









-
Soil salinization is an increasingly severe global challenge, significantly limiting plant growth and reducing crop yield. According to the latest Food and Agriculture Organization of the United Nations (FAO) report, salinization affects approximately 1.381 billion hectares of land globally, representing 10.7% of the world's land area[1]. Furthermore, FAO estimates indicate that salinity (high salt content) or sodicity (high sodium concentration) affects 10% of irrigated cropland and 10% of rainfed cropland[2]. The occurrence of soil salinization is mainly due to the accumulation of soluble salts, including sodium (Na+), magnesium (Mg2+), potassium (K+), sulfate (SO42−), and chloride (Cl−), which impair plant function by inducing cellular toxicity and physiological drought in plants[3].
Most plants are glycophytes, which are adapted to low-sodium environments. Only a small group of plants are halophytes, and can thrive in high-salinity soils[4]. When exposed to salinity stress, nearly all plants exhibit physiological responses, such as water loss from roots, disruption of iron uptake, reduction of photosynthetic efficiency in leaves, and inhibition of plant growth. Thus, plants have evolved multifaceted adaptation mechanisms, which mainly include ionic equilibrium, activation of antioxidant enzymes, osmoregulation and protection of small molecule substances, and regulation of plant hormones[3,5−7].
Transcription factors, such as NAC (NAM, ATAF1/2, and CUC2), ERF (ethylene response factor), bZIP (basic leucine zipper), WRKY, MYB, DREB (dehydration responsive element binding), bHLH (basic helix-loop-helix), GRAS (GAI-RGA- and -SCR), and ZFP (zinc finger protein), play crucial roles in regulating plant responses to salt stress[8]. Zinc finger proteins (ZFPs), among the most abundant proteins in plants, mediate responses to various abiotic stresses, including salt, drought, flooding, cold, and high light[9−11]. Characterized by zinc-finger domains, ZFPs coordinate one or more zinc ions to stabilize their structure through cysteine (Cys) and/or histidine (His) residues. Based on the numbers and positions of these Cys/His residues, ZFPs are classified into different types such as C2H2, CCHC, PHD, RING, and others[9,12]. Stress-associated proteins (SAPs), a type of ZFPs, are defined by the presence of both A20 and AN1 zinc-finger domains[13]. Current studies demonstrated that SAPs can form homodimers or heterodimers through their A20 and AN1 zinc-finger domains to interact with other proteins, thereby influencing plant responses to stress[14]. SAP genes are involved in stress responses by regulating stress-responsive gene expression, maintaining cellular homeostasis, stabilizing the cell membrane, and improving photosystem performance[15−17]. However, the specific functions of SAPs in regulating salt tolerance in grasses are far from being understood.
Bermudagrass (Cynodon dactylon) is a major perennial warm-season turfgrass species widely used in parks, home lawns, golf courses, and sports fields as well as for ecological restoration, owing to its vigorous vegetative propagation, extensive stolon spread, and strong tolerance to drought and salinity[18]. SAPs play crucial roles in regulating plant growth and stress responses; however, SAP family members in bermudagrass remain unidentified and functionally uncharacterized. This study aimed to isolate CdSAP2 and elucidate its role and molecular mechanisms in regulating salt tolerance.
-
The sequences of AtSAP2 (At1G51200) from Arabidopsis were used for searching bermudagrass expressed sequence tags (ESTs) from the National Center for Biotechnology Information (NCBI) EST database. Full-length coding sequence of CdSAP2 was amplified from the cDNAs of bermudagrass (Cynodon dactylon cv. Tifway), using primer set (CdSAP2for and CdSAP2rev, Supplementary Table S1) with restriction enzymes SalI and EcoRV recognition sites and cloned into pMD19-T vector. CdSAP2 was excised from the pMD19-T vector with SalI and EcoRV digestion and ligated to pENTR™1A. Linearized pENTR™1A-CdSAP2 was recombined using Gateway™ LR Clonase™ II (Invitrogen, Carlsbad, CA, USA) into the destination vector, pEarlygate103, for Arabidopsis (Ecotype Columbia-0) transformation. The resulting plasmid pEarleyGate103-CdSAP2 was transformed into Agrobacterium tumefaciens strain EHA105 and transformed to Arabidopsis by floral dipping[19]. The Arabidopsis transgenic lines were selected using a glufosinate ammonium resistance assay at a concentration of 20 mg·L−1, and identified by genome PCR and RT-qPCR using the specific primers of the CdSAP2 coding sequence (Supplementary Table S1).
Bioinformatic analysis of CdSAP2
-
The molecular weight and conserved domains of CdSAP2 were analyzed using the Sequence Manipulation Suite (SMS) online program (
www.puregion.cn/dcode/calculator/sms/protein_mw.html ) and the Conserved Domain Search Database (CDD,www.ncbi.nlm.nih.gov/Structure/cdd/cdd.shtml ), respectively. Multiple sequence alignment of CdSAP2, AtSAP2, OsSAP4 (Os02g10200), and OsSAP8 (Os06g41010) was performed using BioXM software (version 2.7,https://cbi.njau.edu.cn/BioXM ). The phylogenetic tree was constructed using the neighbor-joining method in MEGA X[20].Transcription activation assay
-
The transcription activation activity of CdSAP2 was carried out using the ProQuest™ Two-Hybrid System with Gateway® Technology (Invitrogen, CA, USA). CdSAP2 and the positive control, pCL1, were recombined into pDEST32 vectors, respectively. The recombined vectors were transformed into yeast (Saccharomyces cerevisiae) strain MaV203 using the EZ-Yeast™ Transformation Kit (MP Biomedicals, CA, USA), and screened on synthetic defined medium lacking leucine (SD/-Leu) agar plates. Single colonies were cultured in SD/-Leu liquid medium and then gradiently diluted and grown on agar plates containing SD/-Leu and SD/-Leu/-His/-Ade to analyze transcription activation activity.
Plant materials and growth conditions
-
Uniform stolons of bermudagrass were cut and planted in plastic containers (41 cm length, 31 cm width, and 15 cm depth) filled with 1/2 Hoagland's nutrient solution[21], then maintained in a controlled growth chamber (MT8070iE, Xubang, Henan, China) with growing conditions maintained at 30/25 °C (day/night), 70% relative humidity, and 14/10-h photoperiod with photosynthetically active radiation of 650 μmol·m−2·s−1 for 2 weeks.
Arabidopsis seeds were surface-sterilized with 75% ethyl alcohol for 30 s and 1.5% sodium hypochlorite for 10 min, rinsed thoroughly (5 to 6 times) with sterile water, and sown on Murashige and Skoog (MS) solid medium[22]. The seeds were stratified at 4 °C in darkness for 3 d, and then transferred to a controlled growth chamber (Xubang, Henan, China) with growing conditions maintained at 23/18 °C (day/night), 70% relative humidity, and 16/8-h photoperiod with photosynthetically active radiation of 200 μmol·m−2·s−1.
