









-
Polygala tenuifolia Willd., a member of the Polygalaceae family, comprises one of approximately 21 genera and over 1,000 species within the family. The dried roots of P. tenuifolia are traditionally consumed as a herbal infusion, commonly known as 'Xian Cha' (Thread Tea) in regions of Shandong and Shanxi provinces, China. Recognized as a key herb in traditional Chinese medicine (TCM), P. tenuifolia was first documented in the Shen Nong Ben Cao Jing (The Classic of Herbal Medicine) for its therapeutic effects in treating mental and neurological conditions, including depression, anxiety, insomnia, and Parkinson's disease. In addition to its listing in the Chinese Pharmacopoeia (2020 edition), P. tenuifolia is also officially recognized in the European Pharmacopoeia (Ph. Eur.), as well as in the pharmacopoeias of Korea, Japan, and in World Health Organization (WHO) monographs[1]. Extensive pharmacological studies have demonstrated that triterpenoid saponins isolated from P. tenuifolia possess sedative, expectorant, antitussive, and anti-inflammatory properties[2,3]. Moreover, recent findings have highlighted notable neuroprotective effects of the plant's aerial parts, particularly in enhancing cognitive function and alleviating memory impairments[4].
For medicinal crops, achieving maximal yields of bioactive constituents remains a significant challenge, intricately influenced by numerous variables including seedling selection, cultivation techniques, field management, harvest timing, primary processing methods, storage conditions, and transportation logistics[5]. Therefore, optimization of primary processing technologies plays a pivotal role in determining the final quality of medicinal crop products[6,7]. In the case of P. tenuifolia, conventional primary processing typically includes impurity removal, drying, water softening, extraction of the woody core, and slicing. However, studies have shown that extended exposure to water, such as through rinsing or soaking, can result in substantial losses of water-soluble active ingredients[8,9]. Importantly, post-harvest processing is not merely a mechanical operation but a dynamic biochemical process involving ongoing chemical and biological transformations even after harvest. Despite its significance, limited scientific literature currently addresses the specific post-harvest processing strategies for P. tenuifolia, leaving the influence of different techniques on its phytochemical profile largely unexplored[10]. Therefore, systematic investigation into the effects of various post-harvest processing methods is urgently needed to identify optimal protocols that preserve or enhance the pharmacologically active components of P. tenuifolia, thereby improving its therapeutic efficacy and product quality.
Currently, three predominant post-harvest processing methods are employed in the production of P. tenuifolia. The first method follows traditional primary processing techniques, wherein dried root-barks undergo rehydration as a critical preparatory step. The second method adopts a fresh-cut approach, which involves directly slicing fresh root-bark into smaller segments without prior drying. The third method resembles the fresh-cut technique but requires that the fresh roots be stored under cool conditions (15−18 °C) for several months before processing. Despite the widespread use of these methods, there remains a notable gap in the literature concerning their impact on the secondary metabolites of P. tenuifolia. To address this gap, quasi-targeted metabolomics offers a promising analytical strategy. This advanced, high-throughput technique enables comprehensive profiling of small-molecule metabolites, allowing researchers to evaluate both compositional and concentration changes in a wide spectrum of bioactive compounds. Utilizing the SCIEX QTRAP® 6500+ mass spectrometer, a triple quadrupole-linear ion trap hybrid instrument, quasi-targeted metabolomics leverages multiple reaction monitoring (MRM) mode to combine the breadth of untargeted metabolomics with the accuracy and sensitivity of targeted approaches[11,12]. Metabolite identification is achieved through an in-house database based on key parameters, including Q1, Q3, retention time (RT), declustering potential (DP), and collision energy (CE), all established using authenticated reference standards. Furthermore, this metabolomic platform integrates liquid chromatography-mass spectrometry (LC-MS) with multivariate statistical tools such as principal component analysis (PCA), and orthogonal partial least squares discriminant analysis (OPLS-DA). This combination forms a robust framework for analyzing chemical variability in biological systems[13]. The methodology's advantages, high throughput, rapid analysis, superior sensitivity, and effective data visualization, make it especially suitable for elucidating chemical transformations and compositional shifts associated with different regions, cultivars, and post-harvest processing techniques[14].
In this study, P. tenuifolia was subjected to three distinct post-harvest processing methods, and quasi-targeted metabolomics analysis was employed to characterize variations in its chemical composition. The primary objective was to elucidate how different processing techniques influence the profiles of functional compounds in P. tenuifolia, thereby offering a theoretical basis for optimizing processing strategies and ensuring quality control (QC) in the production of this traditional medicinal herb.
-
HPLC-grade methanol, phosphoric acid, and acetonitrile were obtained from Fisher Scientific (USA). Analytical-grade sodium hydroxide, n-butanol, and hydrochloric acid were purchased from Macklin Biochemical Co., Ltd. (Shanghai, China). Reference standards, including TF, 3,6-disinapoyl sucrose, and Polygalaxanthone III, were provided by Chengdu Chroma-Biotechnology Co., Ltd. (Chengdu, Sichuan, China). Ultrapure water (Wahaha, Hangzhou, Zhejiang, China) was acquired from a local distributor.
Materials
-
Fresh roots of P. tenuifolia were collected from various regions across Shaanxi Province between April and October 2023. Detailed information on the origins and weights of the samples is provided in Supplementary Table S1. All the samples were identified by professor Hong Zhang. The collected samples were transported to Yulin Guangjitang TCM Development Co., Ltd. (Yulin, China), and Jia County Yongtai Astragalus Planting Co., Ltd. (Yulin, China) for pre-cold storage and subsequent processing. All voucher specimens were authenticated and deposited at the Shaanxi Academy of TCM (Xi'an, China).
Preparation of samples for different post-harvest processing methods
-
All freshly harvested roots of P. tenuifolia were transported to Yulin Guangjitang TCM Development Co., Ltd. for post-harvest processing. In the Control group (C group), the root-barks were separated by manually extracting the wood core from the fresh roots. The obtained fresh root-barks were first dried in an oven at 60 °C for 6 h. After drying, they were rehydrated and conditioned at ambient temperature (21−24 °C) for 12 h, then cut into segments ≥ 3 cm in length according to commercial grading standards for P. tenuifolia root tubers (Yuanzhi Tong). Finally, these rehydrated segments were re-dried at 60 °C for another 6 h (Fig. 1a). In the Fresh group (F group), the fresh root-barks were directly segmented (≥ 3 cm) and immediately dried at 60 °C for 6 h, without rehydration or storage. In the Store group (S group), fresh roots were stored in a cool warehouse at 15–18 °C for 5 months before processing. After storage, root-barks were extracted and treated identically to the Fresh group (Fig. 1a).
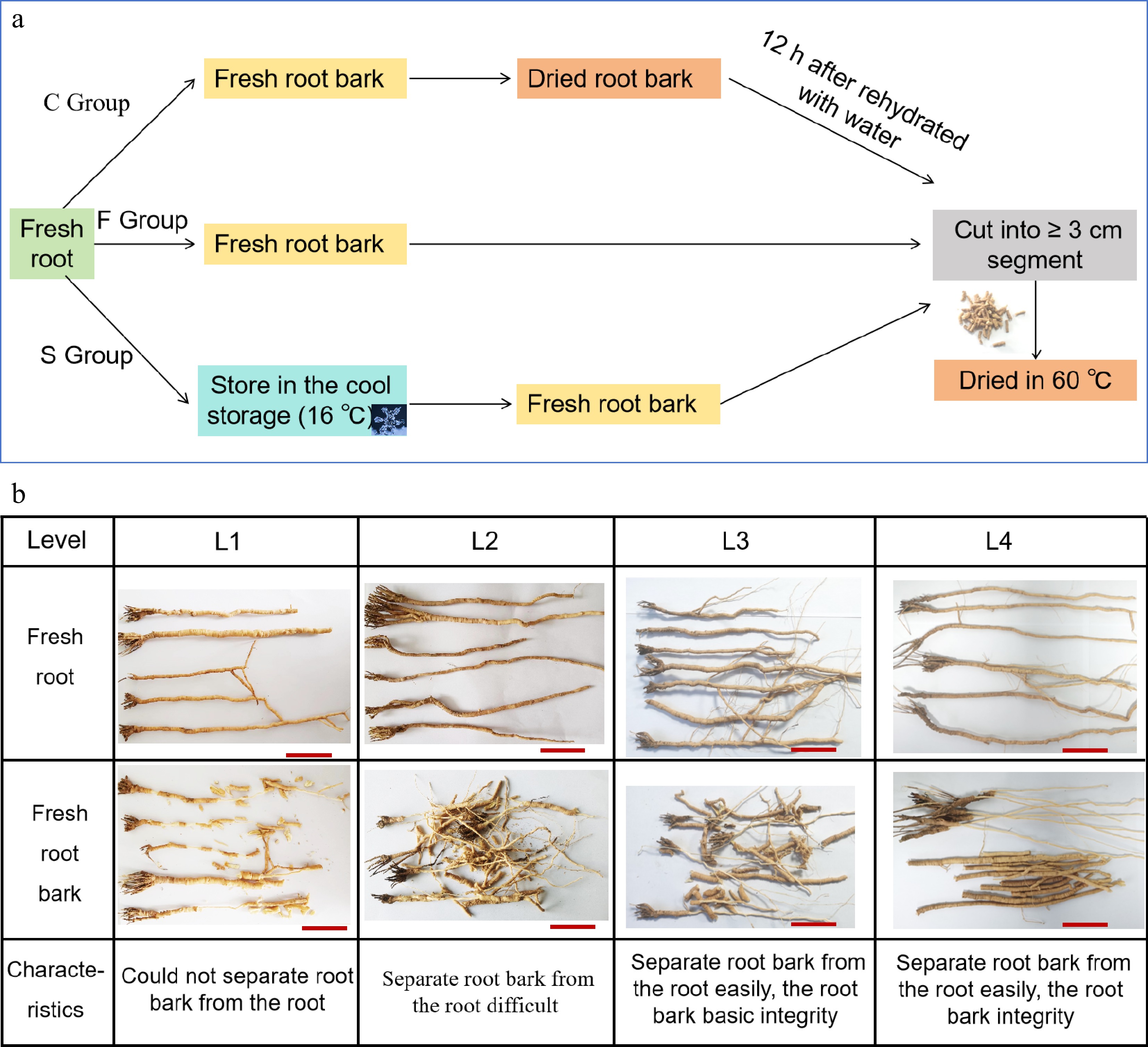
Figure 1.
