






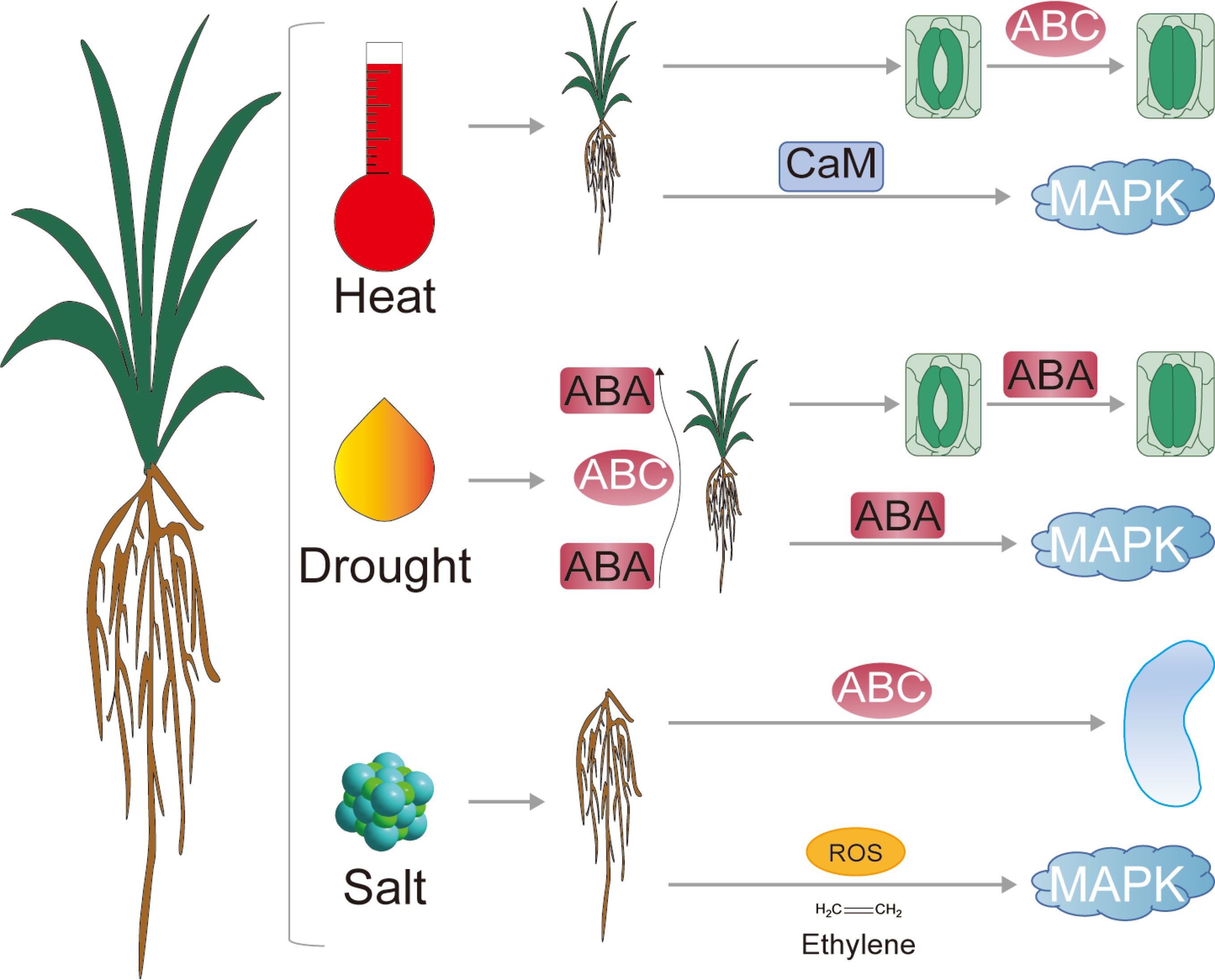



-
The extreme environmental conditions brought about by ongoing climate change exert a profound detrimental influence on the growth and maturation of plant organisms, leading to substantial reductions in agricultural productivity. According to reports from the Intergovernmental Panel on Climate Change (IPCC), between 2011 and 2020, the average surface temperature of the Earth increased by 1.1 °C, compared with 1850−1900. The global temperature rise in the 21st century so far exceeds 1.5 °C, and it will be difficult to control the temperature rise within 2 °C (IPCC, AR6 Synthesis Report: Climate Change 2023,
www.ipcc.ch/report/sixth-assessment-report-cycle/ ). The repercussions of this temperature surge are strikingly evident in agriculture, with estimations suggesting that for every 1 °C increase above 30 °C, the yield of wheat (Triticum aestivum) diminishes by 1%−1.6%[1]. Another direct threat to agriculture in the future is water resource limitations. Projections indicate that by 2050, 40% of global crop production will face drought challenges, impacting 52% of the population, and 45% of the gross domestic product (GDP)[2]. In addition to elevated temperatures and reduced rainfall, salinity is identified as a significant factor contributing to diminished crop yields. About 20% of arable land, and about half of irrigated land are affected by salinization[3]. Salt stress seriously restricts grain yield by affecting the physiological and biochemical pathways of cereal crops. Under salt stress, the yield of wheat was predicted to decrease by 60%[4], the yield of rice (Oryza sativa) by 50%[5], the dry weight of maize (Zea mays) by 51.43%, and the leaf area by 53.18%[6]. Furthermore, it is projected that the global population will reach 10 billion by 2025, exacerbating the existing challenges. To address the increasing demand, it is imperative for global food production to increase by 70% (High Level Expert Forum, FAO, October 2009;www.fao.org ). One of the primary challenges faced by plant scientists and plant breeders is the development of plant varieties that can flourish in environments with limited arable land and water resources, all while maintaining high yield levels under challenging and deteriorating environmental conditions.The utilization of conventional plant breeding techniques has led to significant improvements in crop productivity, and the emergence of enhanced resistance to a range of biotic and abiotic stressors in multiple plant species[7]. Nonetheless, the protracted timeline inherent to this approach renders it inadequate for responding promptly to the burgeoning demand for increased food production[8]. Consequently, the imperative arises for the implementation of more streamlined breeding strategies. A genetic engineering breeding strategy based on key stress-resistant genes to cultivate crops with strong abiotic stress tolerance can greatly shorten the breeding time[9]. Therefore, it is evident that acquiring a thorough understanding of the molecular mechanisms governing plant reactions to alterations in their environment is imperative. This will serve as the basis for identifying and harnessing key genes that confer abiotic stress tolerance, ultimately enabling the cultivation of resilient crops that can withstand adverse conditions[10]. Regrettably, the predominant focus of research into adaptive response mechanisms has been carried out on model plants, which in turn, has resulted in the oversight of potentially novel genes, and mechanism models related to abiotic stress. Thus, it is imperative that we broaden our research scope to non-model plants, and undertake comprehensive investigations into their abiotic response mechanisms, and identify more key genes involved in response to stress.
Pearl millet (Pennisetum glaucum, Cenchrus americanus) is a C4 plant, and it ranks as the sixth most significant cereal crop on a global scale. It also holds significance as a crucial forage grass, playing a pivotal role in ensuring food security for nearly 90 million impoverished people living in high-temperature regions of Africa and Asia (ICRISAT). Pearl millet demonstrates excellent tolerance to abiotic stress factors[11]. Previous studies have observed substantial alterations in pearl millet leaves' relative water volume (RWC), relative conductivity (REC), and malondialdehyde (MDA) content after 21 days of exposure to 40 °C stress. In contrast, maize exhibited significant changes in RWC and increased MDA content after only 4 hours of 40 °C treatment[12]. These findings highlight the greater heat resistance of pearl millet compared to maize. The extraordinary drought tolerance and salinity resistance of pearl millet make it suitable for cultivation in areas where most other crops would struggle to survive or maintain productivity levels. Pearl millet is cultivated in regions characterized by significantly low precipitation levels, typically averaging between 300 and 500 millimeters, whereas agricultural species such as maize or sorghum tend to face considerable failures in most growing seasons[13]. Under conditions of drought, the water use efficiency of pearl millet surpasses that of maize, suggesting that pearl millet demonstrates greater tolerance to drought in comparison to maize[14]. Under salt stress, the yield of pearl millet exceeds that of sorghum by more than a factor of two, indicating that pearl millet is more suitable for cultivation in saline environments[15]. Consequently, pearl millet contains a large number of stress response genes and mechanisms that diverge from those found in conventional crops, making it an ideal material for exploring how non-model crops respond to abiotic stresses[16]. Most research to date has concentrated on the phenotypic, physiological, and genetic analysis of specific genes in pearl millet under abiotic stress conditions[17,18]. Nonetheless, studies on the response mechanisms to abiotic stresses at the transcriptional level are limited. To gain a systematic and comprehensive understanding of the transcriptional dynamics of pearl millet under conditions of high temperature, drought, and salt stress, the transcriptome data of pearl millet was meticulously examined to obtain the transcriptional landscape in response to these three types of stresses. Furthermore, our investigation revealed pathways that respond concurrently to all three stresses, shedding light on the interconnectedness of these stress response mechanisms.