Stress treatments for bermudagrass and Arabidopsis
-
To detect expression patterns of CdSAP2 in response to different abiotic stresses, one-month-old bermudagrass plants were treated with PEG6000 (30% polyethylene glycol 6,000 solution, m/v), 100 μM abscisic acid (ABA), 300 mM NaCl, and 4 °C cold stress for 48 h. Leaf and root samples were collected at 0, 1, 3, 6, 12, and 24 h under different treatments and stored at –80 °C for further analysis.
To compare seed germination phenotype between wild type (WT) and CdSAP2-overexpressing lines under salt stress, seeds from T3 generation of CdSAP2-overexpressing lines and WT were sown on petri dishes (9 cm diameter) containing MS solid medium with 150 mM NaCl. After stratification, the seedlings were photographed, and the seed germination rate was measured after 10 d at growing conditions of 23/18 °C (day/night), 70% relative humidity, and 16/8-h photoperiod with photosynthetically active radiation of 200 μmol·m−2·s−1.
To compare seedling growth between WT and CdSAP2-overexpressing lines under salt stress, ten-day-old seedlings were transplanted to 10 cm × 10 cm square petri dishes containing MS solid medium with 150 mM NaCl. After 10 d of treatment, samples were collected and used for further physiological analysis. In addition, another set of ten-day-old seedlings was transplanted to plastic pots (6 cm diameter) filled with potting soil (Pindstrup Plus, Pindstrup Mosebrug A/S, Denmark) and vermiculite (1:2; V/V). After one month of establishment, plants were exposed to 150 mM NaCl treatment, and the samples were collected at 0, 5, and 10 d of salt treatment and used for further physiological analysis. All the treatments were conducted in a growth chamber under the same environmental conditions as described above.
RNA isolation and RT-qPCR analysis
-
Total RNA was isolated using an E.Z.N.A.® Plant RNA Kit (OMEGA Bio-Tek, Norcross, GA, USA) according to the manufacturer's instructions. Two μg total RNA were reverse-transcribed to cDNA using a PrimeScript™ RT reagent Kit with gDNA Eraser (Takara, Shiga, Japan). The RT-qPCR analysis was performed using a LightCycler 480 II Instrument (Roche Diagnostics, Rotkreuz, Switzerland). The total reaction volume was 20 μL, containing 10 μL of SYBR Green Master mix (Roche), 2 μL of cDNA template, 1 μL forward primer, 1 μL reverse primer, and 6 μL diethyl pyrocarbonate-treated water (Invitrogen, Carlsbad, CA, USA). The thermo-cycling condition was set as follows: initial denaturation at 95 °C for 10 min, followed by 40 cycles of 95 °C for 15 s, 60 °C for 15 s, and 72 °C for 20 s. CdPP2A[23] and AtUBQ10 (AT4G05320) were selected as the reference genes for bermudagrass and Arabidopsis, respectively. Each biological sample has three technical replicates, and the expression level of target genes was calculated using the 2−ΔΔCᴛ method[24]. The RT-qPCR primers were listed in Supplementary Table S1.
Leaf electrolyte leakage, chlorophyll content, photochemical efficiency, and gas exchange parameters
-
Leaf electrolyte leakage (EL) was determined by measuring conductivity with a conductivity meter (Thermo Fisher Scientific, Waltham, USA). EL was calculated as: EL (%) = Ci/Cmax × 100, where Ci is the initial conductivity, and Cmax is the maximum conductivity of the sample after boiling[25].
Leaf chlorophyll content was measured on fully expanded leaves using a SPAD-502 chlorophyll meter (Konica-Minolta, Osaka, Japan). Leaf photochemical efficiency, expressed as the maximum quantum yield of photosystem II (Fv/Fm), was determined using a fluorescence induction monitor (OPTI-Sciences, Hudson, USA). Prior to measurement, leaves were dark-adapted for 30 min. The Fv/Fm ratio was calculated as: Fv/Fm = (Fm−Fo)/Fm, where Fm is the maximal chlorophyll fluorescence yield of the dark-adapted state, and Fo is the minimum chlorophyll fluorescence yield after dark adaptation. Leaf gas exchanges parameters, including stomatal conductance (Gs), net photosynthetic rate (Pn), intercellular CO2 concentration (Ci), and transpiration rate (Tr), were measured using a LI-6400XT Portable Photosynthesis System (LI-COR, Lincoln, USA) under the conditions of CO2 concentration at 400 µmol·mol−1, light intensity at 200 µmol·m−2·s−1, and flow rate at 300 µmol·m−2·s−1.
Contents of Na+ and K+
-
One hundred mg of dried leaf samples were weighed and used for the determination of Na+ and K+ concentrations. The samples were digested in nitric acid and transferred into volumetric flasks filled up to 50 mL with deionized water, and the concentrations of Na+ and K+ were analyzed using an inductively coupled plasma optical emission spectroscopy (ICP-OES; Pekin Elmer, Norwalk, USA) following the protocol described in Mäser et al.[26].
Determination of reactive oxygen species
-
The content of hydrogen peroxide (H2O2) was determined using a potassium iodide (KI)-catalyzed colorimetric assay[27]. Half gram of fresh leaf samples was homogenized in 5 mL of 0.1% trichloroacetic acid (m/v) using a mortar and pestle. After centrifugation at 12,000 rpm for 15 min at 4 °C, 0.5 mL of the supernatant was mixed thoroughly with 0.5 mL of 100 mM potassium phosphate buffer (pH 7.0) and 1 mL of 1 M KI solution. After incubation at room temperature in the dark for 1 h, the absorbance at 390 nm was measured, and the H2O2 content was calculated using a standard curve generated from known H2O2 concentrations.
Histochemical localization of superoxide (O2.−) and H2O2 was performed using nitro blue tetrazolium chloride (NBT) and 3,3'-diaminobenzidine (DAB) staining, respectively[28,29]. Leaves from WT and CdSAP2-overexpressing lines were cut and incubated in NBT staining solution (0.5 mg·mL−1) and DAB staining solution (1 mg·mL−1). Samples were then vacuum-infiltrated with their respective staining solutions for 2 h at 25 °C in darkness. After staining, leaf chlorophyll was removed by decolorizing leaves in 95% ethanol using a boiling water bath. The stained tissues were then visualized and photographed under a stereomicroscope (Olympus, Tokyo, Japan).
Contents of proline and soluble sugar
-
The content of proline was quantified using a colorimetric method[30]. Fresh leaf samples (0.2 g) were homogenized in 5 mL of 3% sulfosalicylic acid (w/v) and incubated in a boiling water bath for 10 min. After centrifugation at 12,000 rpm for 10 min, 2 mL of supernatant was mixed with 2 mL of glacial acetic acid and 2 mL of acidic ninhydrin solution. After incubation in a boiling water bath for 30 min and cooling in darkness for 2 h, 2 mL of toluene was added to the mixed solution. Following a 2-h incubation in darkness, the absorbance of the toluene phase was measured at 520 nm, and proline content was calculated using a standard curve prepared with known proline concentrations.
