









-
As a major global cash crop, melon plays a vital role in agricultural economies worldwide. Among its key market attributes, flesh coloration serves as a primary quality indicator that significantly influences consumer preference and marketability, making it a key focus of agricultural research and breeding programs[1]. The common flesh colors of melons are primarily white, green, yellow, and orange[2]. Research has demonstrated that carotenoids and chlorophyll are the main pigments determining melon flesh color, with orange-fleshed melons being particularly rich in carotenoids, mainly β-carotene[1,3,4]. Currently, over 800 natural carotenoids have been identified in higher plants, animals, fungi, and other organisms[5]. Carotenoids are isoprenoid hydrophobic compounds composed of a C40 hydrocarbon skeleton and are categorized into two primary groups: carotenoids and lutein. Carotenoids possess long conjugated double-bond chains that can contain up to 15 conjugated double bonds. The greater the number of conjugated double bonds, the more intense the red hue of the carotenoid[6,7].
Isoprenoid diphosphate (IPP) serves as a precursor substance for carotenoid synthesis in higher plants[8,9]. However, the IPP synthesis pathway in plants is inconsistent, with two primary pathways: cytoplasmic and plastidic[10,11]. In melons, carotenoids are predominantly synthesized via the plastid pathway[1,12,13]. Within the plastid, a molecule of glyceraldehyde 3-phosphate combines with a molecule of pyruvate and undergoes a series of catalytic reactions, ultimately generating IPP[14,15]. Three molecules of IPP and its isomer dimethylallyl diphosphate (DMAPP) condense through a series of catalytic reactions to form geranylgeranyl pyrophosphate (GGPP), which contains 20 carbon atoms. GGPP serves as a direct precursor substance for the synthesis of carotenoids (carotenes) and various isoprenoids, including gibberellins, chlorophylls, tocopherols, chlorophyll quinones, and plastoquinones[16]. Two molecules of GGPP are condensed by phytoene synthase (PSY) to form colorless phytoene, which subsequently undergoes dehydrogenation by phytoene desaturase (PDS) to form ζ-carotene and continues to dehydrogenate, ultimately forming the red pigment lycopene[2,17,18].
Lycopene represents a crucial branching point in the anabolism of carotenoids, with two main branches of the cyclisation pathway: ε-cyclisation and β-cyclisation[19,20]. Lycopene ε-cyclase (LCYE) catalyzes the conversion of lycopene to σ-carotene, which is further catalyzed by lycopene β-cyclase (LCYB) and LCYE to form α-carotene and ε-carotene. Subsequently, α-carotene is metabolized to form lutein[21]. In an alternative pathway, lycopene undergoes β-cyclisation to form γ-carotene, followed by the LCYB-catalyzed formation of β-carotene[8,21]. Carotenoid cleavage dioxygenases (CCDs) catalyze the metabolism of β-carotene to various deuterocarotenoids[22,23]. Previous studies have demonstrated that CCD4 determines the accumulation of pigmented substances in plant flowers and fruits[24−27]. Furthermore, the hydroxylation of β-carotene generates zeaxanthin, ultimately synthesizing violaxanthin, the precursor substance of abscisic acid (ABA). The final metabolism of violaxanthin to ABA is catalyzed by 9-cis-epoxycarotenoid dioxygenases (NCEDs)[8,13].
Carotenoids are vital compounds that impart vibrant colors to plants, particularly in melon, where they are considered the primary determinants of flesh color[1,2,28]. The content of carotenoids exhibits significant variation across species and even within the same species[28]. For instance, in melon, watermelon, tomato, carrot, and cauliflower, the diverse fruit colors are primarily derived from the degree of lycopene and β-carotene accumulation[1,29−32]. Previous studies have molecularly characterized two pivotal genes, CmOr and CmPPR1, both functioning within the carotenoid biosynthetic pathway, as critical regulators governing melon flesh coloration[33,34]. Studies have demonstrated that the CmOr gene enhances β-carotene accumulation in orange-fleshed tissues by modulating chromoplast biogenesis and repressing metabolic flux through β-carotene downstream pathways[35]. At the molecular level, the melon OR protein facilitates β-carotene biosynthesis through specific interaction with PSY1, maintaining enzymatic stability via post-translational regulation, thereby enhancing metabolic flux through this biosynthetic pathway[36]. Phylogenetic analyses have functionally validated the cross-species conservation of this regulatory mechanism, with experimental evidence spanning model organisms including Arabidopsis, sweetpotato, tomato, and pepper[18]. Accumulating evidence suggests that differential expression of carotenoid pathway genes drives β-carotene accumulation, which likely serves as the central mechanism underlying orange flesh formation in melon fruits. Furthermore, several transcription factors, such as MAD[37−39], bHLH[40,41], NAC[42], MYB[43,44], WRKY[45,46], and zinc-finger proteins[38], also play crucial regulatory roles in carotenoid anabolism. In melon, CmWRKY and CmNAC are involved in carotenoid biosynthesis by activating the expression of CmPSY1 and CmLCYB[2].
Carotenoids are crucial compounds that determine the color of melon flesh; however, there is a paucity of research on the accumulation and regulatory mechanisms of carotenoids in melon fruit. In this study, a combined analysis of targeted metabolome and transcriptome was conducted on melons with varying fruit colors at different developmental stages. This comprehensive approach systematically investigated the accumulation and regulation of carotenoids in melon fruits, providing initial insights into the mechanisms underlying melon fruit color formation. The findings of this study establish a theoretical foundation for further research on carotenoid biosynthesis and regulation in melon, while also offering valuable theoretical knowledge and genetic resources for melon breeding and quality improvement.
-
This study utilized white-fleshed 'TianYu' and orange-fleshed 'TianJu' melon materials, both of which were high-generation self-crosses developed by the Watermelon and Melon Research Team of Northwest A&F University, Yangling, Shaanxi, China (Cucumis melo var. inodorus). The melons underwent color change 30 d after pollination and reached maturity 45 d post-pollination. 'TianYu' shifted from light green to white, whereas 'TianJu' transitioned from light green to orange upon ripening. The seeds were soaked, germinated, and sown in substrate, with plants cultivated at the Cucurbit Research and Experimental Station, College of Horticulture, Northwest A&F University, Yangling, Shaanxi, China[47], upon reaching the two-leaf stage. The plants received standard water and fertilizer management and were hand-pollinated during flowering, maintaining one fruit per plant. Female flowers were labeled on the day of anthesis, and pulp samples from 'TianYu' and 'TianJu' were collected 15 d (before color change stage) and 30 d (color change stage) after pollination, following photographic documentation. The pulp samples were promptly frozen in liquid nitrogen and stored at −80 °C for subsequent analysis. At each developmental stage, five biological replicates were collected by harvesting one fruit from individual plants. From each fruit, five pulp samples were obtained and pooled to create a composite sample.
Determination of carotenoid content
-
In this study, the carotenoid content of 'TianYu' and 'TianJu' fruit samples at different fertility stages was determined using the method described by Medeiros et al.[48]. The pulp samples, preserved under ultracold conditions, were ground into a fine powder using a grinder. A 0.5-mL extraction solution containing 0.01% BHT (g/mL) (1:1:1 hexane : acetone : ethanol mixture, v/v) was added to 50 mg of the powdered sample. The mixture was vortexed for 20 min at room temperature, followed by centrifugation for 5 min (4 °C, 12,000 rpm). The extraction process was repeated, and the supernatants from both extractions were combined. The extract was then concentrated and re-solubilized with 100 μL of a methanol : methyl tert-butyl ether mixture (1:1, v/v), filtered through a 0.22-μm membrane, and stored in a brown vial for liquid chromatography tandem mass spectrometry (LC-MS/MS) analysis using a YMC-C30 liquid chromatography column. Carotenoid quantification was performed using an HPLC-based analytical platform with authentic standards for 11 metabolites: lutein (CAS:127-40-2), β-carotene (CAS:7235-40-7), violaxanthin (CAS:126-29-4), neoxanthin (CAS:14660-91-4), β-cryptoxanthin (CAS:472-70-8), zeaxanthin (CAS:144-68-3), (E/Z)-phytoene (CAS:13920-14-4), lycopene (CAS:502-65-8), 8'-apo-β-carotenal (CAS:1107-26-2), echinenone CAS: (432-68-8), and canthaxanthin (CAS:514-78-3). The assay was performed in three technical replicates. A Metware Database (MWDB) was constructed based on standards for qualitative analysis of the mass spectrometry data. Quantitative analysis was performed using the Multiple Reaction Monitoring (MRM) mode of triple quadrupole mass spectrometry. The mass spectrometry data were processed using Analyst 1.6.3 and MultiQuant 3.0.3 software to obtain the final carotenoid content data.
