in the root of elephant grass under cold stress
-
CAT (Fig. 8a) and APX (Fig. 8b) enzyme activities were significantly induced by cold stress at 3, 6, 12, and 24 h. Cold stress induced an increase in proline (Fig. 8c) levels, with significant differences observed at 3 and 24 h of treatment. ABA content increased under cold stress, with significant differences observed only at 12 and 24 h of treatment (Fig. 8d).

Figure 8.
The (a) CAT and (b) APX activity in elephant grass under cold stress. The content of (c) proline and (d) ABA in elephant grass under cold stress. *Represents significant difference (p < 0.05) between CS and CK at the same treatment time, ns represent non-significant differences; and data are presented as mean ± SEM (n = 3).
WGCNA analysis
-
To better understand the relationship between physiological responses and transcriptomic changes under low-temperature stress, we conducted WGCNA analysis on 11,414 screened genes and identified 22 distinct modules (Fig. 9a). It revealed that genes in the black and magenta modules were significantly positively correlated with CAT, APX, and ABA, while genes in the brown module showed significantly negative correlations with CAT, APX, and ABA (Fig. 9b). KEGG analysis revealed that genes in the black module were significantly enriched in the plant hormone signal transduction and MAPK signaling pathways (Fig. 9c), while genes in the magenta module were significantly enriched in the biosynthesis of nucleotide sugars and amino sugar and nucleotide sugar metabolism pathways (Fig. 9d). The brown module genes were significantly enriched in ATP-dependent chromatin remodeling and arginine and proline metabolism pathways (Fig. 9e).

Figure 9.
The results of WGCNA of elephant grass under cold stress. (a) Cluster dendrogram, each leaf in the tree corresponds to a single gene, while the primary branches form 22 distinct modules, each represented by a different color. The x-axis denotes individual genes, and the y-axis reflects co-expression distances. Modules were identified through dynamic tree cutting, which segments the dendrogram at significant branching points. The horizontal bar directly beneath the dendrogram illustrates module assignments, with colors distinguishing different modules. (b) Module–trait relationships, each row represents a module, and each column corresponds to a specific physiological indicator. The color of each cell at their intersection denotes the correlation coefficient between the module and the indicator. Yellow indicates a strong correlation between a given module and the physiological indicator. (c) KEGG enrichment analysis of black module genes. (d) KEGG enrichment analysis of black magenta genes. (e) KEGG enrichment analysis of brown module genes.
Transformed yeast strains with enhanced cold tolerance
-
Cold stress was found to induce the expression of NCED (CpB0401574.1), SAPK1 (CpA0601863.1), P5CS (CpB0204924.1), and ADC (CpB0403764) genes in the roots of elephant grass. To evaluate the roles of NCED, SAPK1, P5CS, and ADC in cold tolerance, we generated yeast cells overexpressing these genes. Under non-cold stress conditions, yeast INVSc-1 transformants (pYES2-CpB0401574.1, pYES2-CpA0601863.1, pYES2-CpB0204924.1, and pYES2-CpB0403764.1) exhibited similar growth to the control INVSc-1 (pYES2) (Fig. 10a), indicating that overexpression of these genes had minimal impact on yeast cell growth. However, under cold stress, the growth differences between yeast INVSc-1 transformants (pYES2-CpB0401574.1, pYES2-CpA0601863.1, pYES2-CpB0204924.1, and pYES2-CpB0403764.1) and the control INVSc-1 (pYES2) became more pronounced as the cell dilution rate increased (Fig. 10b). These results suggest that the overexpression of NCED, SAPK1, P5CS, and ADC significantly enhanced the tolerance of yeast cells to cold stress.

Figure 10.
Tolerance to cold stress of NCED, SAPK1, P5CS, and ADC overexpressing yeast cells. (a) Diluted droplets of yeast transformants growth under control. (b) Diluted droplets of yeast transformants growth under cold stress treatment.
Validation of gene expression by RT-qPCR
-
To validate the RNA-Seq results, eight candidate genes were randomly selected for RT-qPCR analysis. These genes are involved in ABA biosynthesis, ABA signal transduction, arginine, and proline metabolism, antioxidant enzyme activity, and the calcium signaling pathway. The results indicate that the expression patterns obtained by both methods are consistent (Fig. 11), confirming the reliability of the transcriptome data.

Figure 11.
Gene expression as determined by RT-qPCR. * Represents the significant difference (p < 0.05) in gene expression between CS and CK treatments at the same stress time. Student's t-test was used to determine significant differences in gene expression levels, and data are presented as mean ± SEM (n = 3).
-
Cold stress imposes a significant risk to plant growth and development and studying cold stress response has important implications for plant distribution and production. To elucidate the response of elephant grass to cold stress, the present study explored the changes induced in elephant grass under cold stress based on transcriptome analysis.
Effects of cold stress on ABA synthesis and the ABA signaling pathway
-
As an important stress hormone, abscisic acid (ABA) plays a significant role in the cold stress response by regulating a series of stress-related genes[23,24,47]. NCED is considered a key rate-limiting enzyme in the ABA biosynthetic pathway[48]. It was found that NCED genes were significantly up-regulated in Citrus macrophylla[49], maize[35], and Japanese apricot (Prunus mume)[50] under low-temperatures. Overexpressing citrus CrNCED1 lines showed enhanced tolerance to dehydration, drought, salt and oxidative stress compared to the wild type[51]. Six NCED genes were significantly induced in the root of elephant grass under cold stress (Fig. 3a, b). It was found that overexpression of the NCED gene (CpB0401574.1) from elephant grass enhanced low-temperature tolerance in yeast (Fig. 10). Meanwhile, most other genes involved in ABA biosynthesis, like crtZ, ZEP, and AAO3 were up-regulated as well (Fig. 3a, b). Consistently, ABA content showed a significant increase at 12 and 24 h of cold treatment (Fig. 8d). It has been proven that exogenous ABA spray increased the cold stress tolerance in elephant grass[52], apple (Malus hupehensis)[53], bermudagrass (Cynodon dactylon)[54], and wheat (Triticum aestivum)[55]. The enhanced tolerance was mainly associated with reduced oxidative damage, improved leaf water status and photosynthesis, and activation of cold response genes[54,56,57]. Hence, we hypothesized that ABA content increased in the root of elephant grass responding to cold stress. However, the mechanism by which increased ABA content regulates cold tolerance in elephant grass remains to be investigated.
Cold stress up-regulated most SnRK2, ABF, and PP2C genes, while down-regulating ABA receptor (PYR/PYL) genes in maize[35]. Similar results were reported in wheat where cold stress also caused the downregulation of PYR/PYL genes and the upregulation of PP2C genes[58]. These agree with our findings where eight PYR/PYL DEGs and 13 PP2C genes were observed under cold stress of elephant grass, of which seven PYR/PYL genes were down-regulated, and 12 PP2C genes were up-regulated (Fig. 3c, d). SnRK2 and ABF positively regulate cold tolerance[59−61]. Overexpression of SnRK2.3 in Arabidopsis thaliana promoted seed germination under cold stress[62]. In addition, cold stress induced ABF gene expression in Argyranthemum frutescens[63] and pepper (Capsicum annuum)[64]. In elephant grass root, most SnRK2 and ABF genes were up-regulated after cold stress (Fig. 3c, d). Additionally, overexpression of the SAPK1 gene from the SnRK2 gene family of elephant grass enhanced the low-temperature tolerance of yeast (Fig. 10). These results, together, suggested that ABA and the ABA signaling pathway demonstrated an important role in the cold stress tolerance of elephant grass roots.
Effects of cold stress on calcium signaling and the MAPK signaling pathway
-
It has been found Ca2+ can activate the low temperature signaling pathway in plants under cold stress[65]. When plants undergo cold stress, the intracellular Ca2+ level is increased in plant cells via CNGCs[66]. Cold stress significantly induced the expression of CNGCs in rice[67]. Similarly, in our study, cold stress induced the up-regulation of four CNGC genes in elephant grass roots (Fig. 4a, b), potentially indicating that cold stress induced an increase in Ca2+ level. Cold stress caused an increase in Ca2+ level and signaling to the Ca2+ sensors calcineurin B-like protein (CBL), calmodulin (CaM), Ca2+/CaM-dependent protein kinase (CCaMK) and CPKs[68], CPKs, CBLs, and CIPKs show to modulate freezing or chilling tolerance in Arabidopsis[69], rice[70,71], and wheat[72]. This is consistent with our study, where 15 CPKs, four CBLs, and 15 CaM genes were significantly up-regulated under cold stress (Fig. 4a, b), suggesting their involvement in the regulatory network of elephant grass root under cold tolerance.