The content of soluble sugar was determined using the anthrone-sulfuric acid method[31]. Half gram of fresh leaf samples was homogenized in 10 mL of deionized water and incubated in a boiling water bath for 10 min. After incubation at room temperature in darkness for 2 h, the homogenate was centrifuged at 12,000 rpm for 10 min. Subsequently, 0.1 mL of supernatant was mixed with 1 mL of deionized water and 4 mL of 0.2% anthrone reagent (w/v). The mixture was incubated in a boiling water bath for 10 min, cooled in darkness for 2 h, and the absorbance was measured at 620 nm. The content of soluble sugar was calculated using a standard curve prepared with known glucose concentrations.
Statistical analysis
-
The statistical significance of all data was subjected to analysis of variance (ANOVA) using SPSS22 software (SPSS Inc., Chicago, IL, USA). The statistical significance of data was assessed utilizing the least significant difference (LSD) test at p < 0.05. All figures were generated using Sigmaplot (version 14.0, Systat Software Inc., London, UK).
-
According to the assembled sequence of SAP family in bermudagrass EST database, a 516 bp homologous sequence was isolated from bermudagrass cDNA. The full-length sequence of CdSAP2 (NCBI accession number: ES299586) encoded a polypeptide of 171 amino acids with a molecular weight of 18.34 kilodaltons. CdSAP2 had an A20 domain (C-X3-C-X11-C-X2-C) at the N terminus and an AN1 domain (C-X2-C-X10-C-X1-C-X4-C-X2-H-X5-H-X-C) at the C terminus, respectively (Fig. 1a, b). Phylogenetic analysis suggested that CdSAP2 exhibited the highest homology with Arabidopsis thaliana AtSAP2, Oryza sativa OsSAP4, and OsSAP8 (Fig. 1c). Transcriptional activity assay indicated that only the positive control (pDEST32-pCL1) yeast cells could grow on the SD/-Leu/-His/-Ade, indicating that CdSAP2 might not have transcriptional activation activity (Fig. 1d).
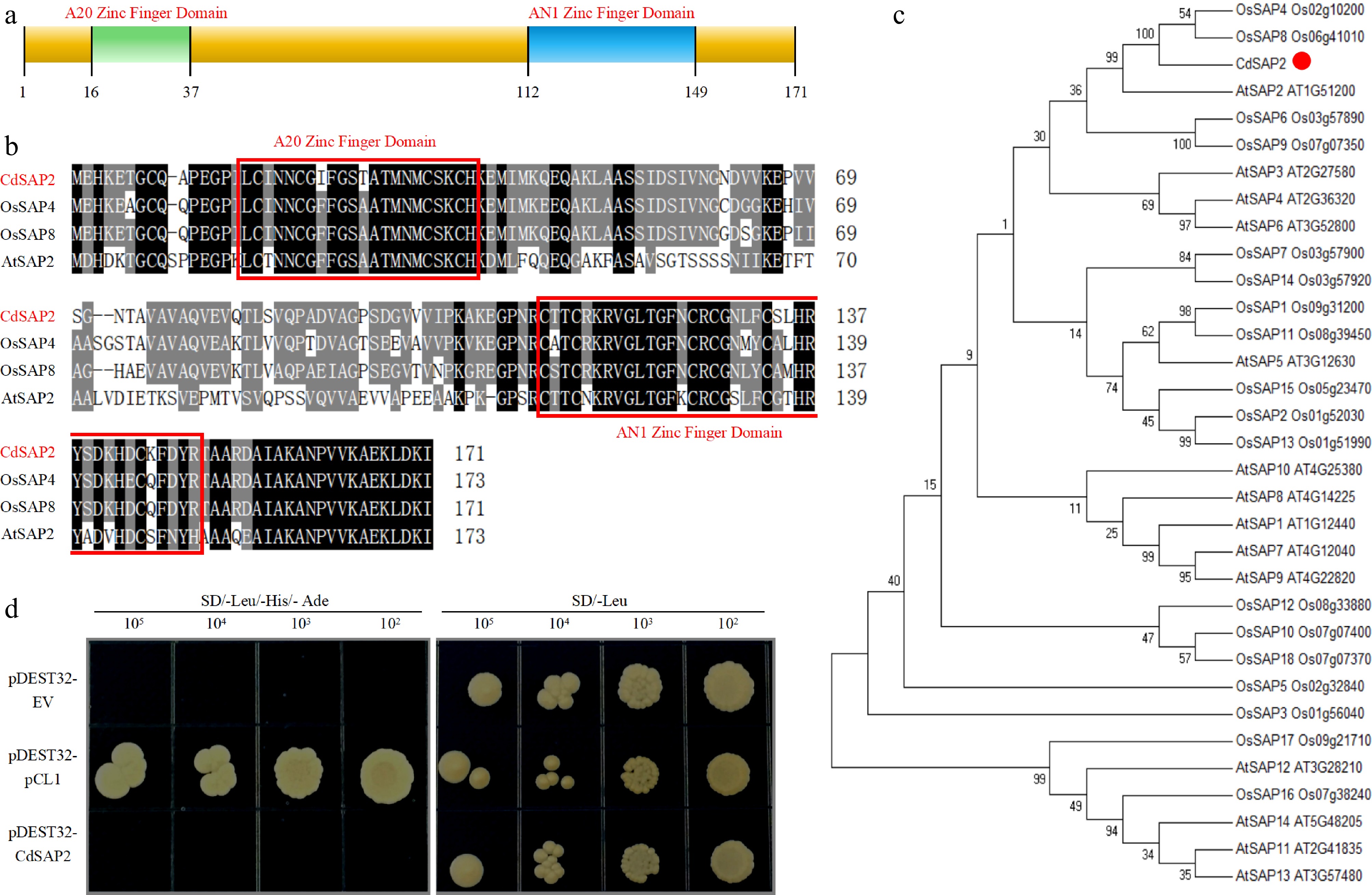
Figure 1.
Sequence analysis, phylogenetic relationship, and transcription activation activity of CdSAP2. (a) The domain composition of CdSAP2. (b) Alignment of amino acid sequences of CdSAP2 with the closest orthologs AtSAP2, OsSAP4, and OsSAP8. The A20 zinc finger domain and the AN1 zinc finger domain are framed in red. (c) Phylogenetic tree of CdSAP2 and 32 additional orthologs. (d) Transcription activation activity of CdSAP2 in yeast cells. The positive control was pCL1, and the negative control was pDEST32 empty vector.