RNA extraction and transcriptome sequencing of melon pulp at different fertility stages
-
Fruit pulp samples were collected from 'TianYu' and 'TianJu' on the 15th and 30th d after pollination, respectively. In this study, total RNA was extracted from pulp samples using the TIANGEN RNAprep Pure kit (DP432) from Tiangen Biotech Co., Ltd. (Beijing, China). Pulp samples were lysed in strict accordance with the kit instructions, with RNA being purified using specific columns and eluted using RNase-free ddH2O. The purity and concentration of the extracted RNA were assessed using the NanoDrop Ultra-Micro UV Spectrophotometer (Agilent Technologies, Santa Clara, USA). RNA integrity was tested using agarose gel electrophoresis (0.8%, 120 V, 400 mA) to ensure that the extracted RNA was suitable for subsequent sequencing. Shanghai Luming Biotechnology Co., Ltd. (Shanghai, China) was commissioned to construct the cDNA library, and the Illumina NovaSeq 6000 platform was employed for bidirectional sequencing of the library and acquisition of raw data. Final clean reads were obtained after quality control of the raw data, and the sequence data were subsequently aligned to the melon reference genome (DHL92, V3.6.1,
http://cucurbitgenomics.org/v2/organism/23 , accessed on 30th September 2024). Subsequently, reads that were mapped less than 10 times, those mapped with more than one genomic region, and those not mapped with the reference genome were removed. Finally, the quantitative expression level of each gene was obtained and analyzed for expression level and functional annotation[47].Differentially expressed gene identification and functional enrichment analysis
-
In this study, RNA sequencing analyses of 'TianYu' before color change stage (TianYu_W) and within color change stage (TianYu_O), as well as 'TianJu' before color change stage (TianJu_W) and within color change stage (TianJu_O) were performed. Four comparison groups were established: TianYu_O vs TianYu_W, TianJu_W vs TianYu_W, TianJu_O vs TianJu_W, and TianJu_O vs TianYu_O. The software DESeq2 (R package, V1.20.0) was employed for comparative combinatorial differentially expressed gene (DEG) analyses, with the p-value (padj) adjusted to control the false discovery rate using the Benjamini and Hochberg method[49]. The ratio of expression between the treatments within a comparison was represented by the fold change provided in the differential expression data. Genes with |log2(foldchange)| ≥1 and Padj ≤ 0.05 were designated as DEGs.
The GO enrichment analysis of the DEGs was conducted utilizing the Wallenius non-central hypergeometric distribution-based GOseq R package[47]. The GO annotation proteome employed in the analysis was obtained from the UniProt-GOA database (
www.ebi.ac.uk/GOA , accessed on 30th September 2024). The KEGG pathway enrichment analysis of the DEGs was performed using the KOBAS V3.0 software (http://bioinfo.org/kobas/download/ , accessed on 30th September 2024).Quantitative real-time PCR (qRT-PCR) and expression validation
-
To verify the transcriptome results, a set of genes was selected for qRT-PCR analysis. cDNA synthesis from RNA for transcriptome sequencing was performed using the TIANGEN FastKing cDNA First Strand Synthesis Kit (KR116) from Tiangen Biotech Co., Ltd. (Beijing, China). Primers were designed using Primer Premier 6.0 software, and synthesized by Tsingke Biotechnology Co., Ltd. The relative expression of candidate genes was calculated by utilizing the housekeeping melon gene, Cmactin2 as the internal reference. All primer information is listed in Supplementary Table S1. The qRT-PCR reaction consisted of 1 µL of cDNA template (80 ng/µL), 0.5 µL of each primer, 10 µL of 2 × SYBR qPCR Mix, and 8 µL of ddH2O, yielding a total volume of 20 µL. The qRT-PCR amplification conditions were set to 94 °C for 3 min, followed by 40 cycles of 94 °C for 10 s and 60 °C for 30 s. Three technical replicates were performed for each sample, and relative expression was calculated using the 2−ΔΔCᴛ method[47].
Data analysis
-
The data were analyzed using IBM-SPSS V19 software, which facilitated the examination of mean variations at different confidence levels. To determine statistical significance, the researchers employed analysis of variance (ANOVA) and Student's t-test methods.
-
Both 'TianYu' and 'TianJu' are classified as thick-skinned melon varieties, whose fruit ripen 45 d after pollination. Before the onset of color change, the flesh of both 'TianYu' and 'TianJu' exhibited a light green hue (Fig. 1a & c). As the fruit continued to develop, both varieties commenced the process of flesh color alteration 30 d post-pollination. During the color change stage, the flesh of 'TianYu' fruit underwent a transition from light green to white, whereas the flesh of 'TianJu' fruit gradually shifted from light green to an orange coloration (Fig. 1b & d).
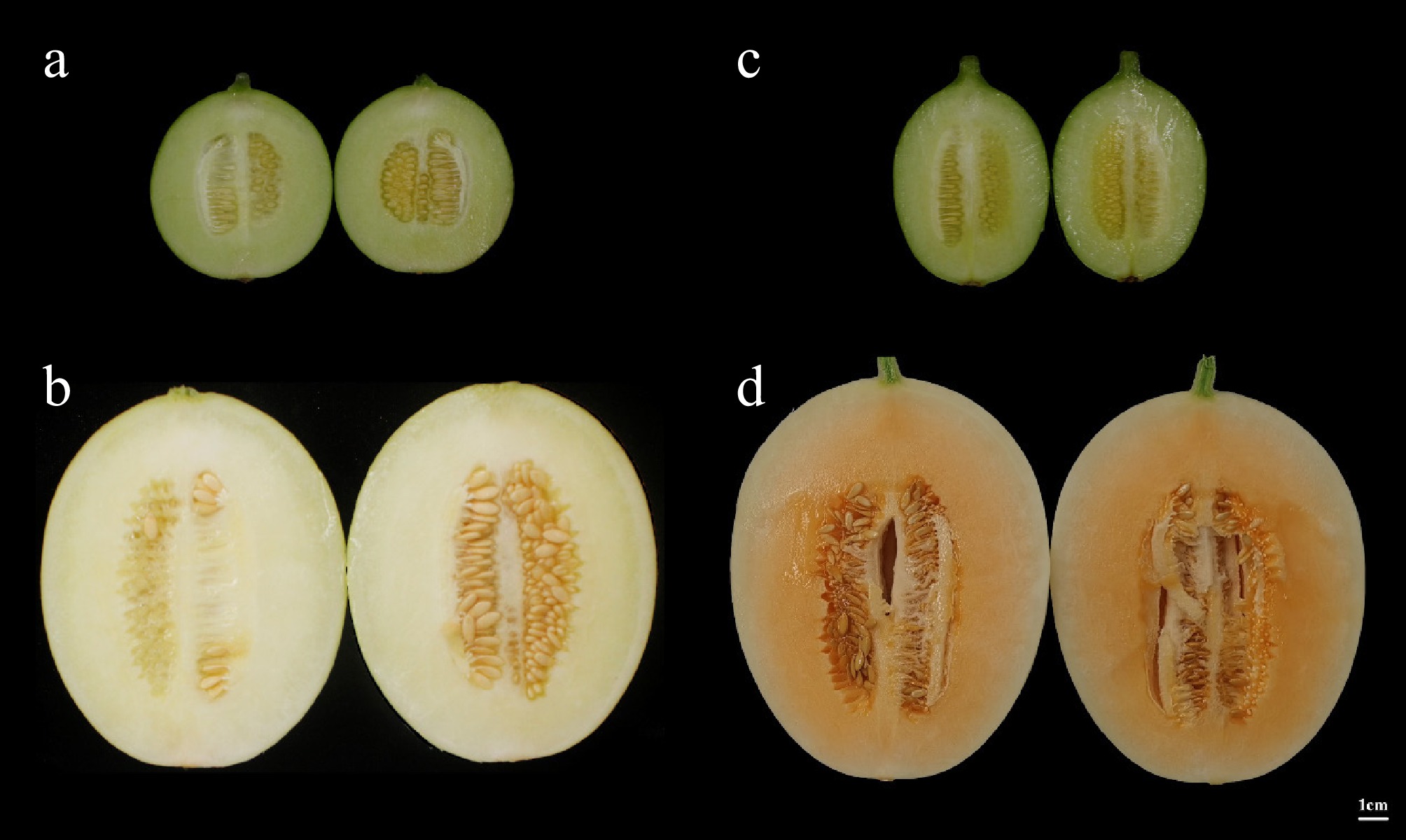
Figure 1.
Flesh color of 'TianYu' and 'TianJu' at different stages of fertility. (a) Represents the color of 'TianYu' flesh at 15 d post-pollination; (b) depicts the color of 'TianYu' flesh at 30 d post-pollination; (c) illustrates the color of 'TianJu' flesh at 15 d post-pollination; (d) portrays the color of 'TianJu' flesh at 30 d post-pollination.
Carotenoid content of fruit flesh at different fertility stages of 'TianYu' and 'TianJu'
-
To investigate the relationship between carotenoid content and flesh color in melons, we determined the contents of 66 carotenoids in 'TianYu' and 'TianJu' melons at two stages: before color change and during color change. A total of 19 carotenoids were detected in the pulp samples of 'TianYu' and 'TianJu' at different fertility stages: β-carotene, (E/Z)-phytoene, lycopene, lutein dimyristate, zeaxanthin, violaxanthin, neoxanthin, lutein, echinenone, lutein dilaurate, violaxanthin-myristate-caprate, β-cryptoxanthin, 8'-apo-beta-carotenal, lutein myristate, canthaxanthin, violaxanthin dilaurate, violaxanthin myristate, and violaxanthin-myristate-laurate (Fig. 2). Among the 19 carotenoids detected, β-carotene and lutein exhibited substantial differences in content between the two stages. Specifically, both β-carotene and lutein content significantly decreased during the process of 'TianYu' flesh color change. In contrast, during the process of 'TianJu' flesh color change, lutein content significantly decreased, while β-carotene content significantly increased to 10.35 ± 0.47 μg/g (Fig. 2). These findings suggest that β-carotene and lutein play a crucial role in the color change of melon flesh, with the accumulation of β-carotene potentially being the key reason responsible for the orange flesh color. Furthermore, we hypothesize that the significant accumulation of (E/Z)-phytoene, lutein dilaurate, violaxanthin-myristate-caprate, and β-cryptoxanthin also contributes to the transformation of 'TianJu' flesh into an orange color (Fig. 2).