-
We collected 192 transcriptome datasets of Tifleaf3 leaves and roots under normal conditions (CK), high temperature (40 °C/35 °C), drought (20% PEG 6000), and salt (100 mM/L NaCl). This data was deposited in the Short Reads Archive (SRA) from 2020 to 2023 (Supplementary Table S1).
Read mapping and differential expression gene identification
-
Trimmatic (version 0.36) was employed for the purpose of eliminating sequencing adapters and low-quality nucleotide sequences from the raw data[19]. Filtered sequences were identified using FastQC software version 0.12.0 (
www.bioinformatics.babraham.ac.uk/projects/fastqc/ ), and only high-quality reads were retained for subsequent analysis. The reference genome utilized in the present study was derived from the previously published pearl millet genome (PRJNA749489,www.ncbi.nlm.nih.gov/bioproject/PRJNA749489/ )[12], and an index file was created using Kallisto software (version 0.48.0, kallisto indx), for subsequent analysis. Kallisto enables direct comparison of RNA-Seq data with indexed data, facilitating the quantification of gene expression levels for individual samples (kallisto quant -b 100). This feature greatly streamlines the analysis process, and saves significant time[20]. Ultimately, the analysis of differential expression was conducted through the txiimport[21] and DESeq 2[22]. Genes that exhibited |log2(treatment group/CK)| ≥ 1 with p.adj-values less than 0.05 were considered to be significantly differentially expressed.Gene function annotation and transcription factor identification
-
Gene functions are annotated in the following four ways. The DIAMOND program was used with an E cutoff value of 1e-5[23] to query the NCBI non-redundant protein database (NR)[24]. Protein families were predicted using the software HMMER (version 3.1) (
www.hmmer.org )[25], based on the Pfam database (Pfam:http://pfam.sanger.ac.uk ). The Blast2GO program (www.blast2go.com )[26] was used for GO annotation based on NR annotation (cut-off E value ≤1e-10). The KEGG database (version 53) (www.kegg.jp/kegg/kegg1.html )[27] was searched by BLAST to determine the pathway in which genes were involved, and the expected cut-off value was 1 × 10−5.The transcription factor (TF) family is predicted by iTAK software (version 1.7a)[28] with default parameters. The Pearson correlation coefficient was utilized to predict co-expressed genes, based on transcript per million (TPM) values.
GO and KEGG enrichment analysis
-
GOseq R was used in the GO enrichment analysis of DEGs[29]. GO terms with adjusted p values < 0.05 were considered statistically significant. Benjamini and Hochberg methods were used to adjust p value. KEGG enrichment results were determined using KOBAS 3.0 (p-value < 0.05)[30].
-
The transcriptome of pearl millet subjected to high-temperature stress conditions was obtained for examination[12,16,31]. Transcriptome analysis resulted in the identification of 11,541 DEGs in leaves, and 14,021 DEGs in roots of pearl millet. This finding suggests a higher number of heat stress-responsive genes in the roots of pearl millet compared to the leaves (Fig. 1a).
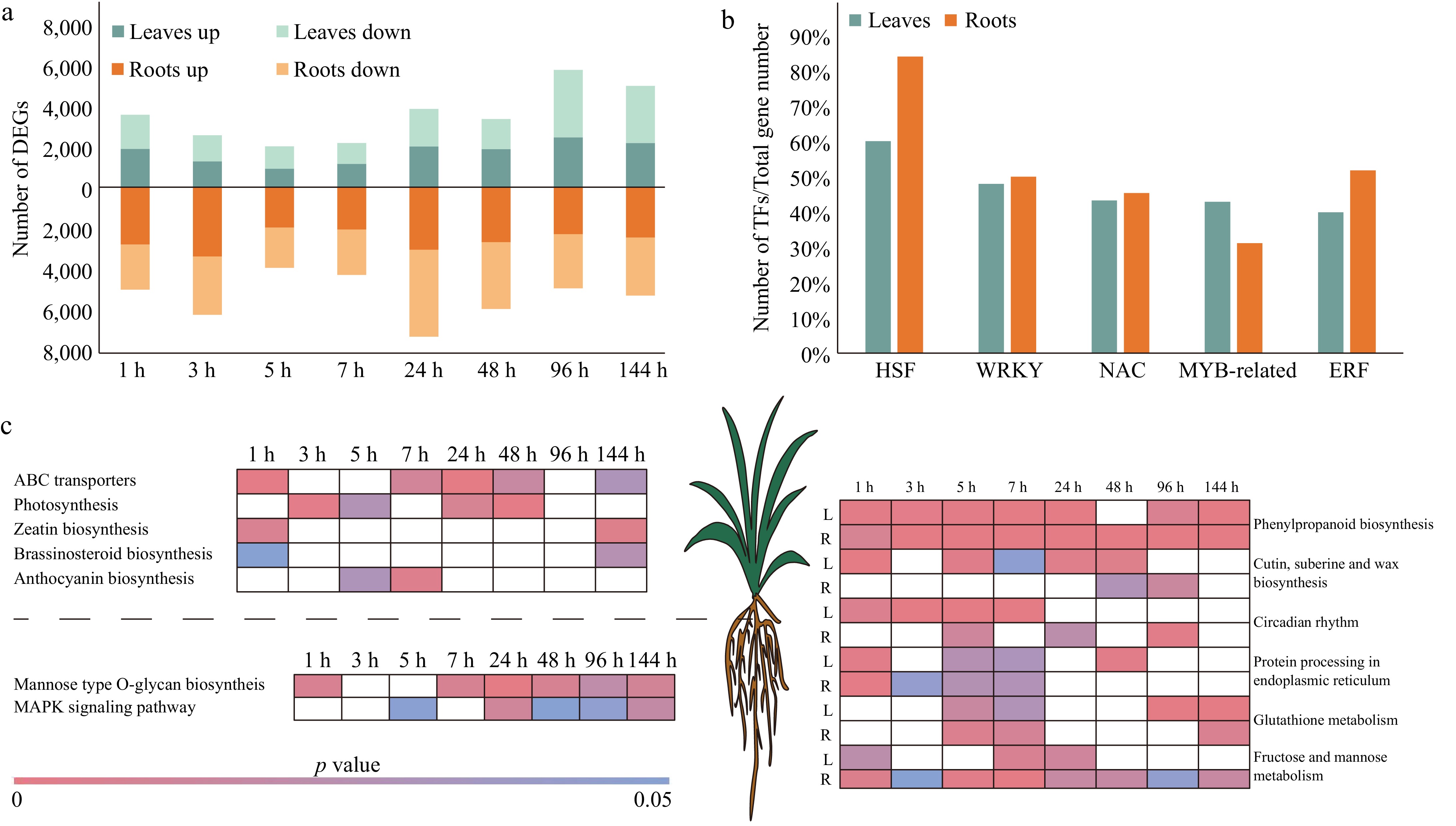
Figure 1.
DEGs in leaf and root of pearl millet at eight time points under high-temperature stress. (a) Number of DEGs in the leaves and roots of pearl millet under high temperature stress. Green and yellow represent leaves and roots respectively, and light and dark colors are used to distinguish up-regulated and down-regulated DEGs. (b) The percentage of differentially expressed HSFs, WRKYs, NACs, MYB-relatives, and ERFs. Green and yellow represent leaves and roots respectively. (c) KEGG enrichment analysis of DEGs. White indicates p value > 0.05.