Expression pattern of CdSAP2 in response to abiotic stresses
-
To reveal stress-dependent-specific expression profiles of CdSAP2, bermudagrass seedlings were exposed to different abiotic stresses, including drought (30% PEG6000), 100 mM ABA, salt (300 mM NaCl), and 4 °C cold stress. The expression level of CdSAP2 in leaves was significantly up-regulated after exposure to 12 h of PEG, ABA, and salt treatments, while decreased at 24 h of PEG, ABA, and salt treatments. Under 4 °C cold stress, CdSAP2 showed significant higher expression levels in leaves at 1 and 6 h compared to at 0 h of stress (Fig. 2). CdSAP2 showed a significant down-regulation pattern when exposed to PEG, ABA, salt and cold treatments in roots at 1, 3, and 6 h (Fig. 2).
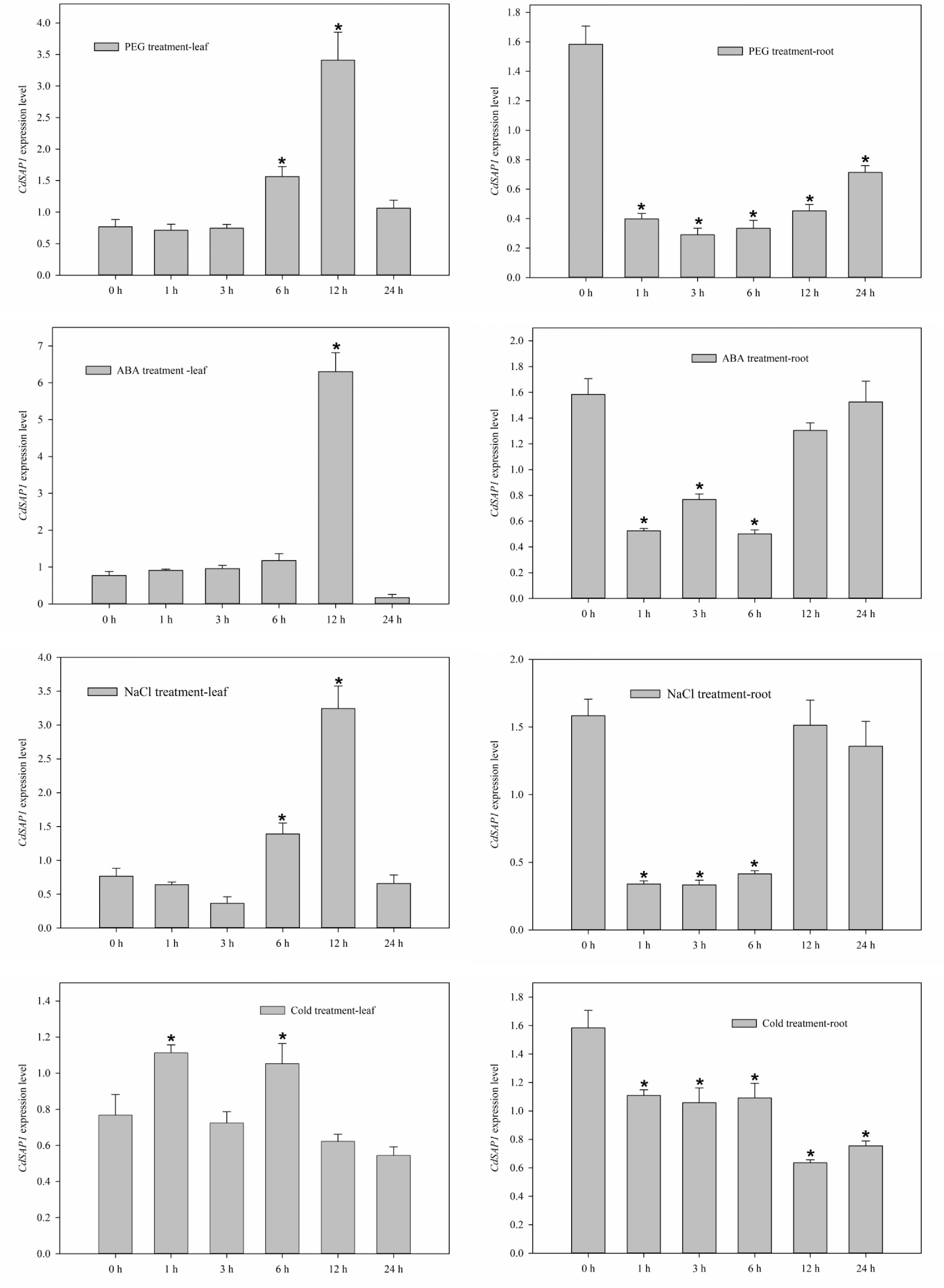
Figure 2.
Expression patterns of CdSAP2 in response to PEG-6000 (30%), ABA (100 mM), NaCl (300 mM), and cold (4 °C) treatments. Data shown were mean values ± SE of three independent biological replicates. Asterisks indicate significant differences according to Fisher's protected LSD test (p < 0.05).
CdSAP2-overexpression improved salt tolerance of transgenic Arabidopsis
-
To investigate the function of CdSAP2 in regulating plant salt tolerance, CdSAP2-overexpressing transgenic Arabidopsis plants driven by cauliflower mosaic virus 35S promoter were generated by flower-dipping-mediated transformation. A total of six independent CdSAP2-overexpressing transgenic Arabidopsis (OE1-OE6) were obtained and confirmed by the presence of the CdSAP2 gene (Supplementary Fig. S1a). In addition, the relative expression level of CdSAP2 suggested that CdSAP2 mRNA was accumulated in the transgenic plant lines, in which OE4 and OE5 lines were selected for further salt tolerance analysis (Supplementary Fig. S1b).
The T3 generation seeds of two CdSAP2-overexpression lines and WT were sown on MS medium containing 150 mM NaCl. Under non-stress conditions, there were no significant differences in phenotype between CdSAP2-overexpressing lines (OE4 and OE5) and WT (Fig. 3a, b). CdSAP2-overexpression significantly enhanced salt tolerance of transgenic Arabidopsis seedlings (OE4 and OE5), as indicated by significantly higher seed germination rate and better growth performance under salt stress (Fig. 3a, b).
