








-
Pestalotioid species are a group of fungi that cause a variety of diseases in a range of different hosts worldwide. Most pestalotioid species have their asexual morph with appendages bearing conidia and are extensively dispersed in tropical and temperate regions. It is well known that most of the identified pestalotioid species are phytopathogens[1,2]. In contrast, some are endophytes that can produce several bioactive compounds with potential applications in medicine, agriculture, and various industries[1−3]. Due to their ability to switch nutritional modes, many phytopathogenic and endophytic pestalotioid species occur as saprobes inhabiting dead leaves, bark, and twigs[1,4−7]. Fungal taxonomists have increasingly relied on multi-locus phylogeny combined with morphological characteristics for species identification. Pestalotioid taxa were segregated into three genera: Pestalotiopsis, Neopestalotiopsis, and Pseudopestalotiopsis in the family Sporocadaceae, which was introduced by Corda in 1842[8,9]. The conidial morphology of Pestalotiopsis, Neopestalotiopsis, and Pseudopestalotiopsis is largely identical, comprising five cells: a basal cell with a single appendage, a hyaline apical cell with one or more appendages, and three pigmented median cells. However, the color of the pigmented cells differentiates these taxa, with Pseudopestalotiopsis and Pestalotiopsis being concolorous, while Neopestalotiopsis exhibits versicolorous pigmentation. Although Pestalotiopsis and Pseudopestalotiopsis share similar conidial morphologies, they can be distinguished by having three slightly darker pigmented cells in Pseudopestalotiopsis[1,4−6]. Sexual morphs are rare among Pestalotiopsis[1], Pseudopestalotiopsis[6] , and Neopestalotiopsis[1,3] species. Distinguishing between the sexual morphs of Pseudopestalotiopsis based solely on morphology is challenging due to their close resemblance to Pestalotiopsis[1,6,7]. Therefore, researchers have increasingly relied on multigene phylogeny combined with morphological characteristics to identify the Pestalotiopsis and allied genera[1,6,10,11].
Members of Neopestalotiopsis are commonly distributed in tropical and subtropical ecosystems worldwide as phytopathogens, endophytes, or saprobes[1,3]. Furthermore, common phytopathogens of Neopestalotiopsis are associated with a variety of plant diseases, including shoot and twig dieback[12,13], leaf spots[14−16], crown rot[17], leaf blight[17−19], fruit rot[20,21], flower drying[22,23], and leaf fall diseases[24−26]. They also contribute to postharvest diseases in fruits such as mango, rose apple, mangosteen, strawberry, and avocado[20,27,28]. Over the past few years, a significant number of Neopestalotiopsis species have been described, highlighting the remarkable species diversity within this genus. The genus currently comprises 127 taxa (Mycobank 2025:
www.mycobank.org/page/Home ). Further investigations of pestalotioid fungi are likely to uncover additional Neopestalotiopsis species and provide insights into their ecological interactions.Garcinia mangostana (mangosteen) and A. muricata (soursop) are tropical fruit trees, distributed mainly in tropical and subtropical countries, including Sri Lanka. Economically important plants such as G. mangostana (mangosteen) and A. muricata (soursop), cultivated in Sri Lanka, are also susceptible to pestalotioid phytopathogens. Unraveling the molecular phylogeny of pestalotioid species causing leaf lesions in G. mangostana and A. muricata in Sri Lanka has practical implications for disease management, agriculture, biodiversity, and economic conservation. Molecular diagnostic tools can be used for the early detection of plant diseases and timely intervention, helping to mitigate the economic losses associated with pathogen-associated infections. Furthermore, accurate identification of the pathogens using detailed molecular and morphological identification is essential for developing effective biocontrol methods to manage the diseases they cause, thereby supporting economic development. Most pestalotioid taxa in Sri Lanka are recognized as phytopathogens with no host specificity. Therefore, this study aims to identify pestalotioid species associated with the leaves of G. mangostana and A. muricata in Sri Lanka based on comprehensive molecular phylogenetic analyses and morphological characterization to establish the taxonomic placement of the identified pestalotioid species. Furthermore, the present study evaluates the biocontrol potential of T. atroviride and Daldinia eschscholtzii against the isolated pestalotioid species using dual culture plate assays.
-
Symptomatic fresh leaves of G. mangostana (mangosteen) and A. muricata (soursop) were collected from Kadawatha, Gampaha District, Sri Lanka, and transferred to the laboratory using labelled zip-lock plastic bags. The micro-morphological characters of the specimens, such as conidial length, width, length, colour of median cells, and number of apical appendages, were examined and photographed using an Optika Trinocular Phase Contrast Microscope (B-510ASB, Italy) with a digital camera. All microscopic measurements were made with the Tarosoft image framework (v. 0.9.0.7).
Single spore isolation was carried out following the method described by Senanayake et al.[14]. Germinated conidia were aseptically transferred into fresh PDA plates and incubated at 28 °C to obtain pure cultures. Cultures were incubated at room temperature (28 °C) for 7–14 d. Cultures were later transferred to PDA slants and stored at 4 °C for further studies. Generated cultures were deposited at the culture collection of the Genetics and Molecular Biology Unit, University of Sri Jayewardenepura, Sri Lanka. Colony characteristics were recorded from cultures grown on potato dextrose agar (PDA). Herbarium specimens were deposited in the herbarium of the Department of Botany (USJ-H) and the Genetics and Molecular Biology Unit (USJ-GMBU) of the Sri Jayewardenepura University of Sri Lanka. Fungal isolates were deposited in the Culture Collection of the Genetics and Molecular Biology Unit (GBMUCC), University of Sri Jayewardenepura, Sri Lanka.
DNA extraction, PCR amplification, and agarose gel electrophoresis
-
Approximately 75–100 mg of the scraped axenic mycelia was transferred into labeled 1.5 mL microfuge tubes. Genomic DNA was extracted from the mycelia using the CTAB method, following the protocol described by Thambugala et al.[29]. Three loci (ITS, tub2 and tef1) were amplified, β-tubulin (tub2) with primers Bt2a/Bt2b[30]; internal transcribed spacer region of ribosomal DNA (ITS: ITS5/ITS4)[31] and the translation elongation factor 1-alpha gene (tef1: EF1-526F/EF1-1567R)[32]. The amplification reactions were performed in 25 μl volumes containing 8.5 μl of sterilized H2O, 10.5 μl of Go Taq® Hot Start Green Master Mix (mixture of Go Taq Hot Start Polymerase, dNTPs, and optimized buffer (Promega)), 2 μl of 25 mM MgCl2, 1 μl of each forward and reverse primer, and 2 μl of DNA template (1.2 μg/ml). The PCR thermal cycle program for ITS amplification was provided initially at 94 °C for 3 min, followed by 35 cycles of denaturation at 94 °C for 1 min, annealing at 55 °C for 1 min, elongation at 72 °C for 1 min, and final extension at 72 °C for 10 min. The PCR thermal cycle program for tub2 and tef1 gene amplification was provided with the same, except for the annealing temperature, which was used as 57.5 °C and 56 °C, respectively. The PCR results were observed on 1.2% agarose electrophoresis gels stained with ethidium bromide under ultraviolet light. Subsequently, PCR product purification and Sanger sequencing were performed at Genelabs Medical (Pvt) Ltd, Sri Lanka.