(a) Preparation of samples for different post-harvest processing methods, (b) and the standards of the fresh root-bark.
Relative water content analysis
-
The relative water content of the root samples was calculated as previously described by Vincenzi et al. [15]. Briefly, fresh tissue samples (4.0–5.0 g) were weighed and recorded as W1 (sample weight before drying). The combined weight of the tissue sample and a pre-weighed glass weighing bottle before drying was recorded as W2. After drying the samples at 105 °C for 6 h, the combined weight of the dried sample and the weighing bottle was recorded as W3. The relative water content (%) was calculated using the following equation:
$ \mathrm{Relative\ water\ content\ ({\text{%}})=\dfrac{W_2\ -W_3}{W_1}\ \times100{\text{%}}} $ Standards of the fresh root-bark
-
To accurately evaluate the quality of the fresh root-barks, they were classified into four grades based on the difficulty of wood-core removal and the structural integrity of the root-barks during processing (Fig. 1b).
Metabolites extraction for HPLC analysis
-
All dried segments of P. tenuifolia were crushed using a grinder (CS-200 T, 50 Hz, 1,200 W), sieved through a 45-mesh screen, and stored at 4 °C. The extraction of TF was performed according to the Chinese Pharmacopoeia (2020). Briefly, 1.0 g of the powdered sample was placed into a conical flask with a stopper, mixed with 50 mL of 70% methanol, and subjected to ultrasonic-assisted extraction (40 kHz, 400 W) for 1 h at 30 °C using an ultrasonic processor (SB5200-DT, Ningbo Xinyi Ultrasonic Equipment Co., Ningbo, China). A 25 mL aliquot of the supernatant was collected and evaporated to dryness. The residue was hydrolyzed with 50 mL of 10% sodium hydroxide solution under reflux for 2 h. After cooling to room temperature, the pH of the solution was adjusted to 4–5 using hydrochloric acid. The solution was then extracted three times with water-saturated n-butanol (50 mL per extraction). The n-butanol fractions were combined and evaporated to dryness. The residue was dissolved in 25 mL of methanol, and the final solution was filtered through a 0.22 μm membrane for further analysis.
The extraction of 3,6-disinapoyl sucrose and Polygalaxanthone III was carried out following the method described in the Chinese Pharmacopoeia (2020). Briefly, 1.0 g of the powdered sample was placed in a conical flask with a stopper and extracted with 25 mL of 70% methanol by heating under reflux for 1.5 h. After cooling to room temperature, the solution was shaken thoroughly and filtered. The resulting filtrate was then passed through a 0.22 μm membrane filter and used for further analysis.
HPLC analysis
-
An aliquot (10 μL) of each extract was injected into a high-performance liquid chromatography system equipped with ultraviolet detection (HPLC-UV; Osaka Soda, Agilent, USA). Separation was performed on an Agilent 5 TC-C18 column (250 × 4.6 mm, 5 μm particle size). For the quantification of TF, the mobile phases consisted of (A) methanol and (B) 0.05% aqueous phosphoric acid. The elution was carried out isocratically with 70% A and 30% B for 15 min. The flow rate was maintained at 1.0 mL/min, and the column temperature was set at 28 °C. For the determination of 3,6-disinapoyl sucrose and Polygalaxanthone III, the mobile phases were (A) acetonitrile and (B) 0.05% aqueous phosphoric acid. The gradient elution was programmed at 18% (A) and 82% (B) for 50 min, with a flow rate of 1.0 mL/min and a column temperature of 30 °C, as specified in the Chinese Pharmacopoeia (2020).
Quasi-targeted metabolomics analysis
-
Quasi-targeted metabolomics was conducted by Novogene Bioinformatics Technology Co., Ltd. (Beijing, China) using an ExionLC™ AD system (SCIEX) coupled with a QTRAP® 6500+ mass spectrometer (SCIEX). Approximately 100 mg of each tissue sample was ground in liquid nitrogen, and the homogenate was resuspended in prechilled 80% methanol, followed by vigorous vortexing. The samples were incubated on ice for 5 min and centrifuged at 15,000 × g for 20 min at 4 °C[16]. Supernatants were collected for analysis. QC samples were prepared by pooling equal volumes of each experimental sample to monitor instrument performance and analytical stability. Metabolites were separated on a Waters XSelect HSS T3 column (150 × 2.1 mm, 2.5 μm) using a 20-min linear gradient at a flow rate of 0.4 mL/min. The mobile phases consisted of (A) 0.1% formic acid in water, and (B) 0.1% formic acid in acetonitrile. The gradient program was as follows: 2% (B) for 2.0 min, ramped to 100% (B) over 15.0 min, held at 100% (B) until 17.0 min, returned to 2% (B) at 17.1 min, and maintained until 20.0 min. The QTRAP®6500+ mass spectrometer was operated in both positive and negative ionization modes. In positive mode, the instrument settings were as follows: curtain gas, 35 psi; collision gas, medium; ion spray voltage, 5,500 V; source temperature, 550 °C; ion source gas 1 and gas 2, both at 60 psi. For negative mode, the ion spray voltage was set to −4,500 V with other parameters unchanged. Data acquisition was performed using MRM, and metabolite identification and quantification were carried out using the Novogene in-house MRM database. Three biological replicates were analyzed per experimental group, and one technical replicate was performed per biological sample.
Raw data files generated from the LC-MS/MS assays were processed using SCIEX OS (version 1.4). Metabolite quantification was performed using Q3 ion data, while metabolite identification was based on Q1 and Q3 ion pairs, RT, DP, and CE. Metabolite annotation was conducted using multiple databases, including KEGG (
www.genome.jpkegg ), the Human Metabolome Database (HMDB,www.hmdb.ca ), and LIPID MAPS (www.lipidmaps.org )[17,18]. For differential metabolite screening, the following criteria were applied: VIP score > 1.0 from orthogonal partial least squares-discriminant analysis (OPLS-DA), FC > 1.5 or < 0.667, and p-value < 0.05.Statistical analysis
-
All experiments were conducted with a minimum of three biological replicates. Statistical analyses were performed using SPSS Statistics software (version 24.0; IBM Corp., Chicago, IL, USA). One-way analysis of variance (ANOVA) and two-tailed Student's t-test were employed to evaluate significant differences between groups. A p-value < 0.05 was considered statistically significant. Correlation analyses were conducted using Pearson correlation coefficients within SPSS.
-
The root-barks of P. tenuifolia are recognized as the primary medicinal component of the plant, and the proportion of root-bark free from xylem serves as a critical quality indicator in the evaluation of P. tenuifolia decoction pieces (Yinpian). Historical texts, including the Compendium of Materia Medica and Lei's Treatise on Preparing Drugs, document the traditional practice of discarding the xylem to reduce processing challenges, while preserving the phloem for therapeutic use.
Recent phytochemical investigations have demonstrated that secondary metabolites such as polygalaxanthone XI (PXXI), 3,6-disinapoyl sucrose, polygalacin D (PGD), tenuifolin (TF), polygalic acid (PGA), and senegenin are present in higher concentrations in the xylem than in the phloem of P. tenuifolia[19]. Consequently, the extraction of xylem from P. tenuifolia is a precise and essential aspect of post-harvest processing. Farmers possess substantial empirical knowledge regarding the optimal timing and techniques for detaching the root-bark from the root. A commonly practiced method involves placing freshly harvested roots on drying platforms until they reach a pliable state conducive to manual separation. Practical experience indicates that maintaining an appropriate range of relative water content in the fresh roots is critical for effective root-bark separation. In this study, four batches of fresh roots (Spt1–Spt4) were collected and dried in a sunroom (Supplementary Table S1). The relative water content was monitored during the separation process, and four levels of separation difficulty were established based on the degree of challenge in removing the wood core while preserving the structural integrity of the fresh root-bark (Supplementary Table S2).
Across all four batches, the relative water content marking the transition from Level 2 (L2) to Level 3 (L3) ranged between 47% and 44%. The boundary between L3 and Level 4 (L4) was identified within a relative water content range of 42% to 37% in samples Spt2, Spt3, and Spt4, whereas in Spt1, this transition occurred between 37.35% and 35.38%. Moreover, the shift from L4 to Level 1 (L1) across all batches consistently took place when the relative water content fell between 33% and 24% (Supplementary Fig. S1). These findings indicate that the ease of root-bark separation is closely associated with the relative water content of fresh roots. Specifically, maintaining a relative water content of approximately 30% to 35% appears optimal for achieving efficient and complete separation of fresh root-bark.