Transcription factor (TF) regulation is essential for plants to effectively respond to various abiotic stresses. Among them, HSFs (heat stress factors), WRKYs, NACs (NAM, ATAF, and CUC), MYB-related, and ERFs (ethylene-responsive factors), have been extensively studied and are known to play crucial roles in mediating plant responses to abiotic stress conditions[32]. Under heat stress, 60% (differentially expressed HSFs/total HSFs, 15/25) and 84% (21/25) HSFs were differentially expressed in pearl millet leaves and roots, respectively (Fig. 1b, Supplementary Table S2). There were 47.96% (47/98) and 50% (49/98) WRKYs differential expressions in pearl millet leaves and roots, respectively, and the duration of upregulation in roots (1−7 h) was longer than that in leaves (1 h). In pearl millet leaves, 43.26% of NACs (61/141) responded to heat stress, and they were significantly up-regulated at 1−48 h. In pearl millet roots, 45.39% of NACs (64/141) responded to thermal stress and were significantly upregulated at 1−5 h. Compared with pearl millet leaves (42.86%, 33/77), 31.17% of pearl millet root MYB-related genes were up-regulated under high temperature stress. A total of 39.88% (67/168, 1−48 h) and 51.79% (87/168, 1−96 h) ERFs were differentially expressed in pearl millet leaves and roots, respectively.
Functional enrichment analysis of DEGs was performed separately for roots and leaves of pearl millet (Fig. 1c, Supplementary Table S3). Under high-temperature stress, DEGs in pearl millet leaves were enriched in other pathways besides tissue-specific pathways 'Photosynthesis' and 'Anthocyanin biosynthesis'. For example, the 'ABC transporters' pathway that regulates stomatal opening and closing was specifically enriched in the leaves of pearl millet[33]. Thirty-eight DEGs encoding ABCG were identified in pearl millet leaves (Supplementary Table S3). DEGs were specifically enriched in the 'Mannose type O-glycan biosyntheis', and 'MAPK signaling pathway' pathways in pearl millet roots. Under high-temperature stress, there were 12 (7 CYP86s (cytochrome P450 subfamily 86), 4 HHT1s (HOTHEAD), 1 PXG (plant peroxygenase) in leaves (1 h), and 22 (7 CYP86s, 1 ACE, 4 PXG, 3 FARs, 6 CYP94s, 1 WSD1) genes in roots (48 h), involved in the 'cutin, suberine, and wax biosynthesis' pathways, that were differentially expressed, respectively (Supplementary Table S3). The genes associated with the 'Protein processing in endoplasmic reticulum' pathway exhibited up-regulation in response to stress after 1 h of high-temperature treatment, and the number of DEGs in roots was higher than that in leaves (Supplementary Table S1). The above results show that the roots of pearl millet respond more strongly to high temperatures than the leaves.
Genes involved in ABA synthesis positively respond to drought stress in pearl millet roots
-
Under drought stress, there were 14,856 and 13,290 DEGs in the leaves and roots of pearl millet, respectively (Fig. 2a). Abscisic acid (ABA) is a typical phytohormone utilized by plants to effectively respond to abiotic stresses, particularly drought stress[34]. Its main function is to regulate stomatal closure, thereby reducing water loss during periods of stress[35]. Plants synthesize ABA through the carotenoid pathway, also known as the indirect pathway[36]. A total of 12 genes encoding ZEP (2 DEGs, zeaxanthin epoxidase)[37], NCED (3 DEGs, 9-cis-epoxycarotenoid dioxygenase)[38], ABA2 (2 DEGs, Abscisic acid deficient 2), and AAO (2 DEGs, aldehyde oxidase)[39], in pearl millet root were differentially expressed at 1, 24, 48, and 144 h after drought stress, and all of them were involved in ABA synthesis in plants (Supplementary Table S4).
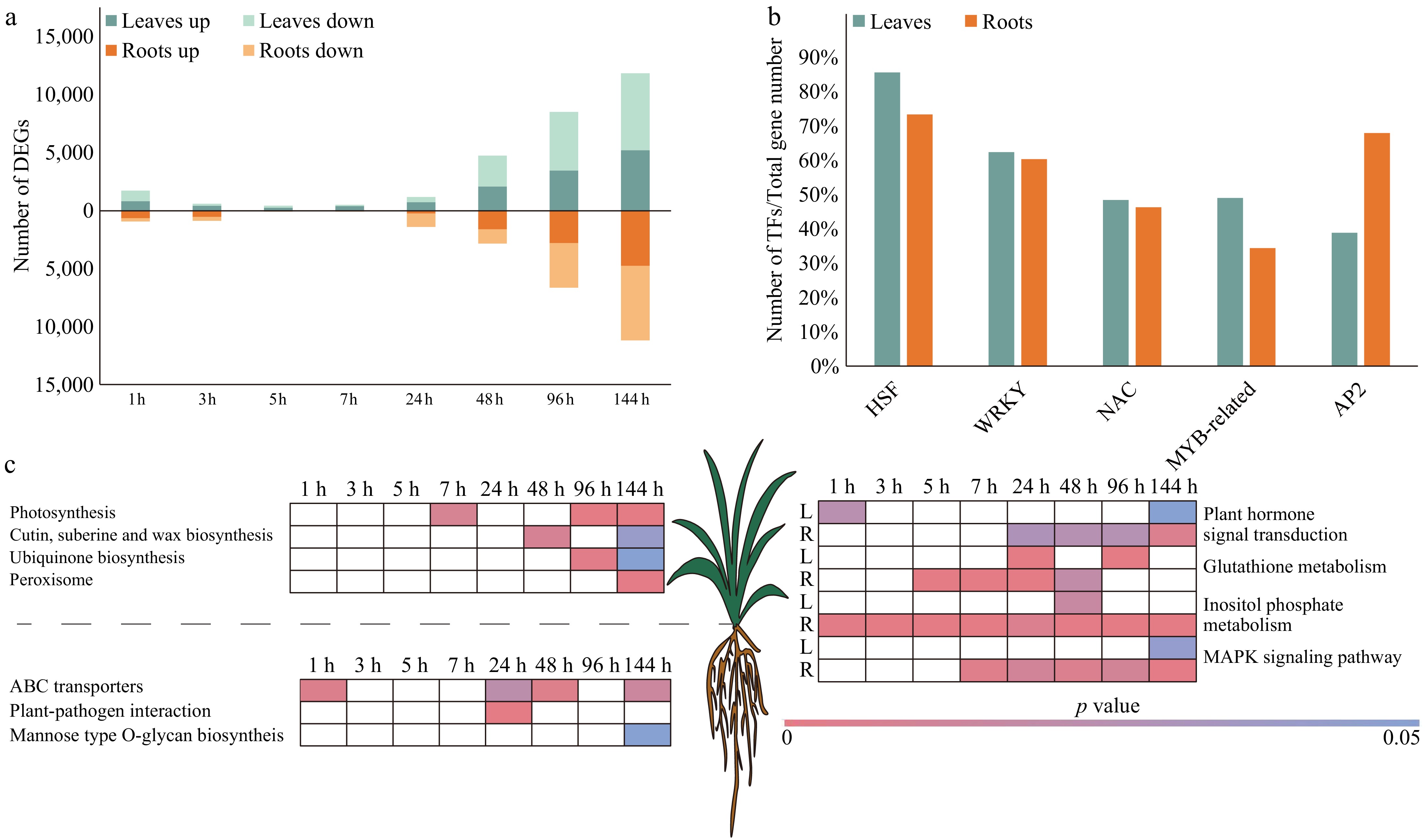
Figure 2.
DEGs in leaf and root of pearl millet at eight time points under drought stress. (a) Number of DEGs in the leaves and roots of pearl millet under drought stress. Green and yellow represent leaves and roots respectively, and light and dark colors are used to distinguish up-regulated and down-regulated DEGs. (b) The percentage of differentially expressed HSFs, WRKYs, NACs, MYB-relatives, and AP2s. Green and yellow represent leaves and roots respectively. (c) KEGG enrichment analysis of DEGs. White indicates p value > 0.05.
Research has shown that the expression levels of TFs change, due to drought stress, with 944 TFs found in pearl millet leaves, and 886 TFs in roots leaves (Fig. 2b, Supplementary Table S2). The active role of HSFs is essential not only for the ability of cells to cope with heat stress, but also for the response to drought stress[40]. In the context of drought treatment, 84% (21/25) and 72% (18/25) of HSFs displayed differential expression patterns in leaves and roots of pearl millet, respectively, and most of them were up-regulated. AP2s (apetala2) are a multifunctional TF family that mediates plant growth, and participates in plant panicle abiotic stress responses. After 24 h of drought treatment, 38.10% (8/21), and 66.67% (14/21) of ERFs responded to stress in pearl millet leaves and roots, respectively. There were 61.22% (60/98), and 59.18% (58/98) WRKYs differentially expressed in the leaves and roots of pearl millet, respectively, which were mainly expressed after 48 h of drought stress. In pearl millet leaves and roots, 47.52% (67/141), and 45.39% (64/141) of NAC expression changed after 48 h of drought stress.
KEGG and GO enrichment analyses were conducted on the DEGs identified in pearl millet leaves and roots, respectively (Fig. 2c, Supplementary Table S4). After five hours of exposure to drought stress, the expression of genes associated with 'Photosynthesis' in the leaves of pearl millet was affected. This shows that drought stress needs a certain amount of time to affect photosynthesis. In addition, genes involved in the 'Peroxisome' pathway were enriched at 144 h. There were 14 DEGs in this pathway, including 7 APXs (ascorbate peroxidase), 4 SODs (superoxide dismutase), 1 DAR (dehydroasrcorbate reductase), and 2 GRs (glutathione reductase) (Supplementary Table S4). Under drought stress, compared with leaves, DEGs in pearl millet roots exhibited significant enrichment in 'ABC transporters' and 'Plant-pathogen interaction' pathways. It was shown that in response to drought stress, the 'Plant hormone signal transduction Pathway', and the 'Phenylpropanoid biosynthesis Pathway' is enriched in both pearl millet leaves and roots.
The expression of genes related to phosphoinositides synthesis in pearl millet root was induced by salt stress
-
There were 4,975 and 14,168 genes differentially expressed in leaves and roots of pearl millet under salt stress (Fig. 3a), respectively. Differential expression of 288 TFs, and 661 TFs in leaves and roots (Fig. 3b, Supplementary Table S2). Under salt stress, the genes encoding HSFs were differentially expressed in leaves (40%, 10/25), and roots (40%, 10/25) of pearl millet, but most of them showed down-regulated expression patterns, which was different from the expression patterns under heat stress and drought stress (Supplementary Table S2). 15.6% (22/141), and 24.82% (35/141) NACs were differentially expressed in leaves and roots of pearl millet under salt stress, respectively. The NAC transcription factors upregulated in leaves exhibited a favorable response during the later phase of salt-induced stress. NACs in roots decreased from 1 to 96 h, and were up-regulated at 144 h. Auxin response factors (ARFs) are a crucial part of plant TFs that play a significant role in various physiological processes, including plant growth, development, and responses to environmental stresses. It is interesting that under salt stress, the amount of differentially expressed ARFs was much higher in pearl millet roots (85.71%, 18/21), than in leaves (9.52%, 2/21) (Supplementary Table S2).
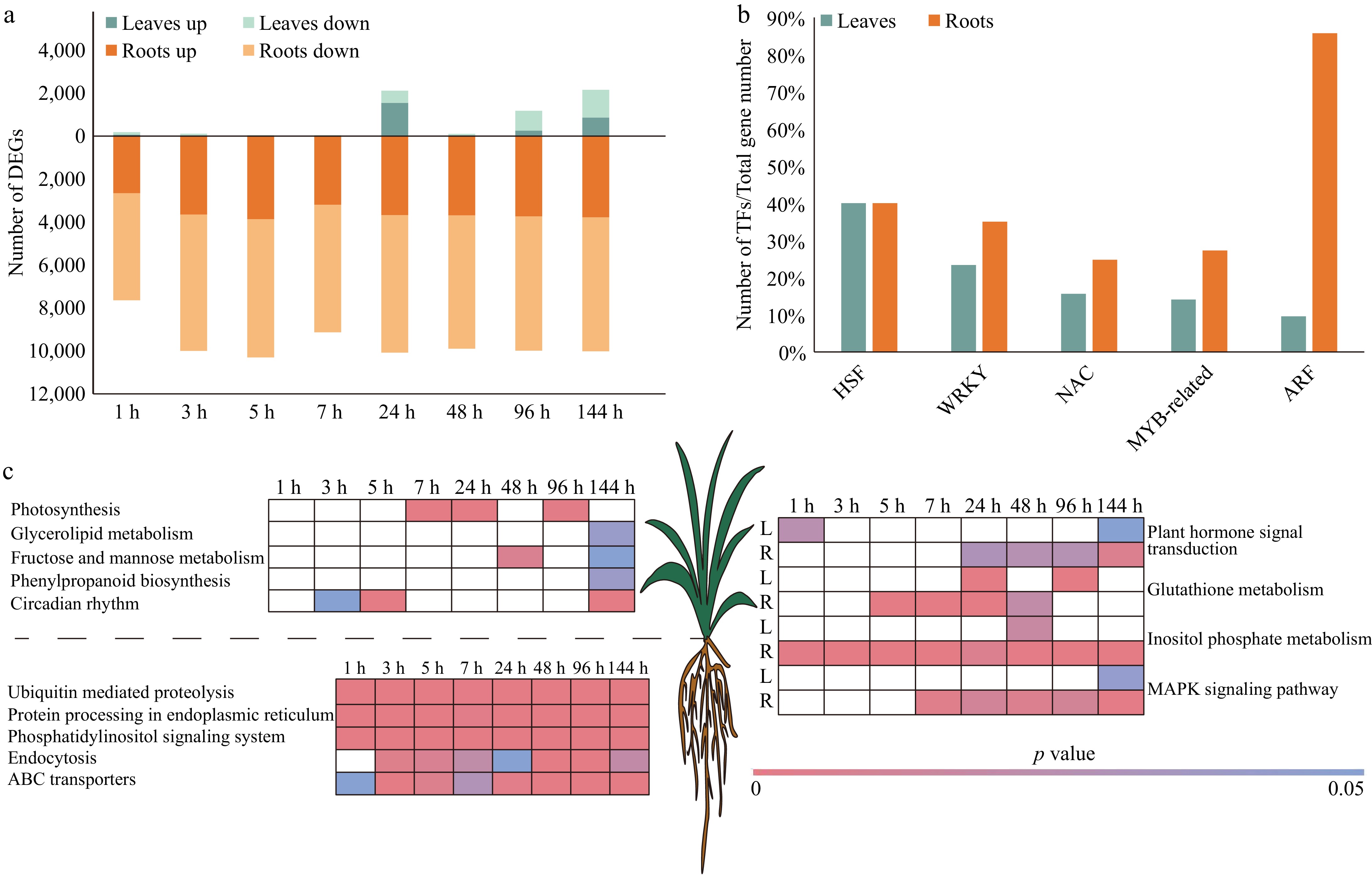
Figure 3.
DEGs in leaf and root of pearl millet at eight time points under salt stress. (a) Number of DEGs in the leaves and roots of pearl millet under salt stress. Green and yellow represent leaves and roots respectively, and light and dark colors are used to distinguish up-regulated and down-regulated DEGs. (b) The percentage of differentially expressed HSFs, WRKYs, NACs, ARFs, MYB-relatives, and ARFs. Green and yellow represent leaves and roots respectively. (c) KEGG enrichment analysis of DEGs. White indicates p value > 0.05.
The results of functional enrichment showed that DEGs in pearl millet leaves was enriched not only in 'Photosynthesis', but also in 'Circadian rhythm', 'Fructose and mannose metabolism', 'Phenylpropanoid biosynthesis', and 'Glycerolipid metabolism' (Fig. 3c, Supplementary Table S5). The genes involved in 'ABC transporters', 'Phosphatidylinositol signaling system', 'Protein processing in endoplasmic reticulum', 'Ubiquitin mediated proteolysis', and 'Endocytosis' pathway was differentially expressed in pearl millet roots under salt stress. As a signal lipid, phosphoinositides (PIs) participated in plant stress response processes[41]. A total of 19 DEGs related to phosphoinositides synthesis in pearl millet were identified. They encode INO1 (2 DEGs)[42], IMPA (1 DEGs, myo-inositol-3-phosphate synthase)[43], INPP5A (1 DEGs, inositol polyphosphate 5-phosphatase)[44], PIK3 (2 DEGs, phosphatidylinositol 3-kinase)[44], PIK4 (2 DEGs, phosphatidylinositol 4-kinase)[44], PIKFYVE (6 DEGs, 1-phosphatidylinositol-3-phosphate 5-kinase)[45], and PIP5K (5 DEGs) (Supplementary Table S5). These DEGs respond positively to salt stress, indicating that pearl millet roots actively synthesize phosphoinositides. The pathways of enrichment observed in the leaves and roots of pearl millet under salt stress include 'Plant hormone signal transduction', 'Glutathione metabolism', 'Fructose and mannose metabolism', 'Inositol phosphate metabolism', and 'MAPK signaling pathway-plant'.