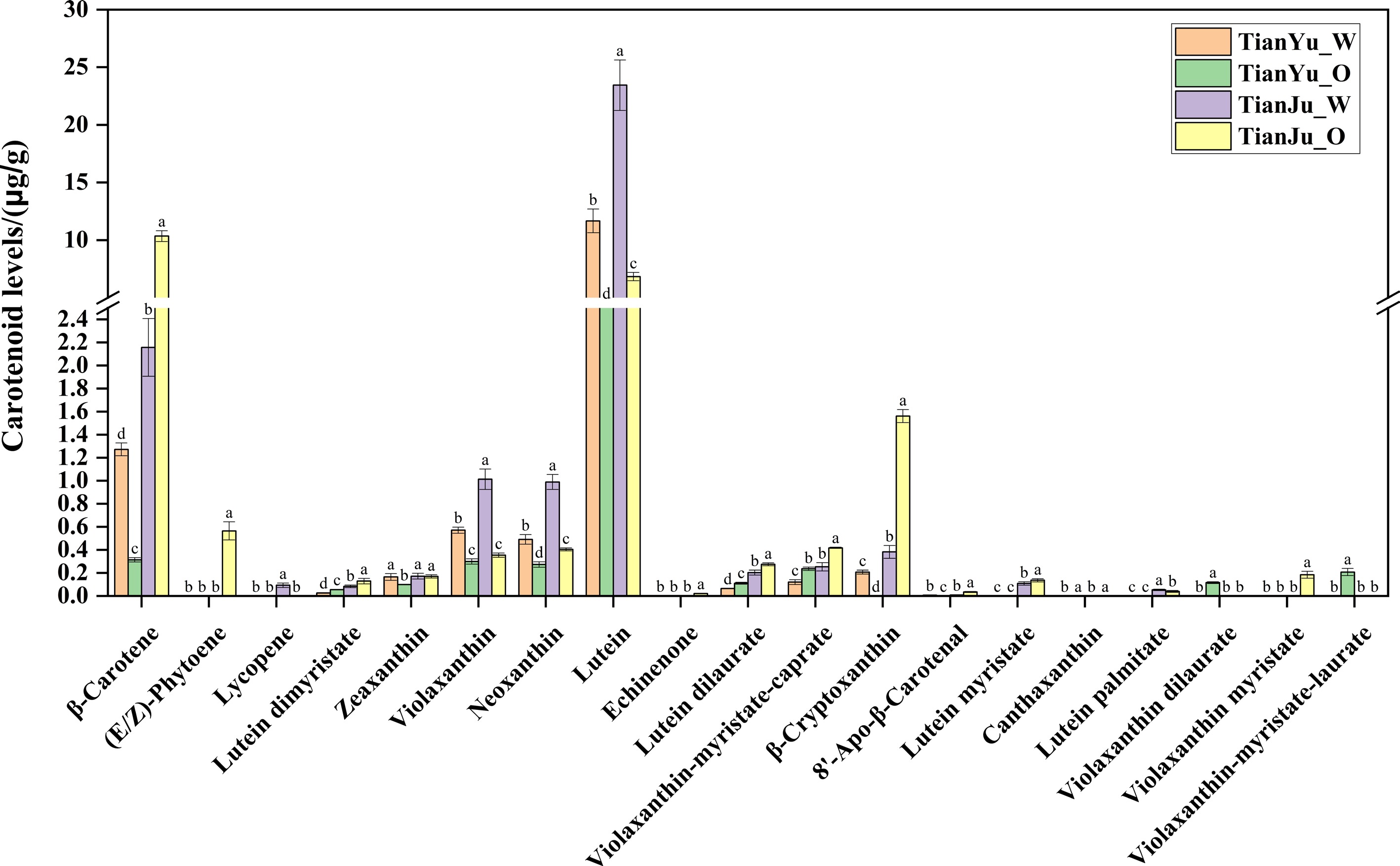
Figure 2.
Contents of various types of carotenoids in the pulp of 'TianYu' and 'TianJu' fruits at different fertility stages. Columns of different colors represent flesh samples from different fertility stages of 'TianYu' or 'TianJu'. The data are shown as the mean ± SD. Different letters indicate statistical significance at p < 0.05 (Duncan's test).
Analysis of raw transcriptome data from melon flesh samples
-
Following sequencing on the Illumina NovaSeq 6000 platform, the transcriptome data were obtained from flesh samples of 'TianYu' and 'TianJu' cultivars at various developmental stages. The process yielded a total of 41.74−54.39 million raw reads. Following the quality control process, a total of 40.92−53.75 million clean reads remained (Supplementary Table S2). Furthermore, the sequencing exhibited a low mismatch rate, with the proportion of Q20 and Q30 bases exceeding 97.48% and 92.79%, respectively. These metrics indicate that the sequencing data generated in this study are of high quality and render it suitable for further bioinformatic analysis (Supplementary Table S2).
Gene expression values from RNA-seq are generally influenced by gene length and sequencing depth. Therefore, the data were expressed as FPKM (Fragments per Kilobase Million) to correct for these factors. Box plots illustrate the distribution of gene expression levels (FPKM) across different samples in this study. The overall distribution of FPKM was relatively uniform, and exhibited satisfactory homogeneity among different samples, indicating the stable quality of the transcriptome data (Supplementary Fig. S1). Furthermore, correlation tests were performed on the biological replicates. The Pearson correlation coefficient (R2) between samples exceeded 0.9, signifying robust biological reproducibility (Supplementary Fig. S2). Principal component analysis similarly confirmed the satisfactory reproducibility of the samples (Supplementary Fig. S3).
Identification and analysis of differentially expressed genes
-
In this study, fruit samples from different fertility stages of 'TianYu' and 'TianJu' were divided into four comparison groups. The genes identified by RNA-seq were compared for expression levels using fold change values, and the significance of differences was determined using t-test and p-value. The DEG screening criteria for this study were |log2(FoldChange)| ≥ 1 and p ≤ 0.05. The highest number of DEGs were identified from the TianJu_O vs TianJu_W comparison group, totalling 5,915 genes, of which 3,127 genes were up-expressed and 2,788 genes were down-expressed (Fig. 3a; Supplementary Table S3). The TianJu_W vs TianYu_W, TianYu_O vs TianYu_W, and TianJu_O vs TianYu_O comparison groups respectively showcased 2,104 (1,145 up-regulated, 959 down-regulated), 4,855 (2,567 up-regulated, 2,288 down-regulated), and 2,346 (1,455 up-regulated, 891 down-regulated) DEGs (Fig. 3a; Supplementary Tables S4−S6).
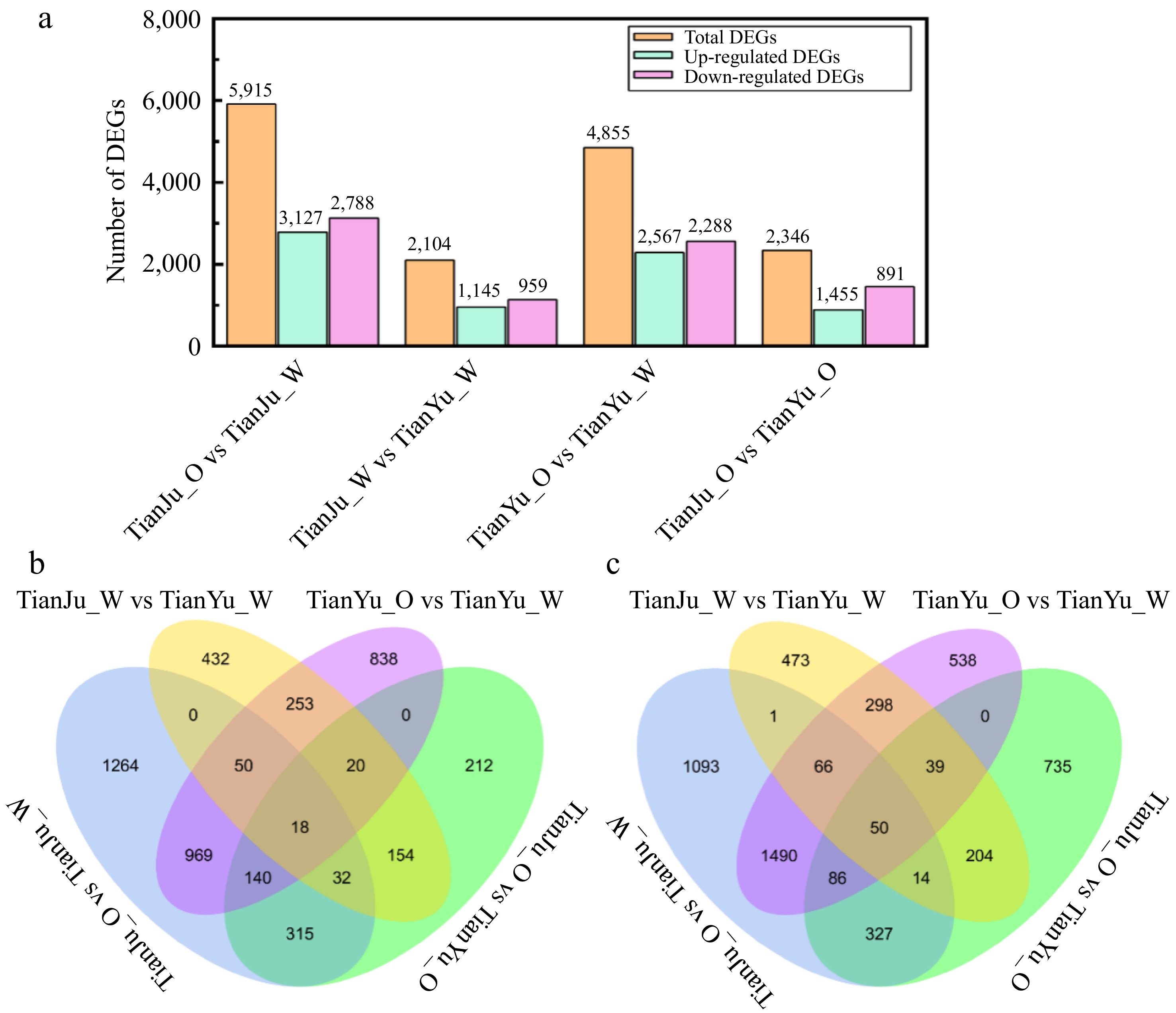
Figure 3.