The concentrations of three internal index components (TF, 3,6-disinapoyl sucrose, Polygalaxanthone III) were found to be independent of the water content in the root-barks that were separated from fresh roots
-
The processing methods employed for medicinal plants, including impurity removal, stacking, steaming, slicing, and drying, play a critical role in determining the concentrations of their active constituents[20,21]. For P. tenuifolia, the moisture content of root-barks derived from fresh roots is a crucial factor influencing post-harvest processing efficiency and product quality. To investigate the relationship between root-bark quality and moisture content, six samples were collected from each batch of fresh roots, and the concentrations of three pharmacologically important marker compounds were analyzed (Table 1). Correlation analyses were then conducted to evaluate the associations between moisture content and the levels of these three marker compounds. The results revealed a very weak negative correlation between TF content and moisture content, with a similar trend observed for 3,6-disinapoyl sucrose (Table 2). In contrast, Polygalaxanthone III exhibited a weak positive correlation with moisture content in samples Spt1, Spt3, and Spt4, while a weak negative correlation was observed in Spt2. Although most of these correlations were statistically significant, the overall associations were weak. These findings suggest that the moisture content of fresh root-bark has minimal influence on the concentrations of TF, 3,6-disinapoyl sucrose, and Polygalaxanthone III, implying that moisture variability during early post-harvest handling may not substantially impact the accumulation of these key bioactive constituents.
Table 1. The content of three internal index components (TF, 3,6-disinapoyl sucrose, Polygalaxanthone III) of the root-bark separated in different water content.
No. Relative water content of fresh root-bark (%) TF (%) Polygalaxanthone III (%) 3,6-disinapoyl sucrose (%) Spt 1-1 33.95 ± 0.15 2.49 ± 0.08 0.23 ± 0.01 1.18 ± 0.10 Spt 1-2 29.75 ± 0.06 3.21 ± 0.41 0.25 ± 0.03 1.06 ± 0.07 Spt 1-3 28.50 ± 0.16 2.71 ± 0.18 0.23 ± 0.02 1.10 ± 0.13 Spt 1-4 23.76 ± 0.12 2.77 ± 0.07 0.22 ± 0.02 1.04 ± 0.09 Spt 1-5 20.99 ± 0.08 2.80 ± 0.20 0.22 ± 0.03 1.08 ± 0.03 Spt 1-6 14.45 ± 0.33 2.70 ± 0.47 0.24 ± 0.02 1.19 ± 0.10 Spt 2-1 46.96 ± 0.22 2.65 ± 0.35 0.22 ± 0.01 1.03 ± 0.07 Spt 2-2 36.06 ± 0.17 2.72 ± 0.34 0.21 ± 0.01 1.15 ± 0.10 Spt 2-3 34.49 ± 0.22 2.34 ± 0.29 0.20 ± 0.01 1.08 ± 0.10 Spt 2-4 28.72 ± 0.17 3.08 ± 0.62 0.24 ± 0.04 1.04 ± 0.01 Spt 2-5 15.89 ± 0.11 2.84 ± 0.34 0.26 ± 0.03 1.10 ± 0.04 Spt 2-6 15.75 ± 0.31 2.64 ± 0.57 0.26 ± 0.02 1.13 ± 0.04 Spt 3-1 29.99 ± 0.20 2.96 ± 0.04 0.21 ± 0.00 0.94 ± 0.02 Spt 3-2 28.81 ± 0.14 3.17 ± 0.45 0.23 ± 0.04 0.92 ± 0.05 Spt 3-3 24.65 ± 0.04 3.65 ± 0.25 0.23 ± 0.01 0.94 ± 0.06 Spt 3-4 23.28 ± 0.08 2.64 ± 0.38 0.19 ± 0.01 0.98 ± 0.12 Spt 3-5 22.98 ± 0.31 3.44 ± 0.30 0.22 ± 0.02 1.00 ± 0.06 Spt 3-6 15.16 ± 0.11 3.21 ± 0.65 0.18 ± 0.01 0.96 ± 0.04 Spt 4-1 39.26 ± 0.21 2.61 ± 0.02 0.27 ± 0.04 1.13 ± 0.11 Spt 4-2 37.72 ± 0.21 2.70 ± 0.47 0.28 ± 0.01 1.09 ± 0.11 Spt 4-3 29.86 ± 0.07 2.71 ± 0.18 0.23 ± 0.01 1.16 ± 0.17 Spt 4-4 28.92 ± 0.10 3.21 ± 0.41 0.27 ± 0.01 0.96 ± 0.05 Spt 4-5 28.77 ± 0.30 2.77 ± 0.07 0.25 ± 0.01 1.08 ± 0.01 Spt 4-6 28.24 ± 0.11 2.49 ± 0.08 0.26 ± 0.02 1.16 ± 0.07 Table 2. Correlation analysis between content of three internal index components (TF, 3,6-disinapoyl sucrose, Polygalaxanthone III) and the water content fresh root cored.
Spt 1 Spt 2 Spt 3 Spt 4 Correlation
(r)Significant
(p value)Correlation
( r)Significant
(p value)Correlation
(r)Significant
(p value)Correlation
(Rho)Significant
(p value)TF −0.003 0.996 −0.262 0.615 −0.128 0.809 −0.086 0.872 Polygalaxanth-one III 0.097 0.855 −0.821 0.045 0.685 0.134 0.464 0.354 3,6-disinapoyl sucroseS −0.128 0.809 −0.461 0.358 −0.486 0.328 0.000 1.000 Specific impact between C group and F group in P. tenuifolia
-
The active constituents of TCM display varying degrees of water solubility. Consequently, prolonged exposure to water, via processing methods such as flushing or immersion, can substantially reduce the concentrations of these bioactive compounds[22,23]. Additionally, drying represents a critical step in the post-harvest handling of Chinese herbal medicines, as it markedly affects both the chemical composition and commercial value of the final product[24]. In this study, the impact of rehydration and re-drying procedures on the quality of P. tenuifolia root-barks were examined by comparing two processing groups: the F group (fresh-cut and directly dried), and the C group (rehydrated and re-dried after initial drying). To assess the potential influence of these processing differences, the levels of three key marker compounds, TF, 3,6-disinapoyl sucrose, and Polygalaxanthone III, were quantified in 18 batches of P. tenuifolia roots collected from northern Shaanxi (Supplementary Table S1). Each batch was divided into C and F groups according to the applied post-harvest method (Fig. 1a). For TF, four out of 18 batches showed higher concentrations in the F group than in the C group (Fig. 2a). Regarding Polygalaxanthone III, seven batches exhibited elevated levels in the F group (Fig. 2b). Notably, 17 out of 18 batches demonstrated significantly higher concentrations of 3,6-disinapoyl sucrose in the F group, with only one exception (Ct2), where the C group had a marginally higher value (Fig. 2c). These findings suggest that the rehydration and re-drying processes employed in the C group may contribute to the degradation or loss of key bioactive constituents, particularly 3,6-disinapoyl sucrose, thus potentially compromising the overall quality of P. tenuifolia products.
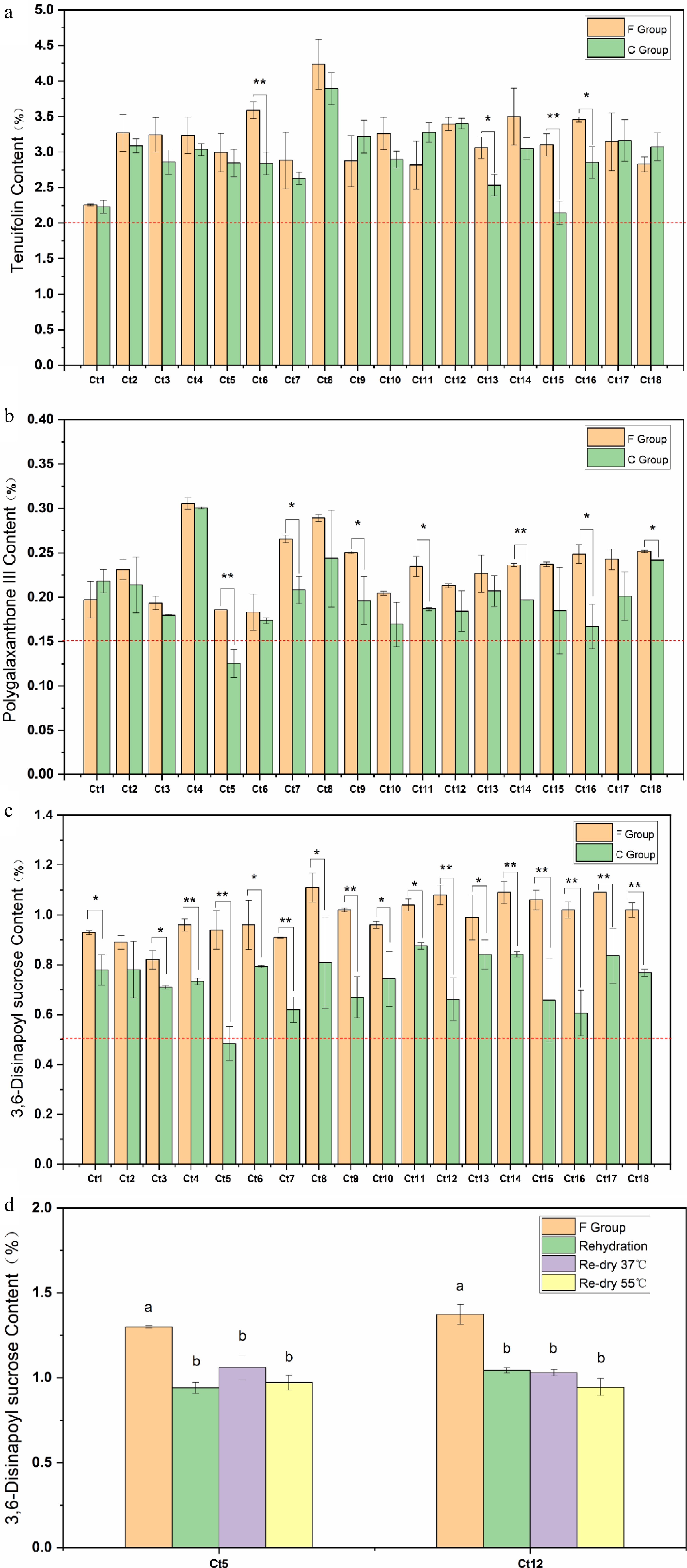
Figure 2.
Content of (a) TF, (b) Polygalaxanthone III, and (c) 3,6-disinapoyl sucrose in F group and C group. (d) Rehydration reduced the content of 3,6-disinapoyl sucrose in P. tenuifolia root bark. Error bars represent the SD of three biological replicates. Different letters indicate significant differences (p < 0.05).
To gain a comprehensive understanding of the differences between the C group and F group, the extract content was assessed and the appearance characteristics of samples evaluated from batches Ct1 to Ct18. Regarding extract content, 12 out of 18 batches displayed lower concentrations in the F group compared to the C group. Notably, five batches (Ct10, Ct12, Ct14, Ct15, Ct18) exhibited a reduction exceeding 5% (Supplementary Table S3). In terms of appearance, no observable differences in color or shape were found between the C and F groups (Supplementary Fig. S2), suggesting that fresh-cut processing does not compromise visual quality. Collectively, these results indicate that fresh-cut processing offers a preferable alternative to traditional processing for P. tenuifolia. Although a slight reduction in extract content was observed in some F group samples, the method preserves higher levels of key bioactive constituents, particularly 3,6-disinapoyl sucrose and Polygalaxanthone III. Moreover, the rehydration and re-drying steps involved in traditional processing appear to adversely impact the chemical quality of P. tenuifolia root-barks. Therefore, the fresh-cut method is recommended for enhancing both efficiency and the preservation of pharmacologically important compounds.
Rehydration reducing the quality of P. tenuifolia root-barks
-
To further investigate the cause of 3,6-DS reduction during post-harvest processing, targeted content analyses were performed on two representative batches, Ct5 and Ct12, which exhibited the most pronounced decreases among the 18 batches evaluated. Rehydrated samples were collected using liquid nitrogen snap-freezing followed by refrigeration-rubbing, while re-drying treatments were conducted at 55 and 37 °C, respectively. The fresh-cut (F group) samples were used as controls. All measurements were standardized based on the dry weight of the samples. The results revealed that the concentrations of 3,6-disinapoyl sucrose decreased by 27.7% and 24.1% in the Ct5 and Ct12 batches, respectively, following rehydration (Fig. 2d). In contrast, the re-drying process had a negligible impact on the compound's content. These findings suggest that the observed differences between the C group and F group are primarily attributable to the rehydration step, which appears to be the critical factor responsible for the degradation of 3,6-disinapoyl sucrose.
The efficacy of fresh processing methods in preserving bioactive ingredients, as compared to traditional techniques, has been explored across various TCM studies. For instance, Cai et al. reported that while the cutting method, cut thickness, and drying approach did not significantly impact the total saponin content in Ophiopogonis Radix, they exerted notable effects on total polysaccharide and flavonoid levels[25]. Similarly, Zhang et al. demonstrated that fresh processing methods significantly preserved four key active compounds, baicalin, wogonoside, baicalein, and wogonin, while maintaining the structural integrity of 18 bioactive ingredients in Scutellaria baicalensis[26]. In contrast, Bai et al. observed that root slicing led to a marked reduction in the total concentration of these four principal constituents[8]. In the context of Glycyrrhiza (licorice) processing, rehydration and re-drying are often associated with traditional techniques, which tend to be more complex than fresh processing. Flavonoids have been identified as the principal pharmacologically active components responsible for licorice's anti-arrhythmic properties, providing cardioprotective effects such as enhancing myocardial contractility and alleviating ischemic injury.
The oligosaccharide esters in P. tenuifolia are inherently chemically unstable, particularly under aqueous and elevated temperature conditions. Under such environments, the ester bonds within these compounds are prone to hydrolysis, resulting in the formation of corresponding secondary glycosides and/or aglycones. Xu et al. reported that 3,6-disinapoyl sucrose decomposes when exposed to aqueous heating, generating sibiricose A6, TF C, and sinapic acid as its main degradation products[27]. Consistent with these findings, the comparative analysis revealed a significantly elevated concentration of sinapic acid in the Control group (C group) relative to the Fresh-cut group (F group), as shown in Supplementary Table S4. Regarding the impact of water content on active ingredient preservation, Sun et al. demonstrated that fresh processing techniques more effectively maintain flavonoid levels in Glycyrrhiza (licorice) compared to traditional methods[28]. They attributed the observed loss of bioactivity in conventionally processed samples to glycosidic bond cleavage facilitated by rehydration and re-drying steps. Similarly, research on Astragali Radix (AR), a leguminous medicinal herb, revealed that optimal water content and controlled sweating time can significantly enhance the levels of polysaccharides and calycosin-7-O-glucoside[29]. Furthermore, fresh-cut processing of AR was shown to stimulate the release of key inflammatory and stress-related markers including TNF, SRC, ER-α, AKT, HSP90, and Caspase-3, in RAW264.7 macrophage cells, thereby indicating a potential enhancement in pharmacological activity[29].
These examples may help elucidate the mechanisms underlying the observed reduction in 3,6-disinapoyl sucrose content following rehydration in P. tenuifolia.
Prolonged cold storage of P. tenuifolia root-barks for several months at temperatures between 15 °C and 18 °C led to the degradation of TF and Polygalaxanthone III, which reduced the quality of the root-barks
-
It has been observed that the fresh roots of P. tenuifolia serve as the raw materials for both the Fresh-cut (F group) and Storage (C group) processing methods. However, the harvesting period for fresh roots is limited to a span of only 3 to 4 months annually. As a result, large quantities of fresh roots are typically stored in a cool environment (15–18 °C) for extended periods. Despite this widespread practice, no systematic research has been conducted to evaluate the impact of prolonged cold storage on the chemical quality of P. tenuifolia roots. To address this gap, fresh roots were stored in a temperature-controlled warehouse in Yulin, China, and the concentrations of three key pharmacologically active compounds, TF, 3,6-disinapoyl sucrose, and Polygalaxanthone III, were monitored. HPLC analysis revealed that the concentration of TF declined steadily during the first 2 months of storage, stabilized over the subsequent 2 months, and then exhibited a further decline in the fifth month (Supplementary Fig. S3a). Polygalaxanthone III levels remained stable throughout the first 4 months but significantly decreased during the fifth month (Supplementary Fig. S3b). In contrast, 3,6-disinapoyl sucrose remained relatively unchanged during the initial 3 months and subsequently increased over the next 2 months (Supplementary Fig. S3c). In addition, a visual comparison of external morphological features, including color and shape, between the F group and C group revealed no apparent differences. However, the chemical analysis clearly indicates that prolonged storage at 15–18 °C leads to a decline in the quality of P. tenuifolia root bark. These results support the conclusion that fresh-cut processing preserves the chemical integrity of P. tenuifolia more effectively than cold storage followed by processing.
A substantial gap remains in the current literature regarding the accumulation of secondary metabolites in the fresh roots of medicinal plants subjected to prolonged low-temperature storage following harvest, particularly over durations extending several months. Paeoniae Radix (peony root), a widely used herbal remedy documented in the Chinese Pharmacopoeia Commission 2020, is incorporated into numerous classical TCM formulations. It is renowned for its anti-inflammatory, antioxidant, anticoagulant, and sedative properties [30]. Zhu et al. reported that storing fresh Paeoniae Radix roots at 4 °C for approximately 1 month resulted in a slight increase in paeoniflorin content, which was likely attributable to the suppression of enzymatic degradation [24]. This storage condition also significantly mitigated discoloration in the roots. In contrast, the present study revealed a biphasic reduction in TF content in P. tenuifolia during cold storage. The first notable decline occurred after 2 months, potentially linked to stress responses induced by low temperature, dehydration, or suboptimal nutrient availability. A second significant decrease was observed at the five-month mark, which may be associated with altered phytohormone dynamics during early root germination. Supporting this hypothesis, comparative metabolomic analysis revealed that roots in the Storage group (S group) accumulated markedly higher levels of Gibberellin 7-1, Gibberellin 20, and Gibberellin 9 than those in both the Fresh-cut (F group) and Traditional (C group) processing groups (Supplementary Table S4).