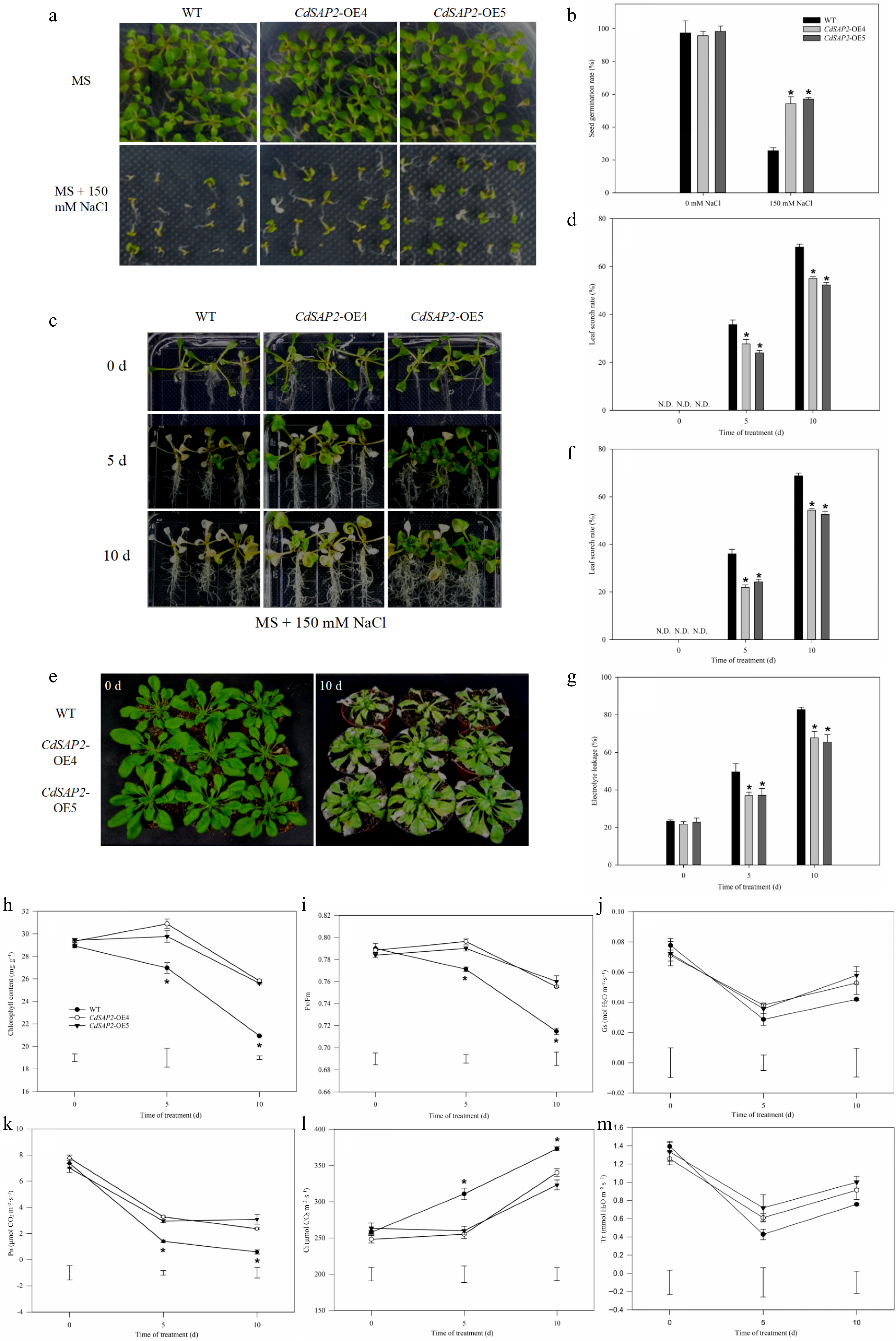
Figure 3.
CdSAP2-overexpression enhanced salt tolerance of Arabidopsis under salt stress. (a) Germination phenotype of WT and transgenic Arabidopsis on MS medium with 150 mM NaCl for 10 d. (b) Seed germination rate of WT and transgenic Arabidopsis on MS medium with 150 mM NaCl. (c) Salt tolerance phenotypes of WT and CdSAP2-overexpressing transgenic Arabidopsis on MS medium with 150 mM NaCl. (d) Statistical analysis of leaf scorch rate. (e) Salt tolerance phenotypes of WT and CdSAP2-overexpressing transgenic Arabidopsis in soil with 150 mM NaCl. (f) Statistical analysis of leaf scorch rate. (g) Statistical analysis of electrolyte leakage. (h) Chlorophyll content. (i) Photochemical efficiency (Fv/Fm). (j) Stomatal conductance (Gs). (k) Net photosynthetic rate (Pn). (l) Intercellular CO2 concentration (Ci). (m) Transpiration rate (Tr). N.D. means not detected. The leaf scorch rate was calculated by estimating the percentage of leaves that were scorched (white or pale color). Data shown were mean values ± SE of three independent biological replicates. Asterisks indicate significant differences according to Fisher's protected LSD test (p < 0.05).
In order to determine the role of CdSAP2 in regulating salt stress tolerance, ten-day-old seedlings of WT and CdSAP2-overexpressing transgenic Arabidopsis lines (OE4 and OE5) were transferred to MS medium with 150 mM NaCl. The results showed that CdSAP2-overexpressing lines had significantly better growth performance and lower leaf scorch rate at 5 and 10 d of salt stress, compared with WT (Fig. 3c, d). To further gain insight into the role of CdSAP2 in regulating salt stress tolerance, 10-day-old WT and CdSAP2-overexpressing lines (OE4 and OE5) were transferred to soil, grown for one-month and then treated with 150 mM NaCl for 10 d (Fig. 3e). The results showed that leaf scorch rate in CdSAP2-overexpressing lines (21.97% in OE4, 24.25% in OE5) was significantly lower than that in WT (36.00%) at 5 d of salt stress (Fig. 3f). Similar results were also observed at 10 d of salt stress. In addition, EL in CdSAP2-overexpressing lines were significantly lower than that in WT at 5 and 10 d of salt stress (Fig. 3g).
Under salt stress, chlorophyll content, Fv/Fm and net photosynthetic rate of CdSAP2-overexpressing lines were significantly higher than those of WT under salt treatment (Fig. 3h, i, k), while intercellular CO2 of CdSAP2-overexpressing lines was significantly lower than that of WT (Fig. 3l). Stomatal conductance and transpiration rate showed no significant differences between WT and CdSAP2-overexpressing lines under salt stress (Fig. 3j, m).
CdSAP2-overexpression stimulated ionic balance, antioxidative defense and osmotic adjustment in responses to salt stress
-
To investigate the physiological changes in CdSAP2-overexpressing transgenic Arabidopsis under salt stress, the contents of ions, reactive oxygen species (ROS), and osmotic adjustment substances were determined. Under 150 mM NaCl treatment, K+ content in WT and CdSAP2-overexpressing lines was significantly decreased, and Na+ content was significantly increased, resulting in increased Na+/K+ ratio, at 5 and 10 d of salt stress. Under salt stress, there was no significant difference in K+ content between WT and CdSAP2-overexpressing plants, while CdSAP2-overexpressing plants exhibited significantly lower Na+ content and Na+/K+ ratio compared with WT (Fig. 4a−c).