CPKs sense changes in the level of cytoplasmic Ca2+ and influence with downstream signaling molecules, including plant hormones and MAPKs, resulting in cold stress adaptations in various plant species[73−76]. It was previously shown that the MEKK1-MKK2-MPK4/MPK6 cascade participated in the positive regulation of cold response and freezing tolerance[77]. Our results revealed that two MEKK1 were up-regulated while one MEKK1 was down-regulated in elephant grass root under cold treatment (Fig. 4a, b). MAP3K17 and MAP3K18 belong to a group of genes regulated by the ABA core signaling module, leading to the long-term activation of MAPKs by MKK3, and enhanced ABA-dependent responses under stress conditions[78]. After cold stress, the MAPKKK17/18, MKK3, and MPK1/2 were significantly induced in the root of elephant grass (Fig. 4a, b). ABA and MAPK cascade may co-regulate cold stress in elephant grass roots. In summary, Ca2+, ABA, and the MAPK signaling pathways may be cooperatively involved in the regulation of cold tolerance in elephant grass.
Effects of cold stress on TFs
-
Transcription factors (TFs) are important in cell differentiation, plant development, and adaptation to environmental changes via regulation of gene expression[79]. A large number of TFs of NAC, WRKY, ERF, MYB, and C2H2 were found to be up-regulated in cold-tolerant peanut (Arachis hypogaea)[80], and rice[81]. Our study demonstrated that the genes encoding most of the TFs were involved in the low-temperature response, and primarily belong to the MYB, bHLH, AP2/ERF, NAC, bZIP, and WRKY families (Fig. 5a). In this study, 42 TFs were differentially expressed at all time points under cold treatments, among which five MYB, four AP2/ERF, four Dof, three bHLH, two NAC, and two C2H2 were significantly induced (Fig. 5b, c). We speculated that these TFs likely play an important role in cold stress regulation. However, their detailed functions in cold stress of elephant grass need to be further characterized.
Effects of cold stress on antioxidant enzymes
-
Cold stress disrupts the electron transport chain, induces membrane lipid peroxidation, and disturbs the antioxidant system, ultimately leading to the accumulation of ROS[82,83]. ROS homeostasis can protect elephant grass from oxidative damage resulting from low temperatures. Consistent with the generation of ROS, the antioxidant system controls the cellular redox regulatory network. SOD catalyzes the conversion of superoxide anions into hydrogen peroxide and H2O[84]. SOD, CAT, and APX were enhanced in cold-tolerant cultivar rice under chilling stress[85]. It was revealed that cold stress induced the up-regulation of GPX and GST genes in rice[47]. In this research, SOD, CAT, GPX, GR, and most of APX and GST were up-regulated in cold treatments of elephant grass root (Fig. 6a, b). Accordingly, the enzyme activities of CAT and APX also significantly increased (Fig. 8a, b). These changes may be related to the enhanced ROS scavenging and thereby, increased cold stress tolerance in elephant roots.
Effects of cold stress on arginine and proline metabolism
-
In plants, arginine is an essential nitrogen storage nutrient, as well as being a precursor for polyamine (PAs) and nitric oxide (NO) biosynthesis. It has been recorded that cold stress significantly induced crucial enzymes related to arginine catabolism in tomato, including arginase, ADC, and NOS (nitric oxide synthase)[86]. Overexpression of ADC enhanced freezing tolerance in potato (Solanum tuberosum)[87] and drought tolerance in Arabidopsis[88]. Similarly, in our study, overexpression of the ADC gene from elephant grass enhanced the low-temperature tolerance of yeast (Fig. 10). Furthermore, six ADC were significantly up-regulated under cold stress in elephant grass (Fig. 7a, b), contributing to the accumulation of putrescine. An exogenous spray of putrescine has been reported to enhance salt tolerance in chickpea (Cicer arietinum)[89] and drought tolerance in alfalfa (Medicago sativa)[90].