Statistics of DEGs in between various comparision groups. (a) Depicts the number of DEGs across the four comparison groups; (b) presents a Venn diagram illustrating the common and unique up-regulated DEGs among different pairwise comparison groups; (c) displays a Venn diagram representing the common and unique down-regulated DEGs across various pairwise comparison groups.
Statistical analysis of DEGs revealed 969 upregulated and 1,490 downregulated genes shared between the color-changing processes of 'TianYu' and 'TianJu' (Fig. 3b & c; Supplementary Table S7−S14). We speculate that these 2,459 genes play a critical role in the color transformation of melon flesh. Additionally, 212 uniquely upregulated and 735 uniquely downregulated genes were identified in the 'TianJu_O vs TianYu_O' comparison (Fig. 3b & c; Supplementary Tables S7−S14). We propose that these genes are key regulators of orange flesh formation in melon.
Transcriptome data reliability validation
-
To confirm the reliability of transcriptome data, we selectively performed qRT-PCR for several DEGs related to carotenoid biosynthesis from the four comparison groups. A total of 16 up-regulated and 12 down-regulated genes were chosen, and the expression trends of all these DEGs were consistent with those revealed by the transcriptome data (Fig. 4). These findings revealed that the transcriptome results obtained in this study are reliable and can be utilized for subsequent analyses.
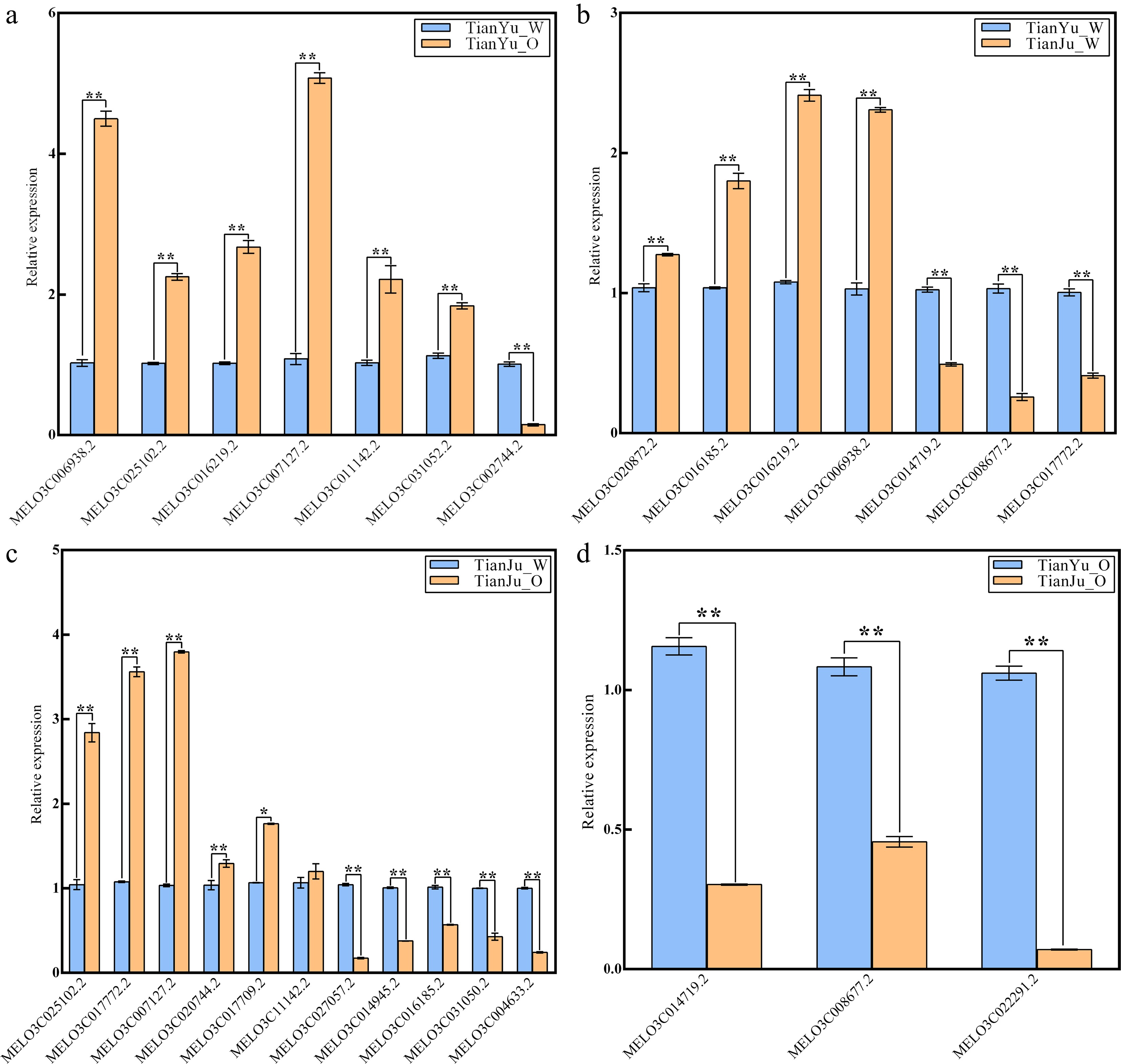
Figure 4.
qRT-PCR Validation of DEGs in different comparison groups. (a) Represents the quantitative expression results of DEGs in the comparison group of 'TianYu_O vs TianYu_W'; (b) illustrates the quantitative expression results of DEGs in the comparison group of 'TianJu_O vs TianYu_W'; (c) demonstrates the quantitative expression results of DEGs in the comparison group of 'TianJu_O vs TianJu_W'; (d) depicts the quantitative expression results of DEGs in the comparison group of 'TianJu_O vs TianYu_O'. Relative expression (Y-axis) was calculated using the 2−ΔΔCᴛ method, with DEGs shown on the X-axis. Bars represent the mean ± SD of three replicates. * and ** denote significant differences in expression levels at p < 0.05 and p < 0.01, respectively (Student's t-test).
Functional enrichment analysis of differentially expressed genes
-
To delineate the molecular events governing carotenoid-mediated pigmentation in Cucumis melo, we performed integrative GO and KEGG analyses on the identified DEGs. The DEGs were analyzed separately and classified into three GO categories molecular function (MF), biological process (BP), and cellular component (CC). Analysis of the GO-annotated TOP30 entries revealed that most of the DEGs were annotated to MF entries (Fig. 5; Supplementary Tables S15−S18). In the TianJu_O vs TianJu_W comparison, the top three MF entries were GO:0048037 (cofactor binding), GO:0140110 (transcription regulator activity), and GO:0003700 (DNA binding transcription factor activity), containing 166 (94 up-regulated and 72 down-regulated), 132 (48 up-regulated and 84 down-regulated), and 130 (48 up-regulated and 82 down-regulated) DEGs, respectively (Fig. 5a). Similarly, in the TianJu_O vs TianYu_O comparison, the top three MF entries were identical to those in TianJu_O vs TianJu_W, containing 83 (34 up-regulated, 49 down-regulated), 73 (14 up-regulated, 59 down-regulated), and 71 (14 up-regulated, 57 down-regulated) DEGs, respectively (Fig. 5d). Comparable results were observed in the TianYu_O vs TianYu_W comparison, with 112 DEGs under both its GO:0003700 and GO:0140110 entries (Fig. 5c). However, the GO:0048037 (cofactor binding) entry was absent in both the TianYu_O vs TianYu_W and TianJu_W vs TianYu_W comparison portfolios (Fig. 5b & c). This analysis revealed that a large number of DEGs were enriched under the functional entries of transcriptional regulation during the melon flesh color transition process, suggesting that key transcription factors play a crucial role in regulating this shift. Notably, the significant enrichment of the GO:0048037 entry exclusively in the comparison group of orange-colored flesh 'TianJu' after discoloration indicates that genes under this entry are potentially key regulators of orange flesh formation.