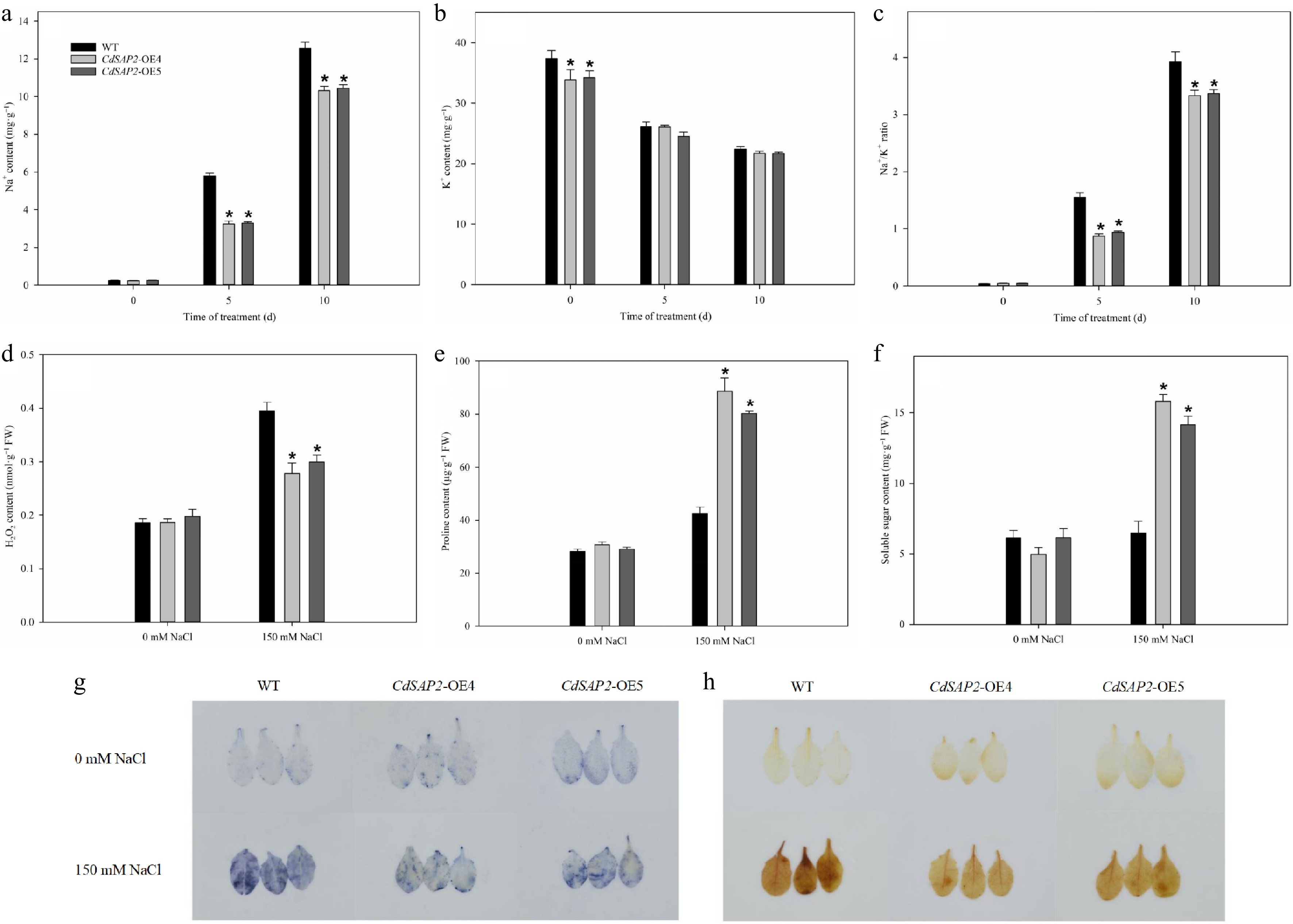
Figure 4.
Overexpression of CdSAP2 affected the content of ions, reactive oxygen species, and osmotic adjustment substances in Arabidopsis leaves under salt stress. (a) Na+ content. (b) K+ content. (c) Na+/K+ ratio. (d) H2O2 content. (e) Proline content. (f) Soluble sugar content. (g) O2·− accumulation by NBT staining. (h) H2O2 accumulation by DAB staining. Data shown were mean values ± SE of three independent biological replicates. Asterisks indicate significant differences according to Fisher's protected LSD test (p < 0.05).
Under salt stress, the accumulation of ROS, such as O2·− and H2O2, was induced in Arabidopsis leaves. However, CdSAP2-overexpression reduced the accumulation of O2·− and H2O2 by salt stress in Arabidopsis (Fig. 4g, h). In addition, the quantification assay also showed that H2O2 contents in CdSAP2-overexpressing lines were significantly lower than those in WT under salt stress (Fig. 4d).
Under non-stress condition, there were no significant differences in the contents of proline and soluble sugar between WT and CdSAP2-overexpressing lines. After 10 d of salt stress, the proline contents of OE4 and OE5 were 108.14% and 88.63% higher than those of WT, while the soluble sugar contents of OE4 and OE5 were 144.46% and 118.50% higher than those of WT, respectively (Fig. 4e, f).
Overexpression of CdSAP2 modulated expression level of genes involved in ionic balance, antioxidant defense, and osmotic regulation
-
Given that CdSAP2-overexpression regulated ionic, antioxidative, and osmotic responses of Arabidopsis, the expression level of genes related with ionic balance (AtSOS1, AtAKT1, and AtNHX1), antioxidant defense (AtAPX, AtSOD, and AtPOD) and osmotic adjustment (AtBADH and AtP5CS) were further analyzed between WT and CdSAP2-overexpressing lines at 0, 6, 12, and 24 h of salt stress. The expression levels of AtSOS1 and AtNHX1 in leaves of CdSAP2-overexpressing lines were significantly higher than those of WT at 12 and 24 h of salt stress, respectively (Fig. 5a, c). The expression levels of AtAPX, AtSOD, and AtPOD in leaves of CdSAP2-overexpressing transgenic Arabidopsis were significantly higher than those in WT plants at 12 and 24 h of salt treatment (Fig. 5d−f). The expression level of AtBADH in OE4 plants was significantly higher than WT at 24 h of salt treatment (Fig. 5g). The expression level of AtP5CS in CdSAP2-overexpressing lines was significantly higher than those of WT at 6 and 12 h of salt treatment (Fig. 5h). The expression level of AtAKT1 didn't show a significant difference between WT and CdSAP2-overexpression lines under salt treatment (Fig. 5b).