Osmotic disruption is a widely observed consequence of cold stress, as low temperatures impair water uptake and cellular homeostasis, leading to osmotic imbalance[91,92]. To counteract this, plants accumulate osmolytes such as proline and putrescine, which help stabilize osmotic pressure, maintain membrane integrity, and protect macromolecules from damage[93,94]. Proline accumulation has been a common response to abiotic stresses in plants, including cold stress. P5CS is one of the key enzymes in the synthesis of proline biosynthesis via glutamate precursors[95]. Genes involved in proline synthesis, like P5CS, PDH1, and P5CDH were significantly up-regulated in cold-tolerant citrus under cold stress[49]. The P5CS was also upregulated under cold stress[35]. In our study, six P5CS were up-regulated while 10 PHA catalyzing proline catabolism were down-regulated, which corresponded to the significant increases in proline contents after 3 and 24 h of cold treatment. Moreover, overexpression of the P5CS gene from elephant grass was found to enhance cold tolerance in yeast (Fig. 10). Osmotic stress is a common feature of salt, drought, and cold stresses, as these conditions disrupt water balance and cellular homeostasis[35,96]. Since putrescine has been shown to enhance osmotic adjustment under salt and drought stress[93,94], its accumulation may similarly contribute to cold tolerance in elephant grass by stabilizing cellular osmotic balance and protecting cellular structures. Collectively, these suggest that accumulation of putrescine and proline probably reduced damage in elephant grass by improving the osmotic adjustment under cold stress.
-
In this work, transcriptome analysis was used to investigate the possible pathways of cold tolerance in elephant grass. We proposed a model in which cold tolerance of elephant grass roots was co-regulated by Ca2+, antioxidant enzymes, osmotic substances, ABA signaling, MAPK cascades, and TFs. Cold stress induced an increase of Ca2+ and ABA, which triggered the Ca2+ signaling pathway, ABA signaling pathway, MAPK cascade, and TFs to respond to cold stress in elephant grass. The pathways work together to form a network that regulates cold tolerance in elephant grass. The present study demonstrated the molecular mechanism of elephant grass in response to cold stress, and provided gene resources for the breeding of cold-tolerant cultivars of elephant grass in the future.
Supported by the Fundamental Research Funds for the Sichuan Province Institute of Animal Husbandry Research (SASA202106).
-
The authors confirm contribution to the paper as follows: experiments conception and design: Hu Y, Mao C, Huang L; data analysis: Sun M, Mao C, Luo D; plant material culturing: Yang D, Jia J, Ding Q, Zhang Y; language modifying of the manuscript: Najeeb A, Liang X; manuscript wrting and revision: Wang X, Ji Y, Zhang J, Liu Y, Huang L, Mao C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files. The raw sequence reads were deposited in the NCBI SRA database under Accession no. PRJNA1007572.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Yuanbin Hu, Chunli Mao
- Supplementary Table S1 Primers used in cloning.
- Supplementary Table S2 Primers used in RT_qPCR.
- Supplementary Fig. S1 Gene Ontology (GO) enrichment analysis of DEGs at different cold treatments time. The DEGs enriched at 3 h (a), 6 h (b), 12 h (c) and 24 h (d) under cold stress.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Hu Y, Sun M, Luo D, Yang D, Zhang Y, et al. 2025. Transcriptome profiling the response of elephant grass (Cenchrus purpureus) root to cold stress. Grass Research 5: e014 doi: 10.48130/grares-0025-0011 

Transcriptome profiling the response of elephant grass (Cenchrus purpureus) root to cold stress
- Received: 02 December 2024
- Revised: 06 March 2025
- Accepted: 13 March 2025
- Published online: 06 May 2025
Abstract: Elephant grass (Cenchrus purpureus) is a significant perennial energy and forage plant, characterized by high biomass, low nutrient needs, and resilience to adverse conditions. However, its sensitivity to cold limits its yield and utilization in agriculture. Understanding the cold stress responses of elephant grass is crucial for breeding cold-tolerant varieties. To investigate this, a multi-temporal transcriptome analysis was conducted, revealing that approximately 1,480, 4,949, 8,433, and 9,005 genes were differentially expressed after cold treatments for 3, 6, 12, and 24 h, respectively. The differentially expressed genes (DEGs) were significantly enriched in pathways related to plant hormone signaling, MAPK signaling, and arginine and proline metabolism. KEGG enrichment analysis indicated that these pathways play a vital role in the cold stress response of elephant grass roots. Several transcription factors (TFs), including MYB, NAC, WRKY, bHLH, bZIP, and AP2/ERF, were also implicated, with five MYB, four AP2/ERF, four Dof, three bHLH, two NAC, and two C2H2 TFs significantly up-regulated during cold stress. Additionally, genes encoding antioxidant enzymes (SOD, CAT, GPX, APX, GST) and those involved in the synthesis of putrescine (ADC) and proline (P5CS) contributed to the cold stress response. This study provides valuable insights into transcriptome networks and potential gene resources for breeding cold-tolerant elephant grass.
-
Key words:
- Transcriptome /
- Cold stress /
- Elephant grass /
- Plant hormone /
- MAPK pathway