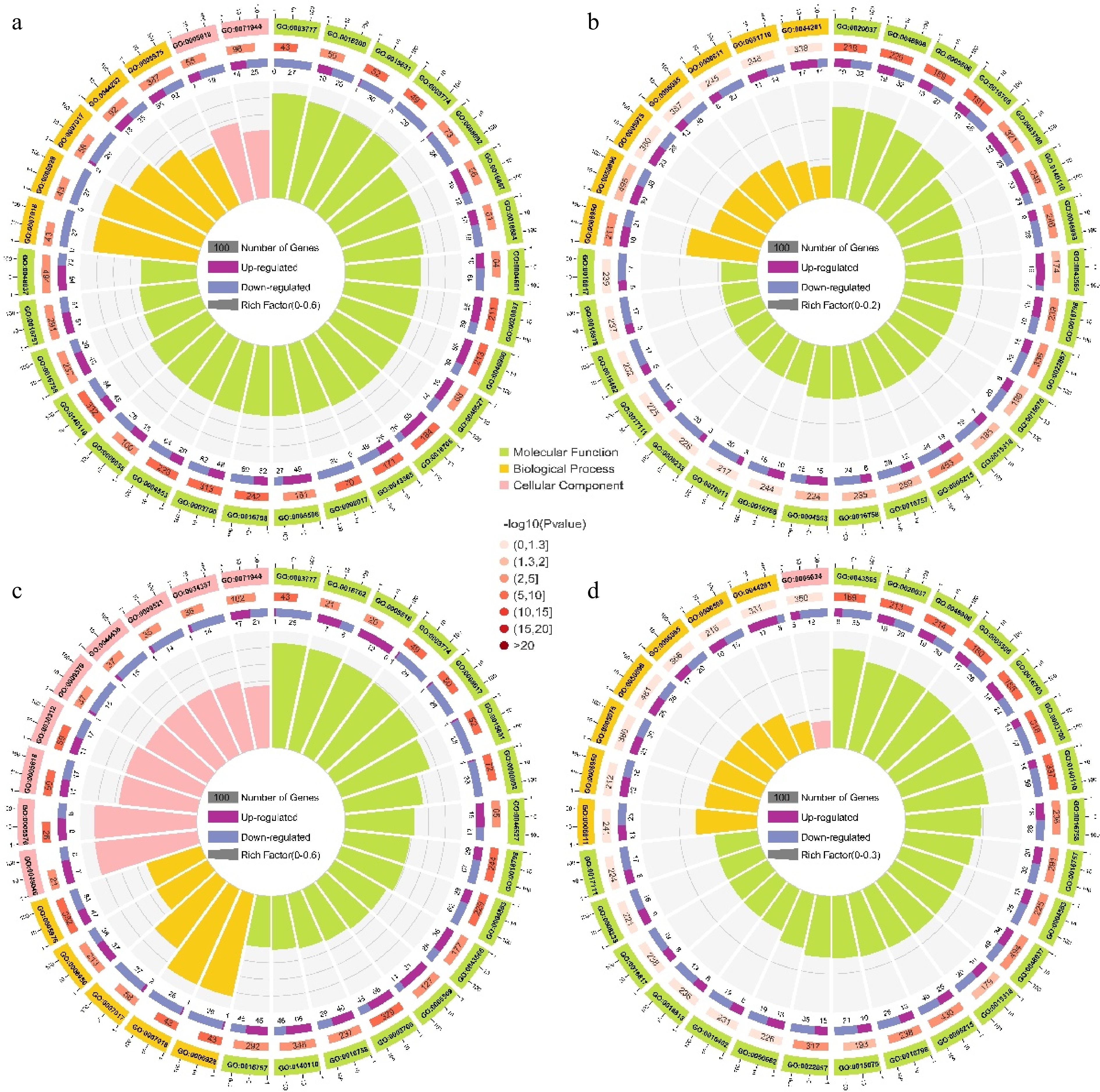
Figure 5.
GO enrichment analysis of DEGs in the four comparison groups. (a) Top 30 GO terms for the TianJu_O vs TianJu_W comparison; (b) top 30 GO terms for the TianJu_W vs TianYu_W comparison; (c) top 30 GO terms for the TianYu_O vs TianYu_W comparison; (d) top 30 GO terms for the TianJu_O vs TianYu_O comparison. The outermost circle represents the scale for the number of genes, with each division denoting a specific enriched GO classification, differentiated by color. The second circle from the outside displays the number of genes in that GO term within the background gene set, along with the corresponding p-value. The third circle depicts the proportion of up- and down-regulated genes via a bar chart. The innermost circle presents the RichFactor value for each GO term, calculated as the ratio of the number of foreground genes to background genes in that term. Each cell of the background auxiliary line represents an increment of 0.1.
To elucidate the biological pathways associated with flesh color shift in melon, we performed a KEGG enrichment analysis of DEGs from four comparison groups, focusing on the top 20 entries (Fig. 6; Supplementary Table S19−S22). The analysis revealed that a substantial number of DEGs were enriched in the cmo04075 (plant hormone signal transduction), cmo04016 (MAPK signaling pathway - plant), and cmo04626 (plant-pathogen interaction) biological pathways, with a significant overlap of DEGs across the four comparison groups (Fig. 6; Supplementary Table S19−S22). Notably, the cmo04141 (protein processing in endoplasmic reticulum), cmo00071 (fatty acid degradation), and cmo00950 (isoquinoline alkaloid biosynthesis) entries were significantly enriched exclusively in the orange flesh of 'TianJu', suggesting a close association between these biological pathways, their enriched DEGs, and the orange flesh color change process of 'TianJu' (Fig. 6a; Supplementary Table S19). Furthermore, cmo00910 (nitrogen metabolism) and cmo00590 (arachidonic acid metabolism) were significantly enriched solely in the TianJu_O vs TianJu_W and TianYu_O vs TianYu_W comparison groups, leading to the hypothesis that these two biological pathways play a crucial role in regulating melon flesh color change (Fig. 6a & c). Additionally, the cmo00270 (cysteine and methionine metabolism) and cmo00920 (sulfur metabolism) pathways were significantly enriched in the TianJu_O vs TianJu_W and TianJu_O vs TianYu_O comparison groups, suggesting their potential involvement in the development of orange flesh (Fig. 6a & d).
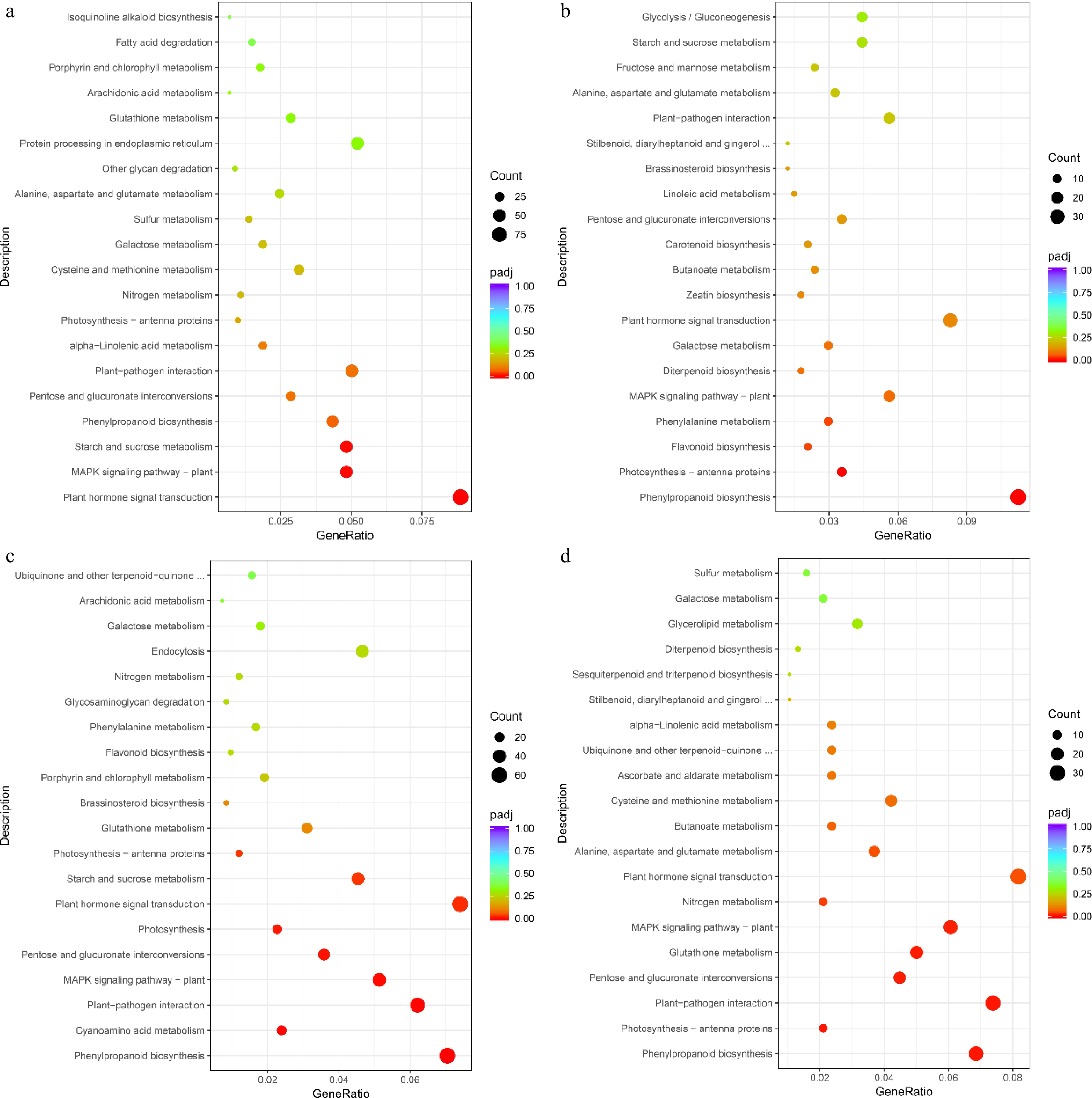
Figure 6.
KEGG enrichment analysis of DEGs in different comparison groups. (a) Top 20 information under KEGG item about TianJu_O vs TianJu_W comparison group; (b) top 20 information under KEGG item about TianJu_W vs TianYu_W comparison group; (c) top 20 information under KEGG item about TianYu_O vs TianYu_W comparison group; (d) top 20 information under KEGG item about TianJu_O vs TianYu_O comparison group. Bubble size represents the number of DEGs contained under that entry.
Analysis of differentially expressed transcription factors
-
Since functional enrichment analyses indicated that transcriptional regulation and transcription factor regulation play important roles in the process of melon flesh color change. Consequently, we summarized and analyzed the differentially expressed transcription factors (TFs) across different comparison groups. We identified and analyzed a total of seven major transcription factor families from four comparison groups (Fig. 7; Supplementary Table S23), among which MYB, WRKY, ERF, and bHLH were the predominant classes in terms of the number of differentially expressed TFs (Fig. 7). The numbers of these four classes of TFs were greater in the TianYu_O vs TianYu_W comparison group than in the other comparison groups. During melon flesh transcoloration (TianJu_O vs TianJu_W and TianYu_O vs TianYu_W), significantly more bZIP, MYB, ERF, and NAC classes of TFs were up-regulated compared with the other two comparison groups (Supplementary Table S23). We hypothesize that these TFs may play a regulatory role in the color change of melon flesh.
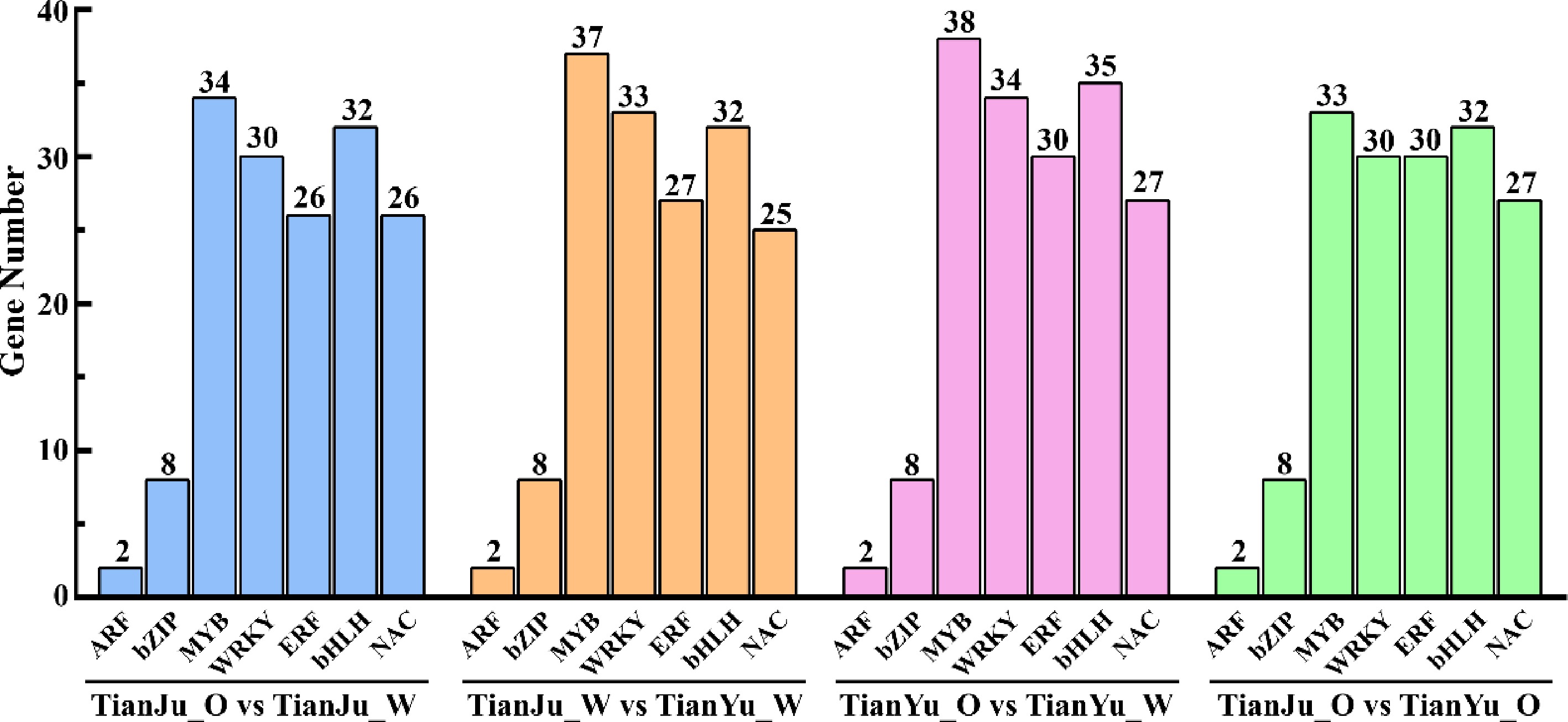
Figure 7.
Differentially expressed TFs across various comparative groups. The four groups of columns, distinguished by color, represent the four comparison groups. The vertical axis denotes the number of differentially expressed TFs, while the horizontal axis enumerates the specific types of TFs.
Analysis of differentially expressed genes involved in carotenoid synthesis and metabolism pathways
-
Analysis of DEGs involved in the carotenoid synthesis and metabolism pathways was conducted to investigate the relationship between melon flesh color change and the synthesis and metabolism of carotenoids. This was based on the observation of notable changes in flesh color and carotenoid content during the ripening process of melon fruits, suggesting that the type and content of carotenoids are key factors influencing the transformation of melon flesh color and the color differences among various fruits.
Analysis of DEGs across the four comparison groups identified 42 genes associated with carotenoid biosynthesis and metabolism. (Fig. 8; Supplementary Tables S3−S6). The plastid pathway serves as the primary pathway for carotenoid biosynthesis in melon, with pyruvate acting as a key substrate for de novo carotenoid synthesis. The findings demonstrate that genes encoding key enzymes (PK, PEPCK, MDH, and NADP-ME) of the pyruvate synthesis pathway exhibited up-regulated expression during the color change of melon pulp (Fig. 8). Concurrently, genes coding for key enzymes (such as PDH) promoting pyruvate metabolism were down-regulated during the flesh color change process (Fig. 8). These results suggest that melon pulp accumulates substantial amounts of pyruvate during the color transfer process, providing sufficient substrate for carotenoid biosynthesis. GGPP serves as the immediate precursor substance for carotenoid synthesis, with two molecules of GGPP condensing to form colorless phytoene through the action of PSY, the first key rate-limiting enzyme in the carotenoid synthesis pathway. The gene encoding PSY, MELO3C25102.2, exhibited up-regulated expression during melon pulp color change (TianJu_O vs TianJu_W and TianYu_O vs TianYu_W), potentially accelerating carotenoid biosynthesis (Fig. 8). It is noteworthy that the genes encoding PDS (MELO3C017772.2) and LCYB (MELO3C020744.2) were up-regulated exclusively in the 'TianJu_O vs TianJu_W' comparison group (Fig. 8), suggesting that the significantly higher β-carotene content in the flesh of ripe 'TianJu' compared to other samples may be attributed to the upregulation of these genes (Fig. 2). Furthermore, in the carotenoid metabolic pathway (especially the β-carotene metabolic pathway), several genes encoding key enzymes such as MELO3C022291.2 (CmCCD7), MELO3C011142.2 (CmCCD8), MELO3C006938.2 (CrtR-b coding gene), and MELO3C007127.2 (CmNCED5a) showed up-regulated expression during flesh color change in 'TianYu' (Fig. 8). The upregulation of these genes would promote the metabolic process of β-carotene, which we hypothesized to be an important reason for the reduction of β-carotene in 'TianYu' pulp (Fig. 2).