The infection of Aspergillus flavus during the three post-harvest processing in P. tenuifolia
-
Aspergillus flavus is a ubiquitous fungal species commonly found in air and soil, capable of rapid dissemination through both waterborne and airborne pathways. Aflatoxin contamination, primarily caused by A. flavus, not only diminishes the quality of root-derived medicinal products such as P. tenuifolia, but also poses substantial risks to consumer safety. In this study, all processed P. tenuifolia samples were analyzed for aflatoxins B1, B2, G1, and G2 by an accredited third-party laboratory. The results confirmed that none of the tested samples contained detectable levels of these aflatoxins. Previous research has shown that temperatures exceeding 20 °C and relative humidity levels approaching 85% significantly increase the risk of A. flavus infestation in P. tenuifolia roots[31]. Notably, the fresh-cut processing method employed in this study utilizes undried raw materials, thereby eliminating the rehydration and moistening steps typically required in traditional processing workflows, steps known to introduce moisture and thus potential fungal contamination. Moreover, immediate sectioning of roots into 3–5 cm segments enhances drying efficiency and prevents the accumulation of localized moisture within the tubular structure of traditionally processed P. tenuifolia roots. These procedural improvements collectively reduce the likelihood of aflatoxin proliferation during both processing and subsequent storage.
Quasi-targeted metabolomics analysis of C group, F group, and S group
Identification of metabolites
-
The quasi-targeted metabolomics approach integrates the high-throughput screening capability of non-targeted metabolomics with the specificity and quantitative precision of targeted metabolomics [32,33]. The fundamental difference among these three metabolomic strategies lies in the databases employed for metabolite identification. Quasi-targeted metabolomics relies on curated in-house databases, such as that developed by Novogene, for metabolite annotation. In contrast, non-targeted metabolomics performs broader searches across publicly available databases, including HMDB, PubChem, METLIN, ChemSpider, and peer-reviewed literature [33,34]. Targeted metabolomics, by definition, focuses on the quantification and profiling of predefined metabolite classes [35]. To investigate metabolite variations in P. tenuifolia root-barks subjected to different post-harvest processing techniques, quasi-targeted metabolomics analysis was performed. A total of 1,743 metabolites were identified, including 885 compounds detected in positive ion mode and 858 in negative ion mode. These metabolites were classified into 17 major categories: amino acids and their derivatives (14.39%), flavonoids (11.61%), organic acids and derivatives (9.37%), lipids (8.96%), organoheterocyclic compounds (7.88%), carbohydrates and derivatives (7.74%), terpenoids (6.65%), phenylpropanoids and polyketides (6.38%), nucleotides and derivatives (6.04%), phenolic acids (5.30%), phenols and derivatives (3.94%), alkaloids and derivatives (3.60%), amines (2.24%), vitamins (1.77%), phytohormones (1.63%), alcohols and polyols (1.15%), and a miscellaneous group of low-abundance compounds (1.36%) (Supplementary Fig. S4, Supplementary Table S5).
QC of quasi-targeted metabolomics data
-
To ensure the reliability and consistency of the data acquisition process, QC samples were prepared by pooling equal volumes from all analyzed batches. The performance of the LC-MS system was assessed using these QC samples. Total ion chromatograms (TICs) for the QC samples in both positive and negative ion modes are presented in Supplementary Fig. S5a and S5b. The overlapping baselines across all three QC replicates in both ionization modes demonstrated minimal instrumental variation throughout the analytical run. Furthermore, PCA revealed tight clustering of the QC samples (Supplementary Fig. S6), indicating high reproducibility and confirming the overall stability of the LC-MS system during the experiment.
Identification and screening of differential metabolites (DM)
-
Metabolomics methodologies are designed to comprehensively profile or quantify a broad spectrum of compounds within biological samples. To identify differentially expressed metabolites DMs among the C group (rehydrated), F group (fresh-cut), and S group (fresh-stored), volcano plots were generated, and DM were screened based on fold change (FC) analysis and Student's t-test. Metabolites with variable importance in projection (VIP) > 1.0, p < 0.05, and FC > 1.5 (upregulated) or FC < 0.667 (downregulated) were considered significant. The results revealed that, compared to the C group, the F group exhibited 239 significantly upregulated and 169 downregulated metabolites (Fig. 3a). In the S group vs C group comparison, 403 metabolites were upregulated, and 100 were downregulated (Fig. 3b). Additionally, comparison between the S group and F group revealed 366 upregulated and 149 downregulated metabolites (Fig. 3c). Detailed results are provided in Supplementary Tables S4–S7. These findings indicate that fresh-root storage under cool conditions induces more extensive metabolic alterations than immediate fresh-cut processing. The larger number of DMs in the S group relative to both the C and F groups suggests that prolonged storage substantially impacts the metabolite composition of P. tenuifolia root-barks.
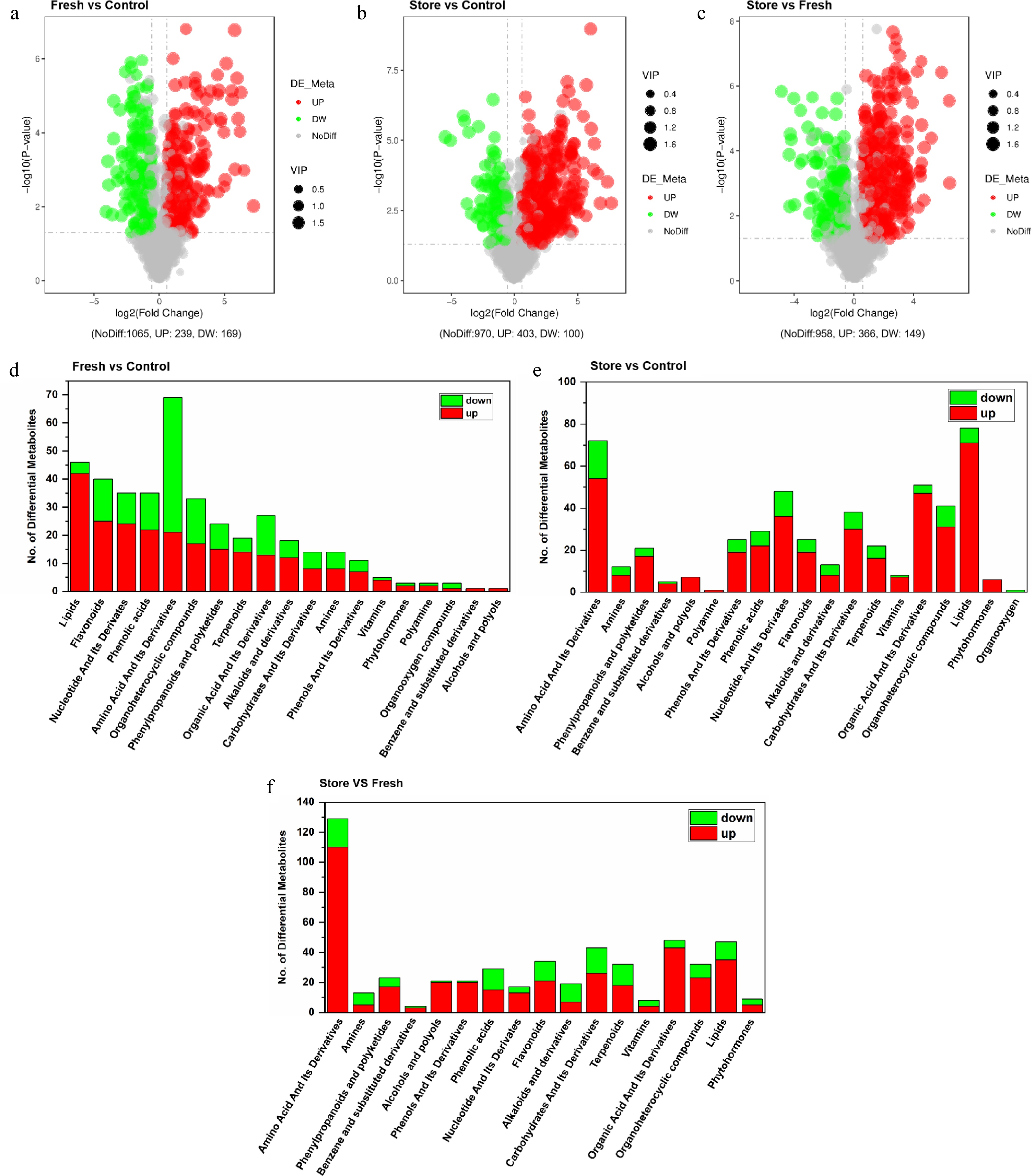
Figure 3.
The different metabolites in P. tenuifolia root-barks processed in three different post-harvest processing were identified and screened. The volcano maps exhibited the DMs between (a) F group and C group, (b) S group and C group, (c) S group and F group. The DMs were screened under the standards of Variable Importance in the Projection (VIP) > 1.0, FC > 1.5, or FC < 0.667 and T test p-value < 0.05. The DMs were displayed by classes between (d) F group and C group, (e) S group and C group, (f) S group and F group.
To further explore the metabolic changes among the different processing groups, the identified DMs were categorized according to their chemical classes. In the comparison between the F group (fresh-cut) and the C group (rehydrated), the largest class of DMs was amino acids and their derivatives, comprising 16.91% (69 out of 408), followed by lipids, which accounted for 11.27% (46/408) (Fig. 3d). Among the differential lipids, 42 were upregulated, and four were downregulated. Notably, 19 of the upregulated lipids were classified as Lysophosphatidylcholines (LysoPCs) and their derivatives (Supplementary Table S4). When comparing the S group (fresh-stored) with the C group, lipids emerged as the most dominant class of DMs, representing 15.50% (78 out of 503) (Fig. 3e). Within this group, 71 lipids were upregulated and four were downregulated, with 21 upregulated metabolites identified as LysoPCs and their derivatives (Supplementary Table S6). Furthermore, a downregulation of specific saturated fatty acids was observed, including docosanoic acid, 2-methoxyhexadecanoate, and capric acid. In contrast, several unsaturated fatty acids such as arachidonic acid, linoleic acid, linolenic acid, vaccenic acid, valproate, eicosapentaenoic acid, and glycerol-3-phosphocholine were significantly upregulated (Supplementary Table S6). Amino acids and their derivatives represented the second-largest class in the S vs C group comparison, accounting for 14.31% (72 out of 503) of the DMs (Fig. 3e). In the comparison between the S group and the F group, Amino acids and their derivatives constituted the largest proportion of DMs, making up 25.05% (129 out of 515), with 110 upregulated and 19 downregulated metabolites (Fig. 3f).
Saponins are the principal bioactive constituents of P. tenuifolia, and metabolomics analysis revealed the presence of various saponin compounds, including ginsenosides (Com_1364_neg), ursolic acid (Com_918_neg), notoginsenosides (Com_1321_neg), Raddeanin A (Com_1334_neg), ziyuglycosides (Com_1301_neg), and saikosaponins (Com_1425_neg). In addition to these glycosylated saponins, several aglycones such as madecassic acid (Com_1172_neg), and betulinic acid (Com_1173_neg) were also identified (Supplementary Tables S4, S6, S7). However, the relative abundance of these saponins across the three processing groups, fresh-stored (S), rehydrated (C), and fresh-cut (F), showed no consistent trend. For instance, ginsenosides were most abundant in the S group, ursolic acid levels peaked in the C group, while notoginsenosides and Raddeanin A were most enriched in the F group (Supplementary Fig. S7). These findings suggest that pharmacological evaluation may ultimately serve as the definitive criterion for assessing the quality of Polygala processed via these three methods.
As is well-established, P. tenuifolia exhibits significant neuroprotective properties, particularly in enhancing learning ability and alleviating memory impairments. Interestingly, the present metabolomic analysis revealed that several unsaturated fatty acids known for their neuroactive functions, such as valproate and eicosapentaenoic acid, were markedly upregulated in the F group (fresh-cut processing). Valproate, in particular, has received FDA approval since 1995 for the treatment of manic episodes and is widely used in the management of epilepsy, bipolar disorder, migraine prophylaxis, and neuropathic pain. Its proposed mechanisms of action include inhibition of γ-aminobutyric acid (GABA) transaminase, enhancement of GABA synthesis, and modulation of voltage-gated ion channels[36].
Amino acids and their derivatives represent a major class of primary metabolites and are widely regarded as important quality indicators in various biological matrices, including fruit and muscle foods. Their composition and abundance are highly sensitive to post-harvest storage conditions, such as temperature, duration, humidity, oxygen availability, and endogenous ethylene concentrations[37,38]. In the present study, a marked upregulation of two non-proteinogenic amino acids, L-theanine and GABA, were observed in the F group (fresh-cut processed P. tenuifolia). L-Theanine, best known as the predominant non-protein amino acid in tea leaves, has garnered increasing attention for its potential health benefits, including stress reduction, immune modulation, and antidepressant effects[39]. For instance, Chen et al. reported that administration of 0.3% L-theanine in drinking water significantly reduced serum corticosterone (CORT) levels in neonatal rats, indicating a strong antidepressant-like effect[39,40]. Similarly, GABA, another non-protein amino acid enriched in the F group, plays a pivotal role as an inhibitory neurotransmitter in the central nervous system. It is essential for modulating brain activity, cognitive processes, and behavior, and is involved in neural development via activation of GABAergic signaling pathways[41]. Collectively, these findings suggest that primary metabolites with known neuroactive functions, such as L-theanine and GABA, may contribute to the therapeutic effects of P. tenuifolia. Their enrichment in the fresh-cut group highlights the importance of considering processing-induced variations in primary metabolites when evaluating the functional and pharmacological potential of this medicinal plant.
Expression profiles of the relative contents of metabolites
-
To visually represent the distribution patterns of metabolite variation across the three processing methods, a total of 1,088 DMs were subjected to K-means clustering following data standardization. As shown in Fig. 4a and Supplementary Fig. S8, all metabolites were grouped into eight distinct clusters based on their relative abundance profiles in the P. tenuifolia samples. Specifically, clusters 1 and 2 encompassed 239 metabolites that exhibited relatively low abundance in the Traditional group. In contrast, cluster 3 comprised 90 metabolites with the highest levels detected in the Traditional group. Cluster 4, containing 137 metabolites, demonstrated peak abundance in the Fresh group, whereas clusters 5 and 6, totaling 219 metabolites, were characterized by the lowest levels across all groups. Notably, cluster 7, consisting of 314 metabolites, showed maximal abundance in the Store group, while cluster 8 included 89 metabolites with minimal expression. These clustering results underscore the substantial impact of post-harvest processing methods on the metabolite composition of P. tenuifolia, offering valuable insights into the biochemical consequences of each technique.
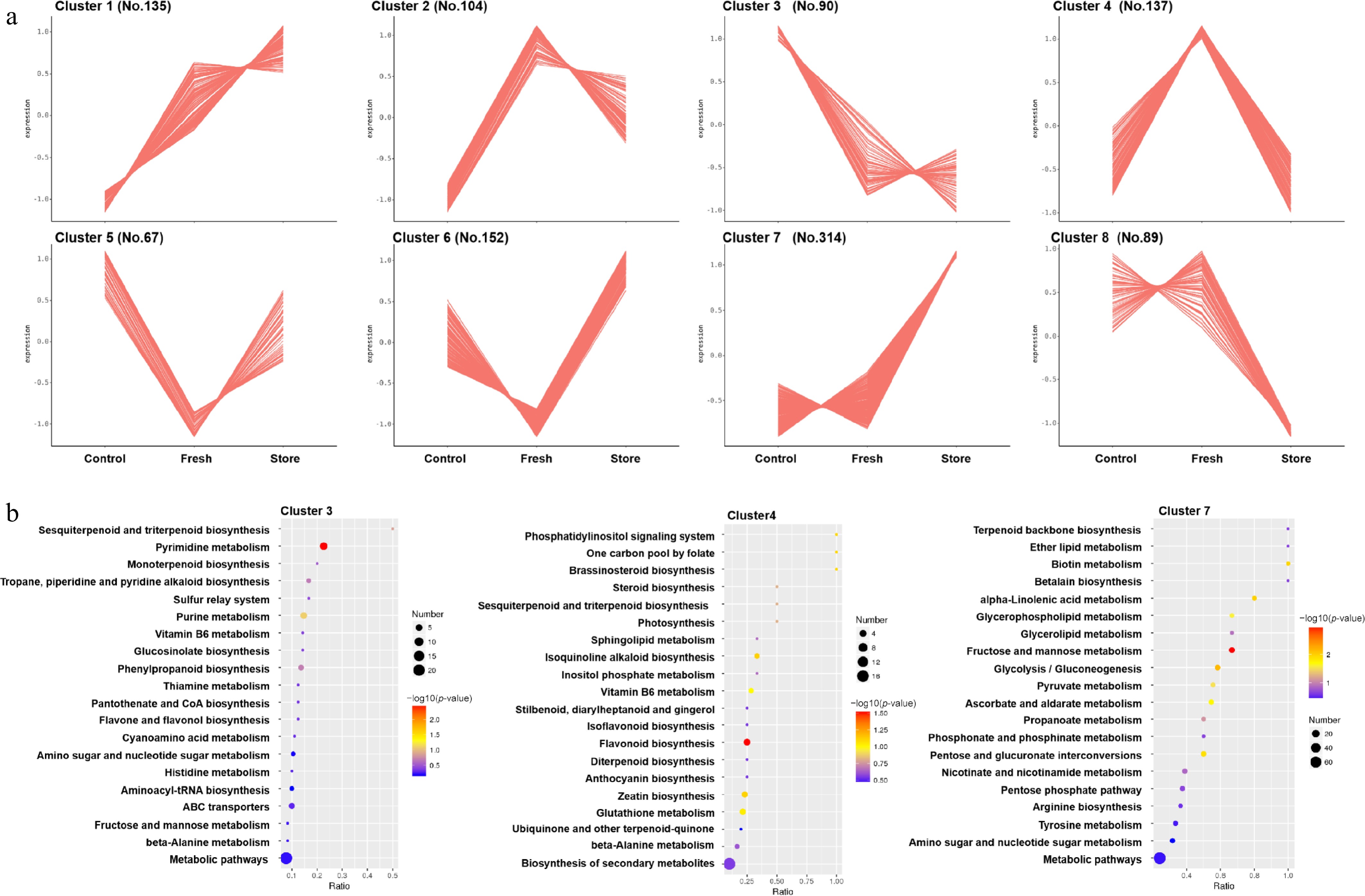
Figure 4.
K-means clustering analysis and KEGG analysis of the DM in the three different post-harvest processing. (a) Clusters of metabolites contents in response to the different post-harvest processing. No means the number of metabolites. (b) KEGG enrichment of the metabolites in cluster 3, cluster 4, and cluster 7. The color and the size of the point represents the p-value and the number of DMs in the corresponding path, respectively. An entire list of the p-value of the metabolites and the enriched KEGG pathway can be found in Supplementary Tables S8 and S9, respectively.
To further elucidate the biochemical mechanisms underlying the three distinct post-harvest processing methods of P. tenuifolia, Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis was performed on the metabolite clusters identified via K-means clustering. Specifically, metabolites from clusters 3, 4, and 7, which demonstrated high accumulation in the Traditional, Fresh-cut, and Store groups, respectively, were selected for the analysis. The results revealed that cluster 3, associated with the Traditional group, was enriched in pathways related to CoA biosynthesis and flavonol biosynthesis, suggesting that traditional processing may enhance the accumulation of flavones and flavonols, compounds known for their pharmacological benefits. In cluster 4, representing the Fresh-cut group, the enriched pathways included signal transduction, triterpenoid biosynthesis, alkaloid biosynthesis, and flavonoid and anthocyanin biosynthesis. This indicates that fresh-cut processing may favor the accumulation of terpenoids, isoquinoline alkaloids, flavonoids, and anthocyanins, all of which are functionally significant for the neuroactive and adaptogenic properties of P. tenuifolia. Meanwhile, cluster 7, which exhibited high metabolite levels in the Store group, was enriched in pathways related to backbone biosynthesis, fatty acid metabolism, and nicotinamide metabolism (Fig. 4b). These findings suggest that lipid metabolism is a key process influenced by long-term storage and may be directly linked to the alteration of aroma during this period. Notably, fresh P. tenuifolia roots are characterized by a distinctive fragrance that diminishes after several months of storage, even though the roots remain viable. This fragrance is composed of a complex array of terpenoids, aromatic compounds, aldehydes, ketones, esters, and other volatile substances. The present results strongly suggest that the modulation of lipid metabolism plays a central role in the transformation of the aromatic profile during storage.
Different post-harvest processing methods influence the phenylalanine related metabolic pathway
-
To specifically highlight the biosynthesis and metabolism of secondary metabolites, primary metabolic and phytohormonal pathways, such as purine metabolism, zeatin biosynthesis, aminoacyl-tRNA biosynthesis, nitrogen metabolism, and selenocompound metabolism, were excluded from the KEGG enrichment analysis. This focused profiling allowed for a more precise evaluation of processing-induced alterations in specialized metabolic pathways. The results revealed that direct processing of fresh root-barks (F group) significantly enhanced pathways associated with butanoate metabolism, cyanoamino acid metabolism, glyoxylate and dicarboxylate metabolism, and pantothenate and CoA biosynthesis when compared to the C group (Fig. 5a). In contrast, long-term storage prior to processing (S group) led to the upregulation of pathways involved in unsaturated fatty acid biosynthesis, isoquinoline alkaloid biosynthesis, butanoate metabolism, and linoleic acid metabolism (Fig. 5b). Notably, pathways associated with phenylpropanoid biosynthesis, including phenylalanine metabolism and flavonoid biosynthesis, were significantly enriched in both the F group and S group relative to the C group. This suggests that both fresh processing and storage modulate phenylpropanoid metabolism, a key pathway responsible for synthesizing a wide range of pharmacologically active compounds. Importantly, phenylalanine and tyrosine serve as critical precursors for phenylpropanoid biosynthesis, which subsequently contributes intermediates for flavonoid biosynthesis, compounds known for their antioxidant, neuroprotective, and anti-inflammatory properties[42].
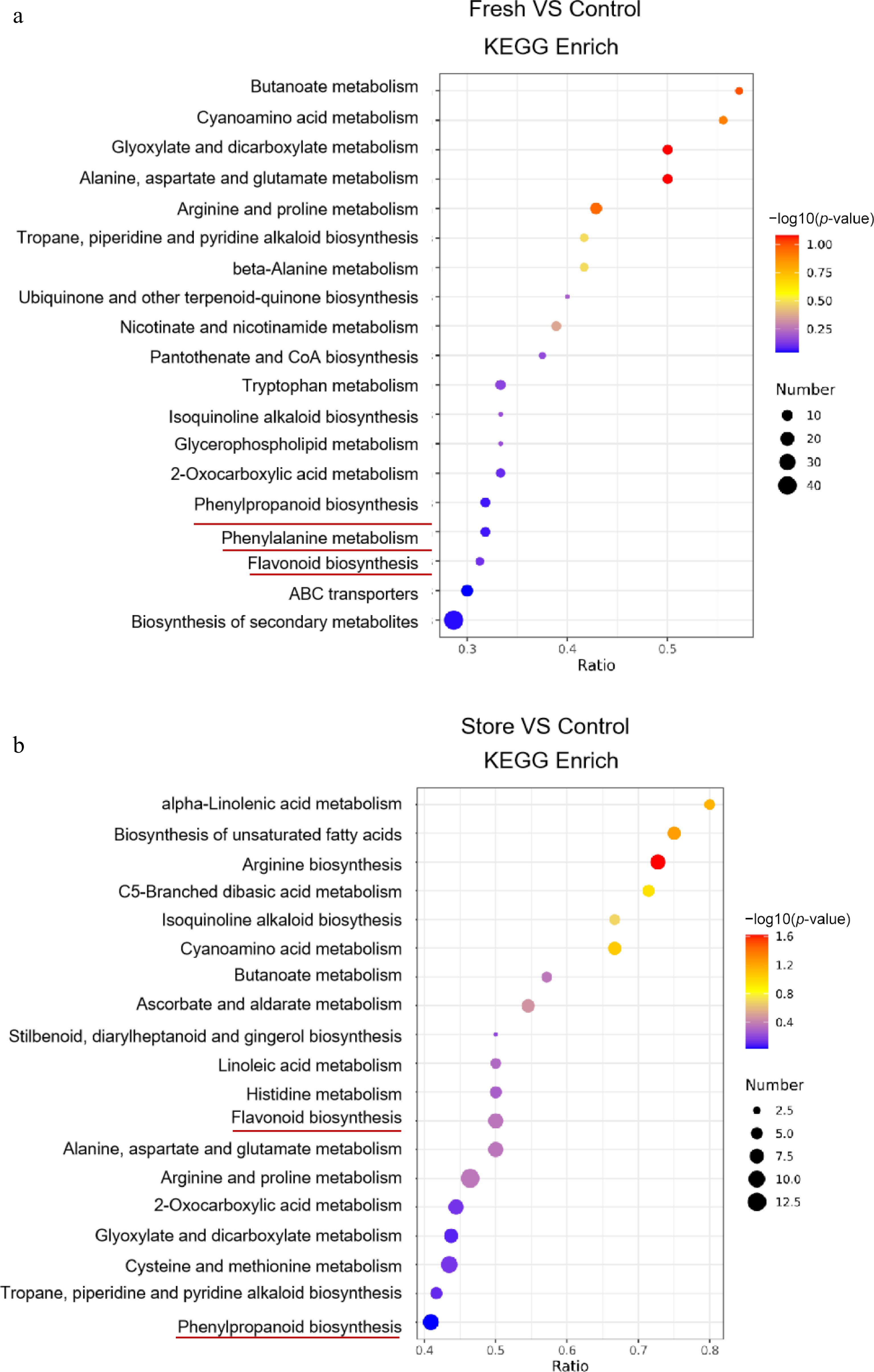
Figure 5.
The KEGG pathway enrichment is based on the DM in F group vs C group, and S group vs C group after removal of some pathways of primary metabolism or phytohormone. Number means the number of metabolites. The color and the size of the point represents the p-value and the number of DMs in the corresponding path, respectively.
To gain a more detailed understanding of the DMs involved in phenylalanine-related metabolic pathways, autoscale normalization of 13 key compounds was conducted for heatmap visualization. The resulting heatmap clearly delineated three distinct groups based on the accumulation patterns of these metabolites. Specifically, sinapine and ferulic acid showed high accumulation exclusively in the C group. In the S group, metabolites such as caffeic aldehyde, L-phenylalanine, p-coumaraldehyde, isoeugenol, and 2-hydroxycinnamate were significantly enriched. In contrast, the F group was characterized by elevated levels of p-coumaric acid, coumarin, coniferin, coniferylaldehyde, spermidine, and 5-O-caffeoylshikimic acid (Fig. 6a). Further pathway integration of these differentially expressed metabolites within the phenylalanine metabolic pathway revealed that phenylalanine, cinnamic acid, and p-coumaric acid, key upstream intermediates, were predominantly accumulated in the S group. Conversely, in the F group and C group, there was a notable enrichment of downstream metabolites such as matairesinol and coniferylaldehyde, indicating enhanced metabolic flux toward phenylpropanoid and flavonoid biosynthesis (Fig. 6b).
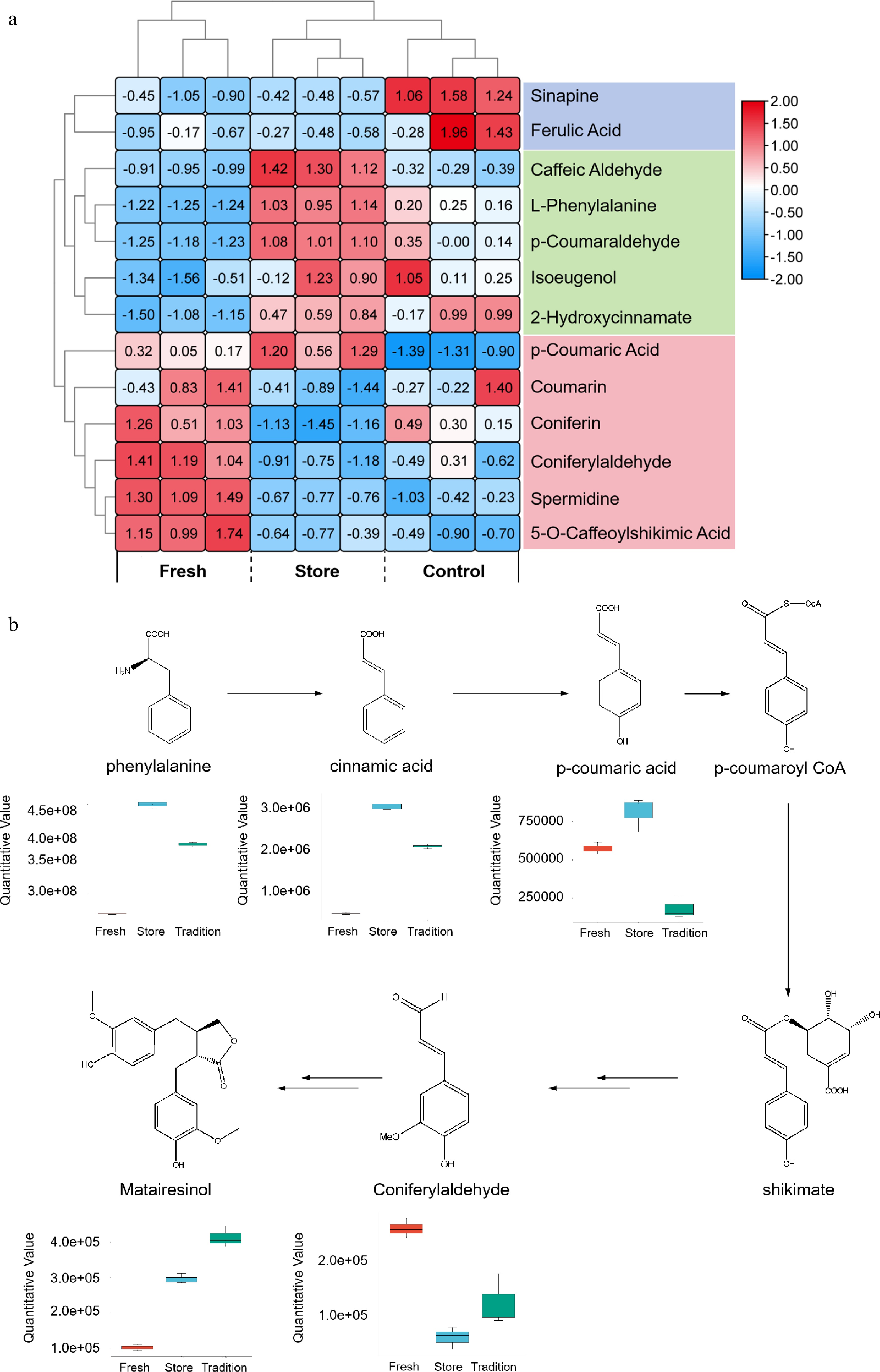
Figure 6.
Various post-harvest processing techniques have a significant impact on the metabolic pathway associated with phenylalanine. (a) Heat maps of the key DMs that are enriched within the phenylalanine-related metabolic pathway. In this representation, each sample is depicted as a column, while each metabolite is represented as a row. The abundance of each metabolite is indicated by a color gradient, with red signifying high abundance and blue indicating low abundance. (b) Schematic representation of the phenylpropanoid metabolic pathway is provided.
The phenylpropanoid pathway generates a wide array of phenolic secondary metabolites, including flavonoids, coumarins, and lignans, which fulfill diverse biological functions such as protection against photo-oxidative damage, regulation of plant development, and defense against pathogens and herbivores[43,44]. The observed enrichment of phenylpropanoid-related metabolites in both the Fresh (F) group and Store (S) group suggests that dehydration and cold storage, respectively, act as abiotic stressors for P. tenuifolia roots. These stress conditions likely stimulate the accumulation of secondary metabolites as part of the plant's adaptive response. A key enzyme in this biosynthetic route is phenylalanine ammonia-lyase (PAL, EC 4.3.1.24), which catalyzes the deamination of phenylalanine to produce trans-cinnamic acid, an essential precursor for downstream phenylpropanoid compounds. PAL is widely recognized as a rate-limiting enzyme in this pathway. Previous studies have shown that cold stress and drought stress can significantly upregulate PAL gene expression in various plant species[45,46]. Based on these findings, it is hypothesized that PtPAL genes are similarly induced during the processing of P. tenuifolia, thereby activating the phenylpropanoid biosynthetic pathway and contributing to the accumulation of stress-responsive secondary metabolites.
-
In this study, three key post-harvest processing methods were systematically applied to freshly harvested P. tenuifolia roots. The present findings demonstrate that fresh-cut processing offers clear advantages over traditional methods, as evidenced by superior levels of three pharmacopoeia-designated marker compounds, higher extract yields, and improved morphological characteristics. It is further recommended that the water content during xylem and phloem separation be carefully controlled within an optimal range, and that storage duration should not exceed 5 months, providing practical guidance for improving production quality and efficiency. Additionally, a quasi-targeted metabolomics analysis across the three processing methods were performed. This analysis revealed that, beyond secondary metabolites, certain primary metabolites may play important roles in the functional properties of P. tenuifolia. These findings offer a novel perspective on post-harvest processing by highlighting the metabolic shifts induced by abiotic stress conditions, such as dehydration and cold storage.
This research was supported by key project at central government level: The Ability Establishment of Sustainable Use for Valuable Chinese Medicine Resources (2060302), Shaanxi Provincial Special Fund for Chinese Medicine (2021-QYZL-01/03), Key Research and Development Program of Shaanxi (2024SF-YBXM-470), General Project of Chinese Medicine Administration of Shaanxi Province (SZY-KJCYC-2023-026), Science and Technology Special Project of the State Administration of Traditional Chinese Medicine (GZY-KJS-2023-036).
-
The authors confirm their contributions to the paper as follows: methodology: Liu S, Zhang J, Gao Y; funding acquisition: Liu Y, Zhang H; project administration: Liu Y; investigation, writing − original draft: Liu S; writing − review and editing: Liang Z, Zhang H; HPLC analysis, resource collection: Liu J; quasi-targeted metabolomics analysis: Liu J; resources, validation: Xu Z; handling of materials: Lei K. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 The information of fresh root of P. tenuifolia.
- Supplementary Table S2 The connection between the relative water content of fresh root and the standards of fresh Root-barks of P. tenuifolia.
- Supplementary Table S3 The content of extracts in C Group and F Group.
- Supplementary Table S4 DM in the F Group vs C Group.
- Supplementary Table S5 The identification of constituents of P. tenuifolia Root-barks.
- Supplementary Table S6 DM in the S Group vs C Group.
- Supplementary Table S7 DM in the S Group vs F Group.
- Supplementary Table S8 An entire list of the p-value and the p-value corrected by the Benjamini procedure of the metabolites.
- Supplementary Table S9 An entire list of enriched KEGG pathway.
- Supplementary Fig. S1 The difficult of Root-bark separated from the roots depended on the relative water content of the fresh roots. X is the time of "sample points". The two point between the gradients yet were belong to different level.
- Supplementary Fig. S2 Appearance characteristics of P. tenuifolia Root-barks with different fresh-processed methods. (a) C Group; (b) F Group; (c) T Group.
- Supplementary Fig. S3 Short-term pri-cold storage at 16 °C would reduce the quality of P. tenuifolia Root-barks to a certain extent.
- Supplementary Fig. S4 The classified of all the 1743 metabolites.
- Supplementary Fig. S5 Total ion chromatograms (TICs) of QC samples in both positive (a) and negative (b) ion modes.
- Supplementary Fig. S6 The PCA plot of all the samples identified.
- Supplementary Fig. S7 Content of Ginsenoside, Ursolic Acid, Notoginsenoside and Raddeanin A in the three processing groups.
- Supplementary Fig. S8 Heatmap cluster of the differential metabolites.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu S, Liu J, Li J, Xu Z, Lei K, et al. 2025. Quasi-targeted metabolomics analysis reveals linkage between chemical composition and different post-harvest processing methods of Polygala tenuifolia. Medicinal Plant Biology 4: e036 doi: 10.48130/mpb-0025-0032 

Quasi-targeted metabolomics analysis reveals linkage between chemical composition and different post-harvest processing methods of Polygala tenuifolia
- Received: 20 May 2025
- Revised: 10 July 2025
- Accepted: 24 July 2025
- Published online: 05 November 2025
Abstract: The dried root bark of Polygala tenuifolia is widely recognized in traditional Chinese medicine (TCM) for its role in promoting cerebral tranquility. However, conventional post-harvest processing methods present challenges in maintaining consistent quality, prompting increased interest in developing improved processing techniques. This study systematically evaluates the impact of three common post-harvest processing methods on the quality of P. tenuifolia. Concentrations of three key bioactive components, tenuifolin (TF), 3,6-disinapoyl sucrose, and polygalaxanthone III, designated as quality markers in the Chinese Pharmacopoeia (2020), were used as analytical indicators. Correlation analysis indicated that separating phloem and xylem water content did not significantly affect the levels of these active compounds in the final product. However, both rehydration and prolonged storage of fresh roots compromised quality: rehydration accelerated the degradation of 3,6-disinapoyl sucrose, while extended storage led to a reduction in TF content. Quasi-targeted metabolomics further revealed significant processing-dependent shifts in the metabolic profile. Fresh-cut samples were enriched in neuroactive primary metabolites such as L-theanine and γ-aminobutyric acid, suggesting a pharmacological basis for the herb's tranquilizing effects. Additionally, the three processing methods differentially affected the biosynthesis of phenylpropanoid compounds: upstream intermediates accumulated in fresh-stored samples, whereas downstream metabolites were more prevalent in samples processed by the other two methods. In conclusion, this study provides a comprehensive comparison of three post-harvest techniques, highlights the advantages of fresh-cut P. tenuifolia, and offers valuable insights for optimizing quality control (QC) and therapeutic efficacy in future research.