









-
The external environmental conditions and internal regulatory factors interactively influence growth and development during the plant life cycle. Elucidating the underlying molecular mechanism of plant growth is necessary to facilitate the utilization of more gene resources in modern breeding engineering. Over the past several decades, numerous pieces of evidence have demonstrated that AINTEGUMENTA-LIKE (AIL) transcription factors (TFs) play a crucial role in regulating plant growth and development processes[1,2]. The AIL genes, which are members of the APETALA 2/ETHYLENE RESPONSE FACTOR (AP2/ERF) superfamily, have been extensively studied in plants[1]. The AIL proteins typically share two AP2/ERF domains connected by a linker region[3]. In Arabidopsis, eight AIL genes, including AINTEGUMENTA (ANT), AIL1, AIL2/BABY BOOM (BBM)/PLETHORA4 (PLT4), AIL3/PLT1, AIL4/PLT2, AIL5/PLT5, AIL6/PLT3, and AIL7/PLT7, have been identified[4−8]. Additionally, the genome-wide identification of the AIL gene family has been conducted in various plant species, including pumpkin, apple, Medicago truncatula, and Brassica species[9−12].
The AIL genes play major roles in embryogenesis, maintenance of meristems, and organ positioning and growth[1,2]. Among them, ANT is specifically required for initiating and maintaining early floral primordia development and regulating organ size[4,5,13]. Loss-of-function of ANT shows smaller organ size, fewer floral organs, abnormal integument, and ovule development, while ectopic expression of ANT causes the formation of enlarged organs[4,5,13−17]. Also, ANT acts together with polarity factor FIL, a YABBY TF, to be involved in organ polarity establishment[18]. During flower development, ANT functions redundantly with AIL6 in regulating floral organ initiation, identity specification, growth, and patterning[19,20]. ANT, AIL6, and AIL7 maintain the cell division and differentiation capacity of the shoot apical meristem; the ant ail6 ail7 triple mutant shoots are prematurely terminated[21]. Moreover, the BBM, AIL4, and AIL6 genes are expressed in the roots and play primary roles in root stem cell niche (SCN) pattern formation, meristem maintenance, and embryogenesis[6−8,22]. The ail3 ail4 ail6 triple mutant exhibits a rootlessness phenotype, and the bbm ail4 double mutant results in embryo lethality[8]. In contrast, overexpression of AIL4 causes ectopic root identity[7]. Overexpression of BBM or AIL5 can induce ectopic embryo formation on meristems, cotyledons, and leaves[6,23−26]. Importantly, BBM also contributes to improving the transformation efficiency[27−29]. In addition, AIL5, AIL6, and AIL7 are also responsible for phyllotaxy stability and lateral root initiation[30−32]. These results suggest that AIL gene functions are diverse but highly conserved in a partially overlapping manner in plant growth and development.
Cucumber, which is consumed as a tender fruit, serves as an important horticultural crop with higher economic value. Given its allogamy characteristic, the cucumber has been used as a model plant for studying sex determination[33−35]. The shape and size of cucumber fruit vary across different geographic populations, but they tend to have a faster growth trait, resulting in a shorter growth cycle[36−38]. Hence, the cucumber also serves as a model species for research on fruit development within the Cucurbitaceae family. Although increasing studies focus on important traits in cucumber, such as sex determination, fruit development, and the brightness of the wart and peel, more work is also required to elucidate the molecular and genetic mechanisms underlying its overall growth and development.
The AIL genes have been extensively characterized in various plant species, especially in Arabidopsis, but their features and functions remain largely unexplored in cucumber. Here, a genome-wide analysis of the CsAIL gene family in cucumber was conducted. Six CsAIL members were identified, and their phylogenetic relationships, gene structures, conserved domains, and motif compositions were systematically characterized. The evolutionary relationships of AIL genes between cucumber and Arabidopsis was also analyzed. Furthermore, the expression profiles of CsAIL genes were well investigated in different tissues and/or the ovaries/fruits at various developmental stages of cucumber. These findings provided important insights into the potential functions of CsAILs during the growth and development processes in cucumber.
-
The cucumber local cultivar 'Xintaimici' and the Nicotiana benthamiana were used in this study. The cucumber plants were cultured in a solar greenhouse, and the Nicotiana benthamiana seedings were grown in a growth chamber at Shandong Agricultural University (Tai'an, Shandong, China) (36°18' N, 117°17' E). Management of cucumber and Nicotiana benthamiana plants were conducted according to standard practices.
Identification and characterization of the CsAIL gene family
-
The eight Arabidopsis AIL proteins were used as queries to identify the CsAIL genes using the BLASTP program in the cucumber genome (
http://cucurbitgenomics.org/v2 ) (accessed on 12 November 2023). The search parameters were set as follows: 'Num Descriptions = 5', 'Num Alignments = 5', and 'e-Value (Expected Threshold) = 1e-2'. Furthermore, all predicted CsAIL sequences were manually confirmed by examining the conserved AP2 domain using Conserved Domain Database (CDD)[39] (www.ncbi.nlm.nih.govStructure/cdd/wrpsb.cgi ), and SMART[40] (http://smart.embl.de ). Chromosomal locations of the CsAIL genes were retrieved from the cucumber genome. Physicochemical properties of CsAIL proteins, including molecular weights (MW) and isoelectric points (pI), were predicted via the ProtParam tool on the ExPASy server[41] (https://web.expasy.org/protparam ). The eight Arabidopsis AIL protein data were obtained from TAIR (www.arabidopsis.org ) (accessed on 12 November 2023).Phylogenetic and synteny analyses of CsAIL genes
-
The protein sequences of CsAILs and Arabidopsis AILs were aligned by ClustalW. A phylogenetic tree was constructed from the multiple sequence alignment with the MEGA 11 software under the Neighbor-Joining method, applying the following parameters: 1,000 bootstrap replicates, Poisson model, and complete deletion. The tree was visualized using the Interactive Tree Of Life (iTOL) (
https://itol.embl.de ) (accessed on 12 November 2023). The synteny analysis was conducted using the Multiple Collinearity Scan toolkit (MCScanX)[42].Analysis of gene structures, conserved motifs, and protein domains
-
Gene structures of CsAILs were visualized by the Gene Structure Display Server[43] (
http://gsds.gao-lab.org ). Conserved motifs of CsAIL proteins were predicted via the MEME online tool[44] (https://meme-suite.org/meme/tools/meme ). Following the multiple sequence alignment of CsAIL proteins, the conserved domains were highlighted by Jalview[45] (www.jalview.org ).Transcriptome analysis of CsAIL genes in cucumber
-
The cucumber transcriptome data analyzed in this study were obtained from the RNA-Seq projects in the Cucurbit Genomics Database (
http://cucurbitgenomics.org/cgi-bin/RNA_project ) (accessed on 15 November 2023). The datasets included:PRJNA80169: roots, stems, leaves, male flowers, female flowers, ovaries, and tendrils of Cucumis sativus var. sativus line 9,930.
PRJNA258122: ovaries of about 2−4 cm in length from cucumber hermaphrodite line 408 (long fruit), and its near-isogenic line 409 (short fruit).
PRJNA431715: ovaries at 6 d before flowering (DBF), and pericarps at 0 DBF from cucumber inbred lines NCG122 (numerous fruit spines), and NCG121 (few fruit spines).
PRJNA319011: seeds at 28, 37, 45, and 60 d after pollination (DAP).
PRJNA271595: growing tips of seedling primary roots of Cucumis sativus Gy14, including meristematic zone (MZ), differentiation zone (DZ), and elongation zone (EZ).
PRJNA437579: root apices (0–5 mm from the tips) of hydroponically grown seedlings (5-day-old) of cucumber cv. Jinchun No. 2 under control conditions and 75 mM NaCl stress for 24 h.
Expression patterns of CsAIL genes were analyzed using the above transcriptome data, with significance thresholds set at false discovery rate (FDR)-adjusted p < 0.05 and fold change (FC) ≥ 1.5. Values represent the means ± standard deviation (SD).
RT-qPCR analysis
-
Samples of ovaries or fruits at different developmental stages were taken into liquid nitrogen and then pulverized. The total RNA was extracted by the Quick RNA Isolation Kit (Huayueyang, Beijing, China), and the cDNA was synthesized from the extracted RNA template using the FastKing gDNA Dispelling RT SuperMix (TianGen, Beijing, China). RT-qPCR was carried out on an Applied Biosystems 7500 real-time PCR system (Applied Biosystems, USA) with the UltraSYBR Mixture (Cwbio, Beijing, China). The UBIQUITIN (UBI) gene served as the internal control. Three biological replicates (each consisting of three technical replicates) were performed for these experiments. Values were the means ± SD. The primers are listed in Supplementary Table S1.
Subcellular localization
-
The CDS of CsAIL1 was cloned and constructed into the Super1300: CsAIL1-EGFP vector. The Super1300-EGFP and Super1300: CsAIL1-EGFP vectors were introduced into Arobacterium tumefaciens GV3101 (pSoup-P19), respectively, and subsequently were co-expressed with the cell nucleus marker NF-YA4-mCherry in young Nicotiana benthamiana leaves. The GFP signals (at 488 nm) and mCherry signals (at 561 nm) were detected using a Nikon A1 laser confocal scanning microscope and captured by the NIS-Elements software (Supplementary Table S2).
Prediction of protein interaction network
-
The CsAIL1 protein sequence was analyzed using the STRING database (
https://cn.string-db.org ) to predict the potential interaction network. Active interaction data were integrated from multiple sources, including curated databases, experimental determination, gene co-occurrence, text mining, and protein homology. -
The Arabidopsis eight AIL proteins were employed as queries to identify the CsAIL homologs in the cucumber genome using the BLASTP program. In total, six CsAIL genes, including CsAIL1, CsBBM, CsAIL3, CsAIL5, CsAIL6, and CsANT were obtained in cucumber after further confirmation of the highly conserved AP2 domain via SMART and CDD analysis (Table 1). The CsAIL genes were named according to the highest protein sequence identification values to the closest Arabidopsis homologs (Table 1, Fig. 1). These six CsAIL genes are distributed across five cucumber chromosomes (chrs) except for chr 5 and chr 7. The sequences of CsAIL proteins ranged from 466 to 650 amino acids (AA), accompanied by predicted molecular weights (MW) that varied from 51.43 to 70.01 kDa. Moreover, the predicted isoelectric point (pI) values ranged from 5.88 to 8.64; notably, CsAIL5 displayed a basic pI (8.64) which differed from the acidic or near-neutral values of other members (Table 1).
Table 1. The classification of CsAIL gene family in cucumber.
Gene name Gene ID Chromosome location Amino acid length (AA) MW (kDa) pI Arabidopsis ortholog CsAIL1 CsaV3_3G031350 chr3: 27057687...27061567 (−) 533 59.07 5.88 AIL1 CsBBM CsaV3_2G010320 chr2: 7285623...7289916 (+) 650 70.01 6.23 BBM (AIL2, PLT4) CsAIL3 CsaV3_4G024120 chr4: 14070320...14073714 (−) 615 67.27 6.34 AIL3 (PLT1)/AIL4 (PLT2) CsAIL5 CsaV3_1G031650 chr1: 18716067...18719027 (+) 466 51.43 8.64 AIL5 (PLT5) CsAIL6 CsaV3_3G006900 chr3: 6134843...6139602 (−) 517 57.00 6.28 AIL6 (PLT3)/AIL7 (PLT7) CsANT CsaV3_6G044340 chr6:26200005...26204175 (+) 629 69.60 7.08 ANT 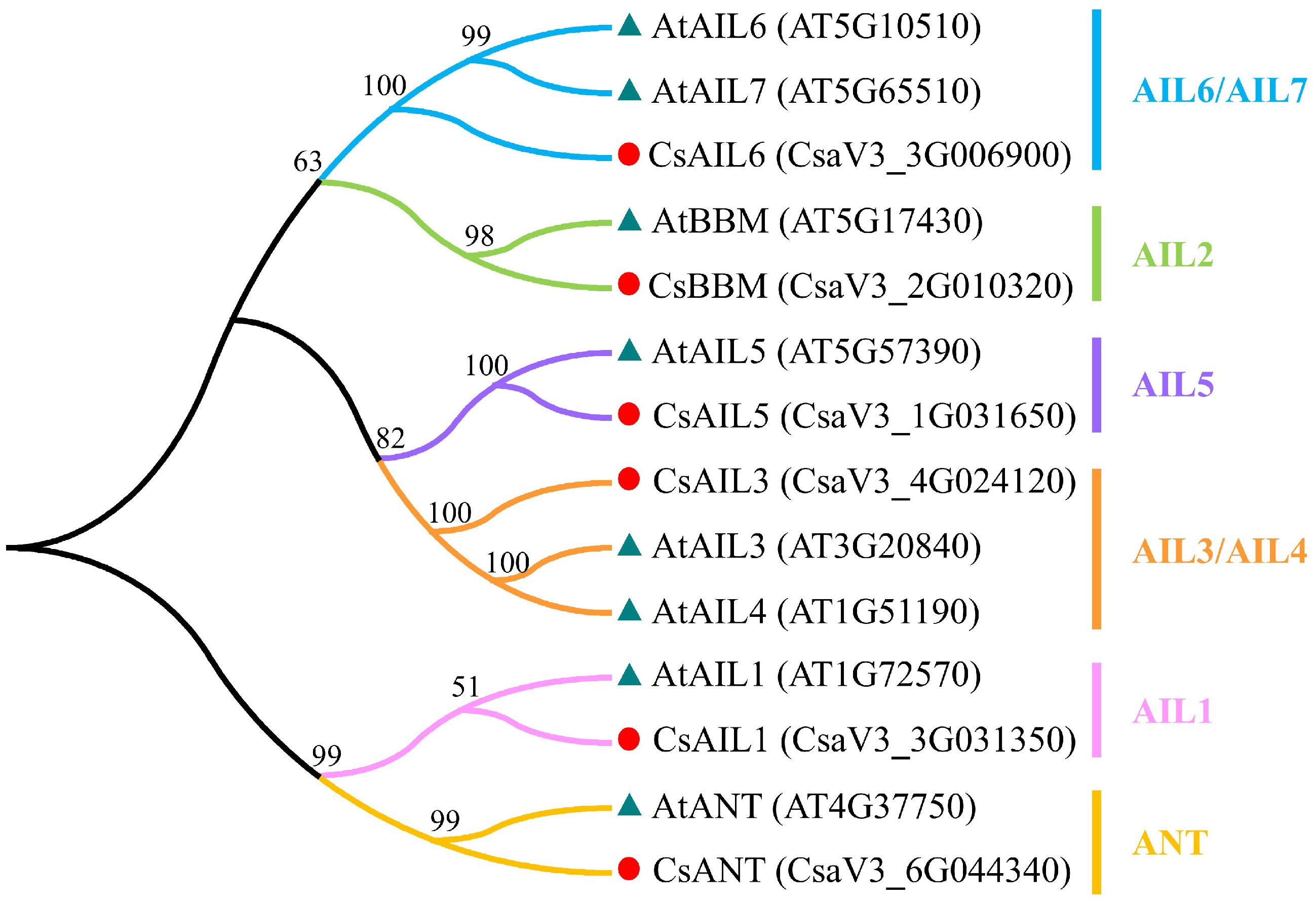
Figure 1.