Partial TFs and pathways simultaneously participate in response to three types of stress
-
To further investigate the similarities and differences of pearl millet in response to three types of abiotic stress, we conducted a Venn analysis on the DEGs under three stresses (high temperature, drought, and salt), and identified a total of 9,024 DEGs that simultaneously responded to the three types of stress. Among them, 1,023 DEGs responded to stress both in the leaves and roots of pearl millet, 237 DEGs specifically responded to stress in the leaves, and 981 DEGs specifically responded to stress in the roots (Supplementary Table S6). Under the three types of stress, 70 TFs were differentially expressed, belonging to 24 gene families (Fig. 4a, Supplementary Table S6). Among them, ERFs, bZIPs, HSFs, and WRKYs account for the most, accounting for 12.87% (9/70), 10% (7/70), 8.57% (6/70), and 8.57% (6/70), respectively. Through co-expression analysis, it was determined that 3,154 genes exhibited potential interaction capabilities with the aforementioned 70 TFs. Of these, 1,161, 925, 820, 550, and 410 genes were found to be co-expressed with GRF, Trihelix, GRAS, C2H2, and MYB-related transcription factors, respectively (Supplementary Table S6).
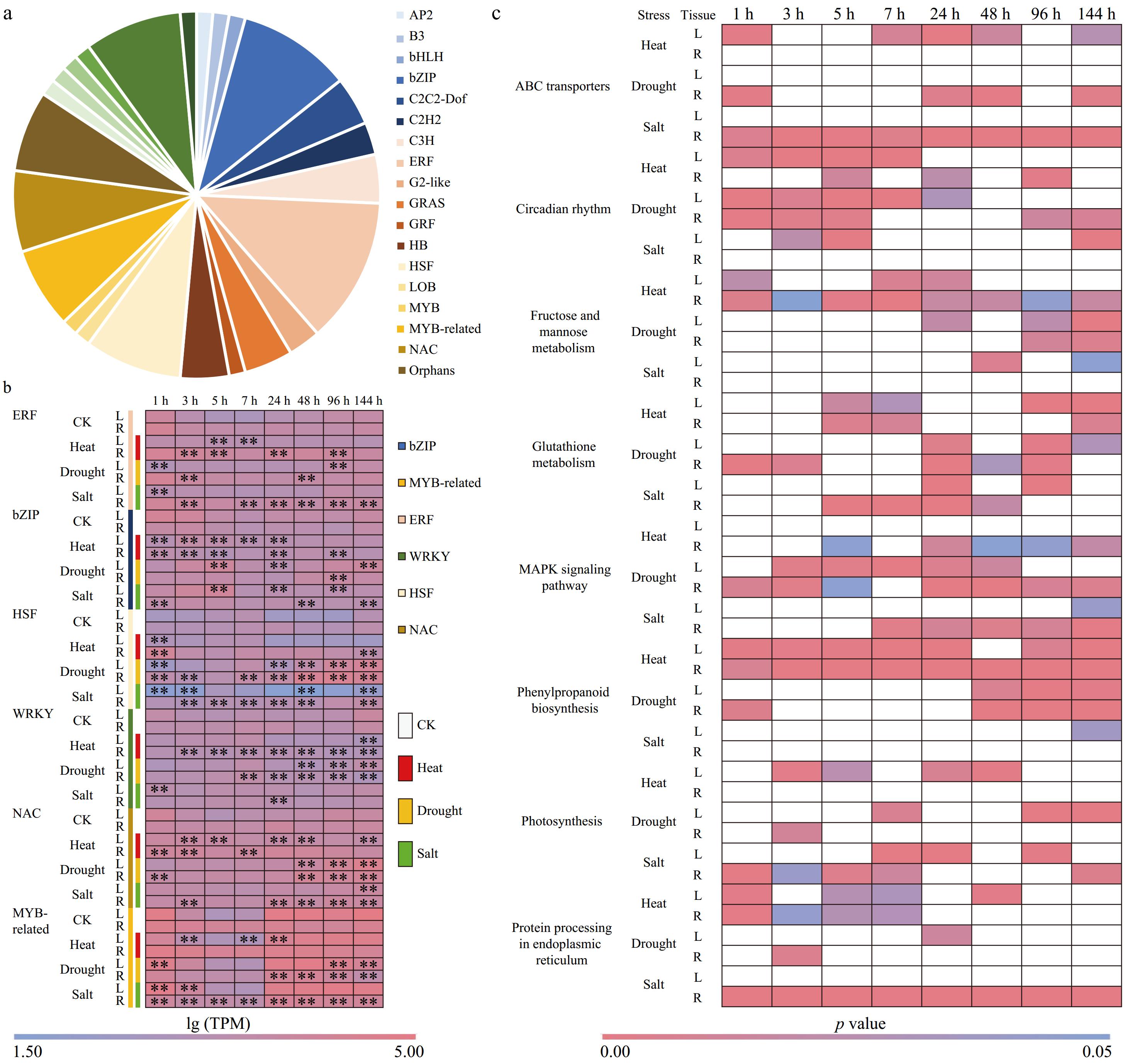
Figure 4.
TFs and KEGG pathways identified both under high temperature, drought and salt stress. (a) The ratio of specific TFs to differentially expressed TFs (70 TFs) in response to the three stresses showed that the proportion of ERFs, bZIPs, HSFs, and WRKYs gene families was 2.87% (9/70), 10% (7/70), 8.57% (6/70), and 8.57% (6/70), respectively. (b) Expression patterns of TFs in leaves and roots of Pearl millet responding to three stresses at eight time points. (c) The KEGG pathway was identified in all three types of stress (high temperature, drought and salt stress). The white square represents p value > 0.05.
We also compared the pathways under the three types of stress and found that pathways 'ABC transporter', 'Protein processing in endoplasmic reticulum', 'MAPK signaling pathway', Phenolpropanoid biosynthesis, 'Glutathione metabolism', 'Fructose and mannose metabolism', 'Photosynthesis', and 'Circadian rhythm' were enriched. This suggests the significance of these pathways in the responses to diverse stress stimuli (Supplementary Table S6). The 'Cutin, suberine and wax biosynthesis', and 'Zeatin biosynthesis pathway', demonstrate significant enrichment in response to heat and drought stress conditions. Additionally, genes associated with the 'Plant hormone signal transduction', and the 'Photoshatidylinositol signaling system pathway', exhibit active responses to drought and salt stress. Genes in the 'Sesquiterpenoid and triterpenoid biosynthesis', and 'Oxidative phosphorylation' pathways are differentially expressed under heat and salt stress. Genes related to 'Brassinosteroid biosynthesis' (heat stress), 'Peroxisome' (drought stress), 'Inositol phosphate metabolism' (salt stress), 'Ubiquitin mediated proteolysis' (salt stress), and 'Endocytosis' (salt stress) exhibited specific responses to heat stress, drought stress, and salt stress, respectively. These findings highlight the specificity of these pathways in response to various stress factors (Supplementary Fig. S1).
-
It is speculated that by 2050 the world population will be more than 9.7 billion, of which more than 65% of the population relying solely on agriculture for their livelihood[46]. Significant abiotic stresses such as extreme temperature, drought, and high salinity have been identified as major factors contributing to a substantial decline in global crop yields, ranging from 51% to 82% annually[47]. An in-depth exploration of the molecular mechanisms governing plant responses to abiotic stress is essential for the identification of key genes that are instrumental in enhancing the stress tolerance of plants. In the present study, we analyzed the transcription data of leaves and roots of pearl millet at eight consecutive time points under high temperature, drought, and salt stress, aiming to comprehensively understand the mechanisms of pearl millet responses to environmental stress. The results showed that: 1) In addition to the protein processes in the endoplasmic reticulum, plant cuticle and ABC transporters played a non-negligible role in pearl millet response to high-temperature stress; 2) Pearl millet response to drought stress is mediated by ABA signaling via the ABC transporter mechanism; 3) Phosphatidylinositol signaling systems are essential in pearl millet response to salt stress; and 4) The pathways that simultaneously respond to the three types of stress have different functions under different stress conditions.
The response pathways of pearl millet to heat stress are diverse
-
The pathway of protein processes in the endoplasmic reticulum has been widely reported to be involved in plant high-temperature stress, but other pathways that cannot be ignored have also been discovered in the study[12]. The DEGs of pearl millet under high temperature stress were analyzed, revealing an enrichment of DEGs in the pathways associated with 'Cutin, suberine and wax' and 'ABC transporters' pathways (Fig. 5a, Supplementary Table S3). Cutin, suberine, and wax, are the main components of the cuticle that covers the cell walls of the outer epidermis of plant organs[48], and play a significant role in facilitating plant responses to environmental stressors[49]. Research has revealed that under high-temperature stress, the plant cuticle can absorb heat, and block the heat transfer between plant organs and the surrounding area, to avoid heat damage to plant cells caused by high temperature[50]. Unsaturated fatty acids generate cutin and suberine-like substances under the action of CYP86s (cytochrome P450 subfamily 86)[51], HTH (HOTHEAD)[52], and PXG (plant peroxygenase)[52]. After 1 h of high temperature stress, the genes encoding cutin and suberine in pearl millet leaves were up-regulated (Supplementary Table S3). Furthermore, alterations in gene expression associated with the biosynthesis of cutin and suberine were observed, along with changes in the expression of genes encoding FARs, CYP94s, and WSD involved in wax synthesis in the roots of pearl millet (Supplementary Table S3). These results suggest that cutin, suberine, and wax have significant effects on plant response to high-temperature stress, possibly with different effects in different tissue types, potentially exhibiting specificity in different tissues[53].
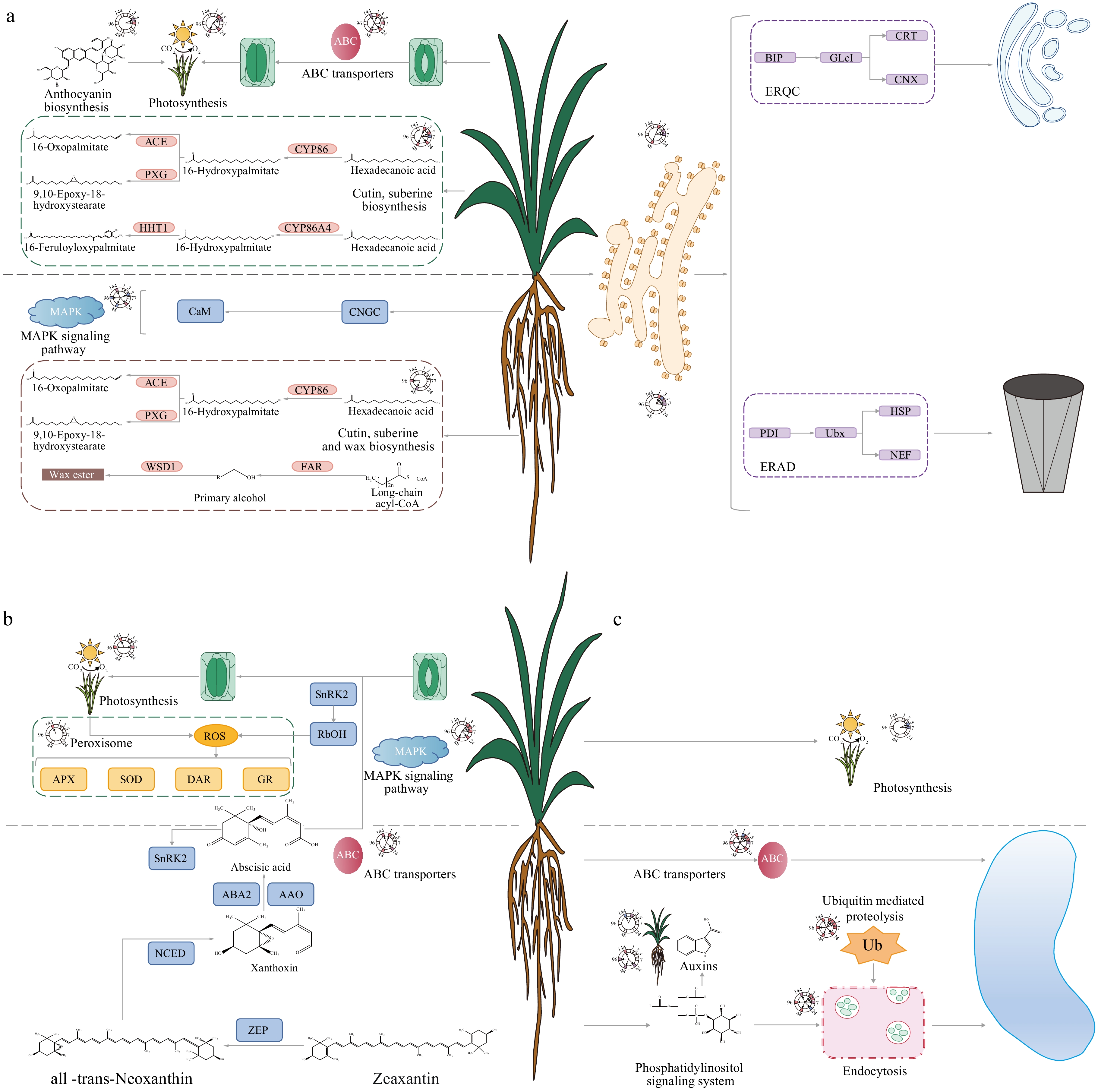
Figure 5.
Mechanism model diagram of pearl millet response to high temperature, drought, and salt stress. (a) Mechanism model of pearl millet response to high temperature stress. Cutin and suberine were synthesized by unsaturated fatty acids in the leaves of pearl millet under the action of CYP86s, HTH, and PXG. Besides Cutin and suberine, wax was synthesized by FAR, CYP94, and WSD in pearl millet root. The stomata of pearl millet leaves were closed by ABCG under high temperature stress. (b) Mechanism model of pearl millet response to drought stress. Epoxycarpenoids in the roots of pearl millet produce xanthoxin under the action of ZEP and NCED, and export it to the cytoplasm. Then, xanthoxin synthesized ABA through ABA2, and AAO. The synthesized ABA is transported by ABCG to the aboveground tissue of pearl millet, and SnRK2 is activated, leading to stomatal closure. In addition, ABA induces a large amount of ROS synthesis through RBOH, leading to the expression of peroxidase scavenging enzymes. (c) Mechanism model of pearl millet response to salt stress. Ins6P (Glucose 6-phosphate) synthesizes Ins (myo-inositol) under the action of INO1 and IMPA. The hydroxyl of Ins are sequentially phosphorylated by a series of PIK3, PIK4, and PIKFYVE kinases to generate three PtdIns monophosphates: PtdIns3P, PtdIns4P, and PtdIns5P. PtdIns4P further generates PtdIns(4,5)P2 under the action of PIP5K. PtdIns(4,5)P2 binds to AP2 protein to form CCPs (clathrin-coated pits). Subsequently, AP2 recruits clathrin and forms a clathrin shell. As the CCVs (clathrin-coated vesicles) mature, the DRP is responsible for separating the vesicles from the PM (plasma membrane). The isolated vesicles form ILVs (intraluminal vesicles) mediated by VPS and are transported to the vacuoles for degradation after ubiquitination.
Plants also regulate transpiration by regulating stomatal movement to cope with high-temperature stress. ABC transporters (ATP-binding cassettes) are a widely recognized family of proteins known for their central roles in various biological processes in plants, including growth, development, stomatal closure, transport of phytohormones, and enhancing tolerance to biotic and abiotic stresses[54]. The overexpression of ABCG25 in the ABC transporter family results in the decrease of stomatal size in plants[55]. Compared with the control conditions, the expression of the ABCG genes in pearl millet decreased under high-temperature stress, which indicated that pearl millet closed stomata by down-regulating ABCG genes to avoid water loss caused by high temperature (Fig. 5a, Supplementary Table S3). Previous studies have found that stomatal closure leads to a decrease in plant photosynthetic efficiency[56]. Under high temperature stress, genes involved in photosynthesis in pearl millet are differentially expressed (29 DEGs), indicating that photosynthesis in pearl millet is affected by high temperature and stomatal closure (Fig. 5a, Supplementary Table S3).
ABA is transported as a signal via ABC transporters to guard cells in response to drought stress
-
ABA is a vital phytohormone that acts as a key mediator in regulating plant reactions to drought stress[34]. ABA is synthesized in root tissue, then released into xylem vessels, and transported to the stem through ABC transporters, which alleviate the negative effects of drought by inducing stomatal closure of guard cells[57]. The synthesized ABA is transported by the ABC transporter to the guard cells to activate SnRK2 (SNF1-related protein kinase 2) to induce stomatal closure[57]. In pearl millet root, 33 ABCGs genes exhibited differential expression in the root tissue of pearl millet. Furthermore, 15 and 14 genes encoding SnRK2 were differentially expressed in the leaves and roots of pearl millet, respectively (Fig. 5b, Supplementary Table S4).
In plant leaves, ABA serves the dual functions of inducing stomatal closure, and generating large amounts of reactive oxygen species (ROS), a process mediated by the respiratory burst oxidase homolog (RBOH)[58]. To a certain extent, ROS acts as a signal molecule, activating different pathways to respond to stress[38]. However, excessive ROS leads to oxidative damage of lipids, proteins, RNA, and DNA molecules, which is toxic to plants[59]. ROS induced by RbOH is cleared by SOD and APX[60]. Eleven DEGs were involved in this process in pearl millet subjected to drought stress (Fig. 5b, Supplementary Table S4). The closure of stomata limits the assimilation rate of CO2, leading to a decrease in photosynthetic electron transfer, inhibition of photosynthesis, and production of ROS in chloroplasts[61]. GR and DAR have been reported to clear ROS in chloroplasts[62]. After exposure to drought stress, three related genes were differentially expressed (Fig. 5b, Supplementary Table S4). The drought stress significantly impacted 83 genes associated with photosynthesis in pearl millet leaves (Fig. 5b, Supplementary Table S4). The findings presented above demonstrate that drought stress induces synthesized ABA in the roots of pearl millet to be transported by ABC transporters to the leaves, which respond to drought stress by mediating stomatal movement in pearl millet leaves.
The phosphatidylinositol signaling system alleviates the negative effects of salt stress through endocytosis
-
PIs are important phospholipids in plants, including PI, PI3P, PI4P, PI(4,5)P2, and PI(3,5)P2[63]. Among these, PI4P and PI(4,5)P2 have been identified as key molecules involved in salt stress responses, acting through mechanisms such as the salt overly sensitive (SOS) pathway, endocytosis, and hormone regulation[64]. The current research investigation revealed that the expression of PIP5s, closely related to the biosynthesis of PI4P and PI(4,5)P2, was increased in pearl millet roots under salt stress (Fig. 5c, Supplementary Table S5). This suggests that pearl millet roots actively synthesize PI4P and PI(4,5)P2, in response to salt stress. This may help transmit salt stress signals to activate other genes involved in endocytosis and stress responses[44].
Endocytosis comprises two pathways, clathrin-mediated endocytosis, and membrane microdomain-associated endocytic pathways[65]. The endocytosis mediated by clathrin is the main pathway in plants. This process is completed by the recognition and recruitment of AP2 (assembly polypeptide 2)[66], the promotion of DRP (dynamin) for plasma membrane separation[67], ubiquitin related VPSs (AAA-ATPase vacuolar protein sorting), UBEs (E2 ubiquitin-conjugating enzyme), and CULs (cullin), and APCs (anaphase-promoting complexes)[68]. A total of 57 DEGs of pearl millet under salt stress were related to this process, encoding AP2 (5 DEGs), DRP (6 DEGs), VPSs (15 DEGs), UBEs (16 DEGs), CULs (6 DEGs), and APCs (9 DEGs) respectively (Fig. 5c, Supplementary Table S5). In summary, pearl millet root synthesizes a large amount of phosphoinositides under salt treatment for signal transduction to mobilize endocytosis and hormones to respond positively to salt stress.
The same pathway has different missions in response to different stresses
-
Improving plant stress tolerance is essential to address the emerging challenges of global food security and meet future food demands[12]. Therefore, it is crucial to explore pathways that respond simultaneously to different stresses. It was found that six pathways were involved in high-temperature, drought, and salt stress in this study. ABC transporters bind to ATP through coupling, hydrolyze ATP, and use the energy released to transfer different substrates such as vitamins, steroids, lipids, and ions across the membrane to regulate cell function (Fig. 6, Supplementary Table S6)[69]. Under high-temperature treatments, the ABCG subfamily present in pearl millet leaves responds to stress by affecting stomatal movement (Fig. 6, Supplementary Table S3)[70]. In contrast to the wild-type, the jat1-2 mutant of Arabidopsis demonstrated a significantly greater accumulation of ABA after treatment with exogenous ABA. Nevertheless, the endogenous ABA levels in both transgenic plants were similar to those found in the wild-type plant, indicating that ABCG16 is involved in the transport of ABA within Arabidopsis plants[71]. A total of 33 ABCGs in pearl millet demonstrated changes in expression in response to drought stress, indicating their possible function as transporters for the hormone ABA (Fig. 6, Supplementary Table S4). Under salt stress conditions, the sodium concentrations in the shoots and roots of AtABCG36-ox plants were noted to be lower compared to those in the wild type, with a notably greater effect in the roots. This finding indicates that AtABCG36 plays a role in enhancing the salt tolerance of plants by regulating sodium accumulation in both shoots and roots[72]. Transcriptome sequencing of pearl millet accession with different salt tolerance revealed that DEGs encoding ABC transporters were dominant in both genotypes[73]. Under salt stress, genes encoding ABCA, ABDB, ABCC, ABCD, and ABCG subfamily proteins in pearl millet root respond positively to salt stress, transporting ions to the extracellular (Fig. 6, Supplementary Table S5).
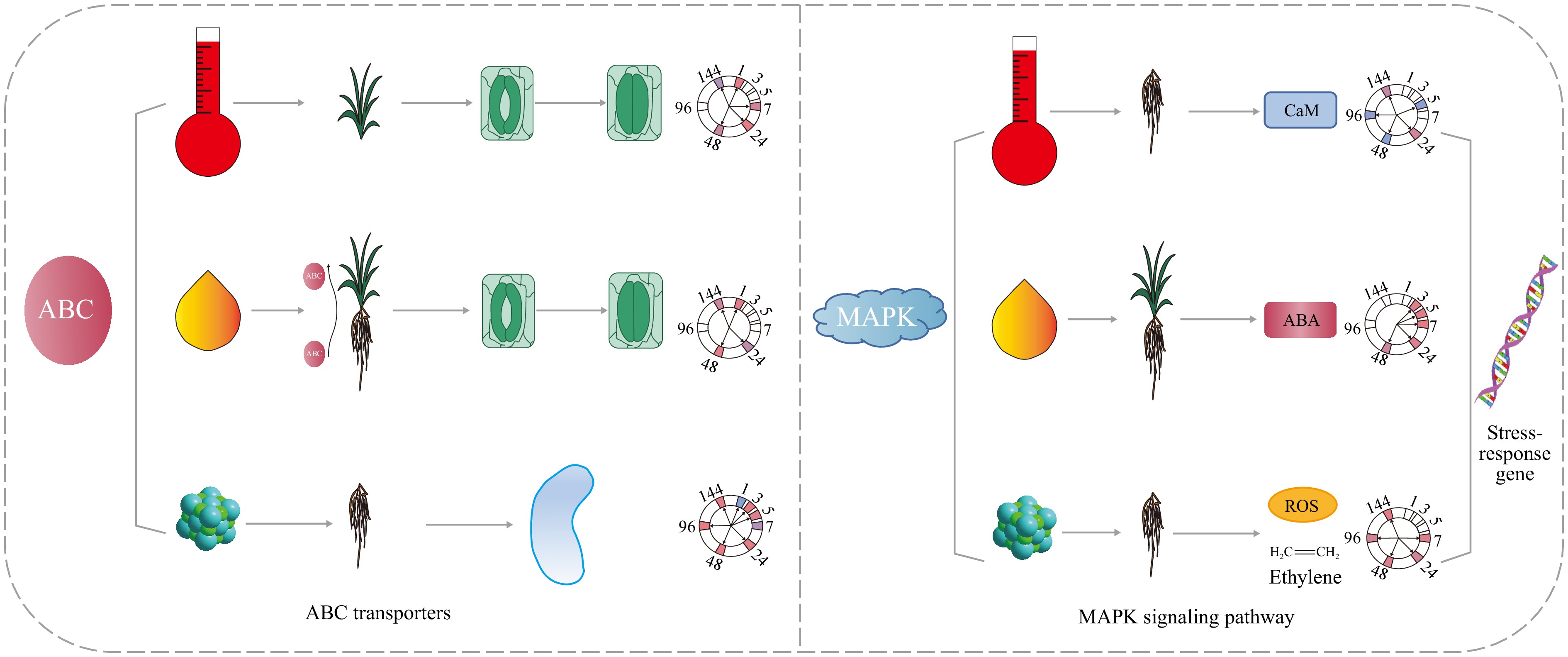
Figure 6.
