









-
Flowers play a crucial role in the reproductive cycle of flowering plants, which attract pollinating insects with their bright hues and unique scents. The pollinators accidentally transmit pollen to the flowers and transfer it to subsequent flowers. These pollinators carry a variety of microorganisms, particularly bacteria, filamentous fungi, and yeasts. The term 'anthophilous' has been used for microorganisms that are flower-associated, suggesting that they may play a role in pollination ecology or plant-microbe interactions[1,2]. Anthophilous microorganisms can be dispersed through the nectar and structures of different flowers, such as petals, stamens, and pistils, which provide a nutrient-rich environment including pollen rich in proteins and amino acids, nectar containing 15 to 75% sugar, and flower tissue. These flower structures support a unique anthophilous yeast community that can adapt to its specific host flower. In addition, the relationship between flowers and their associated anthophilous yeasts represents an important ecological interaction, in which yeasts can influence pollination dynamics, floral scent, and plant reproduction[2−6]. However, variations in floral types and growing sites may influence anthophilous yeast communities, potentially leading to the discovery of valuable, previously undiscovered cryptic taxa. Several studies have established flowers as important habitats for anthophilous yeasts, with the genus Metschnikowia being particularly prominent[2,3,7−9]. Additional anthophilous yeast, and yeast-like fungi, commonly detected in nectar and flower surfaces include the genera Candida, Clavispora, Cryptococcus, Debaryomyces, Filobasidium, Hanseniaspora, Hannaella, Kodamaea, Kwoniella, Meyerozyma, Metschnikowia, Operculina, Papiliotrema, Pichia, Rhodotorula, Sporobolomyces, Starmerella, Sympodiomycopsis, Ustilago, Wickerhamiella, and Yamadazyma[3,8−14]. Furthermore, this list of yeast genera is expected to expand as ongoing research continues to uncover the diversity of anthophilous yeasts and yeast-like fungi.
Thailand, located in a tropical region with diverse ecosystems and climates, has been identified by numerous mycologists as a hotspot for the discovery of new filamentous fungal and yeast taxa. The country's rich biodiversity, including its tropical forests, wetlands, and agricultural areas, offers a unique environment for the growth of a wide range of filamentous fungi and yeasts. Interestingly, the variety of floral species in Thailand likely supports a corresponding diversity of anthophilous yeast. For example, novel anthophilous yeasts include Pichia siamensis from flowers of Justicia fragilis and Ervatamia coronaria in Kanchanaburi Province[15], Candida jaroonii from unidentified flowers in northeastern Thailand[16], C. ratchasimensis and C. khaoyaiensis from unidentified flowers in Khao Yai National Park in Nakhon Ratchasima Province[17], C. wangnamkhiaoensis from Ukshi (Calycoopteris floribunda) flowers[18], C. konsanensis isolated from princess jasmine (Jasminum adenophyllum) flowers[19], and Metschnikowia lannaensis, Wickerhamiella camelliae, and W. thailandensis isolated from Assum tea (Camellia sinensis var. assamica) flowers in northern Thailand[9]. According to preliminary investigations, the discovery of novel anthophilous yeast is still limited, particularly in northern Thailand, a region that represents a significant reservoir of microbial diversity[20]. This area features unique topography, with elevations ranging from 400 to 2,500 meters above sea level, a tropical climate, various forest ecosystems, and rich botanical diversity, all of which create numerous specialized microhabitats conducive to yeast colonization. Over the past decade, several yeast species have been discovered in this region that were isolated from soil, plants, and various substrates, including genera Curvibasidium, Cyberlindnera, Cystobasidium, Debaryomyces, Galactomyces, Kazachstania, Lipomyces, Meyerozyma, Naganishia, Papiliotrema, Rhodosporidiobolus, Rhodotorula, Saitozyma, Saturnispora, Schwanniomyces, Sporidiobolus, Trichosporon, and Wickerhamomyces[9,21−26]. Research on anthophilous yeasts in northern Thailand has further revealed a wide variety of species with potential applications, including plant growth promotion, enzyme production, and the synthesis of volatile compounds. Therefore, this study aimed to isolate anthophilous yeasts associated with various flowers in northern Thailand. Yeast identification was carried out using a polyphasic approach that included morphological, biochemical, and physiological characterization, along with multi-locus phylogenetic analyses. The classification of the anthophilous yeasts obtained in this study is presented in Table 1.
Table 1. Content of anthophilous yeasts in this study.
Phylum: Ascomycota Caval. -Sm. Subphylum: Saccharomycotina O.E. Erikss. & Winka Class: Dipodascomycetes M. Groenew., Hittinger, Opulente & A. Rokas Order: Dipodascales M. Groenew., Hittinger, Opulente & A. Rokas Family: Trichomonascaceae Kurtzman & Robnett 1. Entelexis stigmatis Sipiczki ex Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov., (validation and first record in Thailand) 2. Starmerella etchellsii (Lodder & Kreger-van Rij) C.A. Rosa & Lachance, Int. J. Syst. Evol. Microbiol. 68: 1341 (2018) 3. Starmerella orientalis Alimad., Soudi, F.Y. Bai, S.A. Wang & Q.M. Wang ex Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov., (validation and first record in Thailand) 4. Starmerella thailandica Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 5. Wickerhamiella pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Class: Pichiomycetes M. Groenew., Hittinger, Opulente & A. Rokas Order: Serinales M. Groenew., Hittinger, Opulente & A. Rokas Family: Debaryomycetaceae Kurtzman & M. Suzuki 6. Candida tropicalis (Castell.) Berkhout, De Schimmelgesl. Monilia, Oidium, Oospora en Torula, Disset. Ultrecht: 44 (1923) 7. Kodamaea ohmeri (Etchells & T.A. Bell) Y. Yamada, Tom. Suzuki, M. Matsuda & Mikata, Biosc., Biotechn., Biochem. 59: 1174 (1995) 8. Kodamaea restingae (C.A. Rosa, Lachance, Starmer, J.S.F. Barker, J.M. Bowles & Schlag-Edl.) C.Y. Chai & F.L. Hui, MycoKeys 89: 133 (2022) 9. Meyerozyma caribbica (Vaughan-Mart., Kurtzman, S.A. Mey. & E.B. O'Neill) Kurtzman & M. Suzuki, Mycoscience 51: 8 (2010) 10. Priceomyces siamensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Family: Metschnikowiaceae T. Kamieński ex Doweld 11. Metschnikowia cibodasensis Sjamsur., Oetari, C. Nakash., A. Kanti, Saraswati, & K. Ando, J. Microbiol. Biotechnol. 23: 909 (2013), (first record in Thailand) 12. Metschnikowia koreensis S.G. Hong, J. Chun, H.W. Oh & Bae, Int. J. Syst. Evol. Microbiol. 51: 1928 (2001) Class: Saccharomycetes G. Winter Order: Phaffomycetales M. Groenew., Hittinger, Opulente & A. Rokas Family: Phaffomycetaceae Y. Yamada, H. Kawas., Nagats., Mikata & T. Seki 13. Cyberlindnera fabianii (Wick.) Minter, Mycotaxon 110: 474 (2009) Order: Saccharomycodales M. Groenew., Hittinger, Opulente & A. Rokas Family: Saccharomycodaceae Kudryavtsev 14. Hanseniaspora lachancei Čadež, Poot, Raspor & M.T. Sm., Int. J. Syst. Evol. Microbiol. 53: 1679 (2003), (first record in Thailand) Phylum: Basidiomycota R.T. Moore Subphylum: Agaricomycotina Doweld Class: Tremellomycetes Doweld Order: Filobasidiales Jülich Family: Filobasidiaceae L.S. Olive 15. Filobasidium lannaense Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 16. Naganishia albida (Saito) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 118 (2015) 17. Naganishia diffluens (Zach) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 119 (2015) 18. Naganishia liquefaciens (Saito & M. Ota) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 119 (2015) Order: Tremellales Fr. Family: Bulleribasidiaceae Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout 19. Hannaella pagnoccae Landell, L.R. Brandão, A.C. Barbosa, J.P. Ramos, Safar, F.C.O. Gomes, F.M.P. Sousa, P.B. Morais, Broetto, Leoncini, J.R.A. Ribeiro, Fungsin, M. Takash., Nakase, C.F. Lee, Vainstein, Fell, Scorzetti, Vishniac, C.A. Rosa & P. Valente, Int. J. Syst. Evol. Microbiol. 64: 1975 (2013) 20. Hannaella phyllophila Suruss. & Limtong, Int. J. Syst. Evol. Microbiol. 65: 2138 (2014) 21. Vishniacozyma marinae Q.M. Wang, Stud. Mycol.109: 119 (2024) 22. Vishniacozyma pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Family: Cryptococcaceae Kütz. ex Castell. & Chalm. 23. Kwoniella bestiolae (Thanh, Hai & Lachance) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 137 (2015), (first record in Thailand) 24. Kwoniella heveanensis Metin, K. Findley & Heitman, Mycotaxon 116: 227 (2011) 25. Kwoniella limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 26. Kwoniella saisamorniae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Family: Rhynchogastremaceae Oberw. & B. Metzler 27. Papiliotrema aspenensis (Ferreira-Paim, T.B. Ferreira, Andrade-Silva, D.J. Mora, D.J. Springer, Heitman, F.M. Fonseca, D. Matos, Melhem & Silva-Verg.) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 96: 135 (2020) 28. Papiliotrema flavescens (Saito) Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 126 (2015) 29. Papiliotrema chiangmaiensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 30. Papiliotrema pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 31. Papiliotrema tectonae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Family: Trimorphomycetaceae Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout 32. Saitozyma thailandensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Order: Trichosporonales Boekhout & Fell Family: Trichosporonaceae Nann. 33. Trichosporon asahii Akagi ex Sugita, A. Nishikawa & Shinoda, J. Gen. Appl. Microbiol., Tokyo 40: 405 (1994) Subphylum: Pucciniomycotina R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Class: Agaricostilbomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Order: Agaricostilbales Oberw. & R. Bauer Family: Chionosphaeraceae Oberw. & Bandoni 34. Boekhoutia pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Class: Cystobasidiomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Order: Cystobasidiales R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Family: Cystobasidiaceae Gäum. 35. Cystobasidium benthicum (Nagah., Hamam., Nakase & Horikoshi) Yurkov, Kachalkin, H.M. Daniel, M. Groenew., Libkind, V. de García, Zalar, Gouliam., Boekhout & Begerow, Antonie van Leeuwenhoek 107: 180 (2014), (new habitat record and first record in Thailand) 36. Cystobasidium keelungense C.F. Chang & S.M. Liu, Arch. Microbiol. 201: 31 (2018), (first record in Thailand) 37. Cystobasidium minutum (Cif. & Redaelli) Yurkov, Kachalkin, H.M. Daniel, M. Groenew., Libkind, V. de García, Zalar, Gouliam., Boekhout & Begerow, Antonie van Leeuwenhoek 107: 180 (2014) 38. Cystobasidium thailandicum Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 39. Halobasidium lannaense Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Class: Cystobasidiomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Order: Erythrobasidiales R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Family: Erythrobasidiaceae Denchev 40. Erythrobasidium primogenitum Y.P. Tan, Bishop-Hurley & R.G. Shivas, Index of Australian Fungi 1: 1 (2022), (first record in Thailand) Families Incertae sedis Family: Symmetrosporaceae Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout 41. Symmetrospora chiangmaiensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 42. Symmetrospora hydei Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Class: Microbotryomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw. Order: Incertae sedis Families Incertae sedis 43. Curvibasidium chiangmaiense Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Order: Thailandicolales Kodchasee, Senwanna, J. Kumla & N. Suwannar. Family: Thailandicolaceae Kodchasee, Senwanna, J. Kumla & N. Suwannar. 44. Thailandicolales Kodchasee, Senwanna, J. Kumla & N. Suwannar., ord. nov. 45. Thailandicolaceae Kodchasee, Senwanna, J. Kumla & N. Suwannar., fam. nov. 46. Thailandicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., gen. nov. 47. Thailandicola hydei Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 48. Thailandicola limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Order: Sporidiobolales Doweld Family: Sporidiobolaceae R.T. Moore 49. Rhodosporidiobolus fluvialis (Fell, Kurtzman, Tallman & J.D. Buck) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 181 (2015) 50. Rhodosporidiobolus ruineniae (Holzschu, Tredick & Phaff) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 182 (2015) 51. Rhodotorula paludigena (Fell & Tallman) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 181 (2015) 52. Rhodotorula toruloides (I. Banno) Q.M. Wang, F.Y. Bai, M. Groenew. & Boekhout, Stud. Mycol. 81: 181 (2015) 53. Rhodotorula thailandensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 54. Sporobolomyces thailandensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Subphylum: Ustilaginomycotina Doweld Class: Exobasidiomycetes Begerow, M. Stoll & R. Bauer Order: Exobasidiales Henn. Family: Brachybasidiaceae Gäum. 55. Meira argovae Boekhout, Scorzetti, Gerson & Sztejnb. ex Denchev & T. Denchev, Mycobiota 11: 3 (2021), (first record in Thailand) 56. Meira plantarum Q.M. Wang, Y.Y. Li, M. Groenew. & M.M. Wang, Frontiers Microbiol. 12: 16 (2022), (first record in Thailand) 57. Meira chiangmaiensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 58. Meira limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 59. Meira pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Family: Laurobasidiaceae Pinruan, Sommai, Suetrong, Somrith. & E.B.G. Jones 60. Laurobasidium hachijoense (Y. Otani, Kakish. & Iijima) Kakish., Nagao & Denchev, Phytotaxa 303: 97 (2017) Order: Microstromatales R. Bauer & Oberw. Families Incertae sedis 61. Jaminaea lantanae Q.M. Wang, Y.Y. Li, M. Groenew. & M.M. Wang, Frontiers Microbiol. 12: 22 (2022), (first record in Thailand) 62. Parajaminaea hydei Kodchasee, Senwanna, J. Kumla & N. Suwannarach, sp. nov. 63. Sympodiomycopsis europaea Q.M. Wang, Y.Y. Li, M. Groenew. & F.Y. Bai, Frontiers Microbiol. 12: 22 (2022), (first record in Thailand) 64. Sympodiomycopsis paphiopedili Sugiy., Tokuoka & Komag., in Sugiyama, Tokuoka, Suh, Hirata & Komagata, Antonie van Leeuwenhoek 59: 101 (1991), (first record in Thailand) 65. Sympodiomycopsis hydei Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 66. Sympodiomycopsis limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 67. Sympodiomycopsis saisamorniae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. Class: Ustilaginomycetes Warm. Order: Ustilaginales Bek. Family: Ustilaginaceae Tul. & C. Tul. 68. Anthracocystis heteropogonicola (Mundk. & Thirum.) McTaggart & R.G. Shivas, Persoonia 29: 123 (2012), (first record in Thailand) 69. Moesziomyces antarcticus (Goto, Sugiy. & Iizuka) Q.M. Wang, Begerow, F.Y. Bai & Boekhout, Stud. Mycol. 81: 81 (2015) 70. Moesziomyces bullatus (J. Schröt.) Vánky, Bot. Notiser 130: 133 (1977) 71. Moesziomyces parantarcticus (Sugita, M. Takash., Mekha & Poonwan) Q.M. Wang, Begerow, F.Y. Bai & Boekhout, Stud. Mycol. 81: 81 (2015) 72. Pseudozyma chiangmaiensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 73. Pseudozyma lannaensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 74. Pseudozyma limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 75. Pseudozyma pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. 76. Pseudozyma saisamorniae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. -
Fresh flower samples were collected from Chiang Mai, Chiang Rai, and Phayao Provinces, Thailand between July to October 2024 (Figs 1, 2 and Table 2). Each fresh flower sample was placed in a sterile plastic bag, put on ice and transported to the laboratory within 24 h before sample preparation and subsequent yeast isolation. The excised internal parts of the flowers including carpel stamen, pollen, and nectary were soaked in 5 ml of sterilized 0.85% (w/v) NaCl for 15 min. Then, solution was spread onto yeast malt agar (YMA) plates (1.0% glucose, 0.5% peptone, 0.3% malt extract, 0.3% yeast extract, and 2.0% agar) supplemented with 50 mg/L chloramphenicol, and incubated at 25 °C for 3 d. Each yeast strain was subcultured on YMA, and deposited at −80 °C in the Sustainable Development of Biological Resources (SDBR) Laboratory's Culture Collection within the Faculty of Science at Chiang Mai University, Thailand, the Thailand Bioresource Research Center (TBRC), Thailand, and Guizhou Medical University Culture Collection (GMBCC), Guiyang, China. The holotype of new taxa was permanently preserved in a metabolically inactive state in the Chiang Mai University Biology Department's Herbarium (CMUB), Chiang Mai University, Chiang Mai, Thailand.
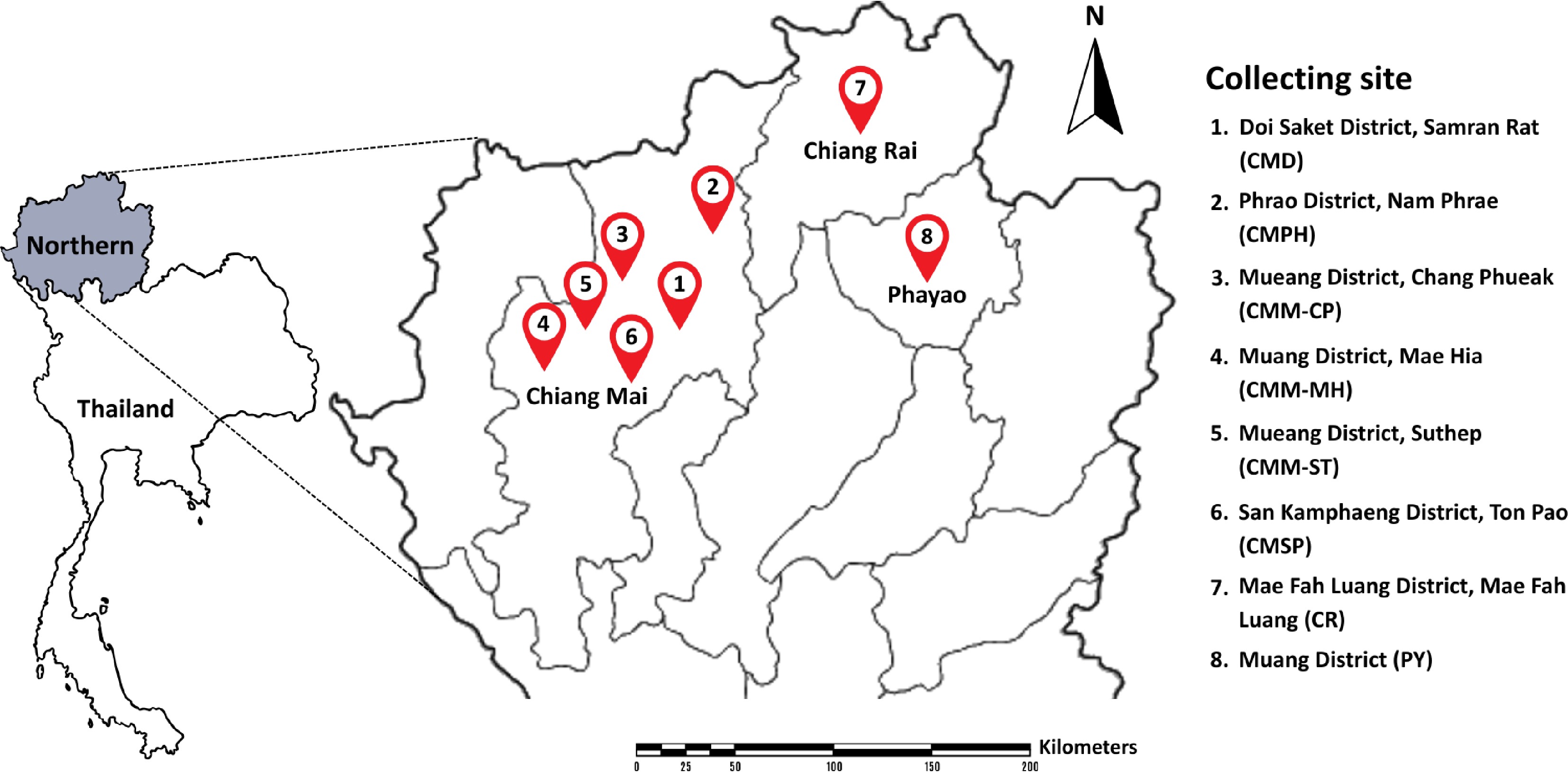
Figure 1.
Location of sampling sites in northern Thailand.

Figure 2.
(a)–(l) Some flower samples collected from northern Thailand, and their (m)–(x) internal parts. (a) Angelonia goyazensis, (b) Bellis perennis, (c) Bougainvillea hybrid, (d) Ixora chinensis, (e) Jatropha integerrima, (f) Melampodium divaricatum, (g) Plumeria rubra, (h) Portulaca grandiflora, (i) Rosa sp., (j) Salvia farinacea, (k) Senna spectabilis, (l) Syzygium jambos. The internal parts of (m) Antigonon leptopus, (n) Citrus japonica, (o) Exacum affine, (p) Ixora chinensis, (q) Jasminum sambac, (r) Nerium oleander, (s) Passiflora caerulea, (t) Portulaca grandiflora, (u) Rosa sp., (v) Salvia farinacea, (w) Senna spectabilis, and (x) Syzygium jambos. Scale bars (a)–(l) = 1 cm, and (m)–(x) = 1 mm.
Table 2. Location of sampling sites and flower species used in this study.
Location sites Flower species Chiang Mai Province Doi Saket District, Samran Rat (CMD) Combretum indicum Phrao District, Nam Phrae (CMPH) Boesenbergia rotunda, Cananga odorata, Canna indica, Crinum asiaticum, Globba winitii, Ixora sp., Musa sapientum, Oncidium sp., Phyllanthus pulcher, Syzygium jambos, Zamioculcas zamiifolia, and Zephyranthes minuta Mueang District, Chang Phueak (CMM-CP) Adenium obesum, Angelonia goyazensis, Antigonon leptopus, Argyranthemum frutescens, Bidens pilosa, Capsicum sp., Citrus japonica, Dolichandrone serrulata, Exacum affine, Jasminum sambac, Mecardonia procumbens, Melampodium divaricatum, Millingtonia hortensis, Murraya paniculata, Nerium oleander, Physalis minima, Plumbago auriculata, Plumeria obtusa, Portulaca grandiflora, Rosa sp., and Salvia farinacea Mueang District, Mae Hia (CMM-MH) Allamanda cathartica, Curcuma sessilis, Hamelia patens, Plumeria rubra, Ruellia tuberosa, and Tecoma stans Mueang District, Suthep (CMM-ST) Alstonia scholaris, Bougainvillea hybrid, Carmona retusa, Ixora chinensis, Passiflora caerulea, Senna spectabilis, and Tectona grandis San Kamphaeng District, Ton Pao (CMSP) Lagerstroemia speciosa, Momordica charantia, and Morinda citrifolia Chiang Rai Province Mae Fah Luang District, Mae Fah Luang (CR) Pentas lanceolata and Thunbergia erecta Phayao Province Mueang District (PY) Catharanthus roseus, Cnidoscolus aconitifolius, Dichorisandra thyrsiflora, Hibiscus rosa-sinensis, Jatropha integerrima, Melampodium divaricatum, Nerium oleander, Ocimum tenuiflorum, Pachystachys lutea, Plumeria pudica, and Thryallis glauca Morphological characterization
-
The morphological characteristics were examined according to the standard methods described by Kurtzman[27], de Vega et al.[11], and Shibayama et al.[13]. Colony morphology of each yeast strain was examined on YMA after incubation at 25 °C for 5 d. The formation of pseudohyphae and true hyphae was assessed through slide culture on potato dextrose agar (PDA) incubated at 25 °C for one month. Ascospore formation was investigated for individual strains and strain pairs on corn meal agar (CMA), PDA, 5% malt extract agar (5% malt extract and 1.5% agar; 5% MEA), V8 agar, and YMA at 25 °C for one month. Micromorphological characteristics were observed using a light microscope (Nikon Eclipse Ni-U, Tokyo, Japan). The sizes of structures such as cells, pseudohyphae, and true hyphae were measured using the Tarosoft® Image Framework program, based on at least 50 measurements for each structure.
Biochemical and physiological characterization
-
All strains of the new yeast taxa were characterized biochemically and physiologically, according to the standard methods described by Kurtzman[27]. Carbon and nitrogen source assimilation tests were conducted in liquid medium. Fermentation of carbohydrates was carried out in a liquid medium using Durham fermentation tubes. Cycloheximide resistance was also performed in a liquid medium. Acid production and urea hydrolysis were investigated using solid media. The effect of temperature on yeast growth was determined by using YMA at various temperatures ranging from 10 to 40 °C. The ability to grow in media of high osmotic pressure was performed using 50% and 60% glucose agar, and 10% and 16% sodium chloride (NaCl) plus 5% glucose medium. The physiological status was used to normalize and compare results across previously published data with varying scales. Positive carbon fermentation was indicated by '+', whereas negative fermentation was indicated by '–'. For carbon and nitrogen assimilations and growth characteristics, '–' indicated no growth, 'w' weak growth, 's' slow growth, 'l' latent growth, 'v' variable growth, '+' strong growth, and 'nd' not determined.
Molecular study
-
Yeast strains were cultivated in 5 mL of yeast malt extract broth (YMB) in 18 × 180 mm test tubes, incubated at 25 °C with shaking at 110 rpm for 48 h. Yeast cells were harvested by centrifugation at 11,000 rpm, and washed three times with sterile distilled water. Genomic DNA was extracted from yeast cells using a DNA Extraction Mini Kit (FAVORGEN, Taiwan, China) following the manufacturer's protocol. Amplification of the D1/D2 domain of the LSU was carried out by PCR with the forward primer NL1, and reverse primer NL4[28]. The ITS region was amplified with the forward primer ITS1, and reverse primer ITS4[29], and SSU was amplified with the forward primer NS1 and reverse primer NS4, the process previously described by Kumla et al.[23]. Additionally, the largest subunit of RNA polymerase II (rpb1) was amplified using the forward primer RPB1-Af, and reverse primer RPB1-Cr; the second-largest subunit (rpb2) was amplified using the forward primer fRPB2-5F, and reverse primer fRPB2-7cR; and the translation elongation factor 1 alpha (tef1-α) was amplified using the forward primer EF1-983F, and reverse primer EF1-2218R. Amplification of these three protein-coding genes was performed as described in Wang et al.[30]. PCR products were checked and then purified using a NucleoSpin Gel and PCR Clean-up Kit (Macherey-Nagel, Germany). The purified PCR products were directly sequenced at the 1st Base Company (Kembangan, Malaysia). The obtained sequences were used to query GenBank via BLAST (
http://blast.ddbj.nig.ac.jp/top-e.html , accessed on 25 August, 2024).Phylogenetic analyses
-
Sequence generated from this study were analyzed with the use of similarity searches retrieved from GenBank based on BLASTn searches of the NCBI nucleotide database (
http://blast.ncbi.nlm.nih.gov ). Reference sequences were selected based on BLAST results and recent publications. Sequence alignments were aligned with MAFFT v.7 (http://mafft.cbrc.jp/alignment/server , accessed on 27 August, 2024)[31], and the alignments were improved where necessary in BioEdit V.7.0.9.1[32]. Maximum likelihood (ML), and Bayesian inference (BI) analyses were conducted using single-locus and concatenated datasets to generate phylogenetic trees.The ML tree analyses with 1,000 bootstrap iterations were conducted via the CIPRES Science Gateway platform[33], using RAxML-HPC v.8 on ACCESS (v.8.2.12)[34], and employing the GTRGAMMA model of evolution. For BI, analyses were performed with MrBayes v.3.2.6[35] to estimate Bayesian posterior probabilities (BYPP)[36,37] by Markov Chain Monte Carlo sampling (BMCMC). MrModeltest 2.3[38] was used to determine the model of nucleotide substitution for each locus. Two independent runs of four simultaneous Markov chains were executed for 1 to 60 million generations (depending on individual settings for the yeast group), trees sampled every 100 generations. When the average standard deviation of split frequencies dropped below 0.01, the analysis was terminated, and the first 25% of the trees, representing the burn-in phase, were discarded. The phylogenetic trees were visualized using FigTree v.1.4.3[39], and edited with Adobe Illustrator CC 2019 (version 23.0.3.585), and Adobe Photoshop CS6 (version 13.0) (Adobe Systems, USA). All sequences generated in this study were submitted to GenBank. All entries are depicted in the phylogenetic tree, along with their corresponding descriptions. New yeast taxa were registered in the MycoBank database[40].
-
Sixty-three flower samples from northern Thailand were used for the isolation of anthophilous yeasts. A total of 187 strains were identified to the species level, based on the threshold of more than 99% sequence identity with the type strain of a described species in ITS region or D1/D2 domain. The classification of taxa in this study follows the Outline of Fungi and Fungus-Like Taxa[41,42], and updated based on recent relevant literature. Based on morphology and multi-locus phylogeny, 89 strains were identified as 38 previous known yeast species. Details on their host, locality, and sequence accession numbers are provided in Table 3, and a phylogenetic tree of each species are presented in the Supplementary File 1. Whereas 98 strains were determined to represent undescribed species, including two with invalidly published names. They were classified into the phyla Ascomycota (22.46%), and Basidiomycota (77.54%), comprising 36 genera in 22 families (Fig. 3). Five families belonging to the phylum Ascomycota were identified, including Debaryomycetaceae (Candida, Kodamaea, Meyerozyma, and Priceomyces), Metschnikowiaceae (Metschnikowia), Phaffomycetaceae (Cyberlindnera), Saccharomycodaceae (Hanseniaspora), and Trichomonascaceae (Entelexis, Starmerella, and Wickerhamiella). While 15 families, and two incertae sedis belonging to the phylum Basidiomycota were identified, including Brachybasidiaceae (Meira), Bulleribasidiaceae (Hannaella and Vishniacozyma), Chionosphaeraceae (Boekhoutia), Cryptococcaceae (Kwoniella), Cystobasidiaceae (Cystobasidium and Halobasidium), Erythrobasidiaceae (Erythrobasidium), Filobasidiaceae (Filobasidium and Naganishia), Laurobasidiaceae (Laurobasidium), Microbotryomycetes genera incertae sedis (Curvibasidium), Microstromatales genera incertae sedis (Jaminaea, Parajaminaea, and Sympodiomycopsis), Rhynchogastremaceae (Papiliotrema), Sporidiobolaceae (Rhodosporidiobolus, Rhodotorula, and Sporobolomyces), Symmetrosporaceae (Symmetrospora), Thailandicolaceae (Thailandicola), Trichosporonaceae (Trichosporon), Trimorphomycetaceae (Saitozyma), and Ustilaginaceae (Acaromyces, Moesziomyces, and Pseudozyma) (Figs 3 and 4).
Table 3. Known yeast species obtained in this study, with their hosts, localities, and GenBank accession numbers.
Species Strain Host Locality GenBank accession number D1/D2 ITS rpb1 tef1-α Ascomycota, Saccharomycotina
Dipodascomycetes, Dipodascales, TrichomonascaceaeStarmerella etchellsii SDBR-CMU641 Passiflora caerulea Thailand, Chiang Mai Province, Mueang District, Suthep PV834436 PV834623 – – Pichiomycetes, Serinales, Debaryomycetaceae Candida tropicalis SDBR-CMU677 Melampodium divaricatum Thailand, Phayao Province, Mueang District PV834442 PV834627 – – Kodamaea ohmeri SDBR-CMU607 Boesenbergia rotunda Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834443 PV834628 – – SDBR-CMU626 Jasminum sambac Thailand, Chiang Mai Province Mueang District, Chang Phueak PV834444 PV834629 – – SDBR-CMU660 Catharanthus roseus Thailand, Phayao Province, Mueang District PV834445 PV834630 – – SDBR-CMU663 Catharanthus roseus Thailand, Phayao Province, Mueang District PV834446 PV834631 – – SDBR-CMU670 Pentas lanceolata Thailand, Phayao Province, Mueang District PV834447 PV834632 – – Kodamaea restingae SDBR-CMU640 Bougainvillea hybrid Thailand, Chiang Mai Province, Mueang District PV834448 PV834633 – – SDBR-CMU672 Crinum asiaticum Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834449 PV834634 – – Meyerozyma caribbica SDBR-CMU644 Pentas lanceolata Thailand, Chiang Rai Province, Mae Fah Luang District, Mae Fah Luang PV834450 PV834635 – – SDBR-CMU676 Melampodium divaricatum Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834451 PV834636 – – Metschnikowiaceae Metschnikowia cibodasensis SDBR-CMU554 Tecoma stans Thailand, Chiang Mai Province, Mueang District, Suthep PV834454 PV834639 – – Metschnikowia koreensis SDBR-CMU553 Plumeria obtusa Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834455 – – SDBR-CMU555 Syzygium jambos Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834456 PV834640 – – SDBR-CMU558 Allamanda cathartica Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834457 – – SDBR-CMU559 Allamanda cathartica Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834458 – – – SDBR-CMU625 Jasminum sambac Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834459 – – – SDBR-CMU643 Pentas lanceolata Thailand, Chiang Rai Province, Mae Fah Luang, Mae Fah Luang PV834460 – – – SDBR-CMU646 Cnidoscolus aconitifolius Thailand, Phayao Province, Mueang District PV834461 – – – SDBR-CMU649 Jatropha integerrima Thailand, Phayao Province, Mueang District PV834462 – – – SDBR-CMU651 Dichorisandra thyrsiflora Thailand, Phayao Province, Mueang District PV834463 – – – SDBR-CMU655 Pentas lanceolata Thailand, Chiang Rai Province, Mae Fah Luang, Mae Fah Luang PV834464 – – – SDBR-CMU657 Nerium oleander Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834465 – – – SDBR-CMU658 Nerium oleander Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834466 – – – SDBR-CMU669 Pentas lanceolata Thailand, Chiang Rai Province, Mae Fah Luang, Mae Fah Luang PV834467 – – – SDBR-CMU671 Bougainvillea hybrid Thailand, Chiang Mai Province, Mueang District, Suthep PV834468 – – – SDBR-CMU673 Ixora chinensis Thailand, Chiang Mai Province, Mueang District, Suthep PV834469 – – – SDBR-CMU674 Ixora chinensis Thailand, Chiang Mai Province, Mueang District, Suthep PV834470 – – – Saccharomycetes, Phaffomycetales, Phaffomycetaceae Cyberlindnera fabianii SDBR-CMU563 Rosa sp. Thailand, Chiang Mai Province, Mueang District, Chang Phueak, PV834471 PV834641 – – Saccharomycodales, Saccharomycodaceae Hanseniaspora lachancei SDBR-CMU560 Antigonon leptopus Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834472 PV834642 – – SDBR-CMU598 Antigonon leptopus Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834473 PV834643 – – Basidiomycota, Agaricomycotina
Tremellomycetes, Filobasidiales, FilobasidiaceaeNaganishia albida SDBR-CMU556 Nerium oleander Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834476 PV834646 – – Naganishia diffluens SDBR-CMU712 Argyranthemum frutescens Thailand, Chiang Mai Province, Mueang District, Chang Phueak, PV834477 PV834647 – – SDBR-CMU716 Argyranthemum frutescens Thailand, Chiang Mai Province, Mueang District, Chang Phueak, PV834478 PV834648 – – Naganishia liquefaciens SDBR-CMU609 Rosa sp. Thailand, Chiang Mai Province, Mueang District, Chang Phueak, PV834479 PV834649 – – Tremellales, Bulleribasidiaceae Hannaella pagnoccae SDBR-CMU576 Curcuma sessilis Thailand, Chiang Mai Province, Mueang District, Mae Hia PV834480 PV834650 – – SDBR-CMU653 Dichorisandra thyrsiflora Thailand, Phayao Province, Mueang District PV834481 PV834651 – – SDBR-CMU683 Plumeria pudica Thailand, Phayao Province, Mueang District PV834482 PV834652 – – SDBR-CMU688 Plumeria pudica Thailand, Phayao Province, Mueang District PV834483 PV834653 – – Hannaella phyllophila SDBR-CMU583 Oncidium sp. Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834484 PV834654 – – Vishniacozyma marinae SDBR-CMU692 Plumeria pudica Thailand, Phayao Province, Mueang District PV834485 PV834655 – – Cryptococcaceae Kwoniella bestiolae SDBR-CMU642 Passiflora caerulea Thailand, Chiang Mai Province, Mueang District, Suthep PV834488 PV834658 – – SDBR-CMU699 Bougainvillea hybrid Thailand, Chiang Mai Province, Mueang District, Suthep PV834489 PV834659 – – Kwoniella heveanensis SDBR-CMU633 Tectona grandis Thailand, Chiang Mai Province, Mueang District, Suthep PV834490 PV834660 – – SDBR-CMU726 Alstonia scholaris Thailand, Chiang Mai Province, Mueang District, Suthep PV834491 PV834661 – – Rhynchogastremaceae Papiliotrema aspenensis SDBR-CMU652 Plumeria pudica Thailand, Phayao Province, Mueang District PV834496 PV834666 – – Papiliotrema flavescens SDBR-CMU668 Pachystachys lutea Thailand, Phayao Province, Mueang District PV834499 PV834669 – –
Trichosporonales, TrichosporonaceaeTrichosporon asahii SDBR-CMU549 Bidens Pilosa Thailand, Chiang Mai Province, Mueang District, Chang Phuea PV834507 PV834677 – – SDBR-CMU656 Nerium oleander Thailand, Phayao Province, Mueang Districtin PV834508 PV834678 – – Pucciniomycotina
Cystobasidiomycetes, Cystobasidiales, CystobasidiaceaeCystobasidium benthicum SDBR-CMU720 Lagerstroemia speciosa Thailand, Chiang Mai Province, San Kamphaeng District, Ton Pao PV834511 PV834681 – – Cystobasidium keelungense SDBR-CMU585 Mecardonia procumbens Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834512 PV834682 – – SDBR-CMU713 Argyranthemum frutescens Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834513 PV834683 – – Cystobasidium minutum SDBR-CMU608 Rosa sp. Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834514 PV834684 – – SDBR-CMU721 Lagerstroemia speciosa Thailand, Chiang Mai Province, San Kamphaeng District, Ton Pao PV834515 PV834685 – – Cystobasidiomycetes, Erythrobasidiales, Erythrobasidiaceae Erythrobasidium primogenitum SDBR-CMU689 Plumeria pudica Thailand, Phayao Province, Mueang District PV834525 PV834695 – –
Microbotryomycetes, Sporidiobolales, SporidiobolaceaeRhodosporidiobolus fluvialis SDBR-CMU562 Rosa sp. Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834532 PV834702 – – SDBR-CMU568 Citrus japonica Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834533 PV834703 – – SDBR-CMU659 Nerium oleander Thailand, Phayao Province, Mueang District PV834534 PV834704 – – Rhodosporidiobolus ruineniae SDBR-CMU593 Zamioculcas zamiifolia Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834535 PV834705 – – SDBR-CMU665 Ocimum tenuiflorum Thailand, Phayao Province, Mueang District PV834536 PV834706 – – SDBR-CMU724 Combretum indicum Thailand, Chiang Mai Province, Doi Saket District, Samran Rat PV834537 PV834707 – – Rhodotorula paludigena SDBR-CMU580 Exacum affine Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834538 PV834708 – – Rhodotorula toruloides SDBR-CMU678 Melampodium divaricatum Thailand, Phayao Province, Mueang District PV834542 PV834712 – –
Ustilaginomycotina
Exobasidiomycetes, Exobasidiales, BrachybasidiaceaeMeira argovae SDBR-CMU709 Morinda citrifolia Thailand, Chiang Mai Province, San Kamphaeng District, Ton Pao PV834546 PV834716 – – SDBR-CMU729 Alstonia scholaris Thailand, Chiang Mai Province Mueang District, Suthep PV834547 PV834717 – – SDBR-CMU730 Alstonia scholaris Thailand, Chiang Mai Province Mueang District, Suthep PV834548 PV834718 – – Meira plantarum SDBR-CMU687 Plumeria pudica Thailand, Phayao Province, Mueang District PV834555 PV834725 – –
Exobasidiales, LaurobasidiaceaeLaurobasidium hachijoense SDBR-CMU615 Syzygium jambos Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834559 PV834729 – – SDBR-CMU616 Syzygium jambos Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834560 PV834730 – – Microstromatales, Microstromatales genera incertae sedis Jaminaea lantanae SDBR-CMU693 Hibiscus rosa-sinensis Thailand, Phayao Province, Mueang District PV834561 PV834731 – – Sympodiomycopsis europaea SDBR-CMU605 Plumeria obtusa Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834564 PV834734 PV941870 PV947475 SDBR-CMU637 Bougainvillea hybrid Thailand, Thailand, Chiang Mai Province, Mueang District, Suthep PV834565 PV834735 – – Sympodiomycopsis paphiopedili SDBR-CMU567 Hamelia patens Thailand, Chiang Mai Province, Mueang District, Mae Hia PV834572 PV834742 – – SDBR-CMU708 Morinda citrifolia Thailand, Chiang Mai Province, San Kamphaeng District, Ton Pao PV834573 PV834743 – – Ustilaginomycetes, Ustilaginales, Ustilaginaceae Anthracocystis heteropogonicola SDBR-CMU551 Bidens pilosa Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834576 PV834746 – – SDBR-CMU589 Antigonon leptopus Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834577 PV834747 – – Moesziomyces antarcticus SDBR-CMU548 Zephyranthes minuta Thailand, Chiang Mai Province, Phrao District, Nam Phrae PV834578 PV834748 – – SDBR-CMU634 Senna spectabilis Thailand, Chiang Mai Province, Mueang District, Suthep PV834579 PV834749 – – SDBR-CMU647 Hibiscus rosa-sinensis Thailand, Phayao Province, Mueang District PV834580 PV834750 – – SDBR-CMU706 Momordica charantia Thailand, Chiang Mai Province, San Kamphaeng District, Ton Pao PV834581 PV834751 – – Moesziomyces bullatus SDBR-CMU571 Citrus japonica Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834582 PV834752 – – SDBR-CMU582 Exacum affine Thailand, Chiang Mai Province, Mueang District, Chang Phueak PV834583 PV834753 – – SDBR-CMU645 Cnidoscolus aconitifolius Thailand, Phayao Province, Mueang District PV834584 PV834754 – – SDBR-CMU675 Melampodium divaricatum Thailand, Phayao Province, Mueang District PV834585 PV834755 – – SDBR-CMU679 Cnidoscolus aconitifolius Thailand, Phayao Province, Mueang District PV834586 PV834756 – – Moesziomyces parantarcticus SDBR-CMU654 Pentas lanceolata Thailand, Phayao Province, Mueang District PV834587 PV834757 – – SDBR-CMU733 Ixora chinensis Thailand, Chiang Mai Province, Mueang District, Suthep PV834588 PV834758 – – SDBR-CMU734 Ixora chinensis Thailand, Chiang Mai Province, Mueang District, Suthep PV834589 PV834759 – – SDBR-CMU732 Ixora chinensis Thailand, Chiang Mai Province, Mueang District, Suthep PV834590 PV834760 – – '−', undetermined sequence. 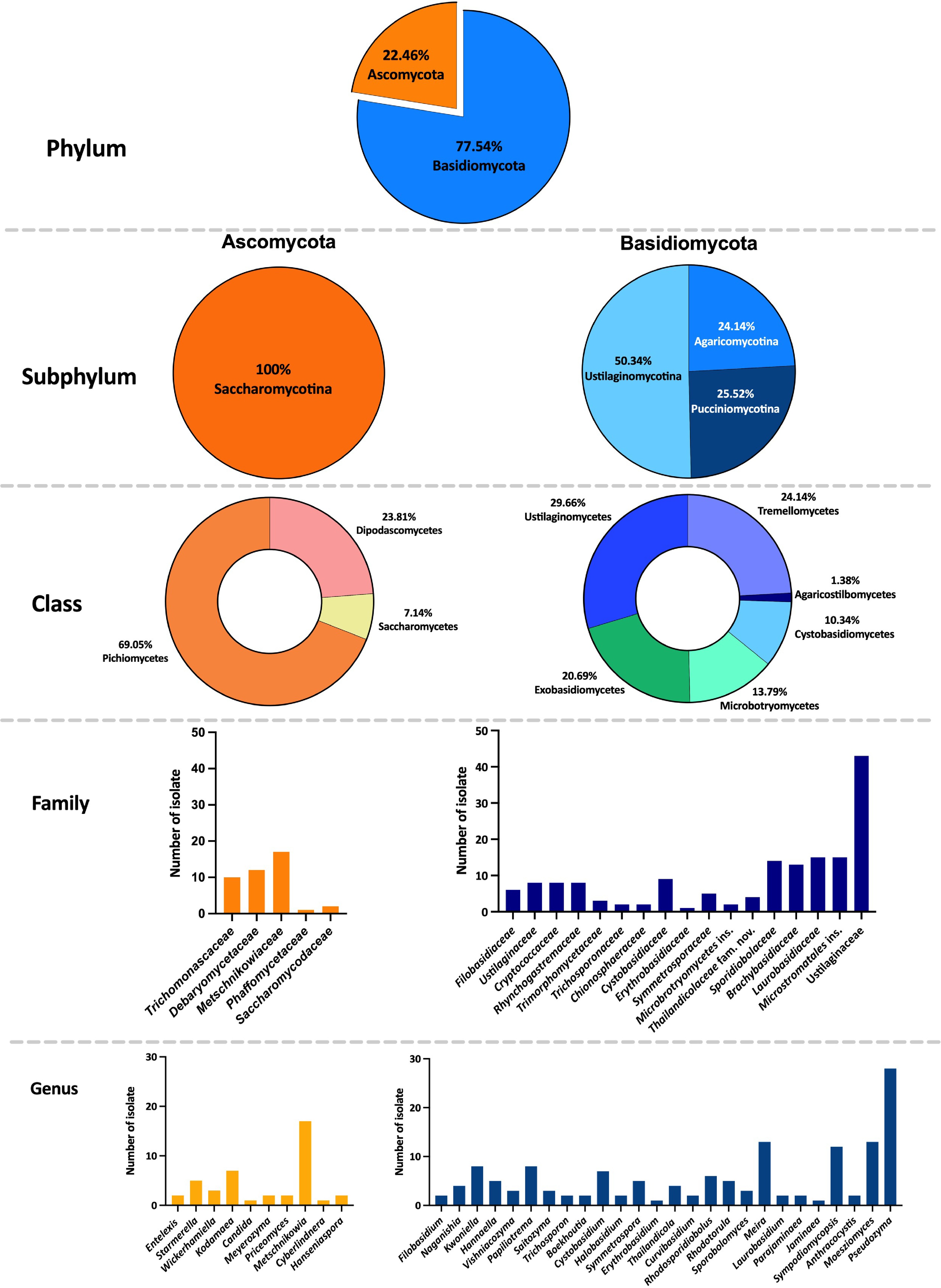
Figure 3.
Classification of anthophilous yeasts isolated from flowers collected in northern Thailand in this study.
The distribution of each yeast species is presented in Fig. 4. The majority of isolated anthophilous yeasts had low to moderate frequency of occurrence (1% to 3%), represented by blue to light green shades, with only a few taxa exceeding 5% frequency of occurrence (Fig. 4a). The most frequent ascomycetous yeast was Metschnikowia koreensis (8.56% frequency of occurrence), which was found in 11 flower samples in five site locations, and the most frequent basidiomycetous yeast was Pseudozyma saisamorniae (4.28% frequency of occurrence), found in eight flower samples presented at three site locations. Other species were found with frequencies of occurrence ranging from 0.53% to 2.67% in each flower species. In addition, Plumeria pudica (PY) harbored the highest number of yeast species, with nine species recorded, followed by Antigonon leptopus (CMM-CP), Curcuma sessilis (CMM-MH), and Tectona grandis (CMM-ST), each of which contained six yeast species (Fig. 4b).
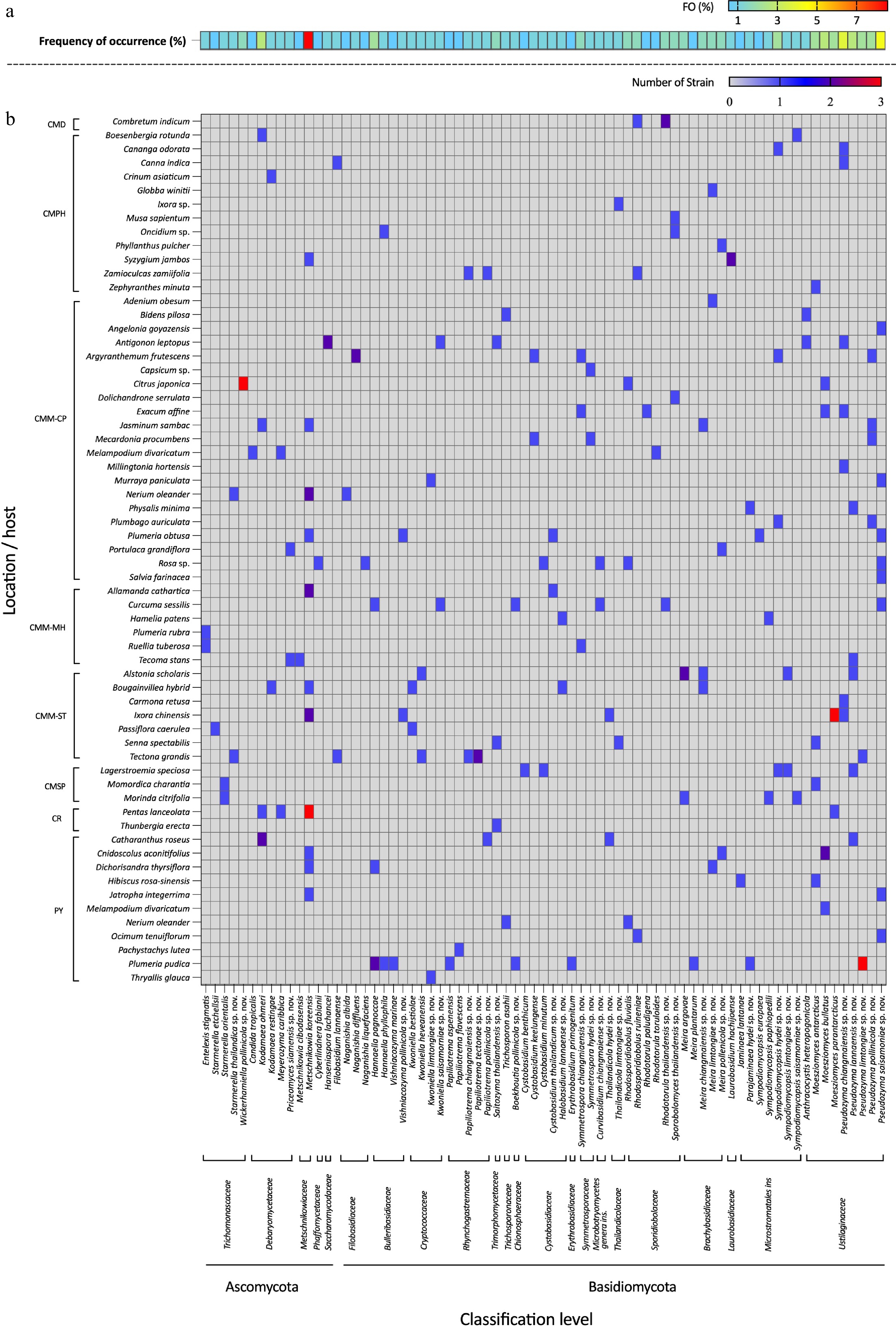
Figure 4.
Heatmap showing the consensus species diversity of anthophilous yeasts isolated from flowers collected in northern Thailand. (a) Frequency of occurrence (FO %) = number of samples, (b) where a particular species was observed, as a proportion of the total number of samples.
Additionally, 38 known species (Table 3), and two newly validated species (E. stigmatis and S. orientalis) were recorded for the first time from flower species. Notably, Cys. benthicum was newly recorded from a plant habitat. Among these, 14 species were recorded for the first time in Thailand including An. heteropogonicola, Cys. benthicum, Cys. keelungense, E. stigmatis, Er. primogenitum, H. lachancei, J. lantanae, Kw. bestiolae, Me. cibodasensis, Mei. argovae, Mei. plantarum, S. orientalis, Sym. europaea, and Sym. paphiopedili, the detail is listed in the Supplementary File 1.
Taxonomy
-
Phylum: Ascomycota Caval.-Sm.
Subphylum: Saccharomycotina O.E. Erikss. & Winka
Class: Dipodascomycetes M. Groenew., Hittinger, Opulente & A. Rokas
Order: Dipodascales M. Groenew., Hittinger, Opulente & A. Rokas
Family: Trichomonascaceae Kurtzman & Robnett
Trichomonascaceae are found in a wide range of habitats, some with ecological distribution patterns indicating close interactions with insects, nectar, and decaying plant matter. Furthermore, species in Trichomonascaceae are economically important in industries (such as food production and cosmetics), medicine, and agriculture. Members of this family typically pyriform to oval in shape, and while some genera form septate hyphae, Wickerhamiella does not[43,44]. Currently, 12 genera are listed in this family including Blastobotrys, Crinitomyces, Deakozyma, Diddensiella, Groenewaldozyma, Limtongella, Spencermartinsiella, Starmerella, Sugiyamaella, Trichomonascus, Wickerhamiella, and Zygoascus[41]. In this study, five yeast species were presented, including Entelexis stigmatis (two strains), Starmerella etchellsii (one strain), S. orientalis (two strains), S. thailandica sp. nov. (two strains), and Wickerhamiella pollinicola sp. nov. (three strains).
Entelexis stigmatis Sipiczki ex Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Supplementary File 1: Fig. S1a)
MycoBank number: MB 860217
≡ Starmerella stigmatis (Sipiczki) ex C.A. Rosa & Lachance, Int. J. Syst. Evol. Microbiol. 68(4): 1342 (2018)
≡ Candida stigmatis Sipiczki. FEMS Yeast Res. 10(3): 364 (2010)
Description – Sipiczki[45]
Holotype – INDIA, Hyderabad District, flower of Magnolia sp., 2007, sin. coll., CBS 11464 holotype, preserved in a metabolically inactive state; isotype CCY 29-179-1 = CBS 11462.
(Candida stigmatis Sipiczki, FEMS Yeast Res. 10[3]: 364 [2010] [nom. inval., Melbourne Code, Art., 40.7]), Starmerella stigmatis C.A. Rosa & Lachance, in Santos, Leon, Barros, Freitas, Hughes, Morais, Lachance & Rosa, Int. J. Syst. Evol. Microbiol. 68[4]: 1342 [2018] [nom. inval., Melbourne Code, Art., 40.7])
Material examined – THAILAND, Chiang Mai Province, Mueang District, Mae Hia in watrakanu flower (Ruellia tuberosa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU591, Chang Phueak, in frangipani flower (Plumeria rubra), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU604. GenBank numbers SDBR-CMU591: PV834437 (D1/D2); SDBR-CMU604: PV834438 (D1/D2).
Ecology and distribution – Magnolia sp. flower in India[45], and flowers of Plumeria rubra and Ruellia tuberosa in Thailand (this study).
Notes – Phylogenetic analyses based on combined D1/D2 domain and ITS sequence data (Fig. 5) showed that Entelexis forms a distinct clade, separate from Starmerella, which is concurred with the results of previous studies[46]. Entelexis stigmatis strains SDBR-CMU591 and SDBR-CMU604 were clustered with the type of species (CBS 11464), and another representative strain (GY9L07). A comparison of D1/D2 domain and ITS indicates that the present strains are not significantly different from E. stigmatis (only one and four differentiated nucleotides, respectively). However, the species of E. stigmatis is currently considered Nom. inval. under Art. 40.7 of the Melbourne Code as listed in Index Fungorum[47]. Therefore, E. stigmatis is validated as a member of Entelexis by providing a registration identifier and presenting corrected type citations, along with references to the original descriptions. The morphological characteristics of the yeast strains in this study are similar to those previously reported, which were identified as E. stigmatis. Additionally, we hereby represent its first isolation from flowers of Plumeria rubra and Ruellia tuberosa, and new geological distribution in Thailand.
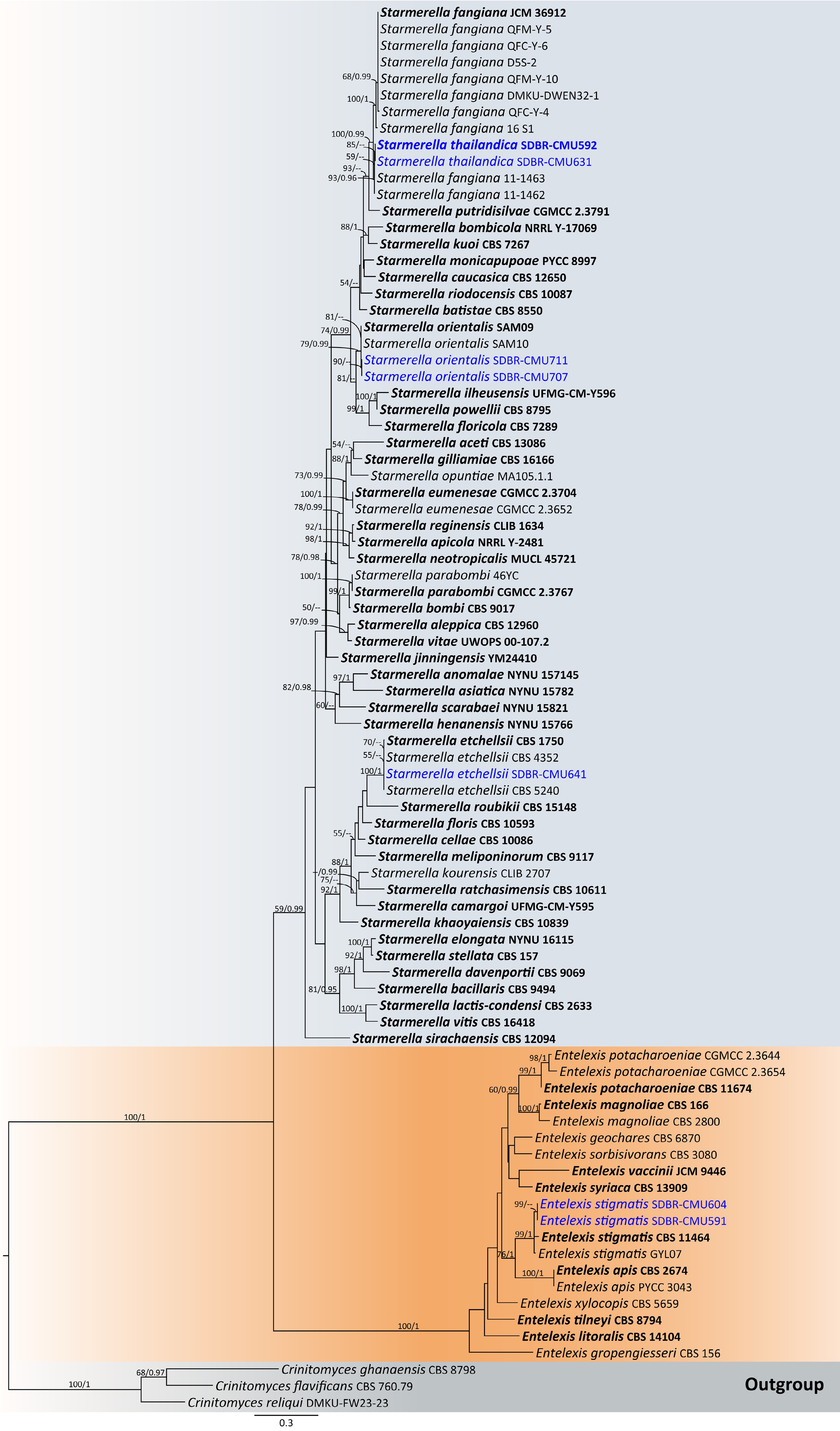
Figure 5.
Phylogram generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing genera Entelexis and Starmerella in Trichomonascaceae. The tree is rooted to Crinitomyces flavificans (CBS 760.79), C. ghanaensis (CBS 8798), and C. reliqui (DMKU-FW23-23). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Eighty-seven strains are included in the combined sequence analysis, which comprise 1,111 characters with gaps. Bootstrap support values for maximum likelihood (≥ 50%, ML, left) and Bayesian posterior probabilities (≥ 0.95, PP, right) are indicated above the nodes. Double dashes (--) denote support values below 50% ML and 0.95 PP. The scale bar represents 0.3 nucleotide substitutions per site. Ex-type strains are shown in bold, and sequences generated in this study are highlighted in blue.
Starmerella orientalis (Alimad., Soudi, F.Y. Bai, S.A. Wang & Q.M. Wang) ex Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Supplementary File 1: Fig. S1c)
MycoBank number: MB860216
Description – Alimadadi et al.[48]
Holotype – IRAN, East Azerbaijan Province, flower of Salsola sp., October 2012, sin. coll., SAM09 holotype, preserved in a metabolically inactive state; isotype IBRC-M 30204 = CBS 14142.
(Starmerella orientalis Alimad., Soudi, F.Y. Bai, S.A. Wang & Q.M. Wang, Int. J. Syst. Evol. Microbiol. 66[3]: 1478 [2016] [nom. inval., Shenzhen Code, Art., 40.7])
Material examined – THAILAND, Chiang Mai Province, San Kamphaeng District, Ton Pao, in balsum pear flower (Momordica charantia), August 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU707, Indian mulberry flower (Morinda citrifolia), August 2024, P. Kodchasee living culture SDBR-CMU711. GenBank numbers SDBR-CMU707: PV834432 (D1/D2), PV834619 (ITS); SDBR-CMU711: PV834433 (D1/D2), PV834620 (ITS).
Ecology and distribution – Salsola sp. flower in Iran[48], and flowers of Momordica charantia and Morinda citrifoli in Thailand (this study).
Notes – Starmerella orientalis was introduced by Alimadadi et al.[48] based on physiological characteristics and phylogeny of rRNA gene sequences. However, the species is considered Nom. inval., Art. 40.7 (Shenzhen) in Index Fungorum[47]. Phylogenetic analyses of a combined D1/D2 domain, and ITS sequence dataset (Fig. 5) show that the present sequence strains, SDBR-CMU707 and SDBR-CMU711, form a clade, clustering with S. orientalis with 78% MLBS and 0.99 BYPP support value. A comparison of D1/D2 domain and ITS region shows that the present strains, SDBR-CMU707 and SDBR-CMU711, are not significantly different from S. orientalis (only one nt substitutions in both regions). Therefore, S. orientalis is validated as a member of Starmerella by providing a registration identifier and presenting corrected type citations, along with references to the original descriptions. In addition, morphological comparison reveals that the present yeast strains correspond to previously characterized strains of S. orientalis[48], representing a new geographical distribution in Thailand, and marking the first isolation from the flowers of Momordica charantia and Morinda citrifolia.
Starmerella thailandica Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 6)
MycoBank number: MB860170
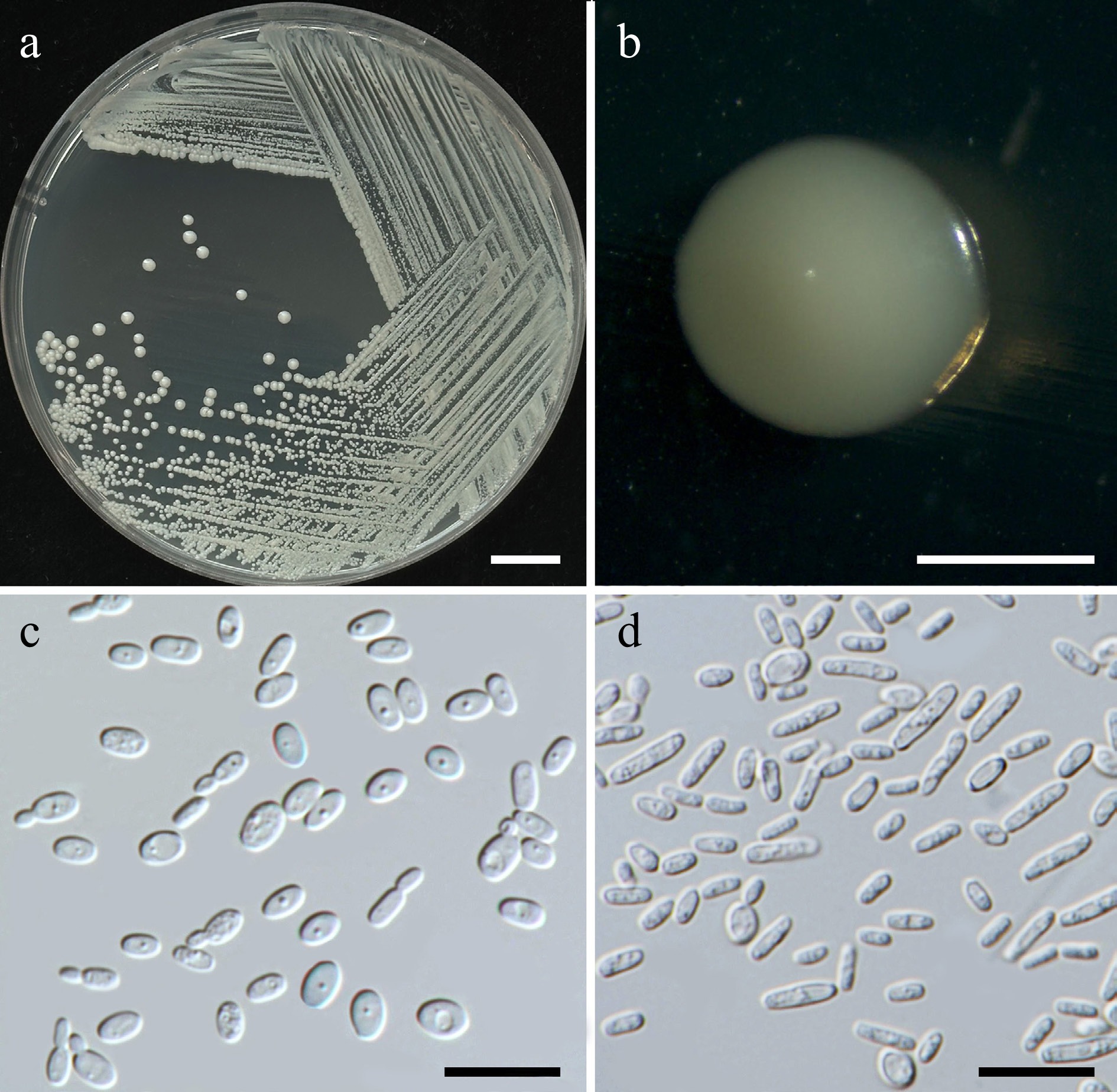
Figure 6.
Morphological characteristics of Starmerella thailandica (SDBR-CMU592, ex-type). (a) Culture, (b) single colony, (c) budding cells, on YMA at 25 °C for 5 d, and (d) elongated cells on PDA at 25 °C after 2 weeks. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The species name 'thailandica' refers to Thailand, the country where the type strain was isolated.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak in oleander flower (Nerium oleander), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype, CMUB40094 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU592 = GMBCC2395 = TBRC21393. GenBank numbers PV834434 (D1/D2), PV834621 (ITS), PX582293 (rpb1), PX582313 (rpb2).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.5–2 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ovoid to ellipsoidal (1.92–3.47 × 3.44–4.92 μm, n = 50), budding is polar. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Ascospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, sorbose, ribose, xylose, sucrose, raffinose, glycerol, D-glucitol, D-mannitol, D-glucono-1,5-lactone, D-gluconate, succinate, citrate, ethanol, and xylitol are assimilated, but N-acetyl glucosamine, L-arabinose, D-arabinose, L-rhamnose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose, melizitose, inulin, soluble starch, erythritol, ribitol, galactitol, myo-inositol, D-glucuronate, D-galacturonic acid, DL-lactate, and methanol are not assimilated. Ammonium sulfate, ethylamine, L-lysine (weak) are assimilated as sole nitrogen source, but potassium nitrate, and sodium nitrite are not assimilated. Cadaverine is variable. Growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose, and 16% NaCl/5% glucose. No growth occurs on media containing, 0.01% cycloheximide and 0.1% cycloheximide. Urease reaction and acid formation are negative. Growth at 10, 20, 25, 35, 37 °C, but not at 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), August 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU631. GenBank numbers PV834435 (D1/D2), PV834625 (ITS).
Notes – Phylogenetic analyses based on a combined dataset of the D1/D2 domain and ITS sequences revealed that S. thailandica (SDBR-CMU592 and SDBR-CMU631) is closely related to S. fangiana strains 11-1462 and 11-1463. Together, they form a sister clade to S. fangiana, including strains JCM36912 (type strain), QFM-Y-5, QFM-Y-6, D5S-2, QFM-Y-10, DMKU-DWEN32-1, QFM-Y-6, and 16S1[49], with 100% BSML and 0.99 BYPP support (Fig. 5). Starmerella thailandica and S. fangiana strains 11-1462 and 11-1463 differ 2 nucleotide substitutions in the ITS region, respectively, suggesting that they may represent the same species. However, further investigation, including detailed morphological and physicochemical characterization of S. fangiana strains 11-1462 and 11-1463, is required. In contrast, S. thailandica SDBR-CMU592 and SDBR-CMU631 differ from S. fangiana clade (including type strain) by 2 nucleotide substitutions and 1 gap (0.22%) in the D1/D2 domain, and by 10–25 nucleotide mismatches (~2.40%–5.84%), including substitutions and deletions, in the ITS region. Likewise, S. fangiana strains 11-1462 and 11-1463 differ from S. fangiana clade by 2 nucleotide substitutions, and 1 gap (0.22%) in the D1/D2 domain, and by 7–25 nucleotide mismatches (~1.88%–5.85%) in the ITS region. Based on the ITS region mismatches, S. thailandica and S. fangiana strains 11-1462 and 11-1463 represent distinct taxa from S. fangiana. Starmerella thailandica can be distinguished from S. fangiana JCM36912 by its ability to assimilate xylose, L-arabinose, D-glucono-1,5-lactone, D-gluconate, and creatine, as well as by its growth in 16% NaCl/5% glucose medium. Moreover, S. fangiana was able to grow at 40 °C, whereas S. pollenicalo cannot[49]. Based on ITS sequence data and phenotypic characteristics, strains SDBR-CMU592 and SDBR-CMU631 are therefore proposed to represent a novel Starmerella species.
Wickerhamiella pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 7)
MycoBank number: MB860171
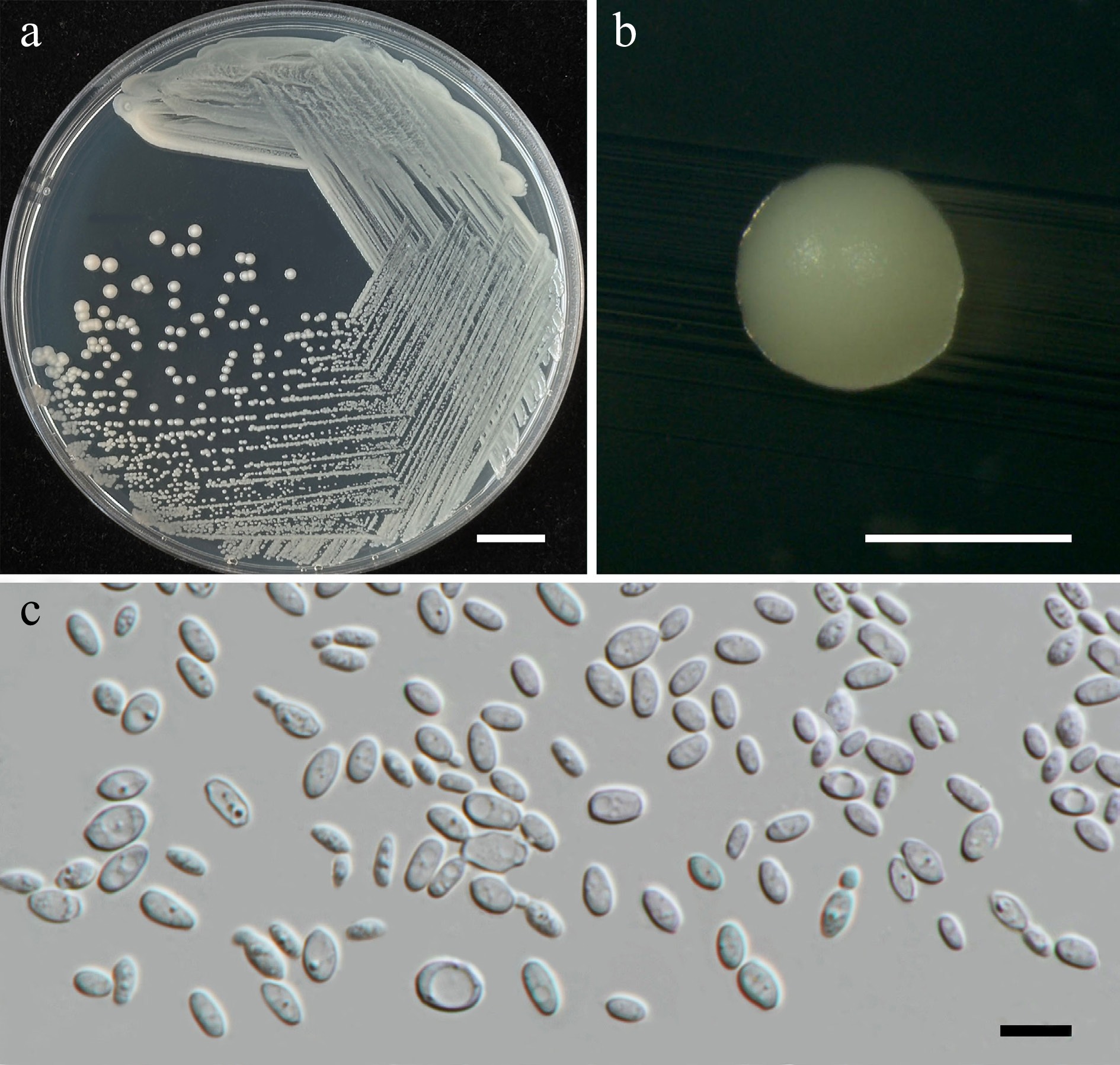
Figure 7.
Morphological characteristics of Wickerhamiella pollinicola (SDBR-CMU569, ex-type). (a) Culture, (b) single colony, (c) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c) = 10 μm.
Etymology – The specific epithet, 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak in kumquat flower (Citrus japonica), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype CMUB40088 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU569 = GMBCC2396 = TBRC21388. GenBank numbers PV834439 (D1/D2), PV834624 (ITS), PX582314 (rpb2).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.5–2 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ovoid to ellipsoidal (2.31–5.42 × 5.15–8.53 μm, n = 50), budding is polar. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Ascospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is positive. D-Glucose, galactose, sorbose, xylose (weak), DL-arabinose (weak), sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, melizitose, glycerol, erythritol, ribose (weak), D-glucitol, mannitol, and D-glucono-1,5-lactone are assimilated, but N-acetyl glucosamine, L-rhamnose, cellobiose, salicin, melibiose, lactose, raffinose, inulin, soluble starch, ribitol, galactitol, myo-inositol, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite (weak), ethylamine (weak), L-lysine (weak), and cadaverine are assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose and 60% glucose. No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is positive. Growth on 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak in kumquat flower (Citrus japonica), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU617 and SDBR-CMU618. GenBank numbers SDBR-CMU617: PV834440 (D1/D2), PV834625 (ITS); SDBR-CMU618: PV834441 (D1/D2), PV834626 (ITS).
Notes – Wickerhamiella pollinicola (SDBR-CMU569, SDBR-CMU617 and SDBR-CMU618) formed a strong clade with 100 MLBS, 0.99 PP support value and clustered as sister to W. azyma (CBS 6826, type species) and W. azymoides (UFMG-R287, type species; Fig. 8). A comparison of D1/D2 domain revealed that W. pollinicola differed from W. azyma and W. azymoides by 4 nucleotide substitutions (0.7% nucleotide divergence) and 9 nucleotide substitutions with 1 gap (1.57% nucleotide divergence), respectively. While W. pollinicola differs by 15 nucleotide substitutions, and 7 gaps (3.54%) from W. azyma, and by 16 nucleotide substitutions and 8 gaps (3.76%) from W. azymoides in ITS region. These differences indicate that the strain represents a new member of the genus Wickerhamiella. Wickerhamiella pollinicola can be distinguished from W. azyma by its ability to assimilate maltose, α-α-trehalose, methyl-α-D-glucoside, melizitose, D-Glucono-1,5-lactone, citrate, ammonium sulfate, and potassium. Wickerhamiella pollinicola can grow at 30 °C, while W. azyma shows maximum growth temperature at 37 °C[50]. Wickerhamiella pollinicola can ferment glucose, whereas W. azymoides cannot, allowing for clear differentiation between the two species[51]. Moreover, W. pollinicola can assimilate D-arabinose, melezitose, potassium nitrate, and sodium nitrite, in contrast to W. azymoides[51].

Figure 8.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing genus Wickerhamiella in Trichomonascaceae. The tree is rooted to Zygoascus hellenicus (CBS 5839) and Z. meyerae (CBS 4099). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Forty-nine strains are included in the combined sequence analysis, which comprise 1,190 characters with gaps. Bootstrap support values for maximum likelihood (≥ 50%, ML, left) and Bayesian posterior probabilities (≥ 0.95, PP, right) are indicated above the nodes. Double dashes (--) denote support values below 50% ML and 0.95 PP. The scale bar represents 0.1 nucleotide substitutions per site. Ex-type strains are shown in bold, and sequences generated in this study are highlighted in blue.
Class: Pichiomycetes M. Groenew., Hittinger, Opulente & A. Rokas
Order: Serinales M. Groenew., Hittinger, Opulente & A. Rokas
Family: Debaryomycetaceae Kurtzman & M. Suzuki
Debaryomycetaceae, the members of this family exhibit polyphyllous budding as their primary form of asexual reproduction. Cells of this family are generally ovate to elliptic, smooth in surface, and of medium size. That widespread ecological distribution and considerable industrial relevance include soil, plants, food products, animal tissues insects, which may function as either beneficial symbionts or occasional opportunistic pathogens[52,53]. Currently, 23 genera are listed in this family including Aciculoconidium, Candida, Danielozyma, Debaryomyces, Diutina, Hemisphaericaspora, Hyphopichia, Kodamaea, Kurtzmaniella, Limtongozyma, Lodderomyces, Metahyphopichia, Meyerozyma, Millerozyma, Nematodospora, Priceomyces, Scheffersomyces, Schwanniomyces, Spathaspora, Suhomyces, Teunomyces, Wickerhamia, and Yamadazyma[41]. In this study, five yeast species were presented, including Kodamaea ohmeri (five strains), K. restingae (two strains), Candida tropicalis (one strain), Meyerozyma caribbica (two strains), and Priceomyces siamensis sp. nov. (two strains).
Priceomyces siamensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 9)
MycoBank number: MB860172
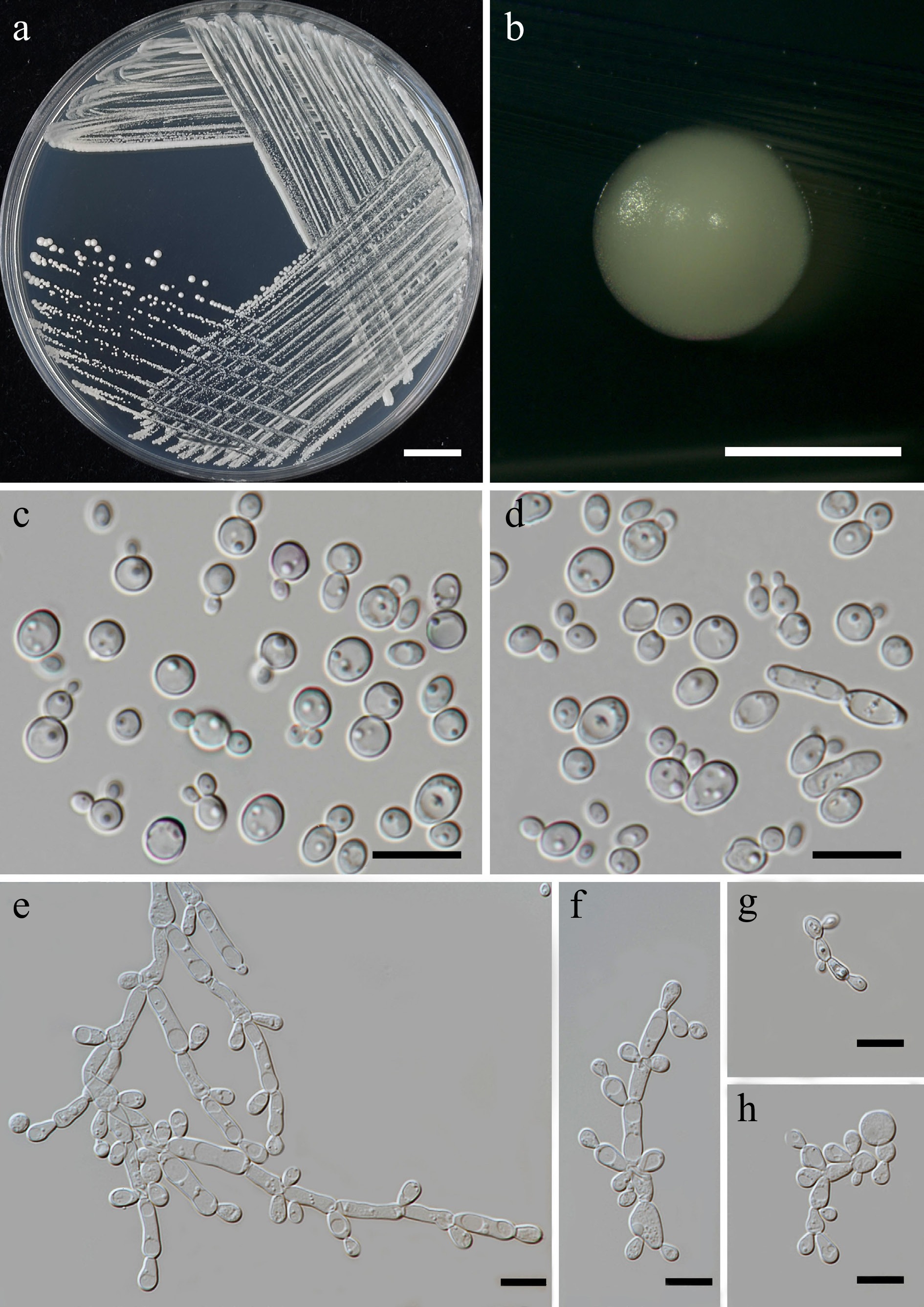
Figure 9.
Morphological characteristics of Priceomyces siamensis (SDBR-CMU614, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 days at 25 °C. (e)–(h) Pseudohyphae and blastoconidia on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c)–(h) = 10 μm.
Etymology – 'siamensis' referring to Siam, the old name of Thailand, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Mae Hia in Yellow elder flower (Tecoma stans), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype, CMUB40099 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU614 = GMBCC2397 = TBRC21399. GenBank numbers PV834452 (D1/D2), PV834637 (ITS), PX622318 (SSU), PX582294 (rpb1), PX582315 (rpb2).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.5–2 mm in diameter), white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spherical, ovoid, ellipsoidal (2.95–4.88 × 3.28–9.5 μm, n = 50). Budding is polar. Blastoconidia in Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae are formed. Ascospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is positive. D-Glucose, galactose, sorbose, N-acetyl glucosamine (weak), ribose, xylose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, melizitose, glycerol, erythritol, ribitol, D-glucitol, mannitol, D-glucono-1,5-lactone, D-gluconate, succinate, citrate, and xylitol are assimilated, but L-arabinose, D-arabinose, L-rhamnose, salicin, melibiose, lactose, raffinose, inulin, soluble starch, galactitol, myo-inositol, D-glucuronate, methanol, D-galacturonic acid, DL-lactate, and ethanol are not assimilated. Ammonium sulfates and ethylamine hydrochloride are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, L-lysine, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose, 10% NaCl/5% glucose, and 16% NaCl/5% glucose. Not growth occurs on media containing 0.01% cycloheximide and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak in portulaca flower (Portulaca grandiflora), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU701. GenBank numbers PV834453 (D1/D2) PV834638 (ITS), PX622319 (SSU).
Notes – In the phylogenetic analyses, two new strains (SDBR-CMU614 and SDBR-CMU701) of P. siamensis formed a separate lineage, sister to P. melissophilus with 100% MLBS and 0.99 PP support values (Fig. 10) and clustered with P. fermenticarens CBS7040 and P. castillae CBS 6053. The present strains differed from those strains by 0.86% nucleotide divergence (5 nucleotide substitutions), 1.36% (8 nucleotide substitutions and 2 gaps) and 3.51% (20 nucleotide substitutions and 2 gaps), respectively, in D1/D2. The ITS sequences of the three strains by 1.59% nucleotide divergence (10 nucleotide substitutions and 4 gaps), 0.79% (5 nucleotide substitutions and 4 gaps), and 4.90% (28 nt substitutions and 16 gaps), respectively, which indicated that strain represents a new member of Priceomyces. The phenotypic comparisons between P. siamensis and P. melissophilus, P. fermenticarens, and P. castillae are shown in Table 4.
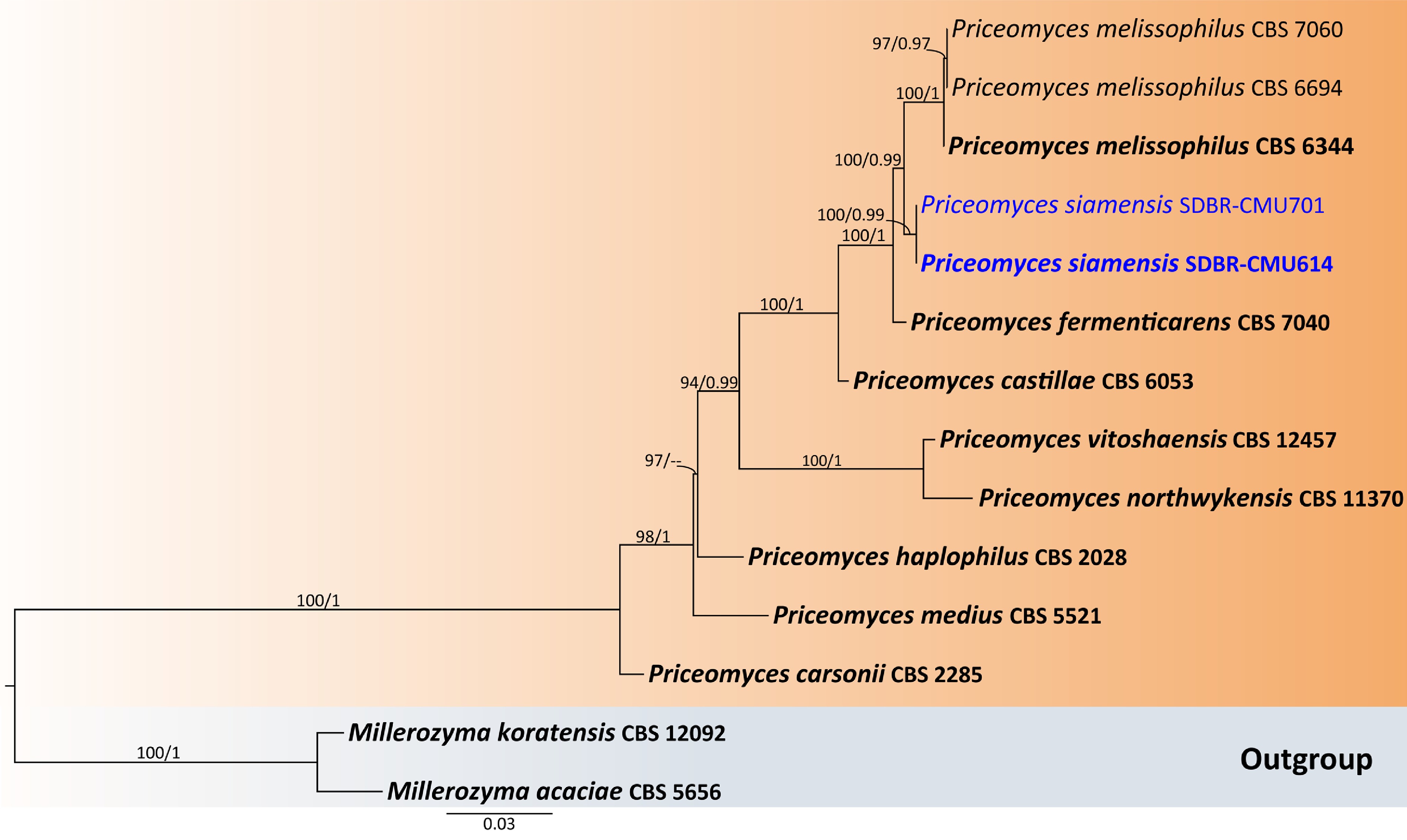
Figure 10.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Priceomyces. The tree is rooted to Millerozyma acaciae (CBS 5656) and M. koratensis (CBS 12092). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fourteen strains are included in the combined sequence analysis, which comprise 1,121 characters with gaps. Bootstrap support values for maximum likelihood (≥ 50%, ML, left), and Bayesian posterior probabilities (≥ 0.95, PP, right) are indicated above the nodes. Double dashes (--) denote support values below 50% ML and 0.95 PP. The scale bar represents 0.03 nucleotide substitutions per site. Ex-type strains are shown in bold, and sequences generated in this study are highlighted in blue.
Table 4. Phenotypic characteristics differentiating Priceomyces siamensis from closely related Priceomyces species.
Characteristics 1 2 3 4 Fermentation Glucose + – – – Carbon assimilation D-Ribose + – + v D-Xylose + – w + L-Arabinose w – w v L-Rhamnose – – – + Sucrose + + – + Maltose + + – + α-α-Trehalose + – – + Cellobiose + w – v Salicin – – + + Melibiose – + – v Lactose – – – + Raffinose – – – v Melizitose + – – + Soluble starch – – – + Erythritol + + + – Succinate + – + + Ethanol – + + + Nitrogen assimilation Potassium nitrate – – – nd Growth characteristics Growth at 37 °C – + + + Strains 1: P. siamensis sp. nov, 2: P. melissophilus[54], 3: P. fermenticarens[55], and 4: P. castillae[27]. Family: Metschnikowiaceae T. Kamieński ex Doweld
The members of this family generally have elongated, elliptic, or needle-shaped cell shapes developed within elongated asci, especially in Metschnikowia[56]. Many species in this group are found in habitats associated with fruits due to their ability to ferment with high glucose and to thrive in high-sugar environments. It is noteworthy that it produces antimicrobial compounds, especially against other fungi, which contribute to the ecosystem[57]. Hyde et al.[41] mention 15 genera are listed in this family including Australozyma, Candidozyma, Clavispora, Danielia, Gabaldonia, Gaillardinia, Helenozyma, Hermanozyma, Isabelozyma, Metschnikowia, Osmozyma, Soucietia, Sungouiella, Tanozyma, and Wilhelminamyces. In this study, two yeast species were presented, including one strain of Metschnikowia cibodasensis, and 16 strains of Me. koreensis (Supplementary File 1).
Class: Saccharomycetes G. Winter
Order: Phaffomycetales M. Groenew., Hittinger, Opulente & A. Rokas
Family: Phaffomycetaceae Y. Yamada, H. Kawas., Nagats., Mikata & T. Seki
Phaffomycetaceae represents exhibit spherical to ellipsoidal cell morphology with vegetative reproduction occurring through multilateral budding[58]. Members of this family exhibit moderate fermentative abilities and can assimilate various carbon compounds. Some species show adaptations to specific environments and are also found in plants, insects, or soil habitats[59]. Currently, four genera are recorded in this family including Barnettozyma, Cyberlindnera, Phaffomyces, and Starmera[41]. In this study, Cyberlindnera fabianii (one strain) was identified (Supplementary File 1).
Order: Saccharomycodales M. Groenew., Hittinger, Opulente & A. Rokas
Family: Saccharomycodaceae Kudryavtsev
Saccharomycodaceae is characterized by its unique vegetative reproduction via bipolar budding, where buds form on broad bases at the cell poles, resulting in the characteristic lemon or apiculate cell shape that distinguishes many members of this group[27]. The members of this family exhibit varying degrees of fermentative capability, with some species being strongly fermentative. Many species demonstrate notable tolerance to acidic environments, explaining their prevalence in fruit-associated habitats and fermentation processes[58]. Currently, two genera are listed in this family including Hanseniaspora and Saccharomycodes[41]. In this study, Hanseniaspora lachancei (two strains) are presented (Supplementary File 1).
Phylum: Basidiomycota R.T. Moore
Subphylum: Agaricomycotina Doweld
Class: Tremellomycetes Doweld
Order: Filobasidiales Jülich
Family: Filobasidiaceae L.S. Olive
Filobasidiaceae, their basidial morphology consists of long, tubular, and often holobasidia-like structures (filobasidia) that develop terminal basidiospores through lateral budding rather than on sterigmata. These basidiospores are typically produced in a row along the elongated basidium[60]. Vegetative reproduction occurs through budding, the cells generally appearing spherical to oval. Additionally, some members produce extracellular polysaccharide capsules and exhibit distinctive carotenoid pigmentation, resulting in orange or red colony coloration. The cell wall composition includes xylose and mannose as predominant carbohydrates[61]. Physiologically, Filobasidiaceae do not ferment sugars but can assimilate various carbon compounds. Currently, six genera are listed in this family including Filobasidium, Goffeauzyma, Heterocephalacria, Naganishia, Syzygospora, and Zyzygomyces[41]. In this study Filobasidium lannaense sp. nov. (two strains), Naganishia albida (one strain), N. diffluens (two strains), and N. liquefaciens (one strain) were presented.
Filobasidium lannaense Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 11)
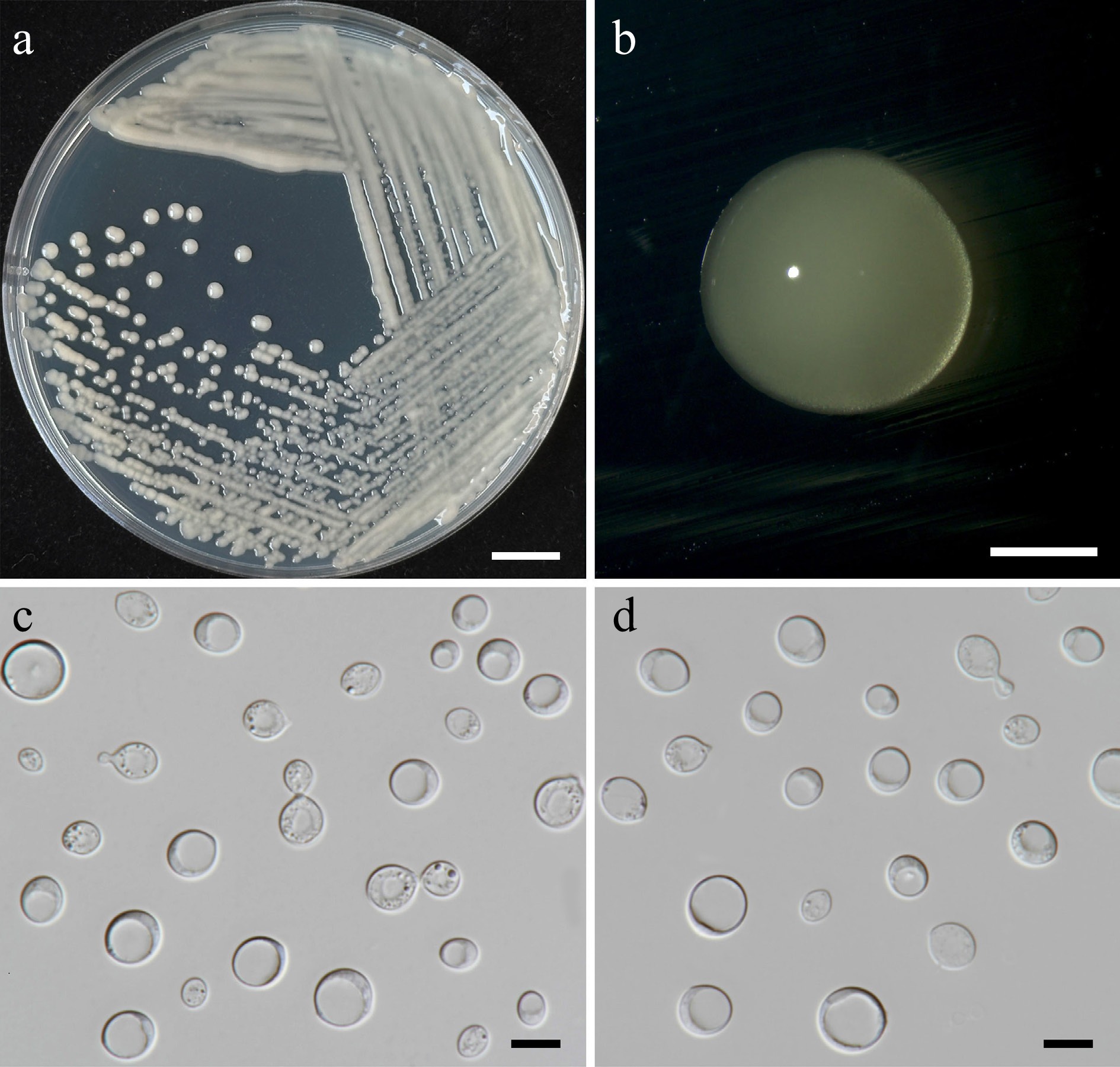
Figure 11.
Morphological characteristics of Filobasidium lannaense (SDBR-CMU628, ex-type). (a) Culture, (b) single colony, (c), (d) budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
MycoBank number: MB860174
Etymology – 'lannaense' referring to the Kingdom of Lanna, the historic name of northern Thailand, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), August 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype, CMUB40102 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU628 = GMBCC2398 = TBRC21403. GenBank numbers PV834474 (D1/D2), PV834644 (ITS), PX622320 (SSU), PX582295 (rpb1), PX582334 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (3–3.5 mm in diameter), white to pale yellow, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are globosal and ellipsoidal (5.06–12.55 × 6.4–13.31 μm, n = 50), budding is polar. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose lactose, raffinose, melizitose, glycerol (or weak), D-mannitol, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate (or weak), D-galacturonic acid, citrate, succinate, ethanol (or weak), and xylitol are assimilated, but L-sorbose, N-acetyl glucosamine, inulin, soluble starch, erythritol, ribitol, D-glucitol, DL-lactate, and methanol are not assimilated. Ammonium sulfates, potassium nitrate (or weak), sodium nitrite, ethylamine hydrochloride and L-lysine are assimilated as sole nitrogen source, but cadaverine are not assimilated. No growth occurs on media containing 50% glucose, and 60% glucose. No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth at 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in canna lily flower (Canna indica), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU681. GenBank numbers PV834475 (D1/D2), PV834642 (ITS), PX622321 (SSU), PX582296 (rpb1), PX582335 (tef1-α).
Notes – A phylogenetic analysis of the combined D1/D2 domain and ITS sequence dataset shows that SDBR-CMU628 and SDBR-CMU681 strains are closely related to Filobasidium pseudomali (NYNU 2111105 and NYNU 22986), F. mali CGMCC 2.4012, F. castaneae NYNU 2111105 and F. globosum (CGMCC 2.5680 and CGMCC 2.5656) (Fig. 12). The D1/D2 sequences differed by 0.16%–1% nucleotide divergence (2–6 nt substitutions) and 2.10%–8.75% (13–53 nt substitutions) in the ITS region. Therefore, a new species, Filobasidium lannaense is introduced in Filobasidiaceae. Additionally, phenotypic differences between F. lannaense and F. pseudomali, F. castaneae, F. mali, and F. globosum are shown in Table 5.
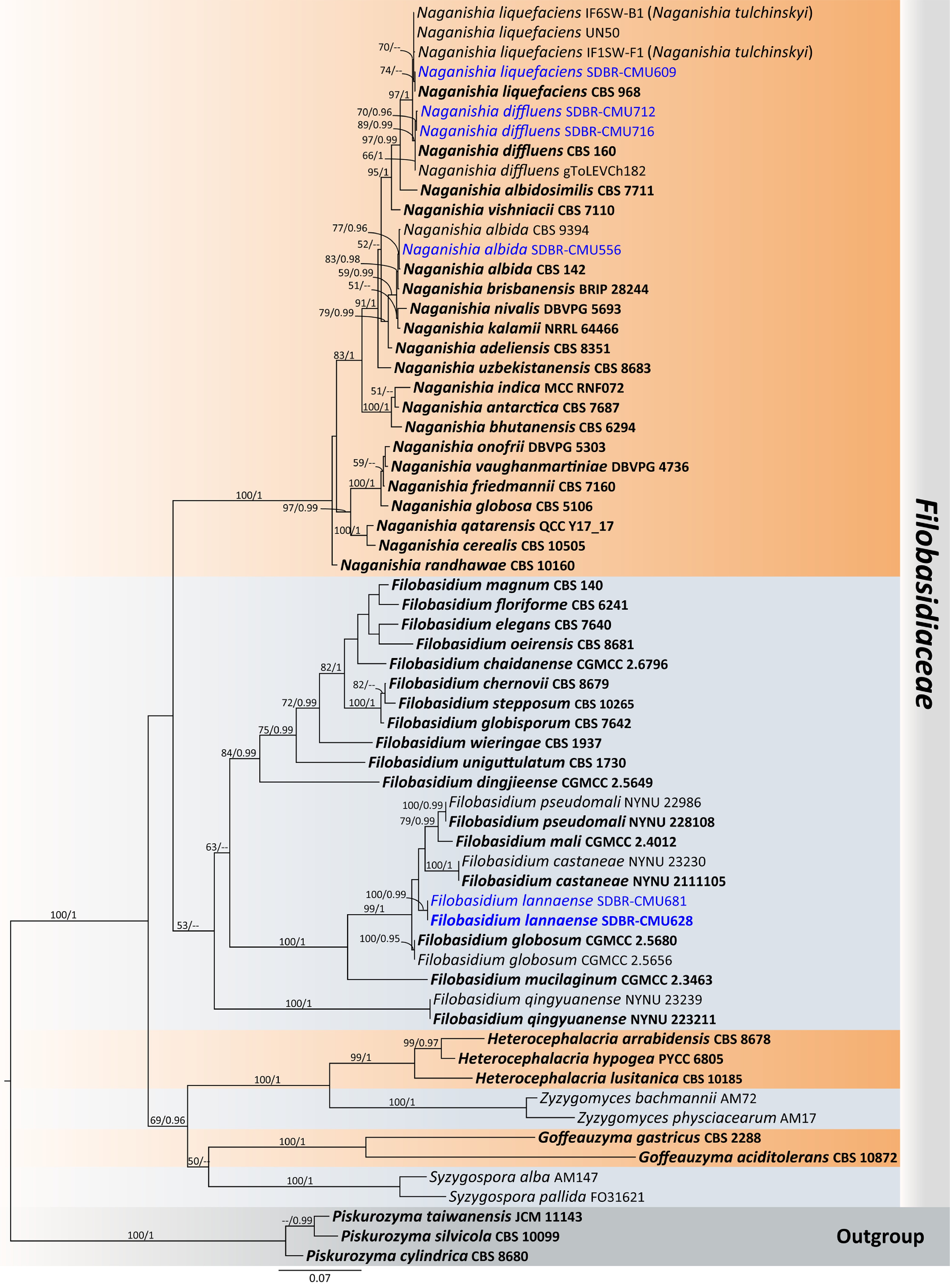
Figure 12.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Filobasidiaceae. The tree is rooted to Piskurozyma cylindrica (CBS 8680), Pi. silvicola (CBS 10099), and Pi. taiwanensis (JCM 11143). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Sixty-four strains are included in the combined sequence analysis, which comprise 1,398 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Table 5. Phenotypic characteristics differentiating Filobasidium lannaense from closely related Filobasidium species.
Characteristics 1 2 3 4 5 Carbon assimilation L-Sorbose – + – + + N-Acetyl glucosamine – + – – – D-Ribose + + – – – D-Xylose + + w w – L-Arabinose + + + w + D-Arabinose + – – – + L-Rhamnose + + w w v Methyl-α-D-glucoside + + w w + Cellobiose + + + w + Salicin + + – w – Melibiose + + + w + Lactose + + + – + Raffinose + + + w + Inulin – + + – + Glycerol w + – – – Ribitol w + – – + D-Glucitol + + – – + Galactitol + + – + + myo-Inositol + + w w + Succinate + + w w + Citrate + + – – + Ethanol w – – w + Nitrogen assimilation Sodium nitrite + + – – + Ethylamine HCl w + + – + Cadaverine – – – + – Growth characteristics Growth at 25 °C + + + + + Growth at 30 °C + – – v + Growth at 35 °C – – – v – Strains 1: F. lannaense sp. nov., 2: F. castaneae[62], 3: F. globosum, 4: F. mali[63], and 5: F. pseudomali[62]. Order: Tremellales Fr.
Family: Bulleribasidiaceae Xin Zhan Liu, F.Y. Bai, M. Groenew. & Boekhout
Bulleribasidiaceae are found on the surfaces of plants, particularly as organisms that reside on the leaf blades. Numerous species exhibit psychrotolerant traits, which enable them to thrive in cold climates[64]. Characterized by this group, colonies can be white, cream, pink, or orange due to the production of carotenoid pigments. The cells are spherical, oval, or elongated. Physiologically, most species do not ferment sugars but display a wide range of abilities to assimilate various carbon compounds. Most are urea positive and can use inositol as a carbon source, a property that aids in species identification[65]. Currently, six genera are listed in this family including Bulleribasidium, Derxomyces, Dioszegia, Hannaella, Nielozyma, and Vishniacozyma[41]. In this study, Hannaella pagnoccae (four strains), Ha. phyllophila (one strain), Vishniacozyma marinae (one strain), and V. pollinicola sp. nov. (two strains) were presented (Fig. 13, Supplementary File 1).
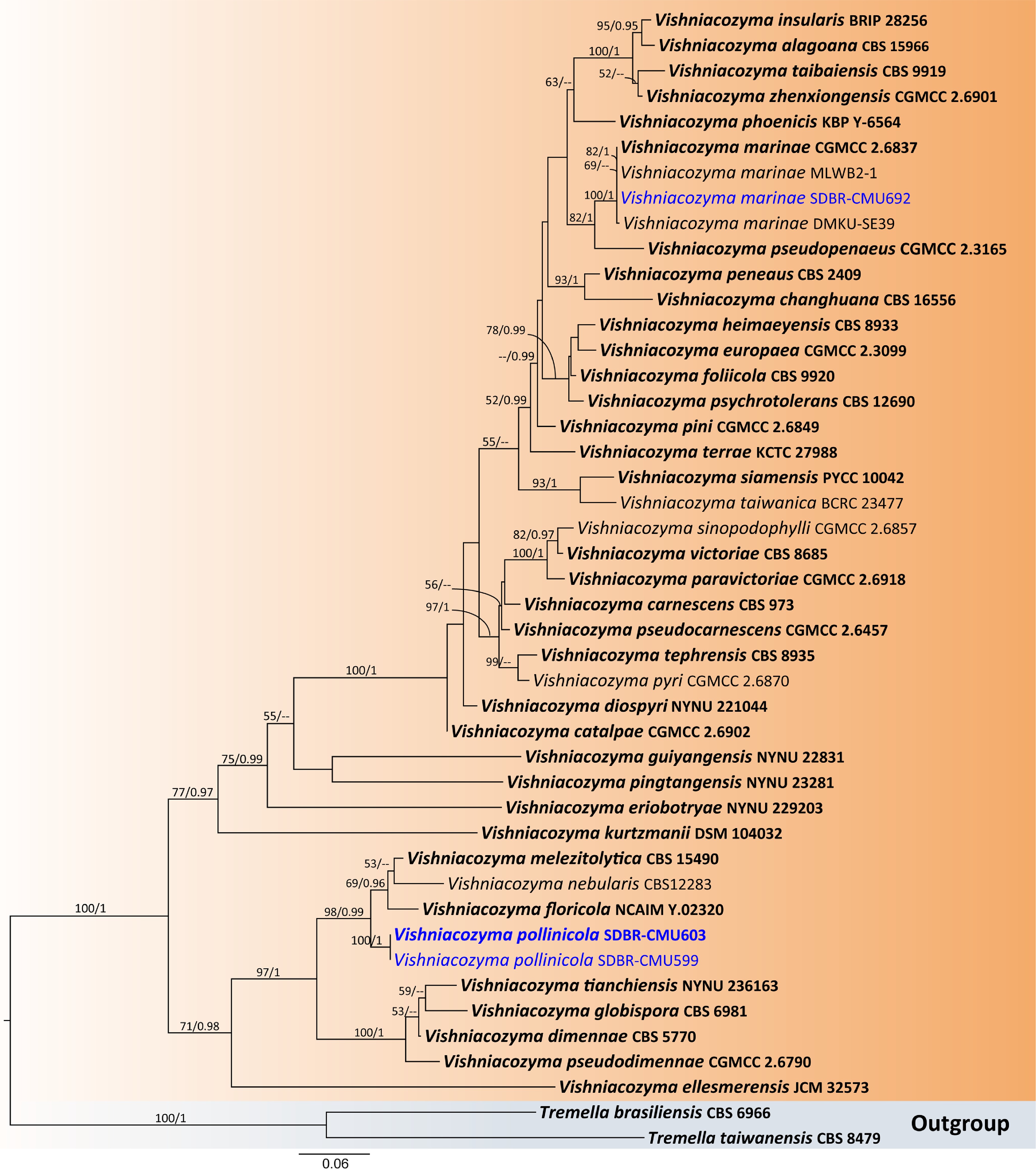
Figure 13.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Vishniacozyma. The tree is rooted to Tremella brasiliensis (CBS 6966) and T. taiwanensis (CBS 8479). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fourty-five strains are included in the combined sequence analysis, which comprise 1,204 characters with gaps. The average standard deviation of the split frequencies of the BI analysis was 0.004360. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Vishniacozyma pollinicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 14)
MycoBank number: MB860175
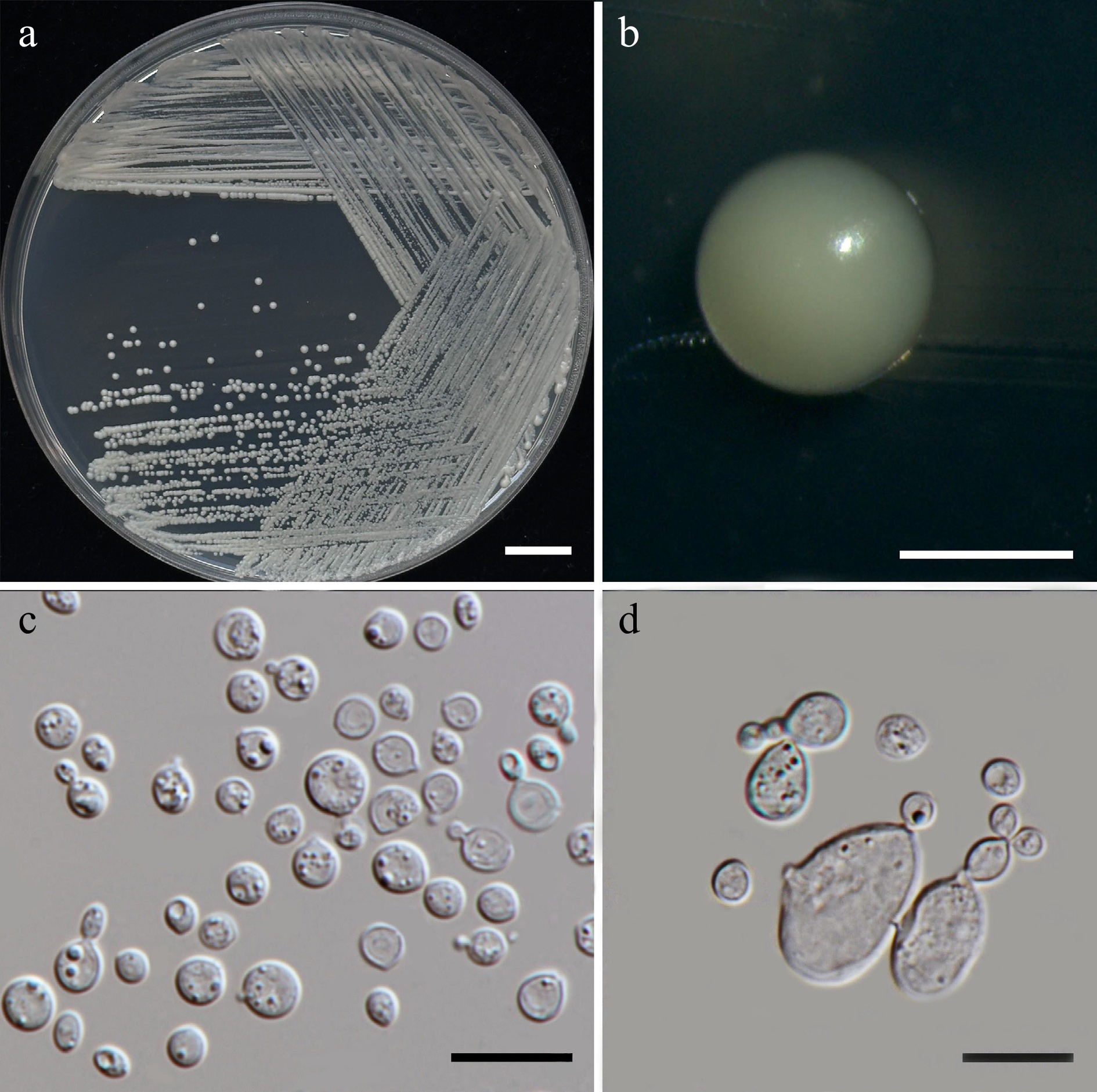
Figure 14.
Morphological characteristics of Vishniacozyma pollinicola (SDBR-CMU603, ex-type). (a) Culture, (b) single colony, (c) cells and budding cells on YMA after 5 d at 25 °C. (d) Rudimentary pseudohypha on YMA after 3 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The specific epithet 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in frangipani flower (Plumeria obtusa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40097 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU603 = GMBCC2399 = TBRC21396. GenBank numbers PV834487 (D1/D2), PV834657 (ITS), PV941856 (rpb1), PV947458 (rpb2), PV947490 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.0–1.5 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroid to ellipsoid (2.55–4.95 × 3.03–5.75 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose (or slow), sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin (or weak), melibiose (or weak), lactose, raffinose, melizitose, D-mannitol, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid (or weak), citrate (or slow), succinate, ethanol, and xylitol are assimilated, but L-sorbose, inulin, soluble starch, glycerol, erythritol, ribitol, D-glucitol, DL-lactate, and methanol are not assimilated. Ammonium sulfates, ethylamine hydrochloride, and L-lysine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and cadaverine are not assimilated. No growth occurs on media containing 50% glucose and 60% glucose. No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth occurs at 10, 15, 25, 30, but is absent at 35, 37, and 40 °C.
Additional strains examined: THAILAND, Chiang Mai Province, Mueang District, Suthep, in Ixora flower (Ixora sp.), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU599. GenBank numbers PV834486 (D1/D2), PV834656 (ITS).
Notes – Phylogenetic analyses of a concatenated D1/D2 domain and ITS sequence dataset show that the present strains (SDBR-CMU599 and SDBR-CMU603) form a separate clade, clustering with V. melezitolytica CBS 15490, V. nebularis CBS 12283, and V. floricola NCAIMY.02320 (98% MLBS and 0.99 BYPP; Fig. 13). The D1/D2 sequences of the present strains differed 1.13%–1.94 % nucleotide divergence with 7 nt substitutions from V. melezitolytica, 12 nt substitutions from V. nebularis and 11 nt substitutions from V. floricola. The ITS sequences also demonstrated divergence 1.79%–2.83% (9–13 nt substitutions and 16–17 gaps) of three species, supporting the recognition of the present strains as a novel species within Vishniacozyma. Furthermore, V. pollinicola can be differentiated from V. melezitolytica with the ability to assimilate melibiose, melizitose, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, citrate, and xylitol[63]. In contrast, V. pollinicola does not assimilate in sorbose, inulin, glycerol, ribitol, and D-glucitol, while V. melezitolytica can grow. Unfortunately, the phenotypic characteristics of V. nebularis and V. floricola were not observed[66], and thus comparisons across the species could not be made.
Family: Cryptococcaceae Kütz. ex-Castell. & Chalm.
Cryptococcaceae, this family can produce polysaccharide capsules surrounding the yeast cells, particularly evident in pathogenic and environmental survival[67,68]. The vegetative reproduction occurs primarily through budding, the cells generally appearing spherical to oval, and some species exhibit distinctive colony morphologies ranging from mucoid due to capsule production to dry[69]. Physiologically, Cryptococcaceae species typically do not ferment sugars but demonstrate varied abilities to assimilate different carbon and nitrogen compounds. Some species produce urease, which serves as a diagnostic. The family includes both environmental saprophytes and opportunistic pathogens of medical significance[70]. Currently, three genera are listed in this family including Cryptococcus, Kwoniella, and Teunia[41]. In this study, Kwoniella bestiolae (two strains), Kw. heveanensis (two strains), Kw. limtongiae sp. nov (two strains), and Kw. saisamorniae sp. nov. (two strains) were presented (Fig. 15).
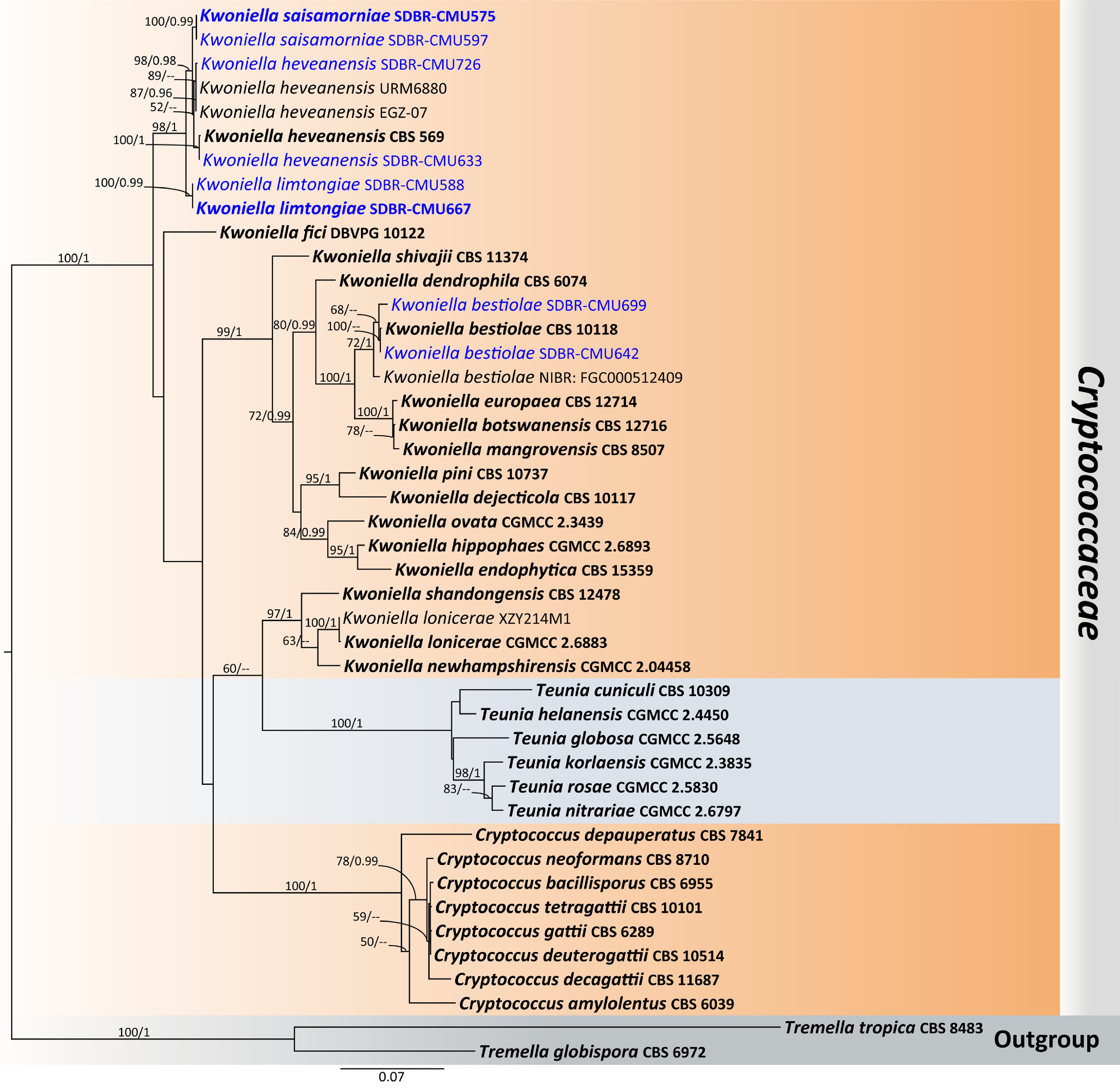
Figure 15.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Cryptococcaceae. The tree is rooted to Tremella globispora (CBS 6972) and T. tropica (CBS 8483). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fourty-four strains are included in the combined sequence analysis, which comprise 1,274 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Kwoniella limtongiae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 16)
MycoBank number: MB860178
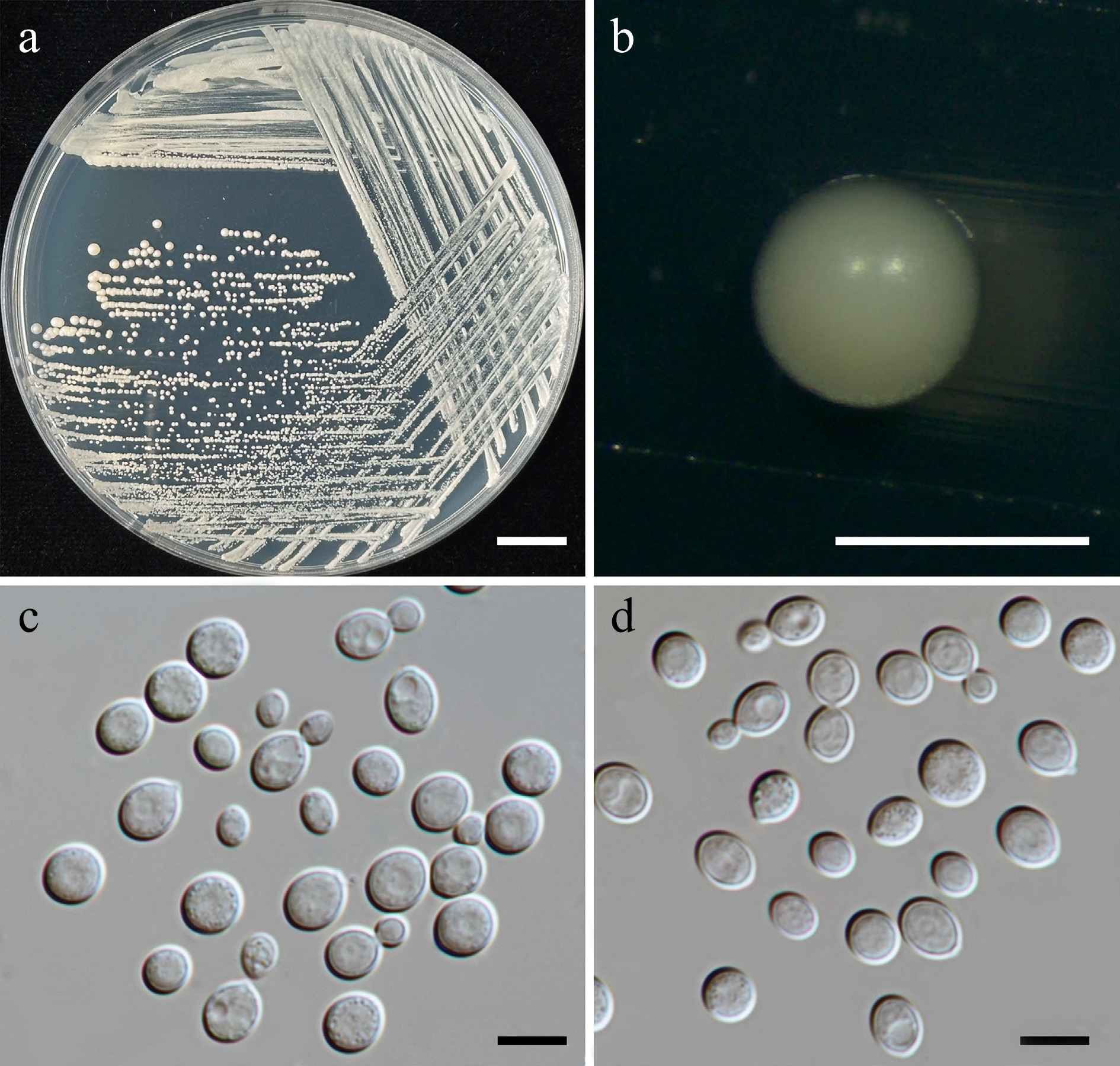
Figure 16.
Morphological characteristics of Kwoniella limtongiae (SDBR-CMU667, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'limtongiae' named in honor of mycologist Savitree Limtong, for her contributions to yeast systematics.
Holotype – THAILAND, Phayao Province, Mueang District, in galphimia flower (Thryallis glauca), August 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype, CMUB40107 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU667 = GMBCC2400 = TBRC21409. GenBank numbers PV834493 (D1/D2), PV834663 (ITS), PX622323 (SSU), PX582317 (rpb2), PX570015 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.2 mm in diameter), yellowish white, smooth surface, entire margin, and convex elevation. The cells are globosal and ellipsoidal, (6.11–8.97 × 5.94–10.38 μm, n = 50), budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, ribose (weak), xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin (weak), lactose, raffinose, melizitose, glycerol (weak), ribitol, D-glucitol, mannitol, galactitol, myo-inositol, D-gluconate, D-galacturonic acid, DL-lactate, succinate, and xylitol are assimilated, but N-acetyl glucosamine, melibiose, inulin, soluble starch, erythritol, D-glucono-1,5-lactone, D-glucuronate (weak), citrate, methanol, and ethanol are not assimilated. Ammonium sulfates, ethylamine hydrochloride, L-lysine, and cadaverine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and creatine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction and acid formation are positive. Growth was observed at 10, 15, 25, 35, and 37 °C but not at 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in Orange jasmine flower (Murraya paniculata), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU588. GenBank numbers PV834492 (D1/D2), PV834662 (ITS), PX622322 (SSU), PX582316 (rpb2), PX570014 (tef1-α).
Notes – Phylogenetic analyses of a combined D1/D2 domain and ITS sequence data demonstrated that two strains of Kwoniella limtongiae (SDBR-CMU667 and SDBR-CMU588) formed a robust clade and clustered with Kw. heveanensis, and Kw. saisamorniae with 98% MLBS and 1 BYPP statistic support (Fig. 15). The D1/D2 sequences of the two strains differed by 0.32% nucleotide divergence (2 nt substitutions), and 2.07% (12 nt substitutions) in ITS region. Furthermore, Kw. limtongiae differed from Kw. saisamorniae by 1.90% nucleotide divergence (11 nt substitutions), indicating that these strains represent a novel species within Kwoniella. Additionally, phenotypic differences between Kw. limtongiae, Kw. heveanensis, and Kw. saisamorniae are shown in Table 6.
Table 6. Phenotypic characteristics differentiating Kwoniella limtongiae and Kw. saisamorniae from closely related Kwoniella species.
Characteristics 1 2 3 Carbon assimilation D-Ribose w + + D-Arabinose + + – Salicin w – + Glycerol w + – Erythritol – + + D-Glucono-1,5-lactone – + nd D-Glucuronate – w + DL-Lactate + + w Citrate – – w Ethanol – – + Nitrogen assimilation Creatine – – + Growth characteristics Growth at 35 °C + w nd Growth at 37 °C + – + Strains 1: Kw. limtongiae sp. nov., 2: Kw. saisamorniae sp. nov., and 3: Kw. heveanensis[69]. Kwoniella saisamorniae Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 17)
MycoBank number: MB860182
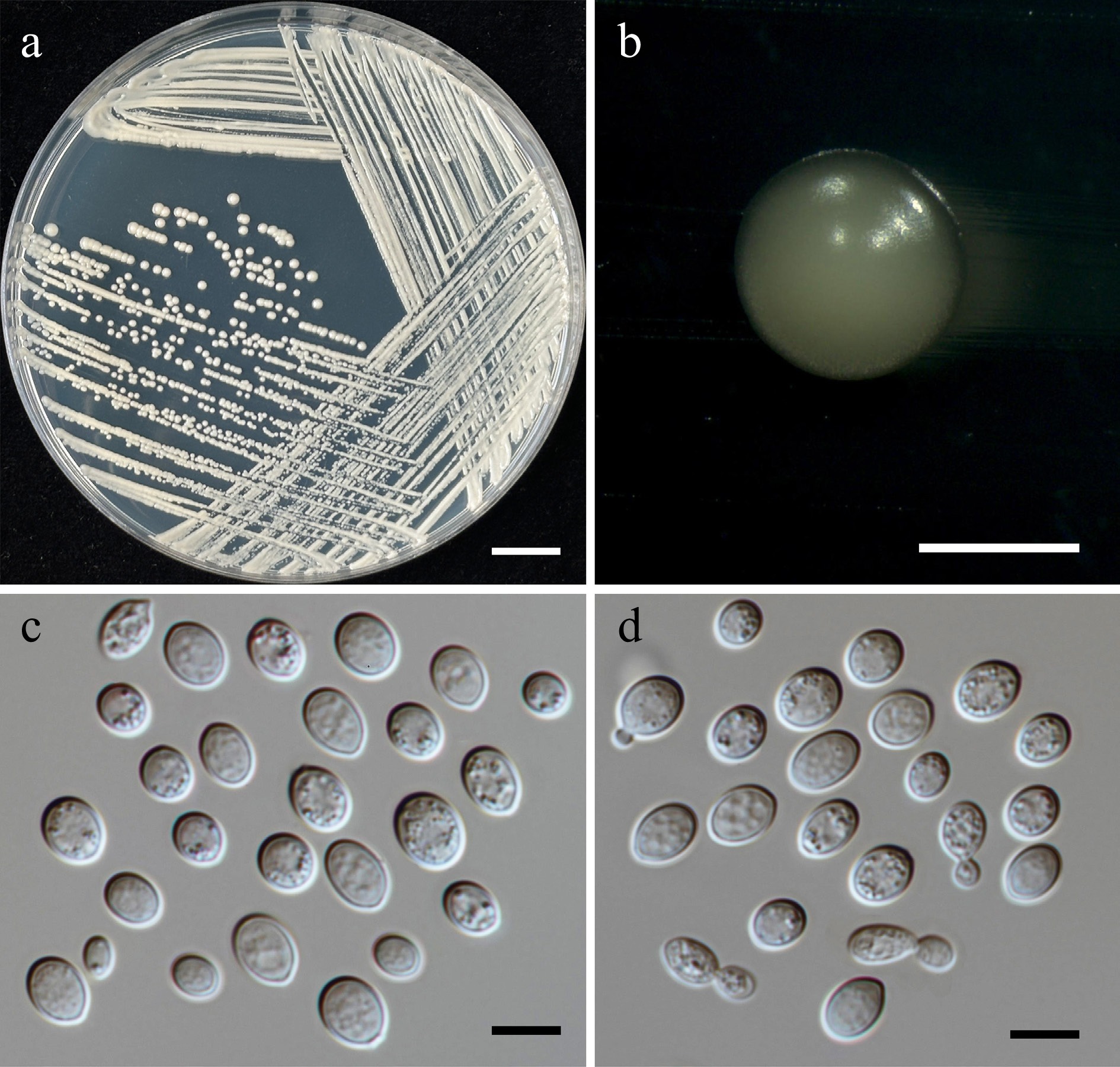
Figure 17.
Morphological characteristics of Kwoniella saisamorniae (SDBR-CMU575, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'saisamorniae' referring to the name of mycologist Saisamorm Lumyong in honor of her 75th birthday.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in Siam tulip flower (Curcuma sessilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype CMUB40090 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU575 = GMBCC2401 = TBRC21390. GenBank numbers PV834494 (D1/D2), PV834664 (ITS), PX622324 (SSU), PX570009 (rpb1), PX582318 (rpb2), PX582336 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.2 mm in diameter), yellowish white, smooth surface, entire margin, and convex elevation. The cells are broadly ellipsoidal, (4.49–9.29 × 5.5–12.1 μm, n = 50), budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, lactose, raffinose, melizitose, glycerol, erythritol, ribitol, D-glucitol, mannitol, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate (weak), D-galacturonic acid, DL-lactate, succinate, and xylitol are assimilated, but N-acetyl glucosamine, salicin, melibiose, inulin, soluble starch, citrate, methanol, and ethanol are not assimilated. Ammonium sulfates, ethylamine hydrochloride, L-lysine, and cadaverine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite and creatine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction and acid formation are positive. Growth was observed at 10, 15, 25 °C but not at 37 and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in confederate vine flower (Antigonon leptopus), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU597. GenBank numbers PV834495 (D1/D2), PV834665 (ITS), PX622325 (SSU), PX570010 (rpb1), PX582319 (rpb2), PX582337 (tef1-α).
Notes – Multi-locus analyses revealed that two strains of SDBR-CMU575 and SDBR-CMU597 formed a clade sister to Kw. heveanensis (Fig. 15). Based on the BLASTn search, the closest match using D1/D2 and ITS sequences is Kw. heveanensis, showing 0.32% nucleotide divergence (1 nt substitutions) and 1.08% (6 nt substitutions), respectively. Moreover, Kw. saisamorniae is different from Kw. heveanensis by 3.27 and 6.09% nucleotide divergence (27 and 56 nt substitutions) in rpb1 and tef1-α, respectively. Thus, we introduce a new species, Kw. saisamorniae, isolated from flowers of Antigonon leptopus and Curcuma sessilis, Thailand. In addition, phenotypic differences between Kw. saisamorniae, Kw. heveanensis, and Kw. limtongiae are shown in Table 6.
Family: Rhynchogastremaceae Oberw. & B. Metzler
The family was established by Metzler et al.[71] to accommodate the unique dimorphic mycoparasite Rhynchogastrema coronatum. The type species demonstrates both yeast-like and filamentous forms depending on environmental conditions and life cycle stage. The vegetative morphology includes both unicellular yeast-like forms and filamentous forms. The cell is spherical to oval and hyphal forms exhibit typical basidiomycetous characteristics including septate hyphae and specialized reproductive structures[71,72]. The ecology of Rhynchogastremaceae is primarily that of mycoparasites, serving important functions in fungal community dynamics. Their parasitic activities can influence host population dynamics and contribute to biodiversity maintenance in fungal ecosystems[73]. Currently, two genera are listed in this family including Papiliotrema and Rhynchogastrema[41]. In this study, Papiliotrema aspenensi (one strain), Pa. chiangmaiensis sp. nov. (two strains), Pa. pollinicola sp. nov. (two strains), and Pa. tectonae sp. nov. (two strains) were presented.
Papiliotrema chiangmaiensis Kodchasee, Senwanna, J. Kumla & N. Suwannar., sp. nov. (Fig. 18)
MycoBank number: MB860184
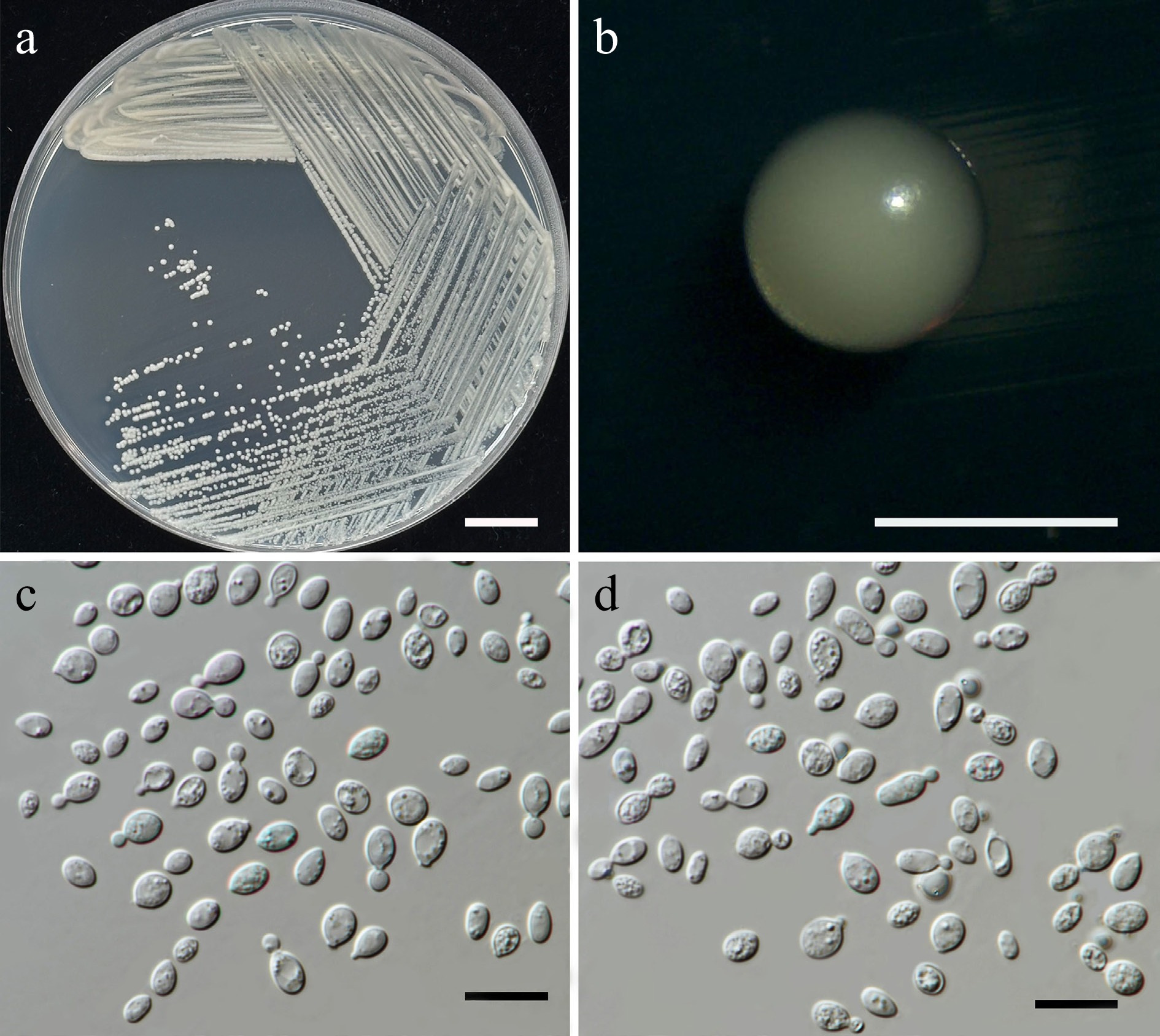
Figure 18.
Morphological characteristics of Papiliotrema chiangmaiensis (SDBR-CMU594, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'chiangmaiensis' referring to Chiang Mai Province, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in Zamioculcas flower (Zamioculcas zamiifolia) July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40095 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU594 = GMBCC2402 = TBRC21394. GenBank numbers PV834497 (D1/D2), PV834667 (ITS), PX622326 (SSU), PV941857 (rpb1), PV947469 (rpb2), PV844824 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1–2 mm in diameter), white to pale yellow, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ellipsoidal and cylindrical (2.67– 4.27 × 3.84–5.76 μm, n = 50), Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose (or weak), ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose, raffinose, melizitose, erythritol, ribitol, D-glucitol, D-mannitol, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, succinate, ethanol (or slow), and xylitol are assimilated, but N-acetyl glucosamine, inulin, soluble starch, glycerol, DL-lactate, citrate, and methanol are not assimilated. Ammonium sulfate, L-lysine, and ethylamine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose. No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction and acid formation are positive. Growth was observed at 10, 20, and 25 °C but 35, and 37 °C are variable.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU627. GenBank numbers PV834498 (D1/D2), PV834668 (ITS), PX622327 (SSU).
Notes – The new species, Papiliotrema chiangmaiensis, strains SDBR-CMU594 and SDBR-CMU627 formed a distinct lineage, closely related to Pa. leoncinii CBS 13918 and Pa. tapputiae BRIP 75038 (Fig. 19). Papiliotrema chiangmaiensis differed from Pa. leoncinii and Pa. tapputiae by 1.04% nucleotide divergence (6 nt substitutions and 1 gap), and 0.82% (5 nt substitutions), respectively, in the D1/D2 domain. While the ITS regions showed 0.2%–1.2% mismatches compared to those known species. Moreover, Pa. chiangmaiensis can be distinguished from Pa. leoncinii with its ability to assimilate methyl-α-D-glucoside, galactitol, and ethylamine. But cannot grow inulin, citrate, and sodium nitrite[74].
Figure 19.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Rhynchogastremaceae. The tree is rooted to Bullera alba (CBS 500) and B. unica (CBS 8290). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fifty-eight strains are included in the combined sequence analysis, which comprise 1,218 characters with gaps. The average standard deviation of the split frequencies of the BI analysis was 0.009286. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Papiliotrema pollinicola Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 20)
MycoBank number: MB860186
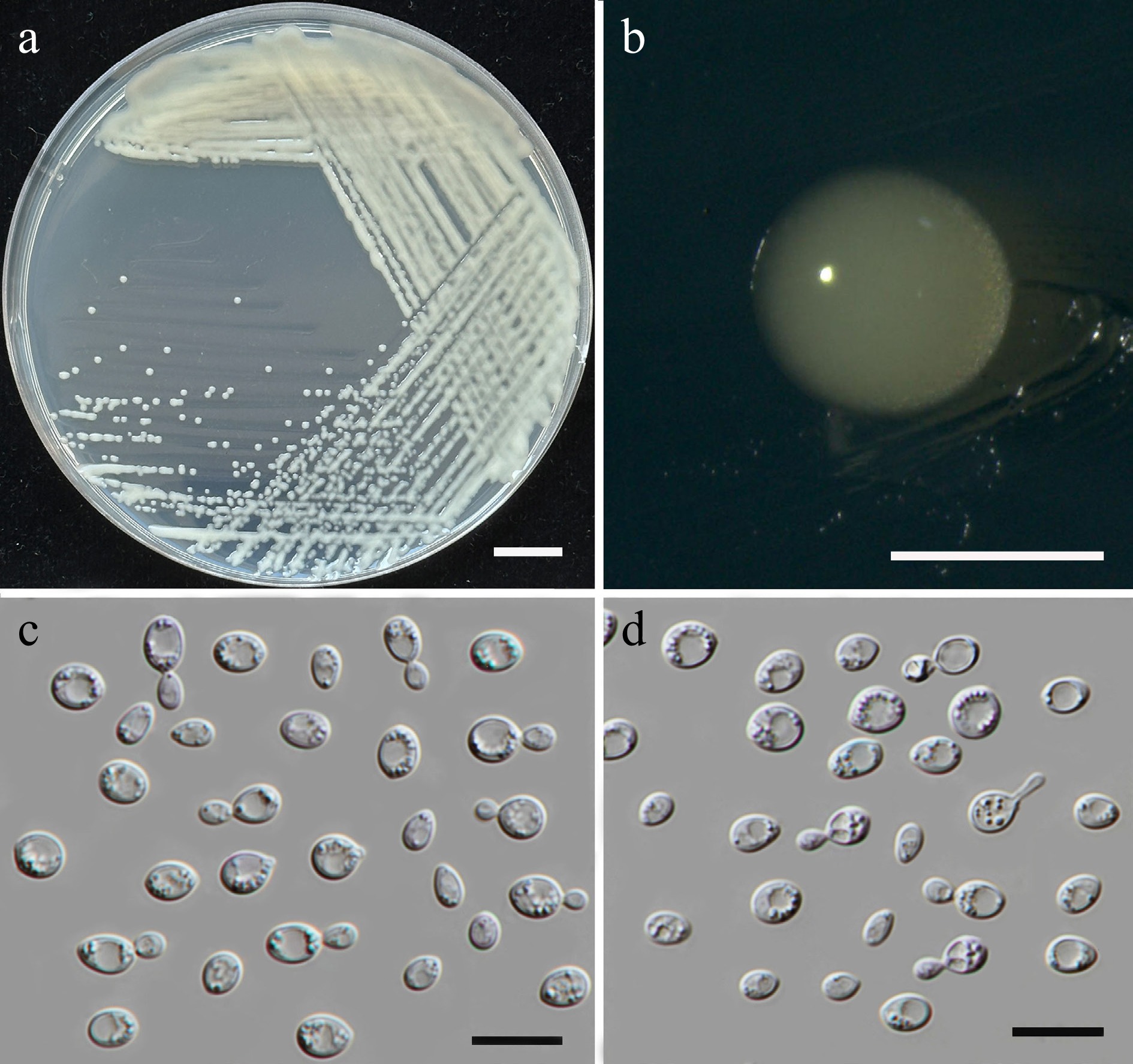
Figure 20.
Morphological characteristics of Papiliotrema pollinicola (SDBR-CMU664, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The specific epithet 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Phayao Province, Mueang District, in vinca flower (Catharanthus roseus), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40106 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU664 = GMBCC2403 = TBRC21408. GenBank numbers PV834501 (D1/D2), PV834671 (ITS), PX622329 (SSU), PX582297 (rpb1), PV947470 (rpb2), PV844825 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.5 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are globose to ellipsoidal (3.1–5.58 × 3.64–5.9 μm, n = 50), occur singly in pairs, or in chains budding. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose, raffinose, melizitose, glycerol, erythritol, ribitol, D-glucitol, mannitol, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, ethanol, and xylitol are assimilated, but methanol, inulin, and soluble starch are not assimilated. Ammonium sulfate, L-lysine, and ethylamine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth at 10, 20, 25, and 30 °C but absent at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in Zamioculcas (Zamioculcas zamiifolia), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU595. GenBank numbers PV834500 (D1/D2), PV834670 (ITS), PX622328 (SSU).
Notes – The new species, Papiliotrema pollinicola (SDBR-CMU664 and SDBR-CMU595) formed a distinct clade with 100 MLBS, 1 BYPP support value and clustered as sister to Pa. rajasthanensis (CBS 10406; type strain, STA1.8 and BRT 572; Fig. 19). A sequence comparison revealed that Pa. pollinicola differed from the type strain of Pa. rajasthanensis by 3.70% nucleotide divergence (38 substitutions and 1 gap) in the D1/D2 domain, 0.56% (3 substitutions) in the ITS region, and 1.96% nucleotide divergence (21 substitutions and 6 gaps) in the rpb2 gene. Based on physiological test, Pa. pollinicola can be distinguished from Pa. rajasthanensis by its ability to assimilate glycerol and DL-lactate. Likewise, Pa. pollinicola was able to grow at 50% glucose, 60% glucose, and 10% NaCl/5% glucose[75].
Papiliotrema tectonae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 21)
MycoBank No: MB860185
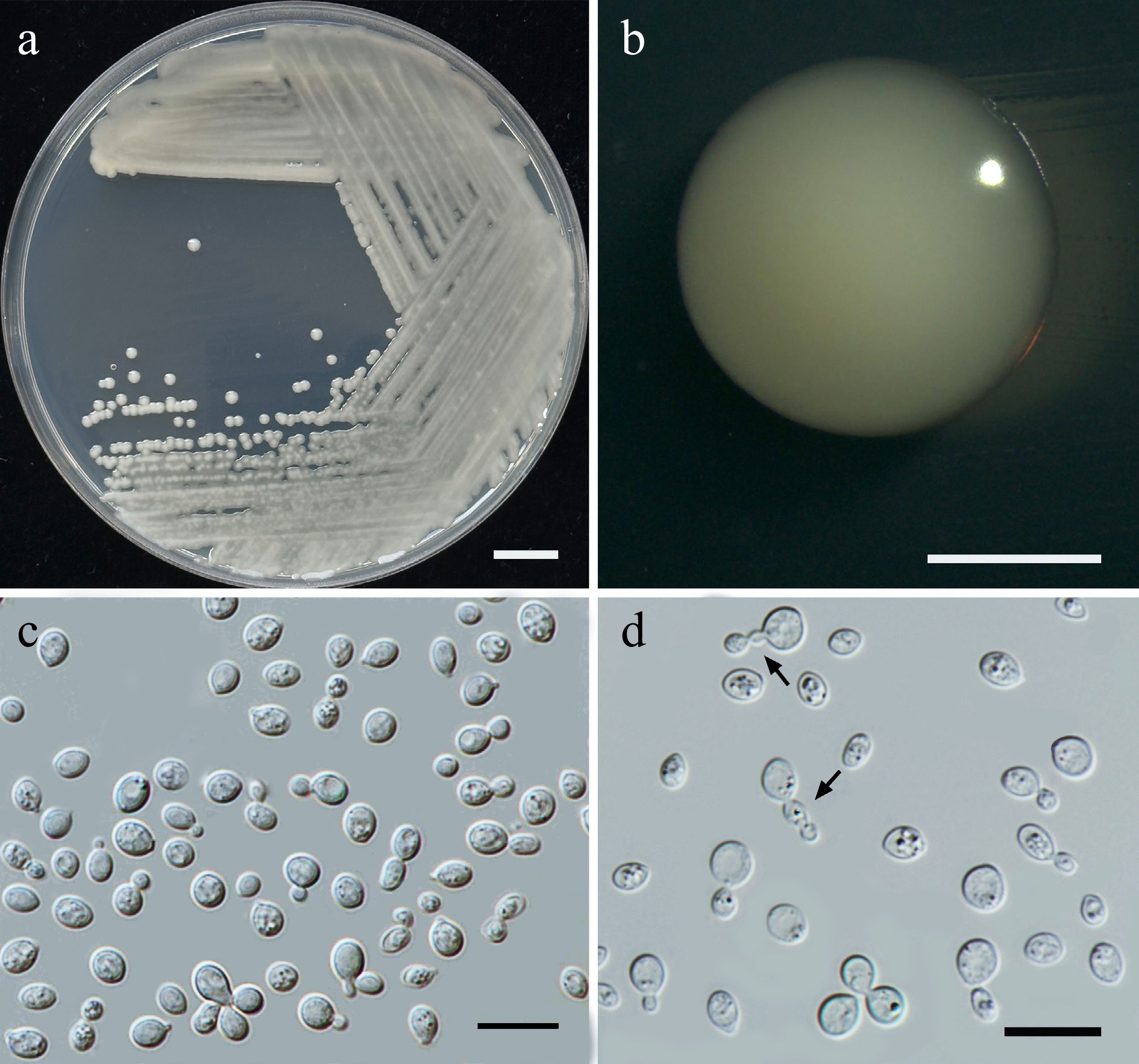
Figure 21.
Morphological characteristics of Papiliotrema tectonae (SDBR-CMU632, ex-type). (a) Culture, (b) single colony, (c), (d) budding cells, and chains of cells (indicated by arrows) on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The specific epithet 'tectonae' refers to Tectona, the plant genus from which the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach holotype, CMUB40103 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU632 = GMBCC2404 = TBRC21404. GenBank numbers PV834503 (D1/D2), PV834673 (ITS), PX622331 (SSU), PV941859 (rpb1), PV947472 (rpb2), PV844827 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1–2 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are globose to ellipsoidal (2.25– 4.75 × 2.85–5.56 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, N-acetyl glucosamine (or slow), ribose (or weak), xylose, L-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside (or slow), cellobiose, salicin (or slow), melibiose (or slow), lactose, raffinose, melizitose, ribitol (or slow), D-glucitol, mannitol, galactitol, myo-inositol (or weak), D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, and xylitol (or slow) are assimilated, but L-sorbose, methanol, ethanol, inulin, soluble starch, glycerol, erythritol, D-arabinose, and L-rhamnose are not assimilated. Ammonium sulfate and L-lysine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, ethylamine and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction and acid formation are positive. Growth at 10, 20, 25, and 30 °C, but absent at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU630. GenBank numbers PV834502 (D1/D2), PV834672 (ITS), PX622330 (SSU), PV941858 (rpb1), PV947471 (rpb2), PV844826 (tef1-α).
Notes – In the phylogenetic analyses, the strains SDBR-CMU632 and SDBR-CMU630 of Papiliotrema tectonae formed a separate lineage, sister to Pa. anmochoreius (CBS 10258) with 75% MLBS and 1 BYPP support values (Fig. 19). The present strains differed from the latter by 2.86% nucleotide divergence (18 nt substitutions) in the D1/D2 domain and 4.75% (25 nt substitutions and 8 gaps) in the ITS region. In addition, Pa. tectonae differs from Pa. anemochoreia by its inability to assimilate inulin glycerol, erythritol, D-arabinose, and L-rhamnose, whereas Pa. anemochoreia can utilize these substrates. Additionally, Pa. tectonae was able to grow on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose[76].
Family: Trimorphomycetaceae Xin Zhan Liu, F.Y. Bai, M. Groenew, & Boekhout
Trimorphomycetaceae represents a morphologically and ecologically distinctive family within the including mixed conidial-basidial fruiting bodies, distinctive twinned conidia in the type genus, and mycoparasite. Trimorphomycetaceae can exist in a variety of growth forms, including the distinctive filamentous and basidiocarp structures and yeast stages. Currently, four genera are listed in this family including Carlosrosaea, Saitozyma, Sugitazyma, and Trimorphomyces[41]. In this study, Saitozyma thailandensis sp. nov. (three strains) was presented.
Saitozyma thailandensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 22)
MycoBank number: MB860188
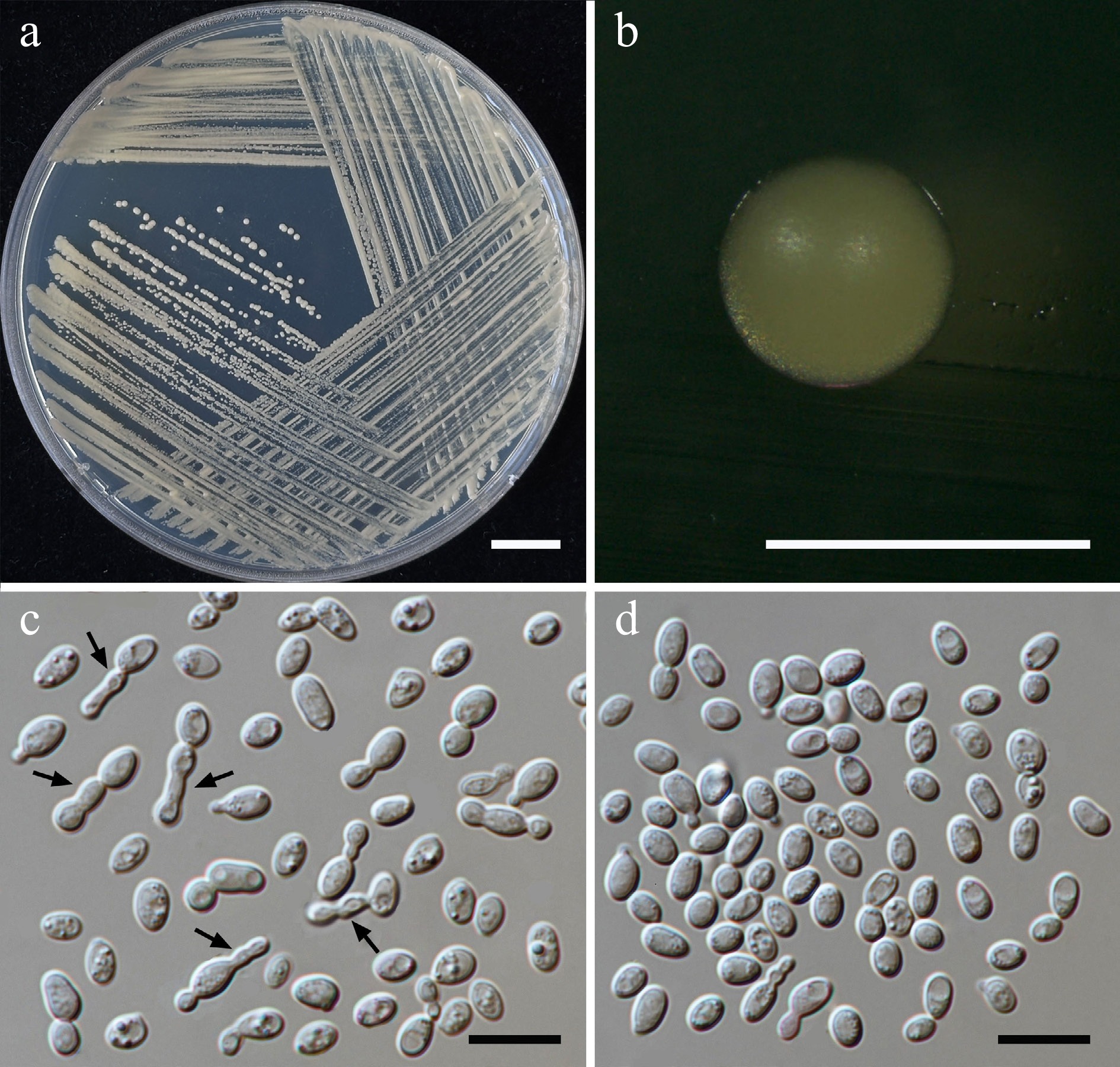
Figure 22.
Morphological characteristics of Saitozyma thailandensis (SDBR-CMU590, ex-type). (a) Culture, (b) single colony, (c),chains of cells (indicated by arrows), (d) budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The species name 'thailandensis' refers to Thailand, the country where the type strain was isolated.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in confederate vine flower (Antigonon leptopus), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40092 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU590 = GMBCC2405 = TBRC21392. GenBank numbers PV834504 (D1/D2), PV834674 (ITS).
Description – The culture on YMA after 5 days at 25 °C, colonies are circular form (0.8–1.0 mm in diameter), pale yellow, smooth surface, entire margin, and convex elevation. The cells are ellipsoidal to cylindrical (1.84–4.6 × 3.65–6.07 μm, n = 50), occur singly, in pairs or in chains budding. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiosporess were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for 1 month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose (weak), raffinose, melizitose, erythritol, ribitol (weak), D-glucitol, mannitol, galactitol, myo-inositol (weak), D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, and xylitol are assimilated, but inulin, soluble starch, glycerol, methanol, and ethanol are not assimilated. Ammonium sulfates, ethylamine hydrochloride, L-lysine, and cadaverine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and creatine are not assimilated. Growth occurs on media containing 50% glucose (weak). No growth occurs on media containing 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is positive (weak). Growth at 10, 20, 25, 35, and 37 °C but absent at 40 °C
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in American cassia flower (Senna spectabilis), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU636; Chiang Rai Province, Mae Fah Luang District, Mae Fah Luang, in bush clock vine flower (Thunbergia erecta), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU698. GenBank numbers SDBR-CMU636: PV834506 (D1/D2), PV834676 (ITS); SDBR-CMU698: PV834505 (D1/D2), PV834675 (ITS).
Notes – Saitozyma thailandensis strains SDBR-CMU590, SDBR-CMU636 and SDBR-CMU698 were placed in the Saitozyma clade (Fig. 23), and closely related to Sa. flava CBS 331, Sa. pseudoflava CBS 15576 and Sa. paraflava CBS 10100. These strains showed which 0.16%, 0.49%, and 1.16% nucleotide divergence (1, 3, and 8 nt substitutions) in the D1/D2 domain, respectively. The ITS region had differences by 1.17%, 1.93%, and 4.15% (6, 10, and 20 nt substitutions). Additionally, after 2 weeks on PDA, the cell presented a few chains and elongated formed and elongated but not eveloped to pseudohyphae and true hyphae (Fig. 22c), and a distinct comparison of the phenotypic characteristics between Sa. thailandensis, Sa. flava, Sa. paraflava, and Sa. pseudoflava are shown in Table 7. Thus, Sa. thailandensis is introduced as a new species.
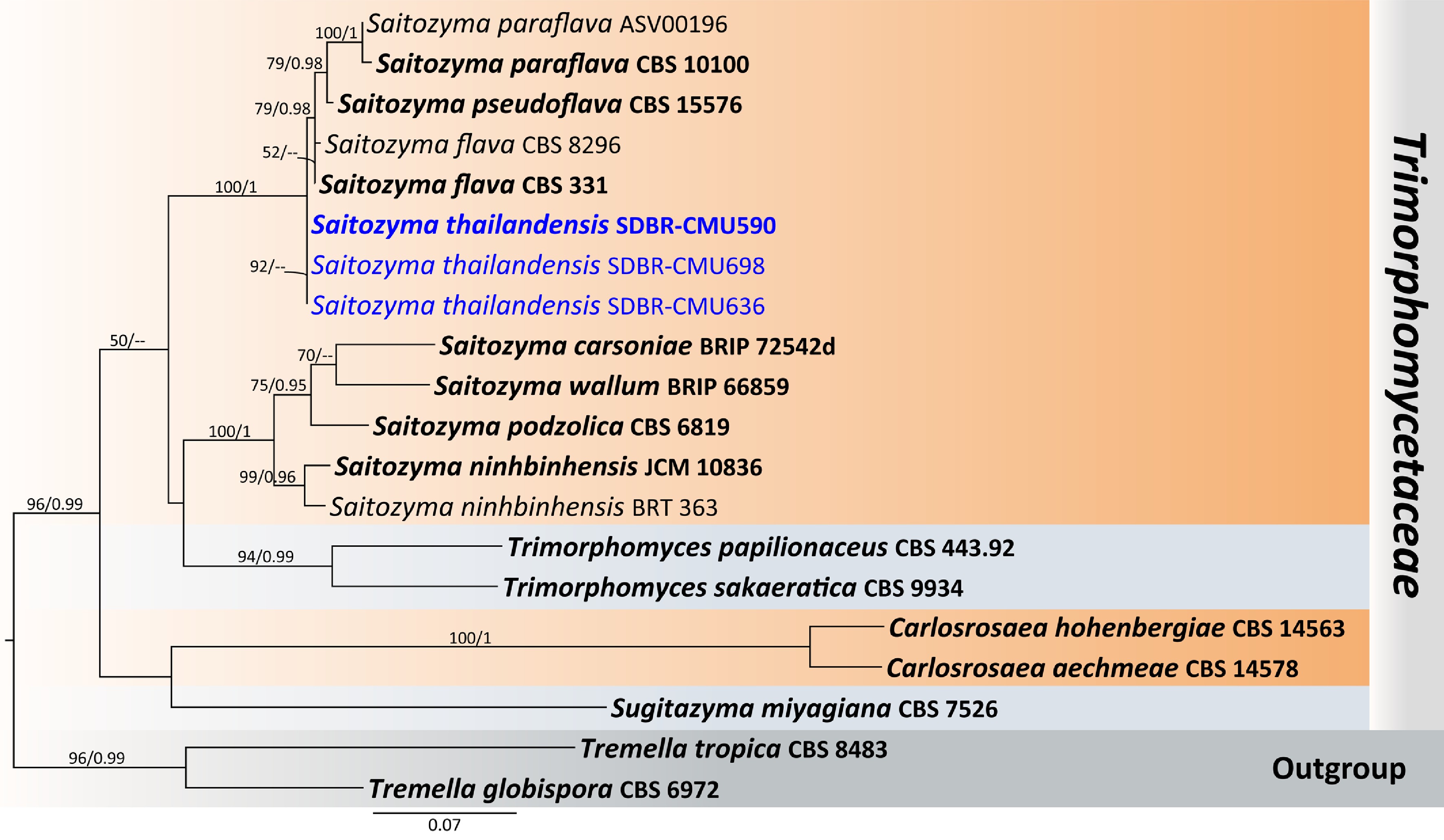
Figure 23.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Trimorphomycetaceae. The tree is rooted to Tremella globispora (CBS 6972) and T. tropica (CBS 8483). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Twenty strains are included in the combined sequence analysis, which comprise 1,149 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Table 7. Phenotypic characteristics differentiating Saitozyma thailandensis from closely related Saitozyma species.
Characteristics 1 2 3 4 Carbon assimilation L-Sorbose + – – – N-Acetyl glucosamine + + + l/w D-Arabinose + w +/w +/w L-Rhamnose + + +/w l/w α-α-Trehalose + + + – Salicin + + – l/w Melibiose + + + +/w Lactose w + + – Inulin – w – +/w Soluble starch – + +/s – Glycerol – – +/w – Erythritol + + – – Ribitol w w + l/w D-Glucitol + + –/+ l/w D-Mannitol + + –/w l/w Galactitol + w –/w + myo-Inositol w + + + DL-Lactate + w +/s – Succinate + + +/s – Citrate + w +/s – Nitrogen assimilation of Ammonium sulfate + – – + Ethylamine HCl + nd – + L-Lysine + – – l/w Growth characteristics Growth at 30 °C + + – + Growth at 35 °C + – nd – Growth at 37 °C + – nd – Growth on 50% Glucose w – – nd Strains 1: Sa. thailandensis sp. nov, 2: Sa. flava[77], 3: Sa. paraflava[78], and 4: Sa. pseudoflava[63]. Order: Trichosporonales Boekhout & Fell
Family: Trichosporonaceae Nann.
Trichosporonaceae is found in a wide range of habitats, including soil, water, and plant surfaces, and are constituents of the normal microbiota of humans and other animals. Members of Trichosporonaceae exist primarily as yeasts but can produce various hyphal structures. The characteristic and taxonomically is the production of arthroconidia - asexual propagules that form through the disarticulation of septate hyphae at the septal junctions. These arthroconidia are typically unicellular, ranging from cubical to barrel-shaped or elongate[60,79]. Currently, nine genera are listed in this family, including Apiotrichum, Cutaneotrichosporon, Effuseotrichosporon, Haglerozyma, Nothotrichosporon, Pascua, Prillingera, Trichosporon, and Vanrija[41]. In this study, Trichosporon asahii (one strain) is presented (Supplementary File 1).
Subphylum: Pucciniomycotina R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw.
Class: Agaricostilbomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw.
Order: Agaricostilbales Oberw. & R. Bauer
Family: Chionosphaeraceae Oberw. & Bandoni
Chionosphaeraceae demonstrate a wide range of habitats ranging from saprotrophic decomposition to mycoparasitic relationships with other fungi. Characteristics of this family exhibit unicellular yeasts or gasteroid basidia with simultaneous basidiospore production per basidium, that allow these fungi to exploit different ecological and environmental[65,80]. Currently, six genera are listed in this family including Ballistosporomyces, Boekhoutia, Chionosphaera, Cystobasidiopsis, Kurtzmanomyces, and Stilbum[41]. In this study, Boekhoutia pollinicola sp. nov. (two strains) were presented.
Boekhoutia pollinicola Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 24)
MycoBank number: MB860189
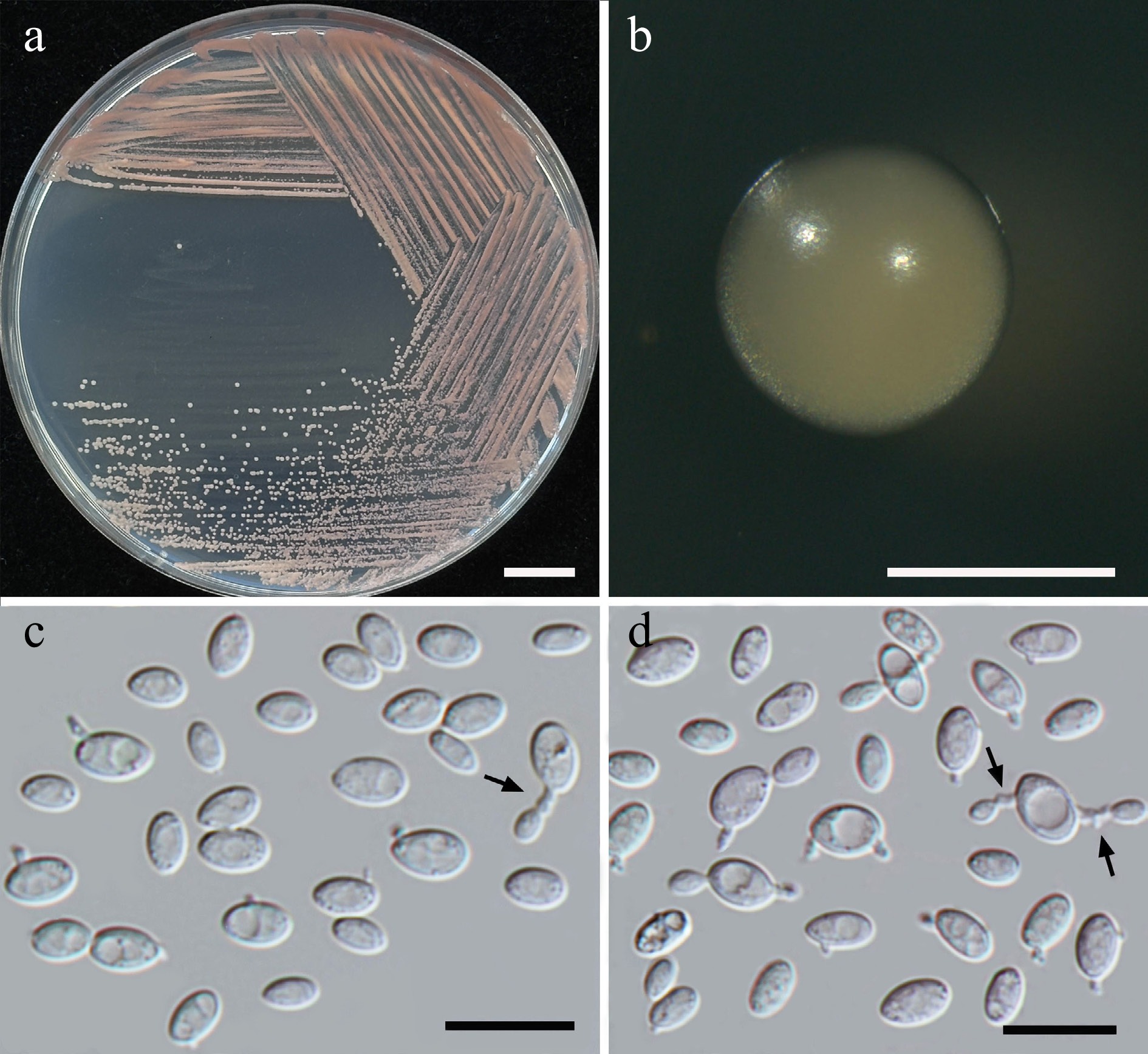
Figure 24.
Morphological characteristics of Boekhoutia pollinicola (SDBR-CMU684, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on sterigma (indicated by arrows) on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – the specific epithet 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Phayao Province, Mueang District, Baan Tom, in bridal bouquet flower (Plumeria pudica), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40109 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU684 = GMBCC2406 = TBRC21411. GenBank numbers PV834510 (D1/D2), PV834680 (ITS), PX622333 (SSU), PV941873 (rpb1), PV947459 (rpb2), PV947486 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1–2 mm in diameter), light orange, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroidal to short ovoidal (1.65–3.54 × 3.77–6.17 μm, n = 50), occur singly or in pairs budding on sterigma. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, ribose, xylose, sucrose, maltose, α-α-trehalose (or slow), methyl-α-D-glucoside (or weak), cellobiose (or slow), melibiose (or latent), lactose (or latent), raffinose (or latent), melizitose, erythritol (or latent or slow), ribitol (or slow), glucitol (weak or slow), mannitol (weak or slow), D-gluconate, DL-lactate, and succinate (weak or latent) are assimilated, but N-acetyl glucosamine, L-rhamnose, salicin, inulin, soluble starch, glycerol, galactitol, myo-inositol, D-glucuronate, D-galacturonic acid, citrate, methanol, ethanol, and xylitol are not assimilated. Assimilation is variable for D-glucono-1,5-lactone, and D-arabinose. Ammonium sulfate, L-lysine, ethylamine hydrochloride, and cadaverine are assimilated as the sole nitrogen source. Potassium nitrate and sodium nitrite are not assimilated. No growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose, 15% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Growth in vitamin-free medium is negative. Acid formation is negative. Urease reaction is positive. Growth at 10, 20, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in Siam tulip flower (Curcuma sessilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, living culture SDBR-CMU621. GenBank numbers PV834509 (D1/D2), PV834679 (ITS), PX622332 (SSU).
Notes – In molecular phylogenetic analysis, two strains, SDBR-CMU621 and SDBR-CMU684, formed a distinct clade, sister to Boekhoutia foliicola JCM 36090 and B. sterigmata CBS 15553 (Fig. 25). The D1/D2 sequences of the two strains differed by 4.80% (27 nt substitutions and 6 gaps) from B. foliicola and 4.55% nucleotide divergence (28 nt substitutions and 6 gaps) from B. sterigmata. The ITS sequences of the strains demonstrated divergence with 8.19% (28 nt substitutions and 23 gaps) from B. foliicola and 7.97% (33 nt substitutions and 29 gaps) from B. sterigmata. These findings suggest that the two strains represent a novel species in the Boekhoutia. Furthermore, they can be differentiated by the ability of B. pollinicola to grow in ribose, melibiose, lactose, erythritol, glucitol, D-gluconate, DL-lactate, and succinate. Additionally, B. pollinicola was able to grow at 30 °C, whereas the maximum growth temperature for B. foliicola and B. sterigmata is 25 °C[63,81].
Figure 25.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Chionosphaeraceae. The tree is rooted to Jianyunia sakaguchii (CBS 8464), Pseudosterigmatospora motuoensis (CGMCC 2.5816), and Sterigmatospora layueensis (CGMCC 2.5817). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Twenty-seven strains are included in the combined sequence analysis, which comprise 1,594 characters with gaps. The average standard deviation of the split frequencies of the BI analysis was 0.001890. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Class: Cystobasidiomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw.
Order: Cystobasidiales R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw.
Family: Cystobasidiaceae Gäum.
Member of Cystobasidiaceae have been isolated from diverse environments, including cold climates, Arctic regions, various plant-associated habitats, dung-inhabiting habitats, and mycoparasitic habitats. In yeast form characterized by nano-meterfusion mycoparasitism with tremelloid haustorial cells and septal pores with cystosomes[65,80] the colony have pastel red or orange colors that produce bioactive compounds like carotenoids with antioxidant properties[82]. Currently, six genera are listed in this family, including Cystobasidium, Begerowomyces, Cystastrum, Halobasidium, Robertozyma, and Queiroziella[83]. In this study, Cystobasidium benthicum (one strain), Cys. keelungense (two strains), Cys. minutum (two strains), Cys. thailandicum sp. nov. (two strains), and Halobasidium lannaense sp. nov. (two strains) were presented (Fig. 26).
Figure 26.
Phylogram generated by maximum likelihood analysis of the combined D1/D2 domain of LSU, ITS, SSU, rpb1, rpb2, and tef1-α sequence data representing Cystobasidiaceae (Cystobasidiales) and Symmetrosporaceae (Incertae sedis) in Cystobasidiomycetes. The tree is rooted to Sakaguchia lamellibrachiae (CBS 9598), S. meli (CBS 10797), and S. melibiophila (JCM 8162). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Seventy-seven strains are included in the combined sequence analysis, which comprise 7,211 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Cystobasidium thailandicum Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 27)
MycoBank number: MB860191
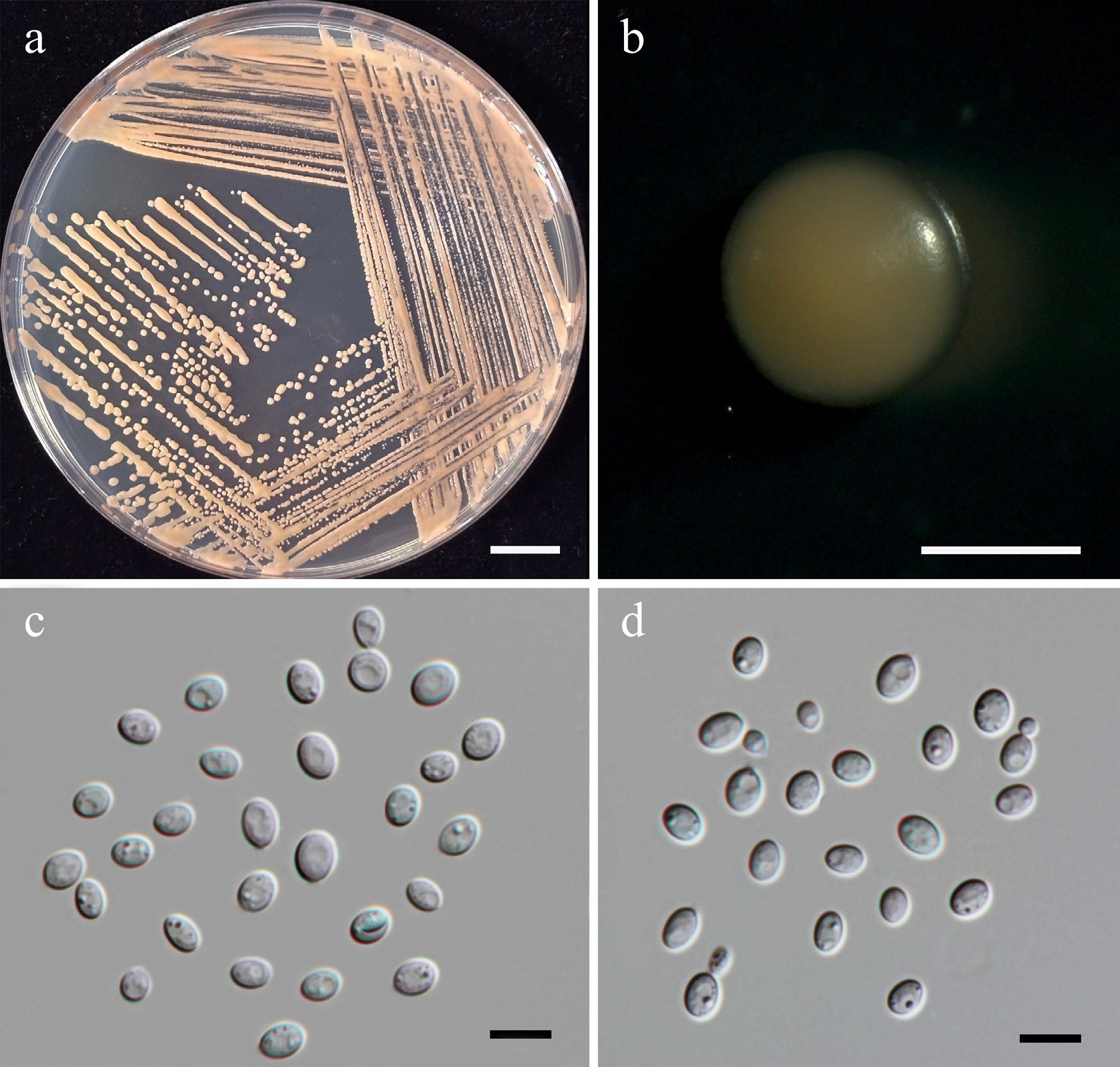
Figure 27.
Morphological characteristics of Cystobasidium thailandicum (SDBR-CMU697, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The species name 'thailandicum' refers to Thailand, the country where the type strain was isolated.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in Golden trumpet flower (Allamanda cathartica), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40111 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU697 = GMBCC2407 = TBRC21414. GenBank numbers PV834516 (D1/D2), PV834686 (ITS), PX622334 (SSU), PX582338 (tef1-α).
Description – Colonies on YMA after five days at 25 °C are circular form (1.5–2 mm in diameter), pastel red, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroidal to ovoidal (4.51–6.89 × 5.47–8.24 μm, n = 50), occur singly and polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for 1 month.
Fermentation of glucose is negative. D-Glucose, galactose, xylose, L-arabinose, sucrose, α-α-trehalose, cellobiose, salicin, lactose, melizitose glycerol, D-glucono-1,5-lactone, D-gluconate, D-galacturonic acid, and succinate are assimilated, but sorbose (or weak), N-acetyl glucosamine, D-arabinose (or weak), maltose, D-ribose, L-rhamnose, methyl-α-D-glucoside, melibiose, raffinose, inulin, soluble starch, erythritol, ribitol, D-glucitol, mannitol, galactitol, myo-inositol, D-glucuronate, DL-lactate, citrate, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfates are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, ethylamine hydrochloride, L-lysine, and cadaverine are not assimilated. No growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in frangipani flower (Plumeria obtusa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU565. GenBank numbers PV834517 (D1/D2), PV834687 (ITS), PX622335 (SSU), PX582339 (tef1-α).
Notes – Cystobasidium thailandicum SDBR-CMU697 and SDBR-CMU565 formed a distinct lineage, closely related to Cys. calyptogenae CBS 9125, CBS 11058, and CBS 11318 in the Cystobasidium clade (Fig. 26). The present strains differed from Cys. calyptogenae by 0.33% nucleotide divergence (2 substitutions) in the D1/D2 domain, 0.89% (5 substitutions) in the ITS region, and 1.96% nucleotide divergence (21 substitutions and 6 gaps) in the tef1-α gene. Based on physiological test, Cys. thailandicum can be distinguished from Cys. calyptogenae by its ability to assimilate L-sorbose, D-arabinose, maltose, raffinose, D-glucitol, D-mannitol, DL-lactate, and citrate. In contrast, Cys. thailandicum does not assimilate cellobiose, lactose, D-glucuronate, and D-galacturonic acid. Likewise, Cys. thailandicum can grow in 50% and 60% glucose medium, whereas Cys. calyptogenae cannot grow under these conditions[84].
Halobasidium lannaense Kodchasee, Senwanna, J. Kumla. & N. Suwannar., sp. nov. (Fig. 28)
MycoBank number: MB860192
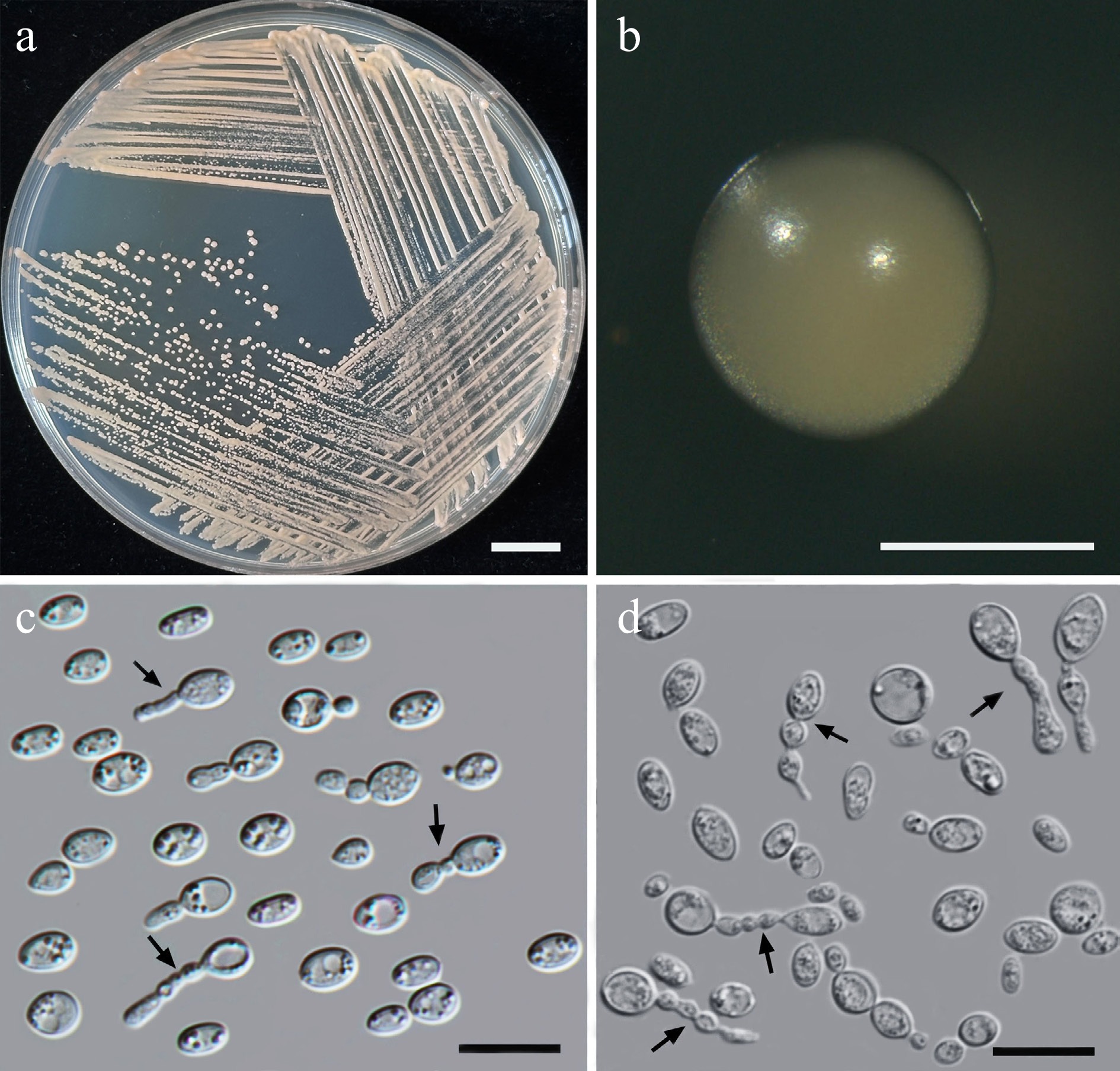
Figure 28.
Morphological characteristics of Halobasidium lannaense (SDBR-CMU612, ex-type). (a) Culture, (b) single colony, (c) chains of cells on YMA after 5 d at 25 °C, and (d) chains of cells (indicated by arrows) on YMA after 3 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'lannaense' referring to the Kingdom of Lanna, the historic name of northern Thailand, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in coloradillo flower (Hamelia patens), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40098 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU612 = GMBCC2408 = TBRC21398. GenBank numbers PV834518 (D1/D2), PV834688 (ITS), PX622336 (SSU), PV941860 (rpb1), PV947457 (rpb2), PV947487 (tef1-α).
Description – Colonis on YMA after 5 d at 25 °C is circular form (1.0–1.5 mm in diameter), pale orange, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroidal to ovoidal (1.69–3.35 × 2.29–5.6 μm, n = 50), occur singly, in pairs, or in chains budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose (or weak), xylose, L-arabinose, D-arabinose (or weak), sucrose, maltose, α-α-trehalose, salicin, raffinose, melizitose, glycerol, D-glucitol (or slow), mannitol, D-glucono-1,5-lactone, D-gluconate, DL-lactate, succinate, and citrate are assimilated, but N-acetyl glucosamine, D-ribose, L-rhamnose, methyl-α-D-glucoside, cellobiose, melibiose, lactose, inulin, soluble starch, erythritol, ribitol, galactitol, myo-inositol, D-glucuronate, D-galacturonic acid, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfate and L-lysine are assimilated as sole nitrogen sources, but potassium nitrate, sodium nitrite, ethylamine hydrochloride, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose, but not on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth at 10, 20, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in bougainvillea (Bougainvillea hybrid), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU639. GenBank numbers PV834519 (D1/D2), PV834689 (ITS), PX622337 (SSU), PX582320 (rpb2), PX582340 (tef1-α).
Notes – Two strains, SDBR-CMU612 and SDBR-CMU639, introduced as a new member of Halobasidium, formed a distinct lineage closely related to Hal. xiangyangense KCTC27810 (Fig. 26). The present strains differed from Hal. xiangyangense by 0.33% nucleotide divergence (2 substitutions) in the D1/D2 domain, 1.28% (7 substitutions) in the ITS region, 6.55% (62 substitutions) in the rpb2 gene, and 2.62% nucleotide divergence (21 substitutions) in the tef1-α gene. Halobasidium lannaense can be distinguished from Hal. xiangyangense with its ability to assimilate raffinose, melizitose, D-glucitol, D-glucono-1,5-lactone, and DL-lactate as well as by its ability to grow on media containing 50% and 60% glucose[85]. Additionally, Hal. lannaense also produced chains of two or three buds on YMA at 25 °C for 5 d (Fig. 28c), and developed elongated buds after 3 weeks (Fig. 28d), which is the unique character of Halobasidium, but not present Y-shaped groups of cells like Hal. xiangyangense[85].
Class: Cystobasidiomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss, & Oