








-
Competition between crops and weeds for soil resources is a pervasive challenge in agriculture, often necessitating the excessive use of herbicides to ensure optimal crop growth. With advancements in technology, the market share of amino acid-based organophosphorus herbicides has rapidly increased, establishing them as the leading group of herbicides[1,2]. Among these, glyphosate (a type of herbicide with amino and phosphate functional groups) dominates the market[3,4]. Chemically known as N-phosphomethyl glycine (C3H8NO5P), commercial forms of glyphosate include products such as Roundup and glyphosate amine salts.
Typically found in salt form, glyphosate exhibits stable chemical properties and is highly water-soluble. It is a post-emergent, non-selective systemic herbicide[5], which functions by inhibiting the enzyme 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS). This inhibition disrupts the biosynthesis of essential aromatic amino acids, including tyrosine and phenylalanine, ultimately halting protein synthesis and leading to plant death[6].
Glyphosate herbicides are typically applied through spraying, after which the chemical enters the soil system either via direct contact or through water runoff from irrigation and rainfall. Once in the soil, glyphosate is primarily degraded through two main pathways. The first pathway involves the breaking of carbon-nitrogen (C-N) bonds, producing aminomethyl phosphonic acid (AMPA), which is subsequently mineralized into small molecules such as methylamine, formaldehyde, and phosphoric acid, ultimately yielding carbon dioxide and ammonium. The second pathway relies on the breaking of the carbon-phosphorus (C-P) bond, resulting in the formation of sarcosine and phosphoric acid, which are further degraded into glycine and formaldehyde[7,8].
Although the degradation percentage of glyphosate in topsoil is relatively high, its cumulative effects and ecotoxicological impact can still cause varying degrees of environmental harm. Research conducted by the European Chemicals Agency has shown that glyphosate negatively affects the survival of aquatic organisms[9,10]. Furthermore, in agricultural soils with prolonged glyphosate application, concentrations ranging from 270 to 690 μg/kg have been detected[11]. Studies have shown that earthworms, widely regarded as key indicators of soil health, are adversely impacted by long-term glyphosate use, resulting in decreased biomass, density, species diversity, and activity[12]. Research also indicates that soils without glyphosate application support significantly higher earthworm biomass compared to glyphosate-treated soils[13]. In addition, glyphosate has been shown to influence microbial activity associated with soybean roots, ultimately inhibiting plant growth[14]. Furthermore, it alters the composition of soil microbial communities, indirectly impacting soil nutrient availability.
Mycoremediation refers to the use of fungi to degrade or extract pollutants from the environment[15,16]. Several fungal genera, such as Aspergillus, Fusarium, Mucor, Penicillium, Scopulariopsis, and Trichoderma, have demonstrated potential for glyphosate degradation[8,17]. However, most studies in this field have been conducted under laboratory conditions. Agaricus subrufescens, a basidiomycete from Agaricaceae, produces edible and medicinal basidiomata known for their anti-tumor, antioxidant, and hypoglycemic properties[18]. As a thermophilic and oxygen-loving secondary decomposer fungus, A. subrufescens is a prolific producer of laccase, making it a promising candidate for soil pollutant removal. It also colonizes substrates previously processed by other microorganisms with relative ease[19].
Previous research has demonstrated that the cultivated substrate of A. subrufescens can degrade the herbicide Metsulfuron-methyl[20]. Additionally, spent A. subrufescens compost can be repurposed as a growing fertilizer for crops, further enhancing agricultural productivity[21]. Under laboratory conditions, A. subrufescens mycelium was found to degrade 44.68% of the insecticide cypermethrin within 15 d, suggesting its potential as a sustainable solution for restoring agricultural soils[22]. Given its cultivation characteristics and enzyme production, A. subrufescens is an ideal subject for further research. However, the enzyme and microbial community mechanisms underpinning the in-situ compost-based mycoremediation process remain poorly understood and require further investigation.
Fungal enzymes such as dehydrogenase (DHA) and fluorescein diacetate (FDA) hydrolase are key indicators of soil degradation performance and microbial bioactivity. The remarkable potential of fungi in bioremediation lies in their extensive mycelial networks and powerful extracellular enzyme systems, including laccase (Lac), manganese peroxidase (MnP), and lignin peroxidase (LiP)[15]. These enzymes exhibit low substrate specificity, enabling them to transform and metabolize persistent organic pollutants such as polycyclic aromatic hydrocarbons. Due to the non-specificity and broad-spectrum capabilities of fungi and fungal enzymes, they play a crucial role in bioremediation by altering the soil microbial community structure[23]. Glyphosate content in soil can lead to shifts in microbial composition and diversity while also affecting soil nutrient availability. Microbial degradation is one of the most effective strategies for reducing glyphosate levels in soil, with microbial community activity and structure playing a central role in its dissipation[24].
In this study, we aim to investigate the benefits of mycoremediation by analyzing the glyphosate degradation percentage, key enzyme activity, and microbial community composition following the application of fungal inoculation and compost in a glyphosate-contaminated coffee plantation. We found that inoculation of A. subrufescens and addition of compost substrate promote the degradation of glyphosate by boosting key enzyme activity and altering microbial community composition. This study provides a novel perspective for the bioremediation of glyphosate-contaminated soil.
-
The experimental site is located in Nandaohe village, Nanping town, Pu'er, China (22°46′ N, 100°58′ E; elevation: 1,064 m), one of the earliest and most densely planted coffee-growing areas in China.
The climate data for the site were collected through a meteorological instrument, and the record shows a maximum temperature of 36.2 °C and a minimum temperature of 2.4 °C, with an annual average temperature of 19.9 °C. The monsoon season spans May to October, bringing an annual rainfall of 1,385.5 mm and an annual relative humidity of 75%. These favorable conditions, characterized by abundant sunlight, heat, water resources, and high soil organic matter content, make Nandaohe village an ideal location for coffee cultivation.
Fungal strain and preparation
-
The Agaricus subrufescens strain used in this study was obtained from the Culture Preservation Center at the Kunming Institute of Botany, as described by Hu et al.[22]. The strain stock, stored at −80 °C, was revived and cultured for further use. Malt Extract Agar (MEA, 50 g/L) was used as the growth medium. All inoculation tools were sterilized by flame on a sterile mycology workbench to ensure aseptic conditions. After cooling, small mycelial fragments were taken from the mother culture and transferred onto the MEA medium using a sterilized inoculating spatula. The medium was then sealed with parafilm and incubated at a constant temperature of 28 °C.
For purification, portions of the mycelium were carefully selected using a sterilized syringe needle and transferred to a fresh MEA medium. The A. subrufescens mycelium underwent 1–2 rounds of purification to ensure culture purity and mass production of fungal hyphae.
The purified A. subrufescens mycelium, grown on MEA medium, took approximately 15–20 d to fully colonize the petri dishes in a constant-temperature incubator. Once fully colonized, the mycelium was cut into small pieces on a sterile workbench using a sterilized knife. These mycelial fragments were then transferred into the sterilized wheat-filled bottles and incubated at 28 °C. It took approximately 28–33 d for the mycelium to completely colonize the wheat substrate. Aseptic techniques were carefully maintained throughout the entire process to prevent contamination during spawn production.
For the spawn production, wheat was selected as the growth substrate. The wheat grains were first soaked overnight to allow them to absorb sufficient water, then boiled in warm water until fully expanded without breaking. After boiling, the wheat was drained for around 1 h until its water content reached approximately 60%. The wheat was placed into 500 mL glass bottles with breathable lids and sterilized at 121 °C for 40 min.
Degradation experiment
-
Compost preparation involved producing 1 ton of material with the following composition: 43% wood chips, 43% bagasse, 6% cow manure, 6% wheat bran, 1% calcium phosphate (CaP2H4O8), and 1% quicklime. The composting process consisted of two main stages. During the first stage, thermophilic bacteria decomposed the raw material, raising the temperature to 80 °C and leading to ammonia release. Regular turning of the compost heap facilitated ammonia evaporation, with the remaining 40% consumed by microorganisms. The entire composting process took approximately 30 d[22]. Moisture levels were carefully maintained throughout, and the pile was turned regularly. The fermented compost had a distinctive odor, which was free of ammonia or other pungent smells, and exhibited a loose, non-sticky texture with a water content of around 65%. After fermentation, the compost was allowed to dissipate heat and cool down before being applied to the field. Each square meter of soil received approximately 10 kg of compost.
The experiment comprised five treatments: untreated soil (S), soil with compost (SC), soil with compost and A. subrufescens inoculation (SCA), soil with compost and high glyphosate concentration (SCH), and soil with compost, A. subrufescens, and high glyphosate concentration (SCAH). The glyphosate concentrations were based on typical field application rates: 120 mL of glyphosate diluted in 15 L of water for standard application, with high concentration achieved by spraying three times the standard amount. The experiment was conducted across 15 plots, each measuring 70 cm × 10 m, with three replicates per treatment.
To prepare the plots, the soil was loosened, and the glyphosate solution was sprayed onto the soil surface. A 20 cm thick layer of compost was evenly spread over the soil. The A. subrufescens spawn was distributed across the compost at a rate of 2–3 bottles per square meter and gently compacted to ensure proper contact. A 3–5 cm layer of soil was added on top, and soil moisture content was adjusted to approximately 75%. During the mycelium growth period, water was regularly applied to maintain consistent moisture levels in the overlying soil.
Soil samples were collected from the underlying soil beneath the fungal matrix using a 15 cm soil sampler at four intervals: the beginning of inoculation (day 0), the middle of degradation (day 30), and the end of degradation practice (day 60). Samples were collected for each replicate, sealed in sterile plastic tubes, and stored in a freezer at -20 °C for further analysis.
Glyphosate content analysis
-
To analyze glyphosate content, 10 g of soil or ground coffee fruit sample was weighed and transferred to a centrifuge tube. The 50.0 mL of sodium phosphate and sodium citrate mixture was added, and the sample was subjected to ultrasonic treatment for 30 min to facilitate extraction. After extraction, the sample was centrifuged, and the supernatant was collected. Subsequently, 50.0 mL of n-hexane was added, and the mixture was shaken and allowed to stand. The aqueous phase was extracted again using an additional 50.0 mL of n-hexane.
A 1.0 mL aliquot of the purified solution was taken, to which 0.12 mL of sodium tetraborate solution and 0.2 mL of 9-fluorene methyl chloroformate solution were added. The solution underwent derivatization for 4 hr and was then filtered for analysis[25]. Blank samples were prepared using the same extraction and derivatization processes with quartz sand (particle size of 0.3–0.7 mm).
Standard curves were generated using standard solutions with concentrations of 10.0, 20.0, 50.0, 100, 200, and 500 μg/L. The standard solutions followed the same derivatization steps as the original soil samples.
Chromatographic analysis was performed using a C18 column (250 mm × 4.6 mm, 5 μm) at a temperature of 35 °C. The injection volume was 20 μL, and the flow rate was set to 1 mL/min.
The degradation percentage of pesticides was calculated using the following formula:
$ \mathrm{DG\;({\text{%}} )\;=\;(G0−Gt)/G0} \;{\times }\;{100{\text{%}} } $ where, DG is the glyphosate degradation percentage; G0 is the glyphosate concentration on day 0; and Gt is the glyphosate content at specific time intervals.
Soil enzyme activity analysis
-
Soil enzyme activity in the samples was measured using an ELISA kit on days 30 and 60. The detection principle for each enzyme (FDA, DHA, Lac, MnP, and LiP) involved the use of purified, enzyme-specific antibodies immobilized as solid-phase antibodies. The respective enzymes (FDA, DHA, Lac, MnP, and LiP) were added sequentially to bind with their corresponding antibodies, forming an enzyme-antibody complex. In this process, the HRP enzyme catalyzes a color reaction with TMB, initially producing a blue that then changes to yellow[26]. The specific steps taken followed the ELISA kit instructions. The intensity of the color reaction is directly proportional to enzyme activity. Absorbance (OD value) was measured at a wavelength of 450 nm, and the enzyme activity in each sample was quantified by plotting a standard curve.
Microbiological analysis
-
A soil sample weighing 200−500 mg was combined with 1× PBS solution and centrifuged at 10,000 rpm for 3 min. The supernatant was then collected for subsequent analysis. DNA extraction was carried out according to the instructions provided in the kit manual.
For fungal DNA amplification, the ITS1-ITS2 region of the fungal internal transcribed spacer (ITS) was targeted using the forward primer ITS1F (5'-CTTGGTCATTTAGAGGAAGTAA-3') and the reverse primer ITS2R (5'-GCTGCGTTCTTCATCGATGC-3'). Bacterial 16S rRNA was amplified in the V3-V4 region using the forward primer 341F (5'-CCTACGGGNGGCWGCAG-3') and the reverse primer 805R (5'-GACTACHVGGGTATCTAATCC-3')[27].
The DNA concentration for the PCR reaction was measured using the Qubit 4.0 DNA Kit. The PCR mixture (30 μL) included 15 μL of 2×Hieff® Robust PCR Master Mix, 1 μL of forward primer, 1 μL of reverse primer, 3 μL of sample DNA, and 11 μL of ddH2O. The PCR procedure consisted of the following steps: predenaturation at 95 °C for 5 min, denaturation at 95 °C for 1 min, annealing at 60 °C for 1 min, extension at 72 °C for 1 min, repeated for 30 cycles, followed by a final extension at 72 °C for 7 min.
The library was verified by agarose gel electrophoresis, and DNA concentration was measured using a fluorescence quantifier. After pooling the samples in equal quantities, they were sequenced using the Illumina MiSeq platform (
www.illumina.com ) to assess microbial diversity, community composition, and abundance variations. High-throughput sequencing and data analysis were performed by Shanghai Shenggong Biological Company.Data analysis
-
The collected data on enzyme activities were organized and analyzed using Microsoft Excel and SPSS 26.0 software. Statistical analysis was performed using one-way ANOVA and Duncan's Multiple Range test to identify significant differences among treatments (p < 0.05). Data visualization was carried out using GraphPad Prism 8.0.2.
For microbial community structure analysis, raw sequence data in FASTQ format were processed using FLASH to merge paired-end reads and obtain high-quality sequences[28]. Sequences were aligned and clustered into operational taxonomic units (OTUs) at a 97% sequence similarity threshold using UCLUST[29]. OTUs with an abundance below 0.001% of the total sequenced quantity were excluded from further analysis[30]. Taxonomic classification and subsequent microbial community analyses were based on GenBank taxonomic information for each OTU. Microbial community richness and diversity were evaluated using the Chao1 index[31], while the Shannon index was used to measure evenness by Mothur[32]. Microbial community composition data were clustered based on OTU counts and taxonomic information using Mega[33] and BioEdit[34].
-
Fungal mycelium required approximately 40 d to fully colonize the compost. Soil samples from different treatments were collected on the 60th d and analyzed. The glyphosate degradation percentages under the various treatments are presented in Fig. 1. The initial glyphosate concentrations for normal and high application rates were 1.259 and 4.75 mg/kg, respectively, at the start of the in-situ remediation experiment.
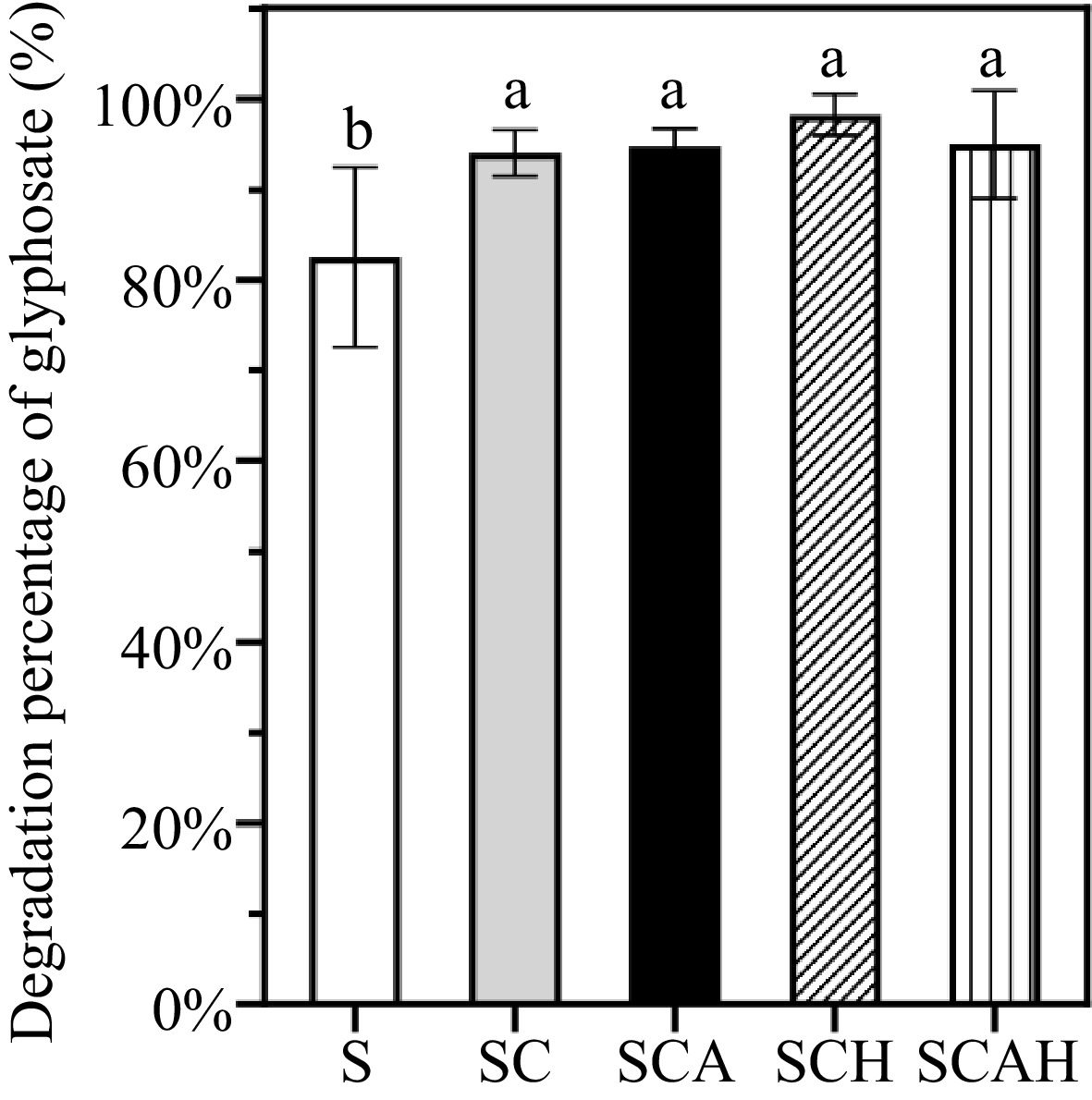
Figure 1.
Degradation percentages of glyphosate in topsoil under different treatments in the coffee plantation. Note: There were three replicates for each treatment (n = 3). Univariate analysis of variance (ANOVA) and Duncan's Multiple Range test were used to assess the significance of the difference. Different letters indicate statistically significant differences (p < 0.05). S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosate concentration.
After 60 d, significant glyphosate degradation was observed across all the treatments. In the untreated control soil (S), the degradation percentage reached 82.52% by day 60. Other treatments exhibited significantly higher degradation percentages compared to the control. The degradation percentages for SC, SCA, SCH, and SCAH were 94.01%, 94.68%, 98.25%, and 94.95%, respectively, indicating that glyphosate in the topsoil was nearly fully degraded by the 60th d.
Notably, there was no significant difference in degradation percentages between SC and SCA or SCH and SCAH on the 60th d (Fig. 1). These findings suggest that both the inoculation of A. subrufescens and the use of fermented compost substrate significantly enhance glyphosate degradation.
Additionally, we tested the glyphosate content in the coffee fruits before the experiment began and one year after the experiment ended during coffee harvest season (November) (Table 1). Glyphosate residues were not detected (< 0.01 mg/kg) in the coffee fruit after 12 months of mycoremediation practice.
Table 1. Glyphosate residue levels in coffee fruits from the experimental site.
Sample code Unit Value (before the experiment begins) Value (12 months after the experiment) 1 mg/kg 0.048 < 0.01 2 mg/kg < 0.01 < 0.01 3 mg/kg 0.074 < 0.01 4 mg/kg < 0.01 < 0.01 5 mg/kg 0.105 < 0.01 6 mg/kg 1.06 < 0.01 7 mg/kg 0.028 < 0.01 8 mg/kg 0.027 < 0.01 9 mg/kg 0.021 < 0.01 Changes in soil enzyme activity
-
Soil samples were collected and analyzed at 30 and 60 d under different treatments, and the changes in enzyme activities are shown in Fig. 2. On day 30, FDA activity was higher in the S, SC, and SCH treatments, while the enzyme activities in the SCA and SCAH treatments were lower. By day 60, FDA activity in the S and SC treatments remained high, reaching 0.33 and 0.31 IU/g, respectively, while the enzyme activities in the other treatments were lower. These results indicate that fungal inoculation or the addition of compost materials can alter FDA activity in soil.
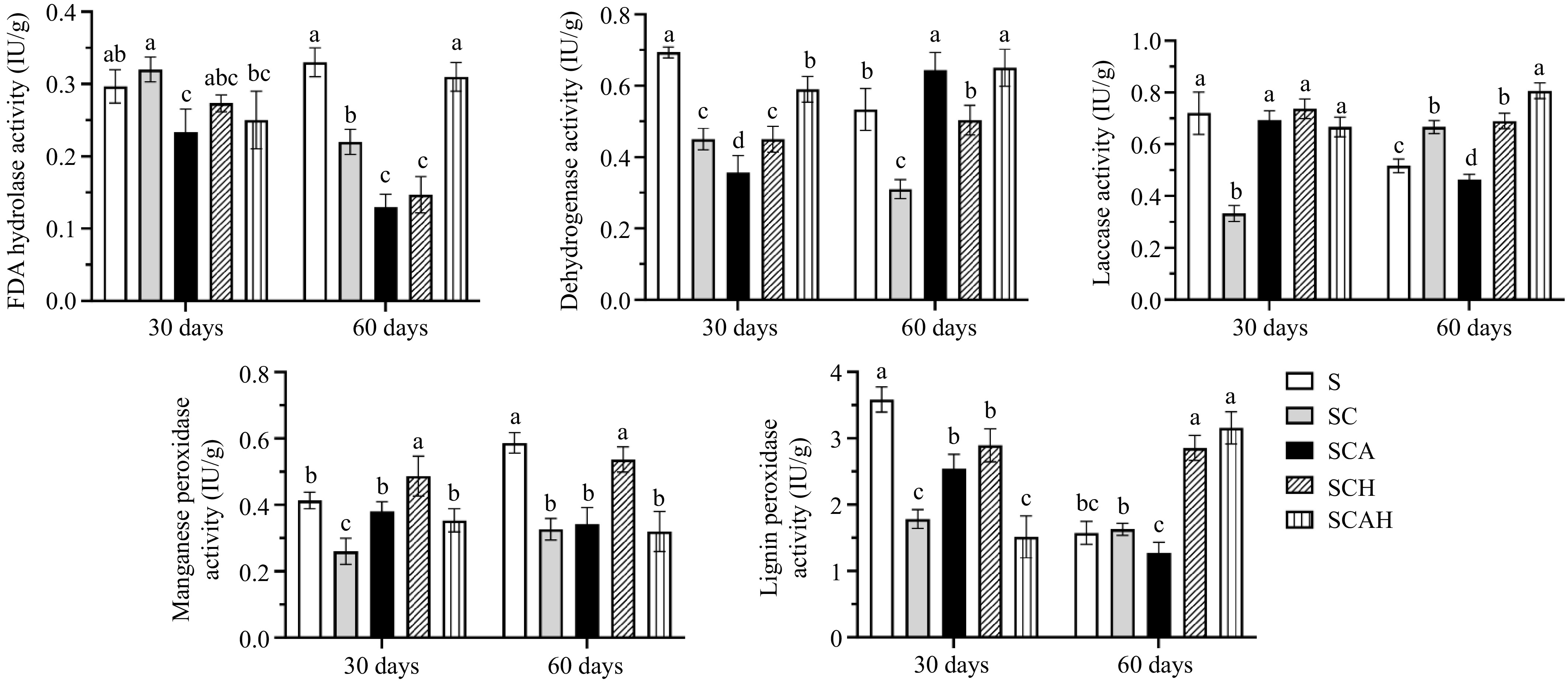
Figure 2.
Changes in soil enzyme activity during cultivation of Agaricus subrufescens in the coffee plantation. Note: There were three replicates for each treatment (n = 3). Univariate analysis of variance (ANOVA) and Duncan's Multiple Range test were used to assess the significance of differences. Different letters indicate statistically significant differences (p < 0.05). S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosate concentration.
Regarding DHA activity, on day 30, the treatments showed lower DHA activity compared to the control (S), with the SCA treatment exhibiting the lowest value at 0.37 IU/g. The DHA activity in the SC and SCH treatments on day 30 was higher than in SCA but lower than in SCAH. These results suggest that both fungal inoculation and compost substrate addition can influence DHA activity. On day 60, the DHA activity in the SCA and SCAH treatments increased significantly, surpassing the activity in the control (S), while the DHA activity in the SC and SCH treatments was lower than in S.
Overall, DHA activity in the control (S) and the SC treatment showed a decreasing trend, whereas in the SCA, SCH, and SCAH treatments, it exhibited an increasing trend, with the increase in SCH being less pronounced between days 30 and 60. This suggests that inoculation with A. subrufescens can increase DHA activity over time, while compost substrate has no significant effect on DHA activity.
Regarding Lac activity, on day 30, no significant differences were observed between the S, SCA, SCH, and SCAH treatments, while the SC treatment had the lowest value at 0.33 U/g. By day 60, Lac activities in the SC, SCH, and SCAH treatments were higher than in the control (S), with SCAH reaching the highest value of 0.81 U/g. These results suggest that A. subrufescens can enhance Lac activity either during mycelial colonization or after a period of compost substrate activity.
For MnP activity, no significant differences were observed between the S, SCA, and SCAH treatments on day 30, although differences were noted in the SC and SCH enzyme activities. By day 60, MnP activity in the control group (S) reached the highest value of 0.59 U/g, followed by SCH, while enzyme activities in SC, SCA, and SCAH were lower. This indicates that fungal inoculation and compost addition influence MnP activity, with the compost substrate having a more pronounced effect over time.
On day 30, LiP activity in the control (S) was significantly higher than in other treatments, with notable differences observed between SC and SCA, as well as between SCH and SCAH. By day 60, LiP activity in SCH and SCAH was significantly higher than in the other treatments. These findings suggest that A. subrufescens inoculation or the addition of compost materials can increase LiP activity.
Soil microbial community structure
Summary
-
High-throughput sequencing results showed the following average number of effective sequences for bacterial communities in five treatments (S, SC, SCA, SCH, and SCAH): 97,652, 131,907, 124,112, 119,511, 128,388, and 129,617, respectively, with an average sequence length of 412 bp. For the fungal community, the average number of effective sequences for the same treatments was 76,961, 169,576, 146,953, 125,044, 137,100, and 142,876, respectively, with an average sequence length of 243 bp.
Richness and diversity of microbial communities
-
The changes in soil microbial community diversity were analyzed after 60 d of cultivation. The soil fungal α-diversity indices are summarized in Table 2, which shows a coverage rate of 99.9% for all tested soil samples, indicating nearly complete coverage within each sample. The Shannon index was higher in the SCAH, S, and SC treatments than in SCA and SCH, indicating that the soil fungal community diversity was greatest in SCAH and lowest in SCH. When comparing the Chao1 indices, the control (S) treatment group had higher values than SC, SCA, SCH, and SCAH, with the SCA group showing the lowest indices. These results suggest that the richness of the soil fungal community decreased under the influence of A. subrufescens.
Table 2. Alpha diversity indices of the soil fungal communities for each treatment.
Alpha index S SC SCA SCH SCAH Shannon 3.25 ± 0.83a 3.22 ± 0.51a 2.81 ± 0.18a 2.68 ± 0.4a 3.27 ± 0.81a Chao1 803.6 ± 120.1a 725.2 ± 43.1a 650.0 ± 82.3a 682.3 ± 48.7a 667.1 ± 17.8a Coverage 99.93% 99.92% 99.91% 99.91% 99.92% Data represent the means ± standard deviations (n = 3). The results of multiple comparisons are marked with different letters to indicate significant differences (p < 0.05). Soil samples were taken from the respective treatments in the coffee plantation at the experimental site in Pu'er, Yunnan. S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosateconcentration. The α-diversity of the soil bacterial community is shown in Table 3, with a coverage rate greater than 98.7% for all tested soil samples. The Shannon index was significantly higher in the SCH treatment compared to SCA and SCAH, with SC following closely behind SCH. The Chao1 index was highest in SCH (mean value of 5,216.8), followed by SC, while SCA exhibited significantly lower values than other groups. In summary, the addition of the compost substrate increased the diversity and richness of the soil bacterial community, while A. subrufescens appeared to reduce bacterial community diversity and richness to some extent.
Table 3. Alpha diversity indices of the soil bacterial communities for each treatment.
Alpha index S SC SCA SCH SCAH Shannon 6.18 ± 0.16abc 6.43 ± 0.31ab 5.98 ± 0.31c 6.57 ± 0.15a 6.03 ± 0.10bc Chao1 4789.8 ± 329.2ab 4905.6 ± 437ab 4245.7 ± 510.4b 5216.8 ± 28.1a 4544.1 ± 372.4ab Coverage 98.85% 98.70% 98.84% 98.76% 98.85% Data represent the means ± standard deviations (n = 3). The results of multiple comparisons are marked with different letters to indicate significant differences (p < 0.05). Soil samples were taken from the respective treatments in the coffee plantation at the experimental site in Pu'er, Yunnan. S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosateconcentration. Classification and composition of microbial community
-
Figure 3 illustrates the fungal community structure and composition of soil samples on day 60. As shown in the figure, Ascomycetes and Basidiomycetes dominated the phylum classification level across all treatments. Compared to the control (S), the relative abundances of Basidiomycetes decreased in the SC and SCAH treatments, while the relative abundance in the SCA and SCH treatments remained relatively unchanged. In contrast, the relative abundances of SC, SCA, SCH, and SCAH increased significantly compared to S, with SC and SCH showing the largest increases. This suggests that fungal inoculation and compost addition can influence the composition of the fungal community, particularly the phylum-level distribution.
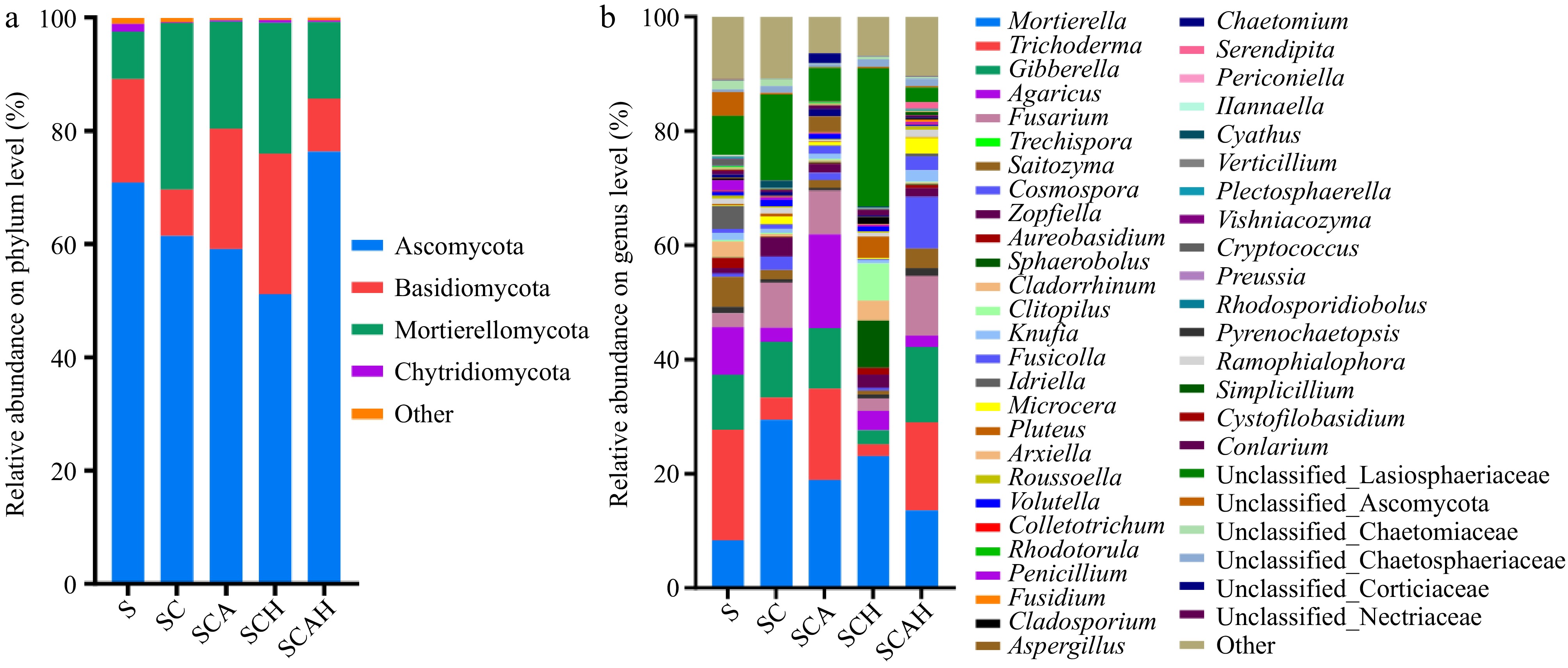
Figure 3.
Composition of soil fungal communities at the (a) phylum, and (b) genus levels. Soil samples were taken from the respective treatments in the coffee plantation at the experimental site in Pu'er, Yunnan. S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosate concentration.
At the genus classification level, four major genera were identified: Agaricus, Gibberella, Mortierella, and Trichoderma. These genera accounted for 8.39%, 9.58%, 8.34%, and 19.40%, respectively, in the S treatment. Figure 3 shows that the relative abundance of Agaricales in the SCA group increased significantly due to A. subrufescens inoculation, while SC, SCH, and SCAH exhibited a decrease. Additionally, the relative abundance of Trichoderma in the S, SCA, and SCAH groups showed minimal change, whereas SC and SCH demonstrated a significant reduction, indicating that compost materials may inhibit the growth of Trichoderma.
In summary, both A. subrufescens inoculation and compost materials influenced the structure and composition of the fungal community, with the compost substrate having a pronounced effect on the growth of Mortierellomycota and Trichoderma.
Figure 4 presents the bacterial community structure. At the phylum level, Proteobacteria, Acidobacteria, Actinobacteria, and Bacteroidetes were the dominant groups across all treatments. Generally, there were no significant changes in bacterial phyla compared to the control (S), regardless of whether fungi were inoculated or compost materials were added. The only exception was the SCAH treatment, which showed a notable increase in Firmicutes.
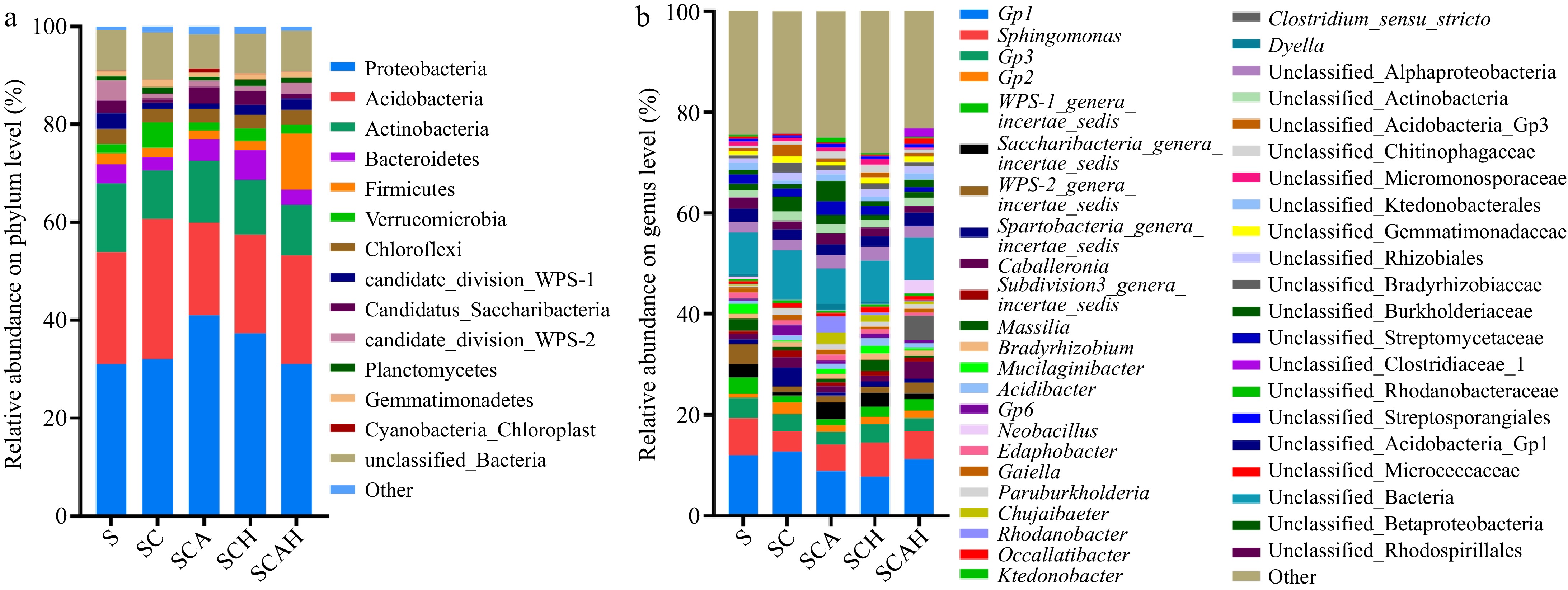
Figure 4.
Composition of soil bacterial communities at the (a) phylum, and (b) genus levels. Soil samples were taken from the respective treatments in the coffee plantation at the experimental site in Pu'er, Yunnan. S, untreated soil; SC, soil with compost; SCA, soil with compost and A. subrufescens inoculation; SCH, soil with compost and high glyphosate concentration; SCAH, soil with compost, A. subrufescens, and high glyphosate concentration.
At the genus level, the predominant genera across all treatments were Gp1, Sphingomonas, Gp3, and Gp2. Changes in these genera were minimal compared to the control (S). In conclusion, A. subrufescens inoculation and the addition of compost substrate did not significantly alter the bacterial community structure or composition at either the phylum or genus level.
-
Glyphosate is a widely used non-selective herbicide that effectively controls weeds but also poses risks to the environment, as it can spread through various environmental vectors such as rain, air, and wind. Due to its chemical properties—including three active functional groups (phosphoric acid, amino, and carboxyl groups)—glyphosate readily binds to soil particles, which reduces its leaching potential but also contributes to its persistence in the soil[35]. Given its extensive use and environmental persistence, the remediation of glyphosate and its behavior in the environment warrant careful consideration and continued research.
The half-life of glyphosate in soil can vary significantly, ranging from 1 to 90 d, depending on several factors, including soil composition, microbial activity, rainfall, and temperature[36]. In our experiments, over 90% degradation of glyphosate within 60 d was observed, consistent with the lower end of the reported half-life under similar environmental conditions. This result suggests that the microbial and soil conditions in our study were conducive to glyphosate degradation, occurring within the typical timeframe for its breakdown in soil.
Glyphosate degradation is primarily carried out by microorganisms, and mycoremediation presents an effective and environmentally friendly solution for mitigating glyphosate contamination[37−39]. Several fungal strains from genera Aspergillus, Fusarium, and Trichoderma have been isolated from soil and shown to thrive on organophosphorus pesticides and herbicides, including glyphosate[40]. For example, one highly efficient glyphosate-degrading strain, Fusarium verticillioides C-2, has been reported to reduce glyphosate concentrations by 72% and 89% in sterile and non-sterile soils, respectively, after 28 days of laboratory incubation with an initial glyphosate concentration of 60 mg/kg. This highlights its potential for bioremediating glyphosate-contaminated soils[41]. Another study found that Aspergillus flavus WDCZ2 degraded 800 ppm of glyphosate with a remarkable efficiency of 99.6%, demonstrating its promise as a candidate for glyphosate bioremediation[42]. In this study, the composition of fungal communities at genus level (Fig. 3) presents an increase of Fusarium, and a decrease of Aspergillus and Trichoderma. This occurs probably due to the competition between fungal-bacteria or among fungal species in the soil ecosystem[43]. Future studies could investigate the interactions among functional microbial species in the mycoremediation process.
In our study, both SCA and SCAH treatments significantly (one-way ANOVA analysis) enhanced the degradation of glyphosate compared to the control (S), with degradation percentages of 94.68% and 94.95%, respectively. This indicates that inoculation with A. subrufescens has strong potential for facilitating glyphosate degradation. The enhanced degradation observed may be attributed to two main factors: first, A. subrufescens mycelium colonizes the soil and acts as a degradation medium, where its unique enzymatic system stimulates changes in soil enzymes and microbial communities, thereby accelerating the breakdown of glyphosate. Second, the compost substrate used in the inoculation process likely interacts with both the fungi and indigenous microorganisms, further promoting glyphosate degradation. These synergistic interactions warrant further investigation to fully understand the mechanisms involved in glyphosate bioremediation through fungal inoculation.
Compost is a biologically active medium, and its addition to contaminated soils can enhance pesticide decomposition by stimulating microbial activity and promoting plant growth[44]. Previous studies have shown that compost not only promotes plant growth but also increases microbial activity, leading to a significant rise in pesticide degradation percentages in the rhizosphere. Additionally, compost improves soil water-holding capacity and organic matter content, which helps reduce pesticide mobility[45]. For example, compost made from cotton residues demonstrated high efficacy in degrading terbuthylazine, illustrating the role of organic matter in compost and microbial community in accelerating herbicide degradation[46].
In the present in situ experiment, glyphosate degradation percentages in the control (S) reached 82.52% by day 60. In comparison, degradation percentages for treatments SC, SCA, SCH, and SCAH reached 94.01%, 94.68%, 98.25%, and 94.95%, respectively. These results indicate that the compost substrate provided an ideal environment for fungal growth, enabling the production of lignin-degrading enzymes that catalyze the breakdown of various compounds. This interaction between the compost substrate and contaminated soil further promoted herbicide degradation.
While monitoring glyphosate content in the soil is vital for evaluating the effectiveness of bioremediation, it is equally important to assess changes in soil enzyme activity. Enzyme activity is a sensitive indicator of ecological shifts, as it can quickly respond to subtle environmental changes[47]. Previous research by Pizzul et al.[48] highlighted the potential of lignin-degrading enzymes in pesticide degradation, demonstrating that the presence of laccase (Lac) and manganese peroxidase (MnP) increased the conversion rate of glyphosate to its metabolite AMPA. In this study, Lac, MnP, and LiP activities varied across different time points. Lac activity in treatments SC, SCH, and SCAH was higher than in the control (S), while MnP activity was lower in SC, SCA, and SCAH compared to S, with the compost substrate having a more pronounced effect on MnP activity at specific time points. Significant differences in LiP activity were also observed, indicating that lignin-degrading enzyme activity is influenced by both A. subrufescens inoculation and the compost substrate in the plantation.
On day 30, FDA activity increased significantly in the presence of the compost substrate. Agaricus subrufescens enhanced DHA activity in the soil, with both SCA and SCAH treatments showing increased activity, while the compost substrate alone had no significant effect on DHA activity. In summary, A. subrufescens may promote glyphosate degradation by enhancing FDA and DHA enzyme activities, while the compost substrate likely influences degradation primarily through increased FDA activity. The combined action of microorganisms significantly boosts FDA and DHA enzyme activities, which are closely correlated with glyphosate degradation.
Glyphosate degradation in soil is primarily facilitated by a variety of microorganisms, especially bacteria, which interact with soil factors to influence the fate of glyphosate[49]. Schnurer et al.[50] found that glyphosate bound to soil becomes more susceptible to microbial degradation. High concentrations of glyphosate have been shown to reduce the number of microbial populations compared to soils treated with normal doses, and glyphosate can further impact fungal populations by disrupting their interactions with other microorganisms[51,52].
In this study, there were no significant changes in the richness and diversity of the fungal community in the SC, SCA, SCH, and SCAH treatments compared to the control (S). However, the bacterial community diversity in SC and SCH was higher than in SCA and SCAH, with the Chao1 indices being highest in SCH. This suggests that the compost substrate enhances the diversity and richness of the soil bacterial community. In contrast, A. subrufescens appeared to reduce bacterial diversity and richness to some extent. During in situ remediation, environmental factors influenced soil microbial communities differently. The compost substrate likely contributed to glyphosate degradation by enhancing bacterial diversity and richness, while A. subrufescens inoculation had a more selective impact on the microbial community.
The bacterial composition and fungal composition in glyphosate-contaminated soil are significantly different from those in non-contaminated soils through PERMANOVA analysis based on a previous study[53]. Glyphosate impacts bacterial communities, and these effects on fungal communities tend to be more pronounced[54]. Guijarro et al.[55] reported glyphosate degradation occurring in soil without significant changes in bacterial community structure, which aligns with the findings of this study. In this study, the relative abundance of Basidiomycota decreased, while Mortierellomycota increased significantly compared to the control (S). This suggests that fungal inoculation and glyphosate contamination have a more noticeable impact on fungal communities. In line with Lancaster et al.[56], the study found that Proteobacteria, Actinomycetes, and Firmicutes dominated the soil environmental communities. In this research, Gp1, Sphingosphingomonas, and Gp3 were the predominant genera across all treatments. Zhang et al.[57] and Fei et al.[58] showed that the diversity of Acidobacteria is related to soil enzyme activity, indicating that glyphosate degradation may be facilitated by its influence on soil enzyme activity during remediation.
-
In coffee plantations, glyphosate was almost completely degraded by the 60th d, with the inoculation of A. subrufescens and the addition of compost substrate significantly promoting this degradation. The cultivation of Agaricus subrufescens resulted in the degradation of glyphosate by altering the microbial community structure and composition, increasing the abundance of glyphosate-degrading bacteria, and significantly enhancing the activities of key soil enzymes such as FDA, Lac, DHA, and LiP. The compost substrate further facilitated the degradation of glyphosate-contaminated soil, primarily by boosting Lac activity through an increase in bacterial community diversity and richness. Additionally, glyphosate residues were not detected in the coffee fruit after one year of mycoremediation, indicating the effectiveness of this approach in reducing glyphosate contamination. Understanding the molecular mechanisms underlying fungal and microbial interactions in glyphosate degradation will provide deeper insights into optimizing this bioremediation strategy.
-
Not applicable.
This study was supported by the Department of Science and Technology of Yunnan, China (Grant Nos: 202303AP140001 and 202202AE090091). We thank Ms. Yan Huang for providing her family coffee farm as the experimental site. We also acknowledge the Rainforest Alliance for its collaboration in the fieldwork.
-
The authors confirm contributions to the paper as follows: writing - draft manuscript preparation: Du Y; writing - manuscript review: Hu Y, Mortimer PE; formal analysis: Du Y, Gui H; validation, methodology: Zhang H, Bandara AR, Gui H, Mortimer PE; resources: Zhang H, Bossolasco L; project administration: Bossolasco L, Pu Y; language editing: Bandara AR, Hu Y, Mortimer PE; conceptualization: Pu Y, Hu Y; supervision, data curation: Hu Y. All authors reviewed the results and approved the final version of the manuscript.
-
The data supporting this study's findings are available from the corresponding authors on reasonal request.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Du Y, Zhang H, Bossolasco L, Bandara AR, Pu Y, et al. 2025. Field cultivation of Agaricus subrufescens Peck lowers soil glyphosate levels, alters soil microbial communities, and boosts soil enzyme activities in coffee plantations. Studies in Fungi 10: e009 doi: 10.48130/sif-0025-0009 

Field cultivation of Agaricus subrufescens Peck lowers soil glyphosate levels, alters soil microbial communities, and boosts soil enzyme activities in coffee plantations
- Received: 20 January 2025
- Revised: 18 April 2025
- Accepted: 23 April 2025
- Published online: 10 June 2025
Abstract: Glyphosate is one of the most widely used non-selective organophosphate herbicides in agriculture. However, its extensive application has resulted in glyphosate accumulation in soil systems and surrounding water bodies, negatively affecting plants, animals, microorganisms, and human health. Compost-based mushroom cultivation has emerged as a promising approach for glyphosate remediation, leveraging the ability of fungal mycelium to extend from compost into contaminated soils and degrade glyphosate. While previous research has focused on laboratory-scale mycoremediation (a form of bioremediation that uses fungi to degrade, remove, or neutralize environmental pollutants from the environment), this study explores the effects of compost and fungal inoculation on glyphosate degradation, microbial community structure, and enzyme kinetics under field conditions. Five treatments were designed in a glyphosate-contaminated coffee plantation: untreated soil (control), soil with compost, soil with compost and Agaricus subrufescens Peck, soil with compost and high glyphosate concentration, and soil with compost, A. subrufescens, and high glyphosate concentration. After 60 d, glyphosate concentrations, microbial communities, and enzyme activities were evaluated. The results indicated that field mushroom cultivation achieved glyphosate degradation percentage of 94%–98%, significantly higher than the 82% observed in the untreated soil, using one-way ANOVA analysis. Fungal inoculation altered soil microbial community structure and significantly increased enzyme activities, such as fluorescein diacetate, dehydrogenase, and lignin peroxidase. Additionally, compost substrate significantly enhanced laccase activity and improved soil bacterial diversity and richness, promoting glyphosate degradation in conjunction with indigenous microorganisms. In conclusion, this study suggests a comprehensive method to integrate waste management, soil health, and bioremediation into agricultural practice.
-
Key words:
- 16S rRNA gene amplicon /
- Bioremediation /
- Coffee industry /
- Herbicide /
- High-throughput sequencing /
- ITS /
- Soil microbiome