Phylogenetic analysis of the AIL proteins between cucumber and Arabidopsis. Red circle and blue-grey triangle represent the AIL proteins from cucumber and Arabidopsis, respectively. Cs, cucumber; At, Arabidopsis.
Phylogenetic and synteny analyses of the CsAIL genes
-
To elucidate evolutionary relationships among CsAIL genes, full-length protein sequences were recruited to construct a phylogenetic tree between cucumber and Arabidopsis using MEGA 11 software. Similar to Arabidopsis, the six CsAILs were divided into six subclades, including AIL1, AIL2, AIL3/AIL4, AIL5, AIL6/AIL7, and ANT, in which each subclade has only one CsAIL gene (Fig. 1). Notably, CsAIL3 was closest to AIL3 and AIL4 in Arabidopsis, and CsAIL6 showed the closest relationship with AIL6 and AIL7 in Arabidopsis (Fig. 1). These results suggest that the CsAIL genes of cucumber possibly share conserved evolutionary relationships with Arabidopsis AIL genes.
Segment and tandem duplications are key mechanisms reflecting plant genome evolution and facilitating gene family expansion[46]. Here, a synteny analysis of the AIL gene family between cucumber and Arabidopsis was conducted. Four CsAIL genes (CsAIL1, CsAIL3, CsAIL5, and CsAIL6) have syntenic relationships with Arabidopsis AIL genes. Among them, CsAIL5 and CsANT had one syntenic gene pair, respectively (Fig. 2). CsAIL3 and CsAIL6 showed two syntenic gene pairs, respectively (Fig. 2), suggesting an absence of any tandem/segment duplication of the two genes in cucumber compared to Arabidopsis. However, no synteny relationship was present between CsAIL1 and AtAIL1, or CsBBM and AtBBM, respectively (Fig. 2), indicating that these two genes between cucumber and Arabidopsis may not undergo the segment and/or tandem duplication events.
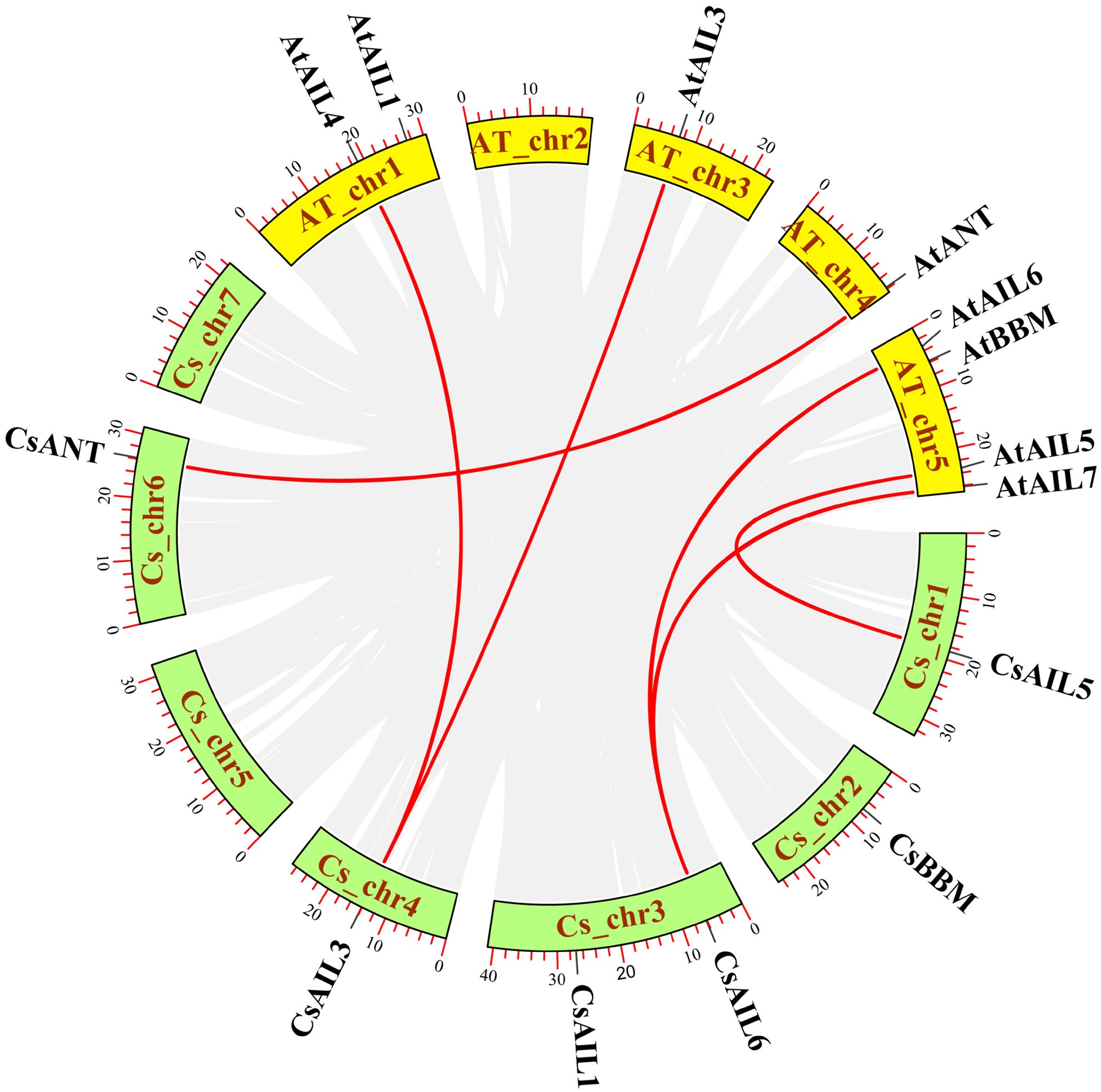
Figure 2.
Synteny analysis of AIL genes between cucumber and Arabidopsis. The synteny gene pairs are highlighted in the red lines. Cs, cucumber; At, Arabidopsis.
Analysis of gene structures and conserved motifs/domains in the CsAIL genes
-
Gene structure diversity and conserved motifs/domains are related to the evolutionary history of a gene family and also reflect the conservation or divergence of gene function to some extent[47]. A comprehensive analysis, which consists of an unrooted evolutionary tree, gene structure characterized by exon-intron distribution, and conserved motifs were conducted. The evolutionary analysis divided CsAILs into three subgroups: (i) CsBBM and CsAIL6; (ii) CsAIL3 and CsAIL5; and (iii) CsAIL1 and CsANT (Fig. 3a), but the genes in same subgroup (e.g., i and ii) did not have a similar exon/intron organization pattern, while the genes in subgroup (iii) showed the same distribution of nine exons and eight introns, respectively (Fig. 3b). Totally, all CsAIL genes displayed differentiation with 7–9 exons and 6–8 introns (Fig. 3b), which is in line with other plant species AIL genes, implying that the cucumber CsAIL gene structures are probably conserved during evolution. Furthermore, the conserved motifs of CsAILs were predicted by MEME analysis. It was observed that most CsAILs share the same motif composition, except that CsAIL5 lacks motif 8 (Fig. 3c, Supplementary Fig. S1). Meanwhile, the multiple sequence alignment results showed that all CsAIL proteins possess two highly conserved AP2 domains (designated AP2-R1 and AP2-R2), which are separated by a conserved linker region (Fig. 4a). Each AP2 domain is composed of YRG and RAYD (which contains an amphipathic α-helix) elements. No significant differences in amino acid residues of AP2 domains existed among all CsAIL proteins (Fig. 4b). Collectively, these results suggest that the CsAIL genes may have conserved functions during the evolution process in cucumber.