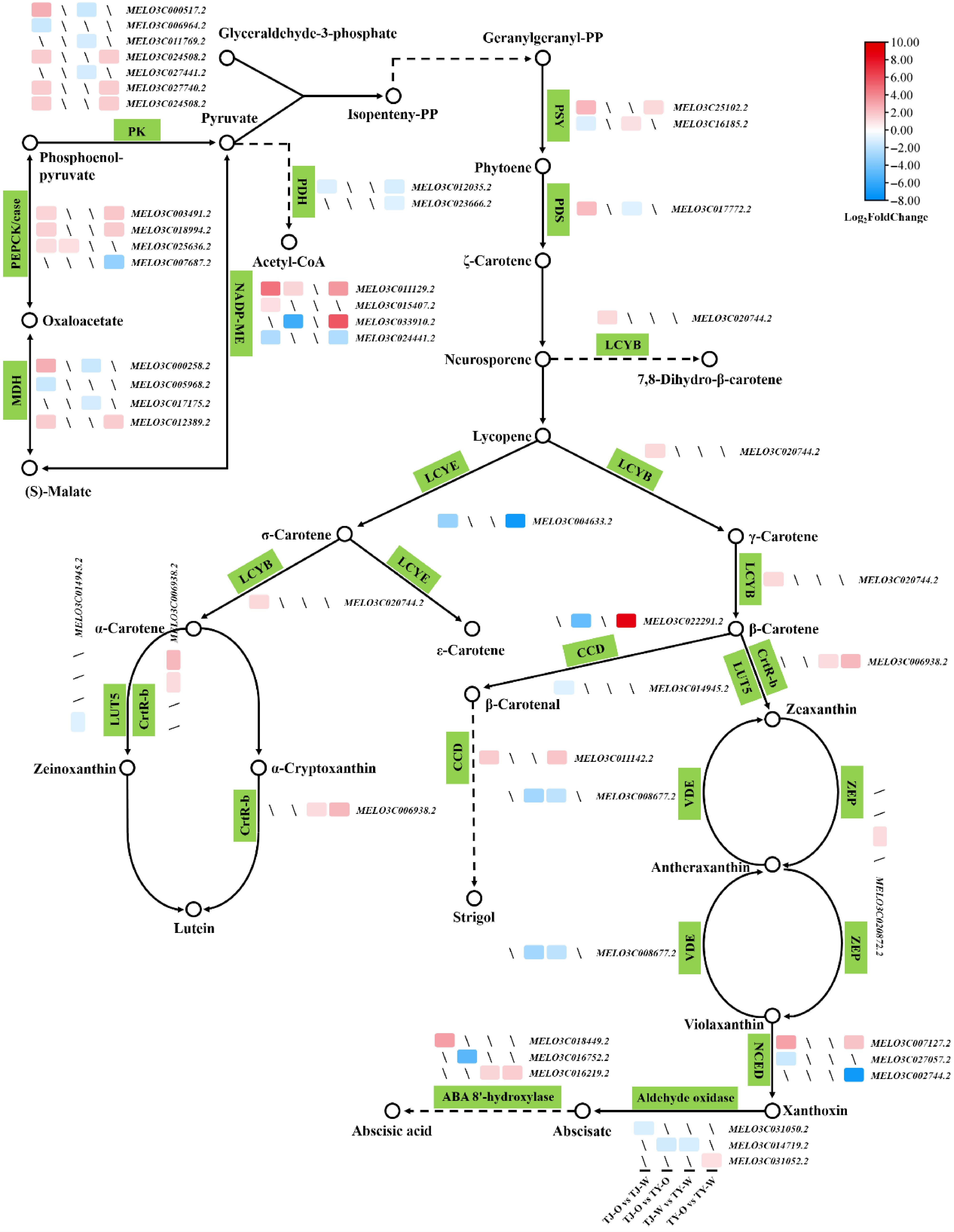
Figure 8.
Expression patterns of DEGs involved in carotenoid biosynthesis and metabolism pathways across various comparison groups. Hollow dots signify molecules or products, while green squares denote catalytic enzymes at each stage. Sharp lines indicate synthesis or promotion, and dashed lines represent the presence of other processes. Differentially colored squares represent DEGs, with mRNA color bars illustrating the Log2(FoldChange) values of the DEGs.
During the color transformation of melon flesh, especially in 'TianJu', a significant accumulation of pyruvate was observed. This accumulation provides an ample substrate for carotenoid biosynthesis. Additionally, the upregulated expression of carotenoid synthesis genes encoding PSY, PDS, and LCYB led to increased accumulation of phytoene, ζ-carotene, γ-carotene, and β-carotene in 'TianJu' pulp (Figs 2 & 8). In contrast, 'TianYu' pulp is more likely to accumulate zeinoxanthin, lutein, and other carotenoids. Additionally, under the influence of PSY, CCD, and NCED, melon pulp will accumulate higher levels of phytoene, strigol, and xanthoxin after ripening (Fig. 8). These compounds are potentially responsible for variations in flesh color and color differences between melon varieties. Furthermore, genes encoding enzymes involved in carotenoid biosynthesis and metabolism were identified as key regulators of melon pulp color change.
-
Flesh color is an essential commodity trait of melon, and its regulatory mechanism has been a focal point of research. In this study, carotenoid components and contents of white flesh melon 'TianYu' and orange flesh melon 'TianJu' were analyzed at various developmental stages. The results indicated that although the carotenoid components were essentially identical in the two melon varieties across different developmental stages, their contents varied. Consequently, carotenoids were identified as the primary factor contributing to the variation in flesh color and the differences in flesh color among diverse melon varieties. Subsequently, we conducted transcriptome analysis of the two melon varieties at distinct developmental stages to investigate the expression of genes associated with carotenoid biosynthesis and metabolism, aiming to elucidate the molecular mechanism underlying carotenoid accumulation and flesh color alteration.
The carotenoid composition and content vary among different plant varieties, with the red, yellow, and orange hues of plants directly correlating to their carotenoid levels[18]. In this study, a total of 19 carotenoids were detected in the flesh samples of 'TianYu' and 'TianJu', with significant differences observed in the contents of β-carotene and lutein (Fig. 2). The β-carotene content in 'TianJu' flesh after color conversion was markedly higher than the content of other carotenoids and substantially greater than that in other samples (Fig. 2). Consequently, it is hypothesized that β-carotene is the primary compound responsible for the orange flesh color, aligning with the findings of previous studies[1,2,50]. Additionally, this study suggests that the orange color of 'TianJu' pulp may also be influenced by (E/Z)-phytoene, lutein dilaurate, violaxanthin-myristate-caprate, and β-cryptoxanthin. In summary, the accumulation of carotenoids is the principal factor underlying the differences in melon flesh color. Therefore, the synthesis and metabolism pathways of carotenoids are critical aspects in the investigation of melon flesh color variation.
In melons, the principal pathway for carotenoid biosynthesis is the plastid pathway, which utilizes pyruvate and glyceraldehyde 3-phosphate as precursors[1,12,13]. Following the color conversion in 'TianJu' samples, the expression of critical rate-limiting enzyme-encoding genes in the pyruvate synthesis pathway was significantly upregulated, including that of MELO3C000517.2 (PK-encoding gene), MELO3C000258.2 (MDH-encoding gene), and MELO3C011129.2 (NADP-ME-encoding gene) (Fig. 8). Consequently, we hypothesize that 'Tianju' possesses a greater abundance of carotenoid synthetic substrates compared to 'TianYu'. PDS is a pivotal rate-limiting enzyme in the carotenoid synthesis pathway. Previous research has demonstrated the direct influence of PDS on β-carotene content[51]. In Arabidopsis, PDS mutations result in a loss of desaturation activity, ultimately impeding the synthesis of β-carotene and other carotenoids[51]. During citrus fruit development, the expression level of the PDS-encoding gene also increases significantly in parallel with the elevation of carotenoid content, suggesting its importance in regulating carotenoid production[52]. In the present study, the expression of the PDS-encoding gene (MELO3C017772.2) was significantly upregulated exclusively in the mature flesh samples of 'TianJu', with expression levels substantially higher than those observed in 'TianJu' samples before color transformation (Figs 4c & 8). Considering the carotenoid content determination results (Fig. 2), we propose that MELO3C017772.2 is a key gene regulating β-carotene synthesis and the development of orange-colored flesh in melons.
Undoubtedly, the elevated expression of β-carotene biosynthesis gene MELO3C020744.2 (the gene encoding LCYB) may also exert a significant regulatory influence on the accumulation of β-carotene and the development of orange flesh (Fig. 8). The LCYB-encoding gene was initially cloned in Arabidopsis thaliana, with lycopene being identified as its catalytic substrate[53]. LCYB exerts the most substantial impact on the partitioning of carotenoid synthesis between the two branches following lycopene synthesis, and its expression leads to an increase in the β-carotene content of maize endosperm[54]. Loss-of-function mutations in the coding region of papaya lycopene β-cyclase resulted in the accumulation of lycopene in the pulp, leading to a reddish coloration[55]. Overexpression of LCYB in sweetpotato tubers altered the tuber color from whitish-yellow to orange-yellow and caused an exponential increase in β-carotene content compared to the control[56]. In this study, the LCYB-encoding gene was upregulated only in the 'TianJu_O vs TianJu_W' comparison group, resulting in a substantial accumulation of β-carotene during the color change process of 'TianJu' flesh. Numerous studies have demonstrated that the accumulation of β-carotene causes pulp or petals to take on an orange color[55−57], suggesting that the orange pulp of 'Tianju' is due to high β-carotene levels. Previous studies have established that CmOr acts as a pivotal regulator of orange pulp development in melon[33]. This regulatory function is mediated through two synergistic mechanisms: enhancing the biosynthesis of the carotenoid precursor IPP, and upregulating the expression of rate-limiting enzymatic genes, such as PSY and LCYB, within the carotenoid biosynthetic pathway[33,58]. These coordinated molecular interactions substantially elevate β-carotene accumulation, thereby driving the development of orange pigmentation in melon pulp[33,58]. The present study identified PSY and LCYB as potential key factors in the orange pulp regulatory pathway, which is consistent with previous findings. In this study, MELO3C017772.2 (PDS) and MELO3C020744.2 (LCYB) were further identified as the key genes for orange pulp formation. Combined with the results of previous studies, we hypothesized that CmOr may function by directly or indirectly regulating the expression of PDS and LCYB. With the gradual improvement of the melon genetic transformation system, our follow-up investigations will implement CRISPR/Cas9-mediated knockout of MELO3C017772.2 and MELO3C020744.2 in melon materials with orange-colored flesh to verify the roles of these two genes in the formation of orange-colored flesh in melon. Furthermore, genes encoding enzymes that promote carotenoid metabolism were upregulated during the color change process of 'TianYu' flesh, such as MELO3C022291.2 (CCD-encoding gene), MELO3C006938.2 (CrtR-b-encoding gene), and MELO3C007127.2 (NCED-encoding gene) (Fig. 8). Consequently, fewer carotenoids accumulated in the flesh of 'TianYu', which may be the primary reason for its white flesh color. Building on these findings, our subsequent investigation will employ overexpression of carotenoid metabolic genes in orange-fleshed melon to elucidate the molecular basis underlying the formation of white flesh phenotype in melon. Subsequently, the knockout and overexpression variants will be analyzed via functional genomics to identify critical regulators of melon flesh regulation, ultimately establishing a comprehensive regulatory network for flesh coloration in Cucumis melo.