The same pathway has different mechanisms in response to high temperature, drought and salt stress. ABC in the leaves of pearl millet responds to high temperature by mediating stomatal movement. ABC transporters in the roots of pearl millet play roles in transporting ABA and ions under drought and salt stress, respectively. The MAPK pathway is activated by Ca2+, ABA, and ROS signals under high-temperature, drought, and salt stress, respectively.
Plant response to abiotic stress mainly includes sensing, signal transduction and amplification, and receptor cell response to stress signals[74]. The MAPK signaling pathway is an important cell signal transduction mode, which is activated by the stimulation of upstream signal molecules, and the signal is amplified by phosphorylation and dephosphorylation step by step[74]. When the temperature increases, CNGC (cyclic nucleotide-gated channels) is activated, and the concentration of Ca2+ in the cytoplasm increases, forming Ca2+ signal[75]. Ca2+ signal was further transduced by CaM, and activated the MAPK pathway[76]. There were a total of 48 genes identified as being involved in the response of pearl millet roots to high-temperature stress (Fig. 6, Supplementary Table S3). Previous research has demonstrated that the expression of AtMAPKKK16-18 is regulated by ABA. Furthermore, plants overexpressing AtMAPKKK18 and AtMAPKK19 display an ABA-sensitive phenotype in response to ABA, suggesting that ABA can modulate the MAPK signaling pathway[77]. In the ABA signaling pathway, PYLs function as receptors of ABA signaling, PP2Cs (protein phosphatase 2C) function as negative regulators, and SnRK2s (SNF1-related Protein Kinases type 2) function as positive regulators in this signaling cascade[78]. Previous research has indicated that after 48 h of drought stress, pearl millet significantly expresses abscisic acid related genes such as the PYL gene family[16]. Under drought stress, 11 DEGs encoding PYL, 13 DEGs encoding PP2C, and 14 DEGs encoding SnRK2s were identified in the roots of pearl millet (Fig. 6, Supplementary Tables S4). The upstream signaling component MAPKKK within the MAPK cascade detects and integrates ROS signals, resulting in the subsequent phosphorylation of MAPKK[79]. In addition, ROS produced under abiotic stress, such as H2O2, can also activate the MAPK pathway. Under salt stress, the genes encoding RbOH in the roots of pearl millet were upregulated, while MEKK1 also showed an upregulated expression trend (Fig. 6, Supplementary Table S5).
To sum up, this study found that even if it is the same stress response pathway, its response organization, manner, and time are different in different stresses. For example, ABC transporters are involved in the response to high heat stress in leaves, while they function in the roots in response to drought and salt stress. Under high temperature, drought, and salt stress, the MAPK pathway is mediated by Ca2+ signaling, ABA signaling, and ROS signaling, respectively (Fig. 6).
-
This study constructed a transcriptome dynamic landscape map of pearl millet roots and leaves under high temperature, drought, and salt stress. The findings of the research indicate that the biosynthesis of cutin, suberin, wax biosynthesis, ABA biosynthesis, and phosphatidylinositol signaling pathways have significant implications in the responses of pearl millet to high temperature, drought, and saline conditions. Although ACB transporters and MAPK signaling pathways are activated in response to all three types of stress, they play different roles depending on the specific stressor.
This work was supported by the National Key R&D Program of China (2024YFF1001300), the Key Project of the Sichuan Provincial Natural Science Foundation (2025ZNSFSC0019), the Modern Agricultural Industry System Sichuan Forage Innovation Team (SCCXTD-2024-16), the Sichuan Province Breeding Research grant (2021YFYZ0013-5) and the Sichuan Province Key Research and Development Project for Young Talents (24MZG0189)
-
The authors confirm contribution to the paper as follows: project design and management, writing − draft manuscript preparation: Huang L, Yan H, Sun M; material collection and processing: Sun M, Zhang A, Jin Y, Lin C; bioinformatics analysis: Sun M, Yan H, Wang X; writing − manuscript revision: Huang L, Luo J, Zhou P, Zhang X. All authors reviewed the results and approved the final version of the manuscript.
-
The reference genome data and transcriptome raw reads are stored in the National Center for Biotechnology Information (PRJNA749489, PRJNA688001, PRJNA689619, PRJNA756390, PRJNA766308, PRJNA801196, PRJNA850644, and PRJNA850645).
-
The authors declare that they have no conflict of interest.
-
accompanies this paper at (https://www.maxapress.com/article/doi/10.48130/tp-0025-0017)
-
Received 12 January 2025; Accepted 7 April 2025; Published online 4 July 2025
-
The study is the first attempt to establish a dynamic transcriptomic landscape of pearl millet under heat, drought, and salinity stress.
The cutin, suberin, and wax biosynthesis, ABA biosynthesis, and phosphatidylinositol signaling specifically contribute to pearl millet's responses to heat, drought, and salt stress, respectively.
Although pearl millet employs similar mechanisms in response to all three stress types, its responses to each stress are distinct, as evidenced by the involvement of ABC transporters and the MAPK pathway regulatory components.
-
# Authors contributed equally: Min Sun, Haidong Yan, Ailing Zhang
- Supplementary Table S1 Overview of the data used for analysis.
- Supplementary Table S2 Expression of stress-related transcription factor families under high temperature, drought and salt stress.
- Supplementary Table S3 Functional enrichment and expression information of DEGs in pearl millet under high temperature stress.
- Supplementary Table S4 Functional enrichment and expression information ot DEGs in pearl millet under drought stress.
- Supplementary Table S5 Functional enrichment and expression information of DEGs in pearl millet under salt stress.
- Supplementary Table S6 DEGs, TFs, and pathway information identified both under high temperature, drought, and salt treatments.
- Supplementary Fig. S1 Pathways identified under high temperature, drought and salt stress.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press on behalf of Hainan University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Sun M, Yan H, Zhang A, Jin Y, Lin C, et al. 2025. The transcriptional dynamic landscape map of pearl millet in response to abiotic stress provides new insights into the responses of plants to stress. Tropical Plants 4: e024 doi: 10.48130/tp-0025-0017 

The transcriptional dynamic landscape map of pearl millet in response to abiotic stress provides new insights into the responses of plants to stress
- Received: 12 January 2025
- Revised: 01 April 2025
- Accepted: 07 April 2025
- Published online: 04 July 2025
Abstract: With the increasing frequency of extreme environmental events and the expansion of the global population, there is an urgent need to explore more stress-resistant genes to develop crops that can withstand extreme weather, to maximize agricultural production. Pearl millet, a significant cereal crop globally, demonstrates remarkable tolerance in harsh environments that are unsuitable for other cereal crops. It possesses unique stress-tolerant genetic resources distinct from those observed in conventional model plants. In our research, we conducted an examination of the transcriptome of pearl millet leaves and roots in response to high temperature, drought, and salt stress at eight time points. This investigation led to the construction of a comprehensive transcriptional landscape illustrating the intricate mechanisms underlying pearl millet's response to abiotic stress. Cutin, suberin, wax biosynthesis, abscisic acid (ABA) biosynthesis, and phosphatidylinositol signaling pathways have significant effects on the response of pearl millet to high temperature, drought, and salt stress, respectively. Although the same pathway was identified in the three treatments, their response to stress differs. Under different stresses, the cargos transported by the ABC transporter are different, and the signaling substances that activate the MAPK signaling pathway are also different. In conclusion, our research offers significant transcriptome mapping that sheds light on the response mechanisms of pearl millet to abiotic stresses. This study also highlights key genes that can be targeted for enhancing crop improvement efforts.
-
Key words:
- Pearl millet /
- High temperature stress /
- Drought stress /
- Salt stress /
- Transcriptional landscape map