Sequencing and phylogenetic analyses
-
For the newly synthesized sequences in this study, a consensus sequence was derived using DNASTAR Lasergene SeqMan Pro v.8.1.3 from the sequences obtained from both forward and reverse primers. Low-quality end fragments of the DNA sequences were removed using the BioEdit program. A total of 120 strains belonging to the Neopestalotiopsis were included to construct the updated phylogenetic tree. The outgroup taxon used was Pestalotiopsis trachycarpicola (OP068).
The DNA sequences were subjected to BLASTn searches, and the reference nucleotide sequences for the phylogenetic analyses were retrieved from GenBank based on published data. Separate single-gene alignments for ITS, tub2, and tef1 were performed using MUSCLE in MEGA11. The alignments were visually inspected and manually corrected when necessary. Single-gene phylogenies were inferred for ITS, tub2, and tef1, followed by a combined multigene analysis to more accurately resolve closely related species. GenBank numbers and culture collection (strains) numbers of the taxa included in the phylogenetic analyses are in Table 1.
Table 1. Details of the Neopestalotiopsis strains included in this study.
Species Culture collection number ITS tub2 tef1 Ref. Neopestalotiopsis acrostichi MFLUCC 17–1754T MK764272 MK764338 MK764316 [53] N. acrostichi MFLUCC 17–1755 MK764273 MK764339 MK764317 [53] N. alpapicalis MFLUCC 17–2544T MK357772 MK463545 MK463547 [54] N. alpapicalis MFLUCC 17–2545 MK357773 MK463546 MK463548 [54] N. amomi HKAS 124563T OP498012 OP752133 OP653489 [33] N. amomi HKAS 124564T OP498013 OP765913 OP753382 [33] N. aotearoa CBS 36754T NR163673 KM199454 KM199526 [1] N. asiatica MFLUCC12–0286T JX398983 JX399018 JX399049 [1] N. australis CBS 114159T KM199348 KM199432 KM199537 [1] N. brachiata MFLUCC 17–1555T MK764274 MK764340 MK764318 [53] N. brasiliensis COAD 2166T MG686469 MG692400 MG692402 [44] N. camelliae–oleiferae CSUFTCC81T OK493585 OK562360 OK507955 [55] N. camelliae-oleiferae CSUFTCC82 OK493586 OK562361 OK507956 [55] N. cavernicola KUMCC 20–0332 MW581238 MW59032 MW590327 [56] N. cavernicola KUMCC 20–0269T MW545802 MW557596 MW550735 [56] N. chiangmaiensis MFLUCC 19–0048 MW248391 MH412725 MW259070 [57] N. chrysea MFLUCC12–0261T JX398985 JX399020 JX399051 [1] N. chrysea MFLUCC12–0262 JX398986 JX399021 JX399052 [1] N. clavispora MFLUCC12–0281T JX398979 JX399014 JX399045 [1] N. clavispora MFLUCC12–0280 JX398978 JX399013 JX399044 [1] N. cocoes MFLUCC 15–0152T KX789687 – KX789689 [58] N. coffeae–arabicae HGUP4015T KF412647 KF412641 KF412644 [59] N. coffeae–arabicae HGUP4109 KF412649 KF412643 KF412646 [59] N. cubana CBS 600 96T KM199347 KM199438 KM199521 [1] N. cubana CSUFTCC37 OK493583 OK562358 OK507953 [1] N. cubana CSUFTCC42 OK493584 OK562359 OK507954 [1] N. cubana GMBUCC 24–001 PP800244 PP889282 – This study N. dendrobii MFLUCC 14–0106T MK993571 MK975835 MK975829 [60] N. drenthii BRIP 72264a MZ303787 MZ312680 MZ344172 [22] N. egyptiaca PEST1/CBS 140162T KP943747 KP943746 KP943748 [61] N. elaeidis MFLUCC 15–0735T ON650689 – ON734012 [62] N. ellipsospora MFLUCC12–0283T JX398980 JX399016 JX399047 [1] N. eucalypticola CBS 264 37T KM199376 KM199431 KM199551 [1] N. eucalyptorum CBS 147684T MW794108 MW802841 MW805397 [63] N. foedans CGMCC 3 9123T JX398987 JX399022 JX399053 [1] N. foedans CGMCC 3 9178 JX398989 JX399024 JX399055 [1] N. formicarum CBS 362 72T KM199358 KM199455 KM199517 [1] N. formicarum CBS 115 83 KM199344 KM199444 KM199519 [1] N. fragariae ZHKUCC 22–0115 ON651145 ON685199 ON685197 [64] N. guajavae FMB0026T MF783085 MH460871 MH460868 [65] N. guajavicola MFLUCC 22–0134T OP802393 OP801770 OP830892 [65] N. hadrolaeliae VIC 47180T MK454709 MK465120 MK465122 [66] N. haikouensis SAUCC212271T OK087294 OK104870 OK104877 [67] N. haikouensis SAUCC212272 OK087295 OK104871 OK104878 [67] N. hispanica FMB–Guv–B3T MW794107 MW802840 MW805399 [63] N. honoluluana CBS 114495T KM199364 KM199457 KM199548 [1] N. honoluluana CBS 111535 KM199363 KM199461 KM199546 [1] N. hydeana MFLUCC 20–0132T MW266069 MW251119 MW251129 [68] N. hyperici KUNCC 22–12597T OP498010 OP765908 OP713768 [33] N. hyperici KUNCC 22–12598 OP498009 OP737883 OP737880 [33] N. Iberica XF6 OM333900 OM350157 OM171265 [63] N. Iberica CBS 147688T MW794111 MW802844 MW805402 [63] N. iranensis P815T KM074048 KM074057 KM074051 [69] N. iranensis P814 KM074045 KM074056 KM074053 [69] N. javaensis CBS 257 31T NR_145241 KM199437 KM199543 [1] N. longiappendiculata MEAN–1315T MW794112 MW802845 MW805404 [63] N. lusitanica MEAN 1320 MW794097 MW802830 MW805409 [63] N. lusitanica MEAN–1317 MW794110 MW802843 MW805406 [63] N. macadamiae BRIP 63737cT KX186604 KX186654 KX186629 [25] N. macadamiae BRIP 63742a KX186599 KX186657 KX186627 [25] N. maddoxii BRIP 72266aT MZ303782 MZ312675 MZ344167 [22] N. magna MFLUCC 12–0652T KF582795 KF582793 KF582791 [1] N. mesopotamica CBS 33686T KM199362 KM199441 KM199555 [1] N. mianyangensis CGMCC 323554T OP546681 OP672161 OP204793 [70] N. mianyangensis UESTCC 22–0006 OP082291 OP235979 OP204793 [70] N. musae MFLUCC 22–0162 OP802388 OP801765 OP830887 [58] N. musae MFLUCC 15–0776T KX789683 KX789686 KX789685 [58] N. natalensis CBS 138.41T NR_156288 KM199466 KM199552 [1] N. nebuloides BRIP 66617T OM417249 ON995130 ON624181 [71] N. nebuloides BRIP 70567 OM417295 ON995131 ON624201 [71] N. olumideae BRIP 72273aT MZ303790 – – [22] N. olumideae BRIP 72283a MZ303791 MZ312684 MZ344176 [22] N. paeoniae–suffruticosae KUMCC 21–0426T OP082292 OP235980 OP204794 [70] N. paeoniae–suffruticosae KUMCC 21–0427 ON426868 OR025933 OR025972 [70] N. pandanicola MFLUCC 22–0144 OP802391 OP801768 OP830890 [57] N. pernambucana RV01T KJ792466 – KU306739 [72] N. perukae FMB 0127 MH209077 MH460876 MH523647 [65] N. petila MFLUCC 17–1738T MK764276 MK764341 MK764319 [53] N. petila MFLUCC 17–1737 MK764275 MK764342 MK764320 [53] N. phangngaensis MFLUCC 18–0119T MH388354 MH412721 MH388390 [57] N. photiniae GUCC 21–0820 OP806524 OP896200 OP828691 [33] N. photiniae GUCC 810T OP498008 OP752131 OP753368 [33] N. piceana CBS 254 32 KM199372 KM199452 KM199529 [1] N. piceana CBS 394 48T KM199368 KM199453 KM199527 [1] N. protearum CBS 114178T JN712498 KM199463 KM199542 [1] N. psidii FMB 0028T MF783082 MH477870 MH460874 [65] N. rhapidis GUCC21501 MW931620 MW980441 MW980442 [34] N. rhizophorae MFLUCC 17–1551T MK764277 MK764343 MK764321 [53] N. rhododendri GUCC 21504T MW979577 MW980443 MW980444 [34] N. rhododendricola KUN–HKAS–123204T OK283069 OK274147 OK274148 [73] N. rosae CBS 101057T KM199359 KM199429 KM199523 [1] N. rosicola CFCC 51992T KY885239 KY885245 KY885243 [74] N. rosicola CFCC 51993 KY885240 KY885246 KY885244 [74] N. samarangensis CBS 115451 KM199365 KM199447 KM199556 [1] N. saprophytica MFLUCC12–0282T JX398982 JX399017 JX399048 [1] N. scalabiensis NJZSN01 OP942341 – – [43] N. scalabiensis GMBUCC 24–002 PP800245 PP889283 PP889285 This study N. scalabiensis GMBUCC 24–003 PP800246 PP889284 PP889286 This study N. sichuanensis SM15–1C MW166232 MW218525 MW199751 [74] N. siciliana AC49 ON117811 ON209164 ON107275 [75] N. siciliana CBS 149117T ON117813 ON209162 ON107273 [75] N. sonneratiae MFLUCC 17–1744 MK764279 MK764345 MN871990 [53] N. sonneratiae MFLUCC 17–1745T MK764280 MK764346 MK764324 [53] N. steyaertii IMI 192475T KF582796 KF582794 KF582792 [1] N. suphanburiensis NGL–2022c isolate TN01T OP497994 OP752135 OP753372 [33] N. surinamensis CBS 450 74 T KM199351 KM199465 KM199518 [1] N. terricola YMD A254/CGMCC 3 23553T OP082294 OP235983 OP204797 [70] N. terricola UESTCC 22.0034 OP082295 OP235983 OP204797 [70] N. thailandica MFLUCC 17–1730T MK764281 MK764347 MK764325 [53] N. thailandica MFLUCC 17–1731 MK764282 MK764348 MK764326 [53] N. umbrinospora MFLUCC12–0285T JX398984 JX399019 JX399050 [1] N. vaccinii 1–22T OQ316613 – OQ342778 [43] N. vacciniicola CAA1055T MW969751 MW934614 MW959103 [43] N. vheenae BRIP 72293aT MZ303792 MZ312685 MZ344177 [22] N. vitis JZB340018T KU140694 KU140685 KU140676 [76] N. vitis JZB340023 KU140699 KU140690 KU140681 [76] N. xishuangbannaensis KUMCC 21–0424T ON426865 OR025934 OR025973 [77] N. xishuangbannaensis KUMCC 21–0425 ON426866 OR025935 OR025974 [77] N. zakeelii BRIP 72282aT MZ303789 MZ312682 MZ344174 [22] Pestalotiopsis trachycarpicola OP068/IFRDCC 2240T JQ845947 JQ845945 JQ845946 [78] Ex-type strains are indicated in 'T'. The strains generated in this study are denoted in red. The phylogenetic analysis was conducted using the maximum likelihood (ML) method, employing RAxML-HPC BlackBox (8.2.4) on the CIPRES portal. The GTR + I + G model of evolution was applied in this analysis[35,36]. Bayesian Inference (BI) was performed using MrBayes v. 3.1.2[37] to evaluate posterior probabilities (PP) by Markov Chain Monte Carlo sampling (MCMC)[38,39]. The posterior probability distribution convergence was ensured by running 1,000,000 generations of Markov chain Monte Carlo (MCMC) with a random seed and a stop value = 0.01, using a MCMC algorithm of four chains. The initial 20% of the sampled data were discarded as burn-in. Phylograms were visualized with FigTree v1.4.0[40] and annotated in Microsoft PowerPoint (2010).
In vitro antifungal activity of Trichoderma atroviride (GMBUCC 24–014) and Daldinia eschscholtzii (UKBC 036) against pestalotioid species using dual culture plate assay
-
Bio–control potential of Trichoderma atroviride (GMBUCC 24–014, Konara et al.[51]) and Daldinia eschscholtzii (UKBC 036, Thambugala et al.[52]) against pestalotioid species was tested. The cultures were obtained from the culture collection of the Genetics and Molecular Biology Unit, University of Sri Jayewardenapura (GBMUCC), and the Culture Collection of the University of Kelaniya (UKBC), Sri Lanka. PDA plates were divided into two portions. Fungal plugs of 4 mm diameter from the isolated fungal phytopathogens (GMBUCC 24–001, GMBUCC 24–002, and GMBUCC 24–003) and the bio–control agents were placed side by side in the two separated portions, known as test plates (Fig. 1). They were triplicated for each pathogen. Plates containing pathogenic fungi without bio–control agents were used as control plates. Both control and test plates were incubated for 10 d at room temperature (25 ± 2 °C). The percentage of inhibition of fungal growth was calculated using the following formula proposed by Imtiaj & Lee[41]:
$ \mathrm{I}{\text{%}} =\dfrac{{\rm r}_{1}-{\rm r}_{2}}{\rm r_1} \times{100} $ where, r1 is the radial growth of the isolated Neopestalotiopsis species in the control plates (cm), and r2 is the radial growth of the Neopestalotiopsis species in the test plates (cm).
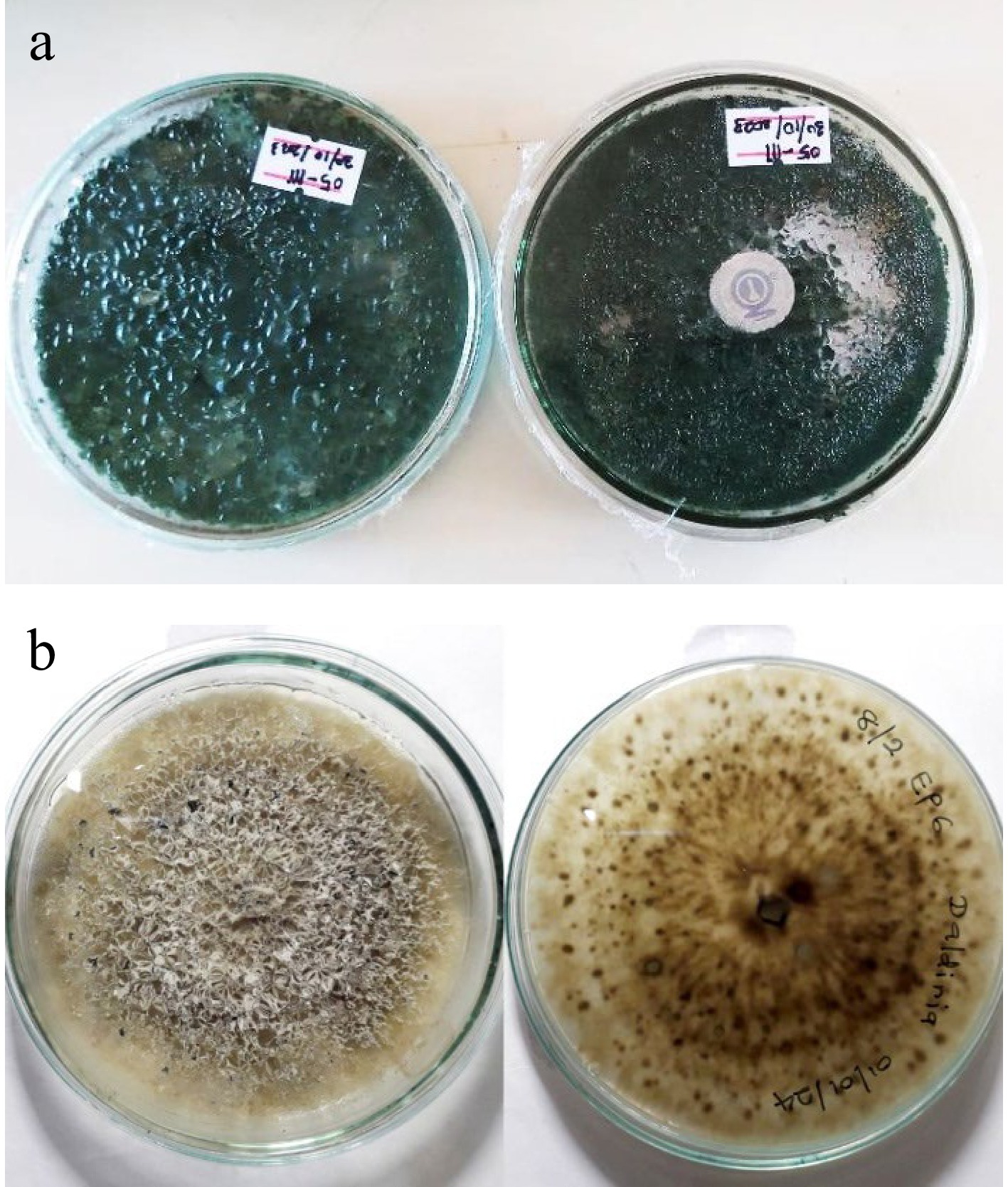
Figure 1.
Pure cultures of bio–control agents used in this study. (a) Trichoderma atroviride (GMBUCC 24–014) isolated from the basidiocarp of Ganoderma angustisporum. (b) Daldinia eschscholtzii (UKBC 036) isolated from the leaves of Camellia sinensis as an endophyte.
-
The combined data set of ITS, tub2, and tef1 sequences comprised 1500 characters with gaps. The RAxML analysis of the combined dataset yielded a best-scoring tree (Fig. 2) with a final ML optimization likelihood value of –8738.944811. The matrix had 736 distinct alignment patterns, with 15.36 % of undetermined characters or gaps. Estimated base frequencies: A = 0.231055, C = 0.266487, G = 0.215875, T = 0.286583; substitution rates AC = 1.042093, AG = 3.330012, AT = 1.383087, CG = 0.619927, CT = 4.040718, GT = 1.000000; proportion of invariable sites I = 0.395452; gamma distribution shape parameter α = 0.657974. The Bayesian analysis resulted in 10,000 trees after 1,000,000 generations. All analyses (ML and BYPP) gave similar results and were in agreement with previous study based on multigene analyses[33].

Figure 2.
Phylogram obtained based on ITS, tub2, and tef1 sequencing data. The tree is rooted with Pestalotiopsis trachycarpicola (OP068). The bootstrap support values for maximum likelihood (≥ 50%) and Bayesian posterior probabilities (≥ 0.95) are shown at the nodes. The type strains are indicated in bold. Newly generated strains are in red.
Taxonomy
-
Neopestalotiopsis cubana Maharachchikumbura SSN, K.D. Hyde & Crous, Studies in Mycology 79: 138 (2014) [MB#809765] Fig. 3.

Figure 3.
Neopestalotiopsis cubana – (GMBUCC 24–001). (a) Leaf lesions on the host. (b), (c) Mycelial growth on PDA above and reverse. (d) Colony sporulating on PDA. (e) Conidiogenesis. (f)–(h) Conidia with appendages.
Associated with leaf lesions of G. mangostana, black spots emerge from the plant epidermis. Sexual morph: Undetermined. Asexual morph: Conidiomata pycnidial in culture on PDA, globose, solitary or aggregated, embedded or semi–immersed, dark brown to black, exuding globose, brown to black conidial masses. Conidiophores are reduced to conidiogenous cells. Conidiogenous cells 5–12 × 2–8 μm (
$ \overline{x} $ $ \overline{x} $ $ \overline{x} $ $ \overline{x} $ Culture characteristics: Colonies on PDA reaching 30–40 mm diameter after 7 d at room temperature (28 °C), with a lobate edge, pale honey–colored, with sparse aerial mycelium on the surface with black, gregarious conidiomata; reverse similar in color.
Known distribution: Cuba[1], Sri Lanka (this study).
Material examined: Sri Lanka, Western province, Gampaha District, Kadawatha, associated with fresh leaves of Garcinia mangostana (Mangosteen); 22 August 2022; Collector – K.M. Thambugala; USJ–GMBU–007; Culture – GMBUCC 24–001.
Notes: In the present phylogeny, the new isolate N. cubana (GMBUCC 24–001) clustered with N. cubana (CBS 600.96) from Cuba[23], suggesting that they are closely related (Fig. 2, Clade A). Clade A contains N. cubana CBS 600.96 (ex-type), LHHN37, LHHN42, GMBUCC 24–001 and N. pandanicola (MFLUCC 22–0144). When compared morphologically, our collection of N. cubana (GMBUCC 24–001) exhibits a slightly smaller conidial size and basal appendage length than N. cubana (CBS 600.96). Specifically, the conidial size of N. cubana (CBS 600 96) measures 22–31 × 7.5–9.0 μm, whereas N. cubana (GMBUCC 24–001) measures 19–27 × 5.5–8.5 μm. The basal appendage length of N. cubana (CBS 600.96) is 22–66 μm, whereas 20–50 μm for N. cubana (GMBUCC 24–001). Additionally, our collection has the same number of apical appendages, around 2–4 compared to N. cubana (CBS 600 96)[1]. In other conidial morphologies and colony characteristics, both strains exhibit similar patterns.
When compared to N. cubana (CBS 600.96), N. pandanicola (MFLUCC 22–0144) exhibits significantly different sizes of conidia and apical appendages. Specifically, the conidial size of N. cubana (CBS 600.96) measures 22–31 × 7.5–9.0 μm, whereas N. pandanicola (MFLUCC 22–0144) measures 16–25 × 4–7 μm. The apical appendage length of N. cubana (CBS 600 96) is 22–66 μm, while it is 27–30 μm for N. pandanicola (MFLUCC 22–0144). Additionally, basal appendage length is 6–11 μm long for N. cubana (CBS 600 96), and 3–7 μm for N. pandanicola (MFLUCC 22–0144). In addition to that, N. pandanicola (MFLUCC 22–0144) has a slight difference in apical appendages with 2–3 compared to 1–2 in N. cubana (CBS 600.96)[1,12,42]. In other conidial morphologies and colony characteristics, both strains exhibit similar patterns.
Neopestalotiopsis scalabiensis J. Santos, S. Hilário & A. Alves, Eur. J. Pl. Pathol. 162: 547 (2021) [MB#841333] Fig. 4.
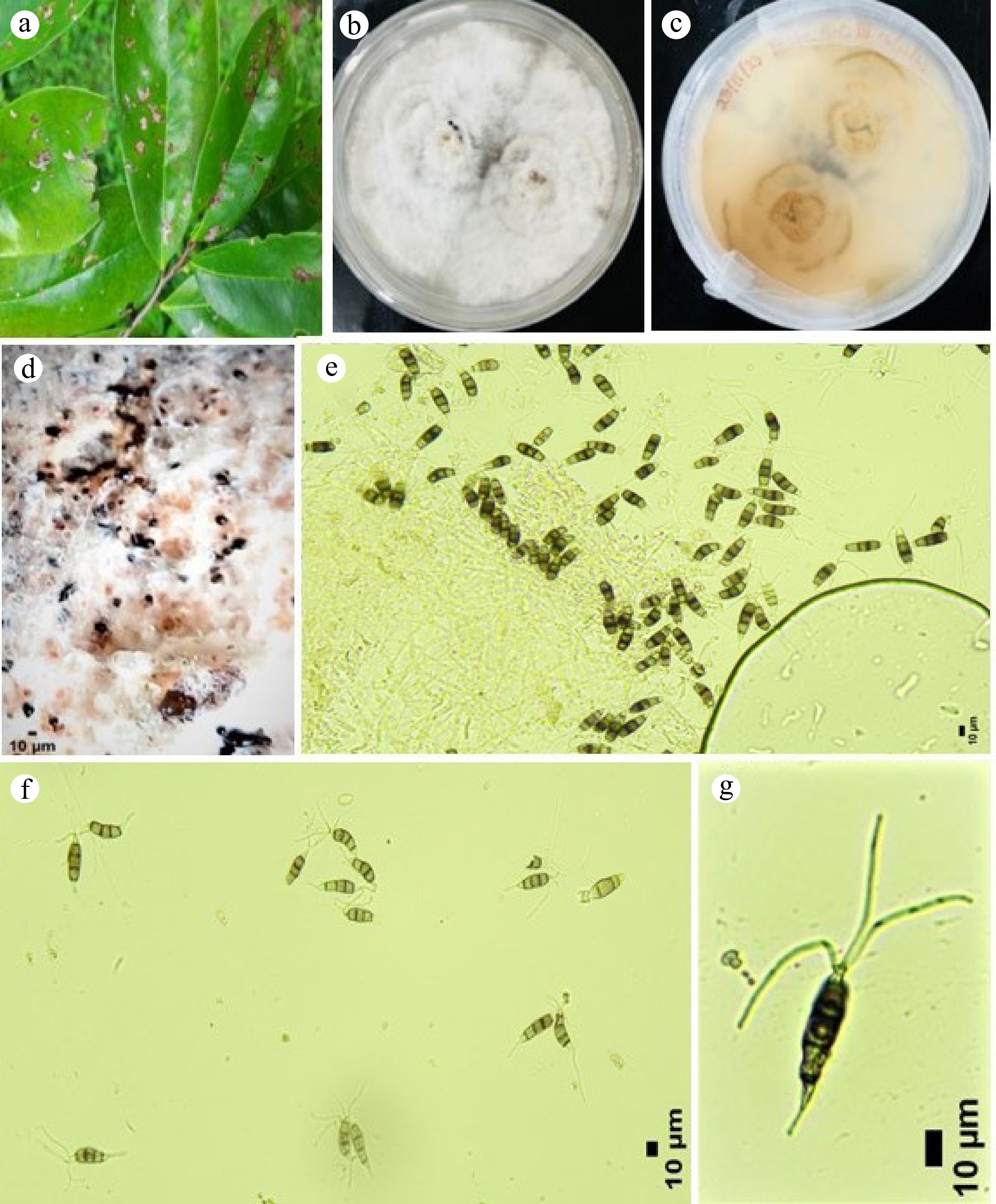
Figure 4.
Neopestalotiopsis scalabiensis – (GMBUCC 24–002 and GMBUCC 24–003). (a) Leaf lesions on the host. (b), (c) Mycelial growth on PDA above and reverse. (d) Colony sporulating on PDA (conidiomata). (e) Conidiogenesis. (f), (g) Conidia with appendages.
Associated with small leaf lesions of Annona muricata, appear as black spots emerging from the plant epidermis surface. Sexual morph: Undetermined. Asexual morph: Conidiomata on PDA pycnidial, globose, solitary, semi–immersed, black, exuding globose, dark conidial masses; Conidiophores rare 15–27 × 3–4 μm (
$ \overline{x} $ $ \overline{x} $ $ \overline{x} $ $ \overline{x} $ $ \overline{x} $ $ \overline{x} $ Culture characteristics: Pycnidia developing on fennel twigs and pine needles, 70–90 mm diameter, after one week at room temperature on PDA. Colonies spreading on PDA with cottony flattened mycelium growing in a radial pattern with white cottony tufts resembling waves foam. Developing rare solitary black conidiomata with age and reverse white to snow white.
Known distribution: Portugal[43], Sri Lanka (this study)
Material examined: Sri Lanka, Western Province, Gampaha District, Kadawatha, associated with leaves of Annona muricata (Soursop); 22 August 2023; Collector – K.M. Thambugala; USJ–GMBU–008, and USJ–GMBU–009; Cultures– GMBUCC 24–002 and GMBUCC 24–003.
Notes: In this study, the strain, GMBUCC 24–002, isolated from A. muricata in Sri Lanka, clustered together with N. scalbiensis (NJZSN01)[29] from Vaccinium corymbosum in Portugal, indicating that they are closely related (ML/MP/PP = 82/1.00) (Fig. 2, Clade B). The strain, GMBUCC 24–003, isolated from A. muricata, clustered together with N. egyptiaca (CBS 140162)[44], indicating that they are closely related (ML/MP/PP = 81/1.00) (Fig. 2, Clade B). The current collections of N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) exhibit shorter basal appendages, measuring 2–10 μm, compared to N. scalabiensis (NJZSN01), which has appendages ranging from 2.1–17 μm. This difference in basal appendage length may be attributed to their distinct geographical origins. Furthermore, these collections of N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) exhibit longer basal appendages, measuring 2–10 μm, compared to N. egyptiaca (CBS 140162), which has appendages ranging from 1.5–5.5 μm and longer apical appendages measuring 6.5–30 μm compared to N. egyptiaca (CBS140162), which has appendages ranging from 11-21 μm. Despite these variations, both strains exhibit similar conidial morphologies and colony characteristics, indicating a close relationship between the Sri Lankan collections (GMBUCC 24–002 and GMBUCC 24–003), the collections from Portugal (N. scalabiensis – NJZSN01) and Egypt (N. egyptiaca – CBS140162).
Dual culture assay
-
Trichoderma atroviride (GMBUCC 24–014) as the biocontrol agent (Tables 2 & 3).
Table 2. In vitro biocontrol estimation of T. atroviride (GMBUCC 24–014) against N. cubana (GMBUCC 24–001) associated with the leaves of G. mangostana.
Day Control (r1/cm) T1 (r2/cm) T2 (r2/cm) T3 (r2/cm) Average (r2\cm) I% 3 0.5 0.3 0.5 0.3 0.37 26.0 5 1.5 1.1 1 1.2 1.1 26.7 7 1.7 1.1 1 1.2 1.1 35.3 10 2 1.2 1 1.2 1.1 45.0 Table 3. In vitro biocontrol estimation of T. atroviride (GMBUCC 24–014) against N. scalabiensis (GMBUCC 24–002, and GMBUCC 24–003) associated with the leaves of A. muricata.
Date Control (r1/cm) T1 (r2/cm) T2 (r2/cm) T3 (r2/cm) Average (r2/cm) I% 3 1.5 0.5 1 1.3 0.93 38.0 5 2 0.5 1.5 1.5 1.16 42.0 7 2.5 0.7 1.5 1.5 1.23 50.8 10 3 0.7 1.5 1.5 1.23 59.0 r1: growth radian of the pathogen (Neopestalotiopsis species) without biocontrol agent (T. atroviride), r2: growth radian of the pathogen (Neopestalotiopsis species) in the presence of biocontrol agent (T. atroviride), C: control, T1/T2/T3: test replicates, I: inhibition percentage. Daldinia eschscholtzii (UKBC 036) as the biocontrol agent (Tables 4 & 5).
Table 4. In vitro biocontrol estimation of D. eschscholtzii (UKBC 036) against N. cubana (GMBUCC 24–001) associated with the leaves of G. mangostana.
Day Control
(r1/cm)T1
(r2/cm)T2
(r2/cm)T3
(r2/cm)Average
(r2\cm)I% 3 0.5 0.3 0.6 0.5 0.46 8.0 5 1.5 1.1 1.3 1 1.13 24.6 7 1.7 1.1 1.5 1.2 1.26 25.8 10 2 1.2 1.5 1.2 1.3 35.0 Table 5. In vitro biocontrol estimation of D. eschscholtzii (UKBC 036) against N. scalabiensis (GMBUCC 24–002, and GMBUCC 24–003) associated with the leaves of A. muricata.
Date Control
(r1/cm)T1
(r2/cm)T2
(r2/cm)T3
(r2/cm)Average
(r2/cm)I% 3 1.5 1 1.3 1 1.1 26.0 5 2 1.2 1.6 1.5 1.36 32.0 7 2.3 1.5 2.0 1.5 1.3 33.6 10 2.5 1.5 2.0 1.5 1.3 46.7 r1: growth radian of the pathogen (pestalotioid species) without biocontrol agent (Daldinia eschscholtzii), r2: growth radian of the pathogen (pestalotioid species) in the presence of biocontrol agent (Daldinia eschscholtzii), C: control, T1/T2/T3: test replicates, I: inhibition percentage. -
Phytopathogenic fungi have been the primary focus of mycological research in Sri Lanka to date[26,45,46]. This emphasis arises from the significant impact of these fungal pathogens on plants and the resulting economic losses. Numerous investigations have been conducted to clarify the variety of pestalotioid species associated with various plants in Sri Lanka[1,47,79]. This study revealed the first record of Neopestalotiopsis species associated with G. mangostana and A. muricata in Sri Lanka. Fruits of Manilkara zapota infected by Pestalotiopsis microspora were first identified in Sri Lanka[48]. Recently, Weerasekara et al.[79] revealed multiple species of Pseudopestalotiopsis associated with grey blight in tea crops [Camellia sinensis (L.) O. Kuntze] in Sri Lanka. Among them, the currently known species of Ps. daweiana, Ps. annellata, and Ps. chinensis were identified based on the molecular data derived from ex-type isolates. Additionally, four new species of Pseudopestalotiopsis viz Ps. petchii, Ps. ratnapurensis, Ps. rossmaniae, and Ps. srilankensis were introduced. Nevertheless, it is evident that economically significant plants such as G. mangostana and A. muricata are also affected by pestalotioid pathogens.
In this study, three gene regions—ITS, tub2, and tef1-α were used for PCR amplification. These universal primers were selected to offer substantial infra– and interspecific variation that makes them effective for species identification and phylogenetic analysis[49]. While this study focused on these three gene regions, future research could benefit from incorporating large subunit rRNA (LSU) and RNA polymerase II second-largest subunit (RPB2) to enhance phylogenetic resolution.
The results revealed the presence of Neopestalotiopsis species—N. cubana (GMBUCC 24–001) associated with G. mangostana leaves and N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) associated with A. muricata leaves. Phylogenetic analysis indicates that these Neopestalotiopsis species from G. mangostana and A. muricata do not form closely related groups, as they are placed in different clades (Fig. 2, Clades A and B). This divergence may be attributed to host specificity. Notably, N. cubana (GMBUCC 24–001) clusters as a sister taxon to N. cubana (CBS 600 96) from Cuba[1], suggesting a close relationship (Fig. 2, Clade A). Despite there being some variations, both isolated strains of N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) exhibit similar conidial morphologies and colony characteristics, indicating a close relationship with the collections from Portugal (N. scalabiensis – NJZSN01) and Egypt (N. egyptiaca – CBS140162) (Fig. 2, Clade B). Further clarification of this species' phylogenetic position would require a detailed examination of morphological characteristics and the inclusion of additional molecular markers.
It is important to note that we did not include pathogenicity testing in this study to confirm the disease-causing potential of the Neopestalotiopsis species isolated from A. muricata and G. mangostana. While Koch's postulates were not fulfilled in the study, the isolates were included in an in vitro evaluation based on the well-documented pathogenic potential of several Neopestalotiopsis species across diverse host plants. However, we recognize that pathogenicity tests are essential to establish a definitive causal relationship between fungal isolates and host symptoms. As such, our findings should be considered preliminary and not conclusive regarding the role of isolates as pathogens. Future work will focus on conducting pathogenicity assays and in planta biocontrol trials to validate the in vitro observations and evaluate the practical application of these fungal biocontrol agents in disease management.
Fungal biocontrol agents not only prevent diseases but also promote plant growth, enhance nutrient utilization efficiency, strengthen plant resistance, and mitigate agrochemical pollution in the environment[50]. In the dual culture assay, both T. atroviride (GMBUCC 24–014) and Daldinia eschscholtzii (UKBC 036) showed significant percentages of inhibition against N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) which is around 59% and 47% respectively (Fig. 5). In addition to that, T. atroviride (GMBUCC 24–014) shows higher biocontrol potential over D. eschscholtzii (UKBC 036) on N. cubana (GMBUCC 24–001), and N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003). This suggests that T. atroviride (GMBUCC 24–014) can be a potential biocontrol agent against N. cubana (GMBUCC 24–001), and N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) compared to D. eschscholtzii (UKBC 036).
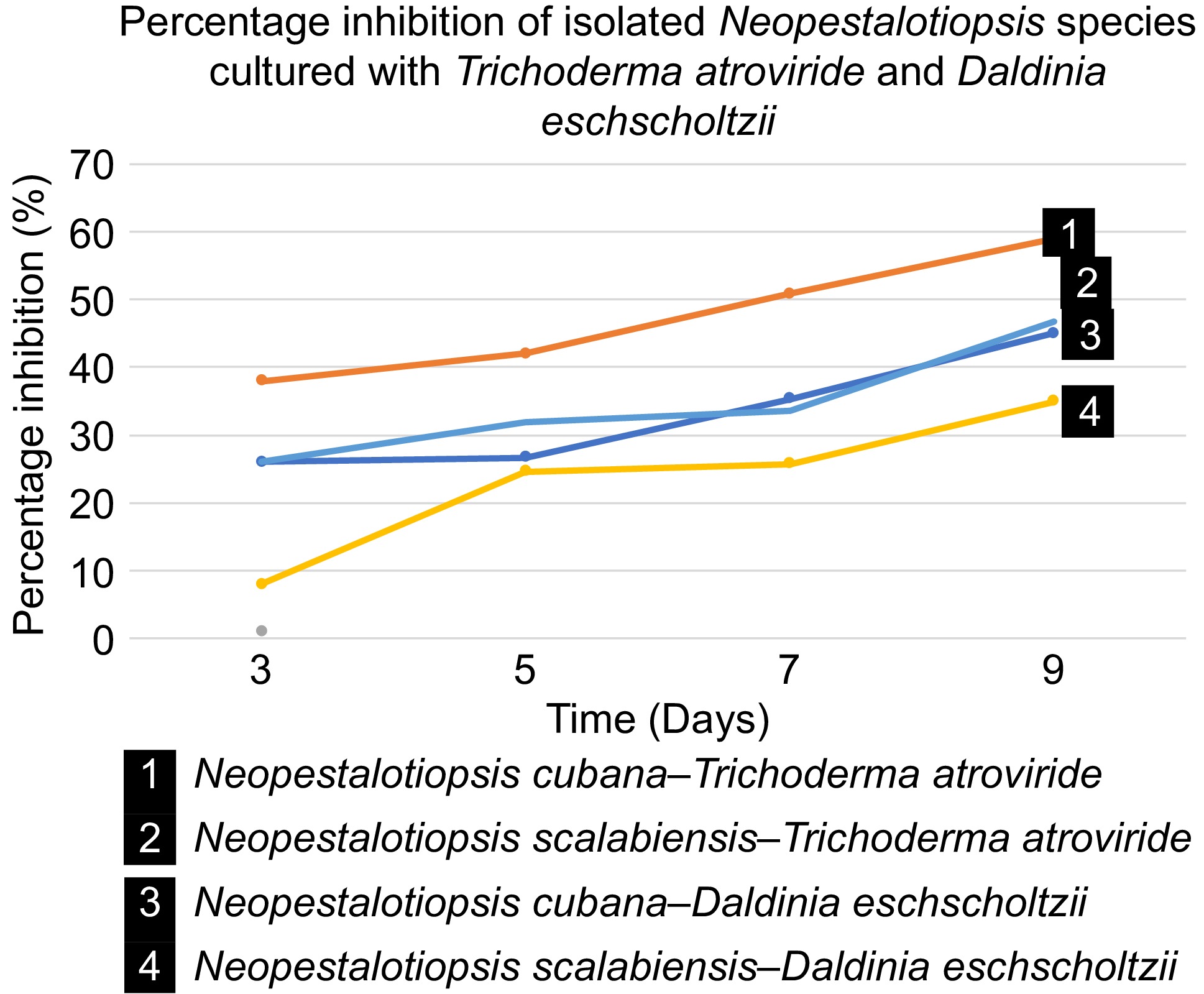
Figure 5.
In vitro biocontrol estimation of T. atroviride (GMBUCC 24–014) and Daldinia eschscholtzii (UKBC 036) against the isolated species (N. cubana – GMBUCC 24–001; N. scalabiensis – GMBUCC 24–002, and GMBUCC 24–003).
The structural complexity and biological activity of the secondary metabolites produced by pestalotioid species have drawn attention to them. Several new secondary metabolites have been isolated and discovered in recent decades[7]. Their bioactivities, which include anticancer, antifungal, antibacterial, and nematocidal activity, as well as medical, agricultural, and industrial applications, were investigated[1,2,7]. It would be beneficial to identify the symbiont pathogens with pestalotioid species since their bioactive metabolites can also have inhibitory action against pestalotioid species.
-
This study has significantly advanced the understanding of the fungi associated with leaf lesions of G. mangostana and A. muricata plants from Sri Lanka, identifying two Neopestalotiopsis species. For the first time, N. cubana (GMBUCC 24–001) from G. mangostana, and N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) from A. muricata reported. The presence of various Pestalotioid fungi indicates that the leaf lesions of G. mangostana and A. muricata are diverse, demanding further research to establish the fungal communities associated with the disease. These insights are crucial for developing better biocontrol agents and management practices in this plant cultivation. T. atroviride (GMBUCC 24–014) shows higher biocontrol potential over Daldinia eschscholtzii (UKBC 036); this suggests that T. atroviride (GMBUCC 24–014) can be a potential biocontrol agent against both isolated species compared to Daldinia eschscholtzii (UKBC 036).
The research was supported by the Center for Plant Materials and Herbal Products Research, University of Sri Jayewardenepura, Nugegoda 10250, Sri Lanka; Rubber Research Institute, Sri Lanka (RRISL), Dartonfield, Agalawatta, Sri Lanka; Centre for Biotechnology (CBT), Department of Zoology, Center for Forestry and Environment, Department of Forestry and Environment Sciences, Faculty of Applied Sciences, University of Sri Jayewardenepura; and the Department of Plant and Molecular Biology, Faculty of Science, University of Kelaniya, Kelaniya 11300, Sri Lanka.
-
The authors confirm contribution to the paper as follows: conceptualization, data curation, and formal analysis: Asfa F, Thambugala KM, de Silva NI; funding acquisition: Liyanage KK, Thambugala KM, Tennakoon S; investigation, methodology, and writing − original draft: Asfa F, Thambugala KM, Daranagama DA, Tennakoon S; supervision: Thambugala KM; writing, review, and editing: Asfa F, Thambugala KM, de Silva NI, Daranagama DA. All authors reviewed the results and approved the final version of the manuscript.
-
The data generated and analyzed during this study are available in this article. DNA sequence data are available in the GenBank database, and the accession numbers are provided in Table 1. ITS: PP800244, PP800245, PP800246, PP859452; tub: PP889282, PP889283, PP889284; tef1-α: PP889285, PP889286.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Asfa F, Thambugala KM, Daranagama DA, de Silva NI, Liyanage KK, et al. 2025. New records of Neopestalotiopsis species associated with leaves of Annona muricata (Soursop) and Garcinia mangostana (Mangosteen) in the Western Province, Sri Lanka, and in vitro evaluation of selected biocontrol agents for their growth suppression. Studies in Fungi 10: e010 doi: 10.48130/sif-0025-0010 

New records of Neopestalotiopsis species associated with leaves of Annona muricata (Soursop) and Garcinia mangostana (Mangosteen) in the Western Province, Sri Lanka, and in vitro evaluation of selected biocontrol agents for their growth suppression
- Received: 26 February 2025
- Revised: 06 May 2025
- Accepted: 06 May 2025
- Published online: 18 June 2025
Abstract: Most pestalotioid species are phytopathogens characterized by conidia with appendages and can also function as endophytes and saprobes. During a morpho-molecular study of plant-inhabiting micro-fungi in Sri Lanka, two pestalotioid species were isolated from the lesions of living leaves of Garcinia mangostana and Annona muricata. Maximum Likelihood and Bayesian Inference analyses of combined internal transcribed spacers (ITS), β-tubulin (tub2), and translation elongation factor 1-α (tef1-α) sequence data were used to determine the phylogenetic affinities of the collected species. Detailed morphological and multi-locus phylogenetic analyses further validated that the collected pestalotioid species belonged to Neopestalotiopsis. Neopestalotiopsis cubana (GMBUCC 24–001) and N. scalabiensis (GMBUCC 24–002 and GMBUCC 24–003) were identified as associated with the fresh leaves of G. mangostana and A. muricata, respectively. This is the first record of the Neopestalotiopsis species associated with these hosts worldwide. This study evaluated the in vitro biocontrol potential of Trichoderma atroviride (GMBUCC 24–014) and Daldinia eschscholtzii (UKBC 036) against Neopestalotiopsis cubana and N. scalabiensis, resulting in significant biocontrol potential of T. atroviride (GMBUCC 24–014) against both species than D. eschscholtzii (UKBC 036). Accurate identification of pestalotioid species is crucial for developing effective biocontrol methods to manage diseases and enhance economic development.
-
Key words:
- Dual culture plate assay /
- Morphology /
- Pestalotioid taxa /
- Multi-locus phylogeny /
- Phytopathogens