Numerous studies have demonstrated the crucial regulatory roles played by various TFs in carotenoid biosynthesis and metabolism[37,59]. Researchers have discovered that specific members of the MYB and WRKY transcription factor families in tomato possess the ability to influence carotenoid synthesis by regulating the activity of PSY[37,59,60]. The WRKY transcription factor CmWRKY49 has been experimentally validated to directly bind the CmPSY1 promoter and transcriptionally activate its expression, thereby orchestrating carotenoid biosynthesis in melon[2]. Furthermore, the involvement of certain NAC transcription factors in the carotenoid metabolic pathway has been established[61−63]. In the present study, we identified a total of seven major classes of transcription factor families (Fig. 7). Considering that the accumulation of carotenoids, particularly β-carotene, may be the primary factor contributing to the orange coloration of 'TianJu', we focused on the relevant transcription factors involved in the transcoloring process of 'TianJu'. Our analysis identified a total of 158 differentially expressed transcription factors in the 'TianJu_O vs TianJu_W' comparison group, among which six (two bZIP53, MYB108, MYB44, WRKY14, and bHLH117) exhibited upregulation exclusively in this comparison group (Supplementary Table S23). Based on these findings, we propose that these six transcription factors play pivotal regulatory roles in carotenoid biosynthesis, especially in promoting β-carotene synthesis.
-
This study elucidates the critical role of carotenoids in the formation of melon flesh color through a comprehensive analysis of the changes in flesh color and carotenoid content during the fruit development of two melons with distinct flesh colors, 'TianYu' and 'TianJu'. The findings reveal that the flesh of 'TianJu' transitioned from light green to orange during the ripening process, which was accompanied by a substantial increase in β-carotene content. In contrast, the flesh of 'TianYu' remained white, and its β-carotene and lutein content decreased. These observations suggest that the change in melon pulp color is attributable to carotenoid accumulation, with β-carotene being the primary contributor to the orange coloration of melon flesh. Furthermore, through RNA-seq analysis, DEGs associated with changes in flesh color were identified, which were found to play crucial roles in carotenoid biosynthesis and metabolic pathways. Notably, the upregulated expression of genes encoding key enzymes in the pyruvate synthesis pathway provides sufficient substrates for carotenoid synthesis. Simultaneously, the upregulation of genes encoding key enzymes, such as PSY, PDS, and LCYB, promoted carotenoid synthesis and accumulation. Among these, MELO3C017772.2 (PDS-encoding gene) and MELO3C020744.2 (LCYB-encoding gene) were identified as the key genes regulating β-carotene synthesis and the formation of orange flesh in melon. Additionally, this study revealed the significant roles of transcriptional regulation and transcription factors in melon flesh color changes through functional enrichment analysis of DEGs. Several transcription factor families, including MYB, WRKY, ERF, and bHLH, exhibited significant differential expression during flesh discoloration, indicating their potential influence on melon flesh color by regulating key genes in the carotenoid synthesis pathway. In conclusion, this study not only provides initial insights into the molecular mechanism underlying melon flesh color change but also highlights the regulatory roles of key genes and transcription factors in the carotenoid synthesis pathway. These findings offer a crucial theoretical foundation for melon quality improvement and breeding, which may also potentially facilitate the development of future genetic engineering approaches to regulate melon flesh color.
This work was supported by the National Natural Science Foundation of China (NSFC: 32402579), Xi'an Science and Technology Program Project (23NYGG0029), the Fundamental Research Funds for the Central Universities (2452021099), and the Xinjiang Uygur Autonomous Region Specialized Program on Innovation Environment Construction (2024D01A111).
-
The authors confirm contribution to the paper as follows: conceptualization, data analysis, experimentation, and manuscript preparation: Wang Z, Cheng D; supervision: Zhang Y; plant growth management: Yao S, Duan J, Hu J; data analysis and trait identification: Zhang S, Wang Z, Wang J, Yang Y; collection and identification of germplasm: Wang Y, Yang X; project implementation and manuscript development: Wei C, Zhang X; funding acquisition, investigation, methodology, and review: Zhang Y, Wang Z, Zhang X, Wei C, Li C. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its supplementary information files.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Zhongyuan Wang, Denghu Cheng
- Supplementary Table S1 List of qRT-PCR primers.
- Supplementary Table S2 RNA-seq raw data summary.
- Supplementary Table S3 DEGs information of TianJu_O vs TianJu_W.
- Supplementary Table S4 DEGs information of TianJu_W vs TianYu_W.
- Supplementary Table S5 DEGs information of TianYu_O vs TianYu_W.
- Supplementary Table S6 DEGs information of TianJu_O vs TianYu_O.
- Supplementary Table S7 TianJu_O vs TianJu_W_up.
- Supplementary Table S8 TianJu_W vs TianYu_W_up.
- Supplementary Table S9 TianYu_O vs TianYu_W_up.
- Supplementary Table S10 TianJu_O vs TianYu_O_up.
- Supplementary Table S11 TianJu_O vs TianJu_W_down.
- Supplementary Table S12 TianJu_W vs TianYu_W_down.
- Supplementary Table S13 TianYu_O vs TianYu_down.
- Supplementary Table S14 TianJu_O vs TianYu_O_down.
- Supplementary Table S15 The information of GO enrichment Top 30 entry for TianJu_O vs TianJu_W comparison combination.
- Supplementary Table S16 The information of GO enrichment Top 30 entry for TianJu_W vs TianYu_W comparison combination.
- Supplementary Table S17 The information of GO enrichment Top 30 entry for TianYu_O vs TianYu_W comparison combination.
- Supplementary Table S18 The information of GO enrichment Top 30 entry for TianJu_O vs TianYu_O comparison combination.
- Supplementary Table S19 The information of KEGG enrichment Top 20 entry for TianJu_O vs TianJu_W comparison combination.
- Supplementary Table S20 The information of KEGG enrichment Top 20 entry for TianJu_W vs TianYu_W comparison combination.
- Supplementary Table S21 The information of KEGG enrichment Top 20 entry for TianYu_O vs TianYu_W comparison combination.
- Supplementary Table S22 The information of KEGG enrichment Top 20 entry for TianJu_O vs TianYu_O comparison combination.
- Supplementary Table S23 List of differentially expressed transcription factors.
- Supplementary Fig. S1 Box plot of sample gene expression distribution.
- Supplementary Fig. S2 Heat map of inter-sample correlation.
- Supplementary Fig. S3 Results of Principal Component Analysis.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Z, Cheng D, Yao S, Zhang S, Duan J, et al. 2025. Joint transcriptomic and metabolomic analyses reveal the mechanism of flesh transcoloration and orange flesh formation in melon. Vegetable Research 5: e024 doi: 10.48130/vegres-0025-0018 

Joint transcriptomic and metabolomic analyses reveal the mechanism of flesh transcoloration and orange flesh formation in melon
- Received: 15 January 2025
- Revised: 28 April 2025
- Accepted: 12 May 2025
- Published online: 03 July 2025
Abstract: Flesh color is a critical commodity trait of melon, directly influencing consumer preference and the fruit's market value. This study investigates the crucial role of carotenoids in determining melon flesh coloration by examining color transitions and carotenoid profiles during fruit development in 'TianYu' (white flesh) and 'TianJu' (orange flesh) melons. Research has demonstrated that the color change in melon flesh results from carotenoid accumulation, with β-carotene being the primary compound responsible for the formation of orange flesh. Transcriptome analysis revealed that 2,459 and 947 differentially expressed genes (DEGs) were involved in melon pulp color change and orange pulp formation, respectively. Among these DEGs, the study identified and analyzed those related to carotenoid synthesis and metabolism. The results indicated that genes encoding key enzymes for pyruvate synthesis, phytoene synthase (PSY), phytoene desaturase (PDS), and lycopene β-cyclase (LCYB) play significant roles in fruit pulp color change and carotenoid accumulation. Notably, the genes encoding PDS (MELO3C017772.2) and LCYB (MELO3C020744.2) were found to be potentially critical genes for β-carotene synthesis and orange flesh formation. Moreover, functional enrichment analysis of DEGs emphasized the importance of transcriptional regulation and transcription factors such as MYB, WRKY, ERF, and bHLH, whose differential expression during color change suggests their potential to regulate carotenoid pathway genes. This study not only elucidates the molecular mechanisms underlying melon flesh color variation but also reveals the potential regulatory roles of key genes and transcription factors, providing valuable insights for enhancing melon quality, informing breeding strategies, and exploring potential genetic engineering approaches for color manipulation.
-
Key words:
- Thick-skinned melon /
- Orange flesh /
- Carotenoid /
- Targeted-metabolomic /
- Transcriptome