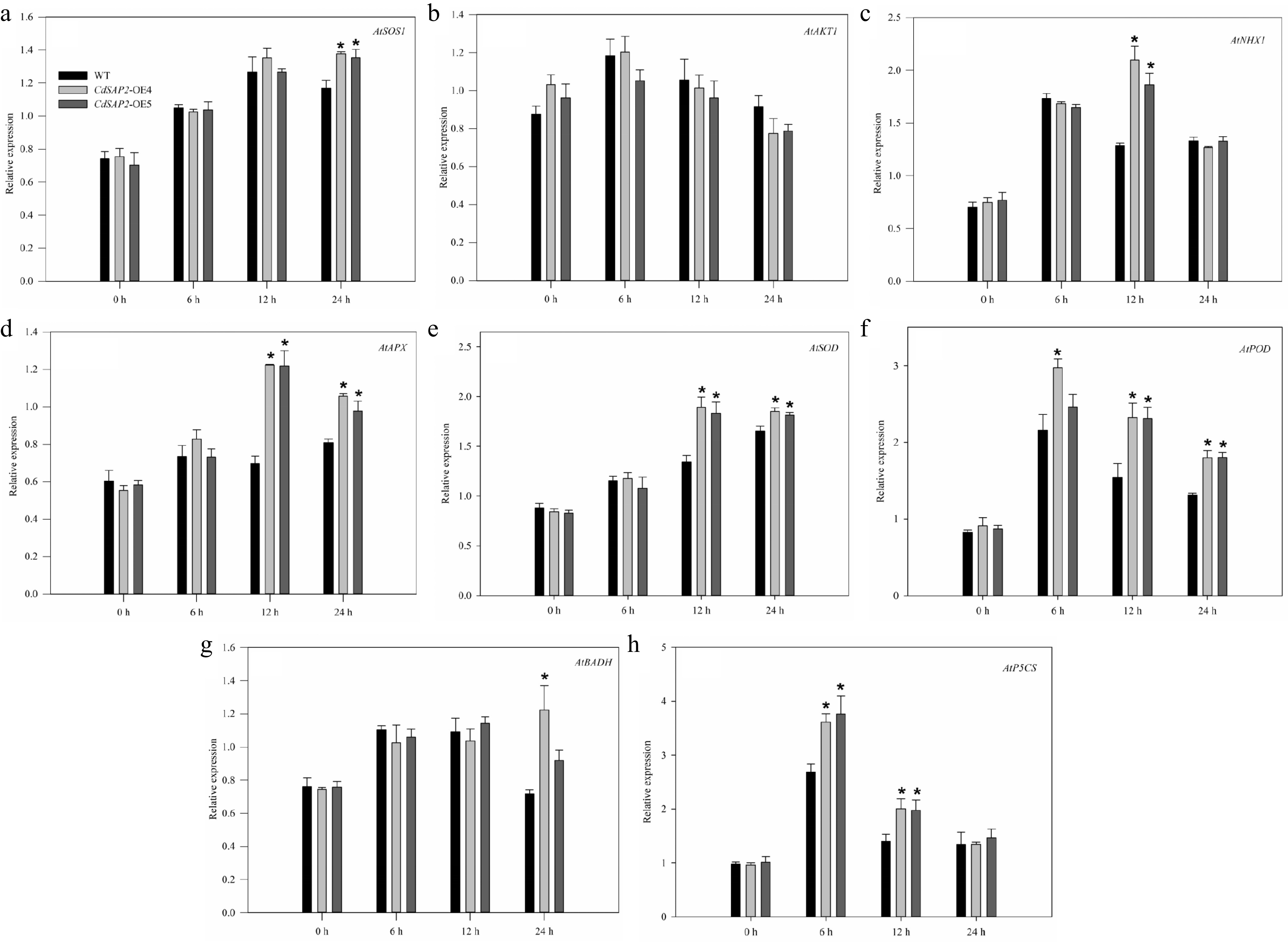
Figure 5.
Overexpression of CdSAP2 affected expression patterns of stress tolerance related genes in Arabidopsis leaves under salt stress. Data shown were mean values ± SE of three independent biological replicates. Asterisks indicate significant differences according to Fisher's protected LSD test (p < 0.05).
-
Abiotic stresses, such as salinity, cold, heat, and drought, significantly limit plant growth and development. Among these, salt stress is one of the most detrimental to crop yield[32]. To mitigate salt-induced damage, plants have evolved various physiological and molecular mechanisms, including maintaining membrane stability, ion homeostasis, antioxidant defense systems, and protecting the photosynthetic apparatus[3,33]. SAPs, a novel class of ZFPs, have been implicated in responses to multiple abiotic stresses. SAP family members have been identified through genome-wide analyses in monocot and dicot plants, including rice (Oryza sativa subsp. indica)[34], Arabidopsis[35], maize (Zea mays)[36], soybean (Glycine max)[37], sweet potato (Ipomoea batatas)[17], and potato (Solanum tuberosum)[38]. Specific members of the rice SAP family, such as OsSAP4[39] and OsSAP8[40], play roles in salt stress tolerance responses. OsSAP4 negatively modulates stress tolerance by influencing osmotic adjustment substances (proline and soluble sugar), gibberellins, and ABA biosynthesis. OsSAP8 has been reported to confer salt tolerance during the germination and seedling stage of rice. Furthermore, evidence indicated that OsSAP6 responds to soda saline-alkaline stress and interacts with OsPK5 to enhance stress tolerance by regulating ROS homeostasis in rice[41]. However, the physiological and molecular mechanisms by which SAPs regulate salt tolerance, particularly their roles in maintaining membrane stability, ionic homeostasis, and photosystem performance, require further characterization.
In the present study, a member of the SAP family, CdSAP2, was isolated from bermudagrass. The CdSAP2 gene contains a 513 bp open reading frame encoding a protein of 171 amino acids. Sequence alignment and phylogenetic analysis revealed that CdSAP2 exhibited high homology to OsSAP4 and OsSAP8 from rice (Fig. 1c). Bioinformatics analysis identified conserved domain A20 and AN1 domains within the CdSAP2 protein, characteristic of SAP family members (Fig. 1a, b). Given that most SAP proteins function in conferring abiotic stress tolerance in transgenic plants[42], the results from the present study suggested that the SAP family is highly conserved not only in amino acid sequence but also in biological function. The results demonstrated that CdSAP2, similar to its homologous genes OsSAP4, OsSAP8, and OsSAP6, was involved in multiple pathways contributing to salt tolerance. Transcriptional activation assays indicated that CdSAP2 might lack transcriptional activation potential (Fig. 1d). This finding is consistent with the previous report that the rice homolog, OsSAP4, also lacks transcriptional activity[39], suggesting that CdSAP2 may not function as a transcription factor in stress responses.
Plants growing in saline environments face two primary abiotic stresses: ionic stress, caused by excessive accumulation of toxic ions such as Na+, and osmotic stress, resulting from decreased external water potential that limits water uptake[32,33]. It is well established that Na+ exclusion and maintenance of a low Na+/K+ ratio are key determinants of plant salt tolerance[43,44]. Maintaining ionic homeostasis under salinity primarily involves limiting Na+ influx and preventing K+ efflux. For instance, in Arabidopsis thaliana, the plasma membrane Na+/H+ exchanger Salt-Overly-Sensitive 1 (SOS1) was phosphorylated and activated under salt stress, facilitating the extrusion of excess Na+ from cells, regulating long-distance Na+ transport from roots to shoots, and maintaining cellular ionic homeostasis[45−47]. Similarly, a tonoplast Na+/H+ antiporter, AtNHX1, has been found to contribute to salt tolerance of Arabidopsis through transporting cytosol Na+ into the vacuole and limiting Na+ translocation from roots to shoots[48−50]. Furthermore, the voltage-gated K+-selective channel protein K+ Transporter1 (AKT1) was phosphorylated and activated for K+ uptake under salt stress conditions[51,52]. In the present study, CdSAP2-overexpressing transgenic Arabidopsis plants exhibited significantly lower Na+ content and Na+/K+ ratio (Fig. 4a, c), coupled with higher expression levels of AtSOS1 and AtNHX1 at 12 and 24 h of salt treatment (Fig. 5a, c). These results suggested that CdSAP2 enhanced salt tolerance in Arabidopsis, potentially by promoting Na+ exclusion and compartmentalization, leading to sustained lower Na+ concentration and Na+/K+ ratio in leaves.
Salt stress typically induces excessive accumulation of ROS in plants, leading to cellular damage and consequently impairing plant growth and development[53]. The plant antioxidant defense system protects cells from oxidative damage by scavenging excessive ROS[54,55]. A key adaptive strategy of this system involves regulating the expression and/or activity of ROS-scavenging enzymes, including ascorbate peroxidase (APX), superoxide dismutase (SOD), and peroxidase (POD)[56,57]. In the present study, CdSAP2-overexpressing transgenic Arabidopsis plants exhibited significantly reduced accumulation of O2·− and H2O2 under salt stress (Fig. 4d, g, h). In addition, the expression levels of genes encoding ROS-scavenging enzymes (AtAPX, AtSOD, and AtPOD) were significantly higher in CdSAP2-overexpressing transgenic Arabidopsis plants compared to WT plants (Fig. 5d−f). This finding is consistent with previous reports that overexpression of a SAP family gene from Sorghum bicolor, SbSAP14, enhanced antioxidant enzyme activities and conferred salt stress tolerance in rice[58]. These results suggested that CdSAP2-overexpression enhanced plant salt tolerance by upregulating the expression of genes involved in the ROS-scavenging pathway, thereby mitigating oxidative damage.
Salt stress impairs plant water uptake capacity, leading to dehydration, loss of cell turgor, and ultimately osmotic stress[32,33]. To counteract osmotic stress, plants accumulate compatible solutes, such as proline, soluble sugar, and glycine betaine, which function as key osmotic regulatory substances in plants[59]. In this study, CdSAP2-overexpressing transgenic Arabidopsis plants accumulated significantly higher contents of proline and soluble sugar than WT under salt stress (Fig. 4e, f). This aligns with the previous observation that GmSAP16-overexpressing Arabidopsis plants also exhibited increased proline accumulation under salt stress and enhanced salt tolerance[37]. The content of proline and soluble sugar is a well-established indicator of salinity tolerance[60,61]. Proline biosynthesis is primarily regulated by pyrroline-5-carboxylate synthetase (P5CS), while betaine aldehyde dehydrogenase (BADH) is a key enzyme in glycine betaine synthesis[62]. Similarly, the expression levels of two genes encoding osmotic adjustment-related proteins (AtBADH and AtP5CS) were also upregulated in CdSAP2-overexpressing plants (Fig. 5g, h). Collectively, these results demonstrated that CdSAP2-overexpression promoted the accumulation of compatible solutes, specifically proline and soluble sugar, in response to salt stress, thereby improving salt stress tolerance in plants.
In summary, a salt-responsive SAP-type zinc finger protein, CdSAP2, was cloned from bermudagrass. Under salt stress, CdSAP2-overexpressing transgenic Arabidopsis plants exhibited significantly increased germination rate, chlorophyll content, Fv/Fm, Pn, and accumulation of proline and soluble sugar compared to WT plants, indicating that CdSAP2 was a positive regulator of plant salt tolerance. Physiological and gene expression analyses revealed that CdSAP2 enhanced salt tolerance by coordinately modulating salt exclusion, osmotic adjustment, and ROS scavenging. CdSAP2 could be a valuable gene resource for molecular breeding programs aimed at improving salt tolerance in turfgrasses and potentially other crops.
This research was funded by the Science and Technology Program of Jiangsu Vocational College of Agriculture and Forestry (Grant No. 2021kj41), National Natural Science Foundation of China (Grant No. 32101437), and China Postdoctoral Science Foundation (Grant Nos 2018T110518, 2017M611842).
-
The authors confirm contribution to the paper as follows: study conception and design: Liu Y, Chen Y; data collection: Liu Y, Liu YN; analysis and interpretation of results: Liu Y, Zhang X, Liu YN, Chen Y; draft manuscript preparation: Liu Y, Zhang X; manuscript revision: Liu J, Yang Z, Chen Y. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article.
-
The authors declare that they have no conflict of interest.
- Supplementary Table S1 Primers used in this study.
- Supplementary Fig. S1 Identification of CdSAP2-overexpressing transgenic Arabidopsis.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Zhang X, Liu Y, Liu J, Yang Z, et al. 2025. Overexpression of stress-associated protein 2 (CdSAP2) from Cynodon dactylon enhances salt tolerance. Grass Research 5: e034 doi: 10.48130/grares-0025-0032 

Overexpression of stress-associated protein 2 (CdSAP2) from Cynodon dactylon enhances salt tolerance
- Received: 07 August 2025
- Revised: 10 October 2025
- Accepted: 14 October 2025
- Published online: 25 December 2025
Abstract: Excess salt reduces soil fertility and limits plant growth and development. Enhancing salt tolerance is crucial for improving land utilization and crop yield. Bermudagrass (Cynodon dactylon) is a widely cultivated warm-season turfgrass worldwide, with high tolerance to heat, drought, and salinity stresses. In the present study, a stress-associated protein 2 (CdSAP2) was cloned from bermudagrass. The domain analysis showed that CdSAP2 had A20 and AN1 zinc-finger domains. Expression pattern analysis revealed that the transcript level of CdSAP2 in leaves was significantly induced by abscisic acid, drought, NaCl, and cold treatments. CdSAP2 overexpression significantly enhanced salt tolerance in Arabidopsis, as manifested by significantly increased germination rate, higher chlorophyll content, photochemical efficiency, and net photosynthetic rate, higher contents of osmotic regulatory substances (proline and soluble sugar), but lower leaf scorch rate, lower electrolyte leakage, lower Na+/K+ ratio, and lower reactive oxygen species accumulation under salt stress. RT-qPCR analysis showed that improved salt stress tolerance by CdSAP2 overexpression might be related to upregulated expression of genes involved in ionic translocation (SOS1, NHX1), antioxidant defense (APX, SOD, POD), and osmotic adjustment (BADH and P5CS).
-
Key words:
- Cynodon dactylon /
- Stress-associated proteins /
- CdSAP2 /
- Salt tolerance