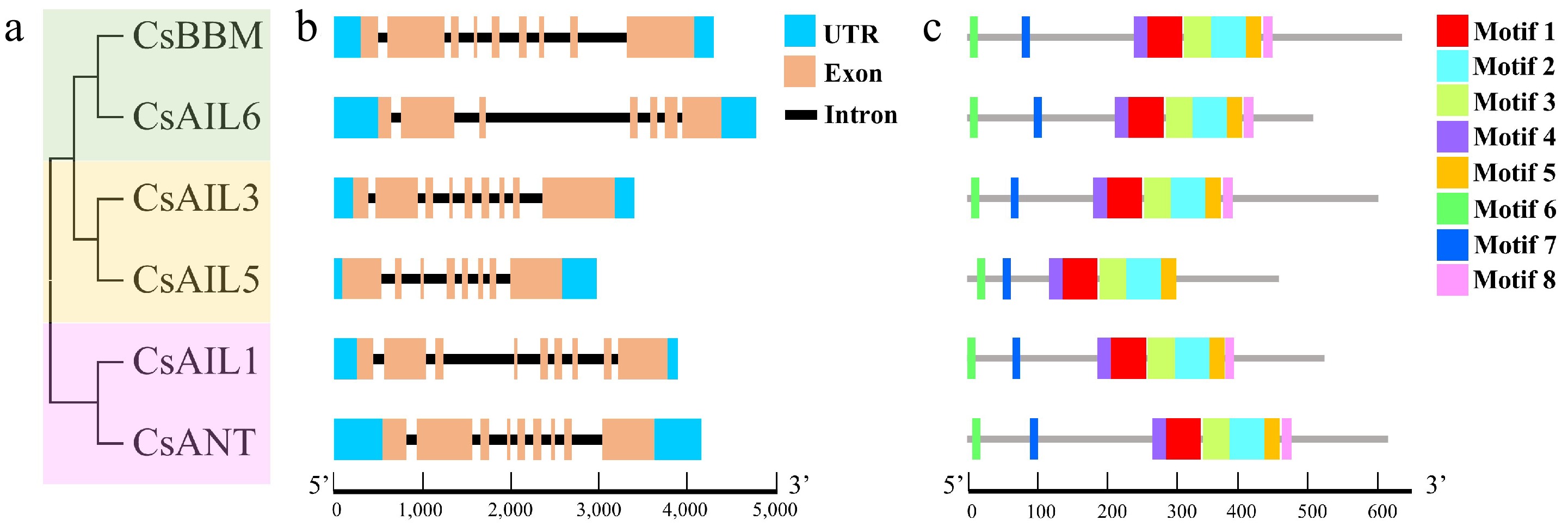
Figure 3.
Phylogenetic clustering, gene structures, and conserved motifs of CsAIL genes. (a) Unrooted phylogenetic tree of CsAIL proteins. (b) Gene structures of CsAIL genes. Untranslated regions (UTR), exons, and introns are indicated by blue boxes, light-brown boxes, and black lines, respectively. (c) Conserved motifs of CsAIL proteins. Distinct motifs are represented by different colored boxes.
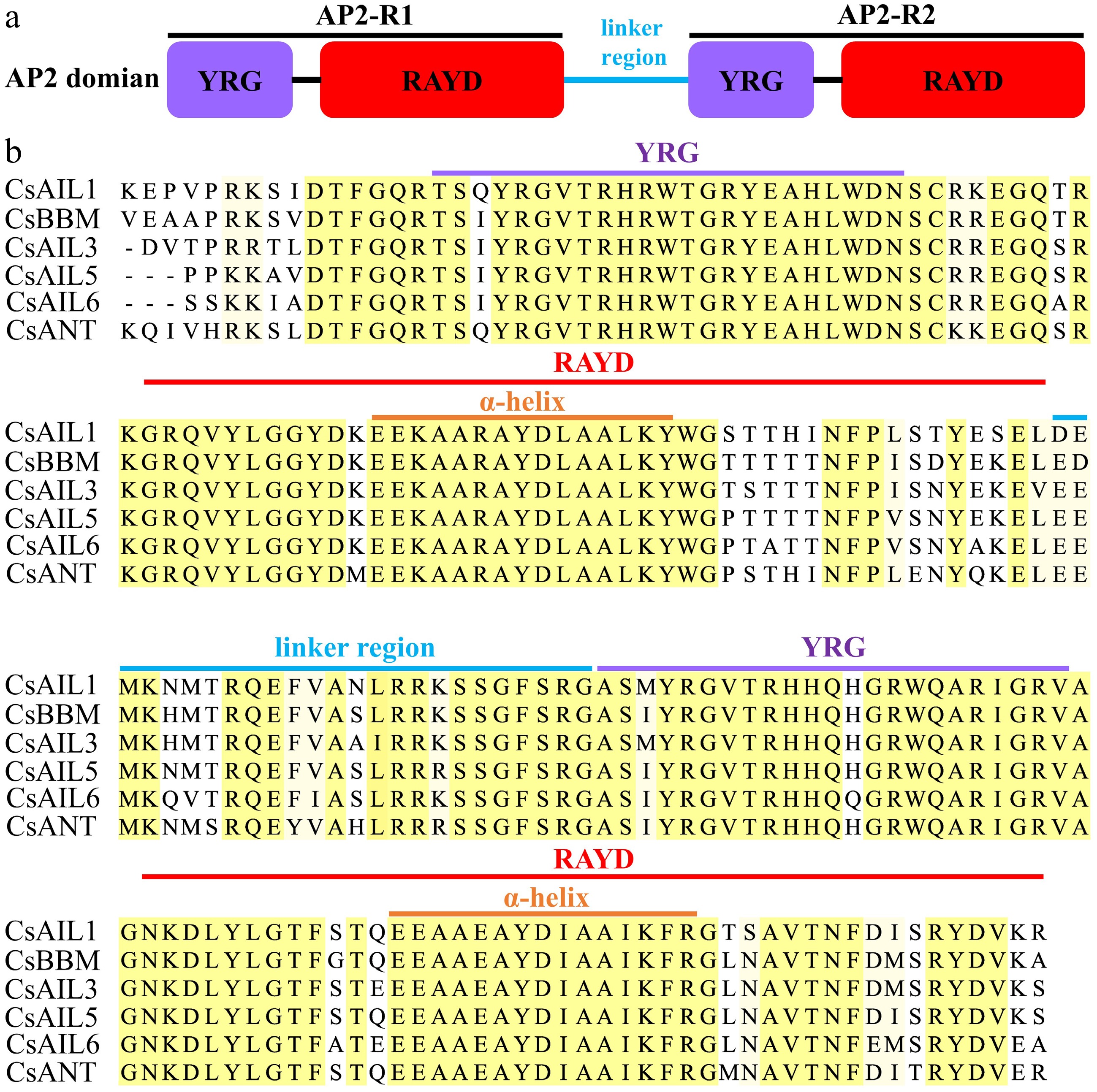
Figure 4.
The conserved AP2 domain of CsAIL proteins. (a) Model of AP2 domain. (b) Typical amino acid residues within AP2 domain.
Expression profiles of CsAILs in cucumber
-
To understand the potential functions of CsAILs during cucumber growth and development, their expression patterns were analyzed across various tissues of cucumber, including roots, stems, leaves, tendrils, male and female flowers, and ovaries. The CsAIL genes exhibited different expression patterns across the examined tissues. As shown in Fig. 5, CsBBM, CsAIL3, and CsAIL6 were specifically expressed in the roots, and no expression was detected in other tissues. CsAIL5 and CsANT were also expressed in the roots. Moreover, CsAIL1, CsAIL5, and CsANT showed higher expression levels in the leaves and ovaries. Meanwhile, CsAIL1 expression was also detected in the tendrils, while CsAIL5 and CsANT were also expressed in the stems. These results imply that CsAIL genes may be involved in the regulation of diverse developmental processes in cucumber.
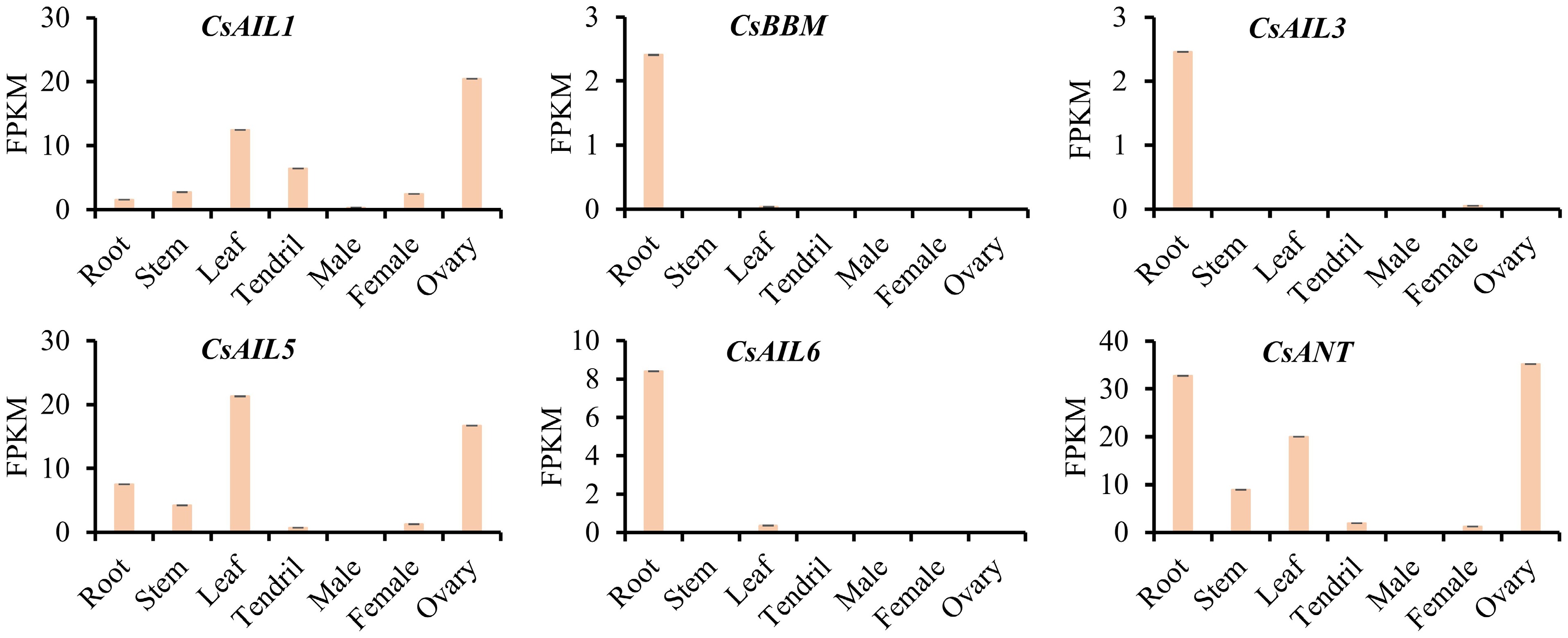
Figure 5.
Expression patterns (FPKM) of CsAIL genes in different tissues of cucumber. The expression data were derived from the PRJNA80169 transcriptome dataset.
CsAIL genes may participate in the development of ovaries, fruit spines, and seeds
-
To further explore the potential roles of CsAIL genes during cucumber ovary/fruit development, their expression levels in two cucumber varieties that have differences in fruit length (408 has a long fruit phenotype, 409 exhibits a short fruit phenotype) were analyzed. Only CsAIL1, CsAIL5, and CsANT were expressed in the ovaries of 408 and 409 (Fig. 6a), and this result is in line with that in Fig. 5. In addition, the three genes showed lower expression levels in 409 than in 408 (Fig. 6a), hinting at their potential roles in fruit elongation. Furthermore, we examined the expression patterns of CsAIL1, CsAIL5, and CsANT in cucumber ovaries/fruits at different developmental stages. As shown in Fig. 6b–d, the three genes showed consistently high expression levels in the ovaries at various developmental stages, but not in the fruits. Given massive cell proliferation at the ovary stage of cucumber[48,49], these results indicate that CsAIL1, CsAIL5, and CsANT possibly contribute to fruit development via cell proliferation.
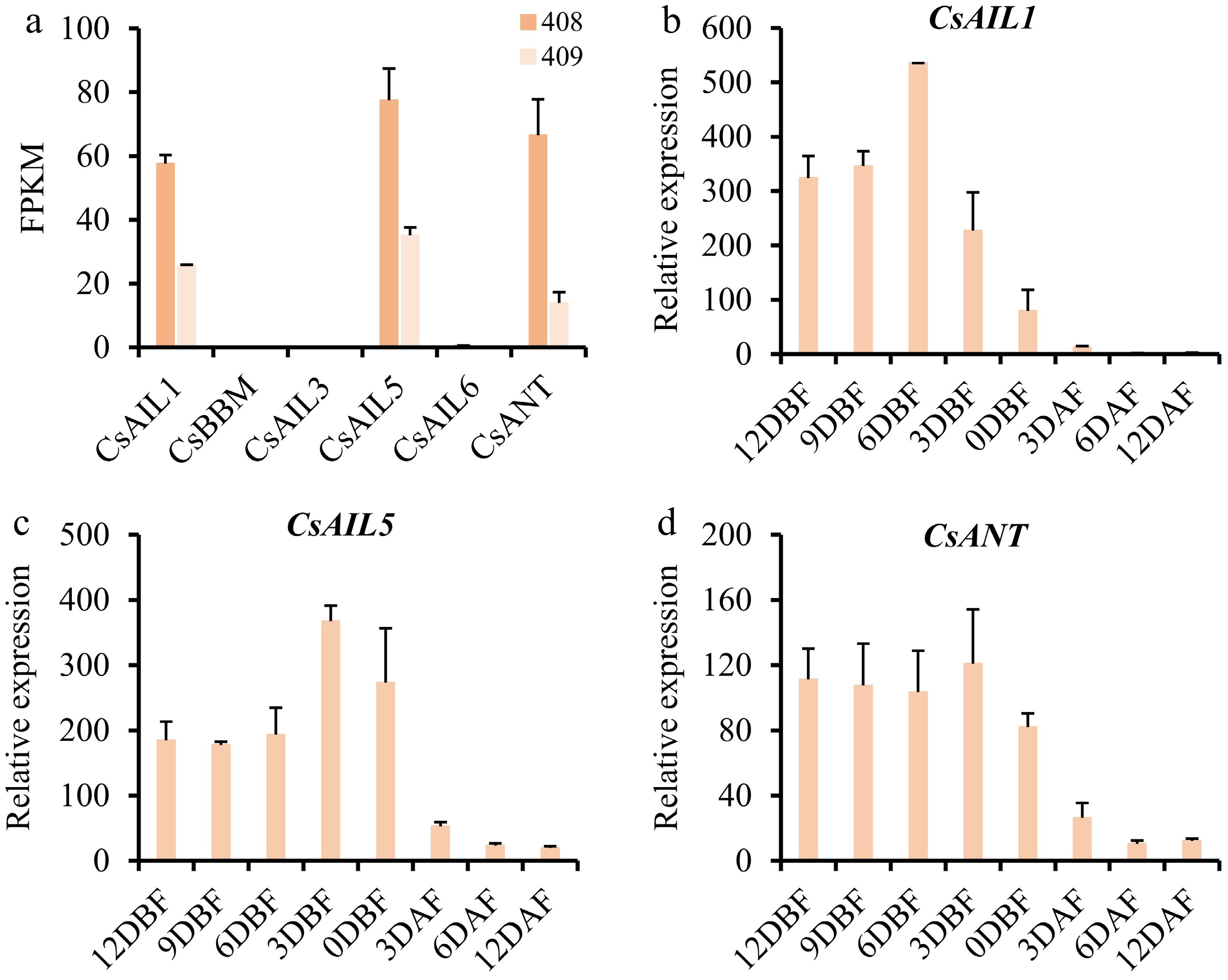
Figure 6.
Expression profiles of CsAIL genes in different stages of ovaries and fruits in cucumber. (a) Expression abundance (FPKM) of CsAILs in the ovaries from long-fruit (408), and short-fruit (409) cucumber lines. Expression data were obtained from the PRJNA258122 transcriptome dataset. (b)–(d) Relative expression levels of CsAIL1, CsAIL5, and CsANT at different developmental stages of ovaries and fruits as determined by RT-qPCR. Error bars represent the SD of three biological replicates. DBF, days before flowering; DAF, days after flowering.
In the two cucumber varieties, NCG121 and NCG122, NCG121 has few fruit spines, while NCG122 fruit harbors numerous fruit spines. We noted that CsAIL5 and CsANT exhibited higher and non-differential expression levels in the ovaries of NCG121 and NCG122, both at the 6 DBF and 0 DBF stages (Fig. 7a). While CsAIL1 was more highly expressed in the ovaries than CsAIL5 and CsANT at 6 DBF stage (Fig. 7a). Interestingly, CsAIL1 expression was also significantly higher in NCG121 ovaries at 6 DBF stage compared to NCG122 (Fig. 7a), implying that CsAIL1 may negatively regulate fruit spine initiation or development in cucumber.
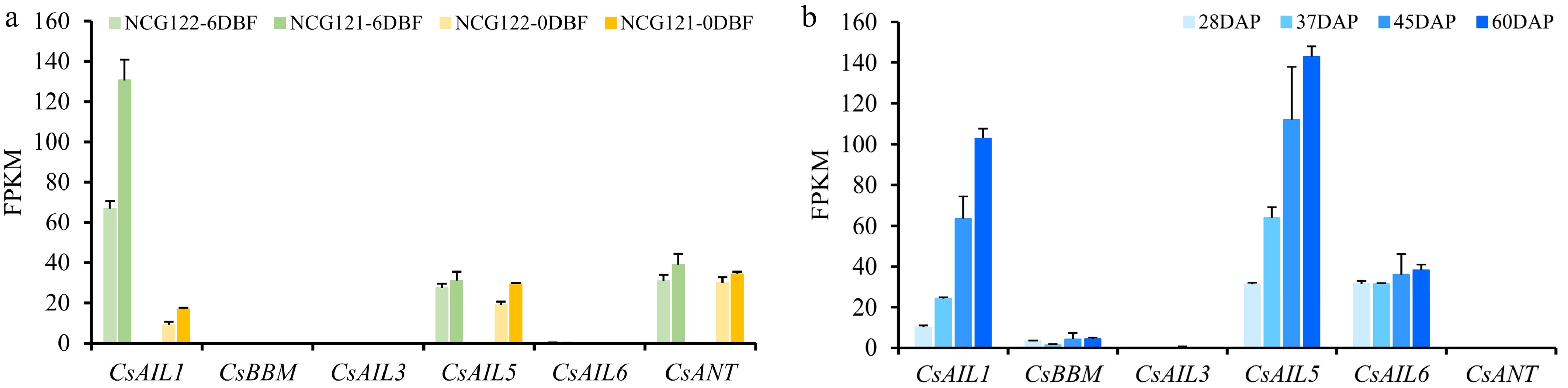
Figure 7.
Expression profiles of CsAIL genes in cucumber ovaries and seeds across developmental stages. (a) Expression levels (FPKM) of CsAILs in ovaries of the NCG121 (few fruit spines), and NCG122 (numerous fruit spines) varieties at different developmental stages. Expression data were obtained from the PRJNA431715 transcriptome dataset. (b) Expression patterns (FPKM) of CsAILs during seed development. Expression data from the PRJNA319011 transcriptome dataset were used in this analysis. Error bars indicate the SD of three biological replicates. DAP, days after pollination.
In plants, AIL genes have been well documented to play crucial roles in embryogenesis[6−8]. Here, the CsAIL gene expression patterns were examined during different stages of seed development (28, 37, 45, and 60 DAP). At the four tested stages, the expressions of CsAIL1 and CsAIL5 progressively increased along with seed development; CsAIL6 exhibited sustained expression, and CsBBM showed a slight expression level. In contrast, no expression of CsAIL3 and CsANT was detected (Fig. 7b). These data indicate that CsAIL1, CsAIL5, and CsAIL6 may participate in controlling embryogenesis during seed development.
Involvement of CsAILs in root development and salt stress response
-
Previous studies showed that AIL genes control root development and rhizotaxis formation[7,8]. Therefore, the expression patterns of CsAIL genes in distinct root zones of cucumber, including the differentiation zone (DZ), the elongation zone (EZ), and the meristematic zone (MZ) were explored. CsBBM, CsAIL3, and CsAIL6 were highly expressed in MZ, and CsAIL6 also exhibited higher expression levels in DZ and EZ (Fig. 8a). These results suggest that CsBBM, CsAIL3, and CsAIL6 may be responsible for meristem maintenance in the roots. Accordingly, CsBBM and CsAIL3 were sensitive to salt stress, as evidenced by their decreased expression levels following salt treatment (Fig. 8b), implying potential roles for these genes in abiotic stress, especially in salt stress. However, CsAIL6 was insensitive to salt stress, as no expression difference of CsAIL6 was found between salt-treated and control plants (Fig. 8b).
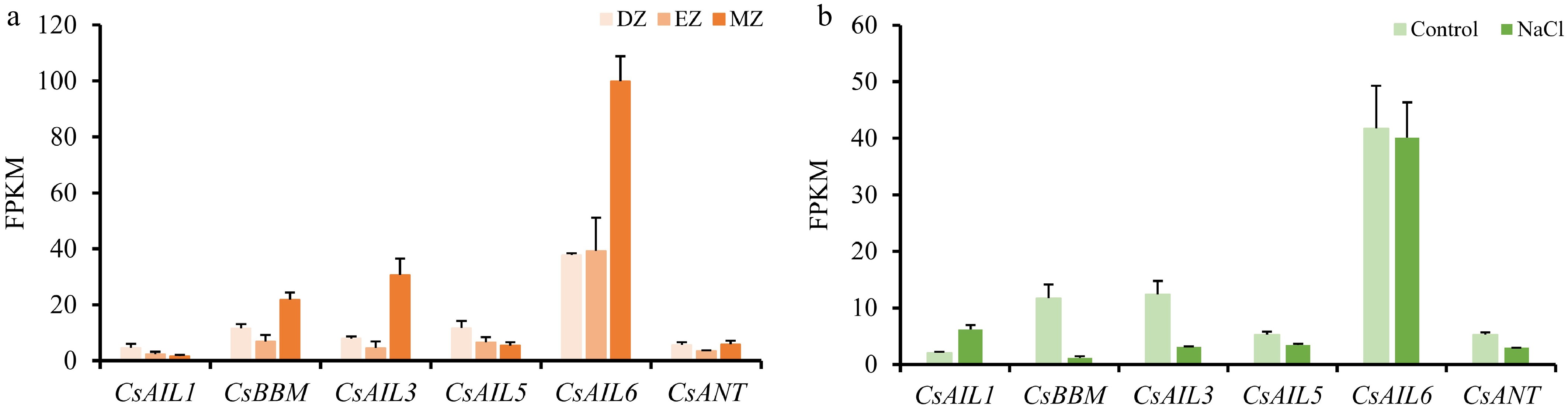
Figure 8.
Expression patterns of CsAIL genes in cucumber roots and under salt stress. (a) Expressions (FPKM) of CsAILs in different root developmental regions. Data were derived from the PRJNA271595 transcriptome dataset. DZ, differentiation zone; EZ, elongation zone; MZ, meristematic zone. (b) Expression levels (FPKM) of CsAILs in roots under control conditions and NaCl treatment. Data from the PRJNA437579 transcriptome dataset were used in this analysis. Error bars represent the SD of three biological replicates.
Subcellular localization and predicted interaction network of CsAIL1 protein
-
Given the potential roles of CsAIL1 with its higher expression level in the ovaries, fruit spines, and seeds, the localization of CsAIL1 in cells and the potential interaction network were further explored. When CsAIL1 was expressed in N. benthamiana leaves, the CsAIL1-GFP fusion protein localized exclusively to the cell nucleus (Fig. 9a). To further understand how CsAIL1 functions during plant growth and development, the possible interaction factors of CsAIL1 were predicted. Among all predicted interaction factors, it was found that CsAIL1 potentially interacts with an ethylene response factor, CsERF13, for which there are no reports in fruit development (Fig. 9b). The CsAIL1-CsERF13 module may provide a new avenue for research into fruit development.
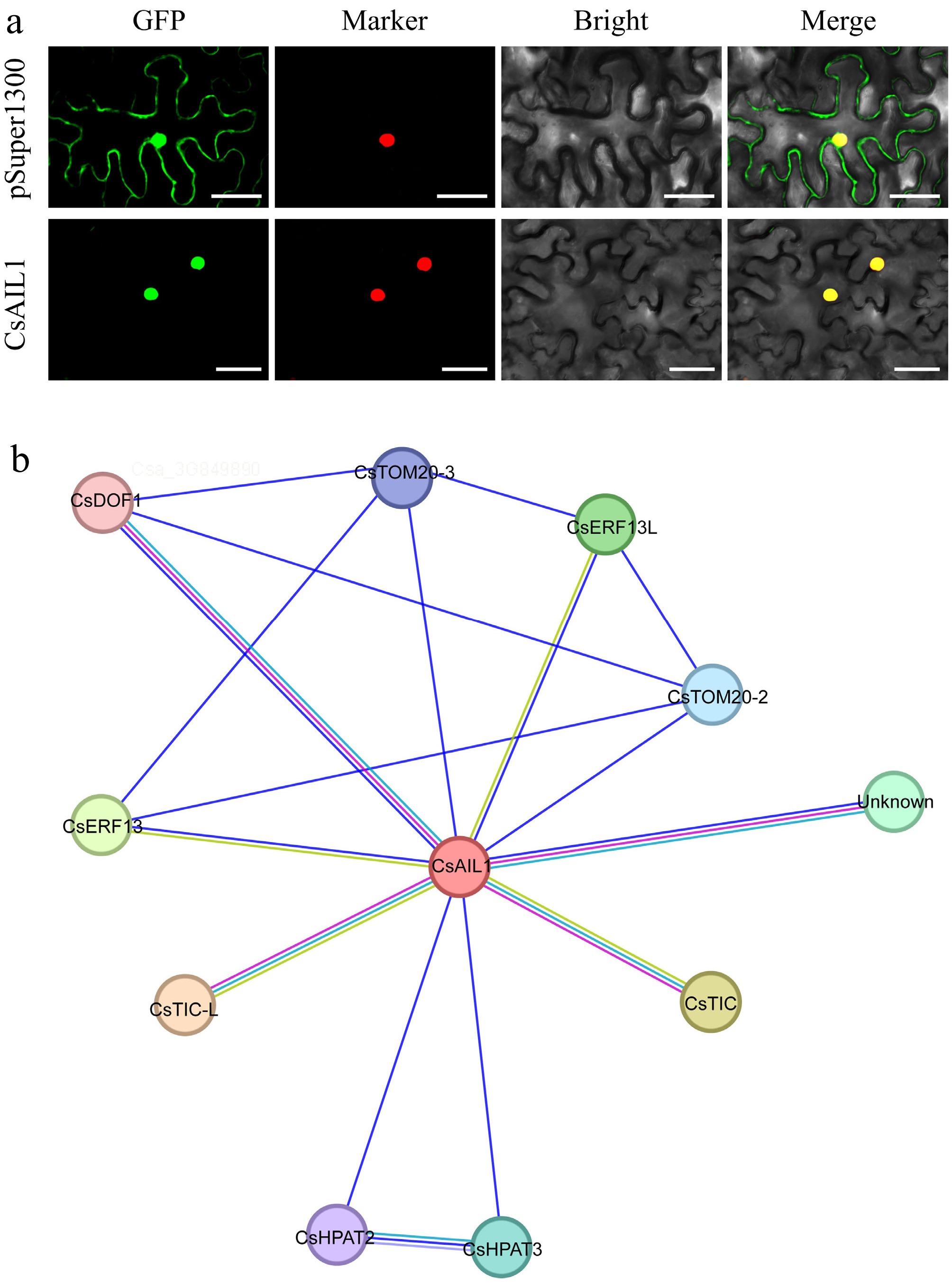
Figure 9.
Subcellular localization and interaction network of CsAIL1 protein. (a) Subcellular localization of CsAIL1 in epidermal cells of N. benthamiana leaves. Bars = 50 μm. (b) Predicted interaction network of CsAIL1 in cucumber. Data sources of predicted interactions are color-coded as follows: purple line, experimentally determined; pearl blue line, from curated databases; blue line, gene co-occurrence; light green line, textmining; light blue-violet line, protein homology.
-
The master regulatory factors, AIL proteins, are involved in multiple developmental processes throughout the plant life cycle[1,2]. However, the functions of CsAILs in the important Cucurbitaceae crop, cucumber, have not been well studied. Herein, six CsAIL members were identified in the cucumber genome; however, the total number of CsAIL genes is fewer than those in Arabidopsis (Table 1). Since gene duplication can contribute to gene family expansion[46], it was speculated that the fewer gene duplication events of CsAIL3 and CsAIL6 might be responsible for the reduced number of CsAIL genes in cucumber compared to Arabidopsis. Furthermore, the synteny analysis results showed that no gene duplication events are observed between CsAIL1 and Arabidopsis AIL1, and between CsBBM and Arabidopsis BBM, respectively (Fig. 2), hinting that the AIL1 or BBM gene is not derived from a common ancestor between cucumber and Arabidopsis, although they are in the same corresponding phylogenetic clade, respectively (Fig. 1). The conserved domains and motifs, which are involved in protein interactions, DNA binding and transcription activity[50], mainly determine the function of the gene. It was found that all CsAIL proteins consist of two highly conserved AP2 domains connected by a linker region (Fig. 4), and share eight conserved motifs except CsAIL5, which lacks motif 8 (Fig. 3). The highly conserved motifs and AP2 domains suggested that the CsAILs might have conserved functions during cucumber growth and development.
To investigate the possible functions of CsAIL genes, the expression patterns of these genes were examined across multiple cucumber tissues. Among the six CsAIL genes, CsAIL1, CsAIL5, and CsANT showed higher expression levels in the ovaries and leaves of cucumber (Fig. 5). The expression pattern of CsANT aligns well with the function of the ANT gene in other plants, suggesting a potentially conserved function of the ANT gene[4,5,13,14,16]. However, no evidence describes the function of the AIL1 gene in plants and the function of AIL5 in ovary or fruit development[1]. Thus, these results suggested that CsAIL1, CsAIL5, and CsANT might control the phyllotaxy and ovary development in cucumber. Moreover, the expression levels of CsAIL1, CsAIL5, and CsANT in the ovaries of 408, which harbors short fruits, are lower than those in 409, which shares long fruits (Fig. 6a). In addition, during the different developmental stages of ovary and fruit, the three genes are mainly expressed in the ovary developmental stage but not in the fruit developmental stage (Fig. 6b–d). Cell proliferation primarily occurred at the ovary developmental stage, and cell expansion appeared during the fruit developmental stage in cucumber[48,49]. Hence, these data further confirmed the potential function of CsAIL1, CsAIL5, and CsANT in cell proliferation during the ovary growth and developmental stage in cucumber.
Numerous pieces of evidence have confirmed that BBM, AIL3, AIL4, and AIL5 play essential roles in embryogenesis[6−8]. However, no expression of CsAIL3 and a relatively lower expression of CsBBM were detected during the seed developmental stages after pollination (Fig. 7b). Previous studies have demonstrated that AIL4 and BBM have shared roles in early embryo development[8,25]. Given that the expression analysis was based on the transcriptome data of seeds at 28 DAP (not the early embryo stage) in this study, and CsAIL3 in cucumber is homologous to AIL3 and AIL4 in Arabidopsis (Table 1 and Fig. 1), it was thus speculated that CsAIL3 and CsBBM also might regulate early embryo development in cucumber, but require further functional verification. In addition, the expression levels of CsAIL1 and CsAIL5 gradually increased except for CsAIL6, with a higher and stable expression level (Fig. 7b), hinting that the CsAIL1, CsAIL5, and CsAIL6 possibly fulfil the predominant roles in the cucumber seed later developmental stage. Interestingly, the CsAIL1 gene also exhibited higher expression in the ovaries at 6 DBF stage in the NCG122 cucumber variety, which has numerous fruit spines, compared to the NCG121 variety, which has fewer fruit spines (Fig. 7a). The fruit spine (also known as trichome) can influence the appearance quality of cucumber fruit and is a fundamental commercial trait in cucumber breeding[51]. Hence, the CsAIL1 might be a promising candidate gene for investigating fruit spine development in cucumber.
Similar to the functions of BBM, AIL3, AIL4, and AIL6 in root development[7,8], their homologs also exhibited specific expression patterns in the roots of cucumber (Fig. 5). Further analysis revealed that CsBBM, CsAIL3, and CsAIL6 are highly expressed in the DZ, EZ, and MZ, especially in the MZ (Fig. 8a), suggesting their potential roles in cell proliferation of the roots. Whereas CsANT had a lower expression level in the three zones (Fig. 8a), hinting at a non-vital function of CsANT in root development of cucumber. Previous studies have suggested that ANT is involved in response to abiotic stress[52,53]. However, in this study, CsBBM and CsAIL3, but not CsANT, were found to be salt-responsive genes, with their transcript levels decreasing under salt stress. In contrast, CsAIL6 maintained expression levels comparable to the control despite showing higher expression than CsBBM and CsAIL3 following salt treatment (Fig. 8b). Thus, these results provided valuable insights into the potential applications of CsAIL genes for enhancing salt tolerance in cucumber breeding.
-
In this study, a genome-wide characterization of the CsAIL gene family in cucumber was conducted. Six CsAIL genes were identified and their gene structures, conserved motifs and domains, evolutionary relationships, and gene duplication events were described in detail. Expression profiles of the CsAIL genes in cucumber were similar to those of their orthologs in other plants, indicating their potentially conserved functions, including leaf development, ovary and fruit development, root development, embryogenesis, and salt stress response. Importantly, CsAIL1 was identified as a candidate regulator of ovary, fruit spine, and seed development in cucumber. This speculation diverges from other plants and requires further functional verification. Moreover, CsAIL genes exhibited distinct expression profiles under salt stress, implicating their potential involvement in abiotic stress responses. Subcellular localization confirmed the nuclear presence of CsAIL1 protein, and interaction network prediction suggested a potential interaction with the ethylene response factor CsERF13. Altogether, these findings lay a promising foundation for future functional studies and targeted breeding strategies, potentially enabling the fine-tuning of cucumber development and stress resilience through CsAIL-mediated regulatory pathways.
This work was supported by the National Natural Science Foundation of China (32302572), and the Central Guiding Local Science and Technology Development Funds of Shandong Province (YDZX2023071).
-
The authors confirm their contributions to the paper as follows: conceived, designed, wrote and revised the manuscript: Yin S, Zhang Y; performed the experiments: Yin S, Chen C; analyzed the data: Yin S, Liu J, Lou A. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed in the current study are available from the corresponding author on reasonable request.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in RT-qPCR.
- Supplementary Table S2 Primers of CsAIL1 used in subcellular localization.
- Supplementary Fig. S1 Conserved motifs in CsAIL proteins in cucumber.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yin S, Chen C, Liu J, Lou A, Zhang Y. 2025. Genome-wide identification and expression analysis of CsAIL gene family in cucumber. Vegetable Research 5: e044 doi: 10.48130/vegres-0025-0044 

Genome-wide identification and expression analysis of CsAIL gene family in cucumber
- Received: 25 August 2025
- Revised: 11 October 2025
- Accepted: 21 October 2025
- Published online: 19 November 2025
Abstract: AINTEGUMENTA-LIKE (AIL) genes encode key transcriptional regulators governing plant growth. However, little is known of CsAIL genes in cucumber (Cucumis sativus L.). Here, six CsAIL genes in the cucumber genome were identified. Bioinformatic analyses revealed high evolutionary conservation of gene structures, protein domains, and motifs among CsAIL members. Expression profiling revealed tissue-specific patterns of CsAILs, hinting at their functional diversification in cucumber growth and development. Notably, CsAIL1 exhibited higher expression levels in ovaries, fruit spines, and seeds, implying its specialized role in these tissues, whereas no functions of AIL1 homologs were reported in other plant species. CsAIL members were also involved in abiotic stress responses, as evidenced by altered expression patterns under salt treatment. Additionally, CsAIL1 protein was localized in the nucleus and was predicted to interact with the ethylene response factor CsERF13. These findings provide a foundational understanding of the CsAIL gene family and contribute valuable insights towards the functional characterization of CsAIL genes in cucumber.
-
Key words:
- AINTEGUMENTA-LIKE /
- Cucumber /
- Evolution /
- Fruit development /
- Gene expression pattern /
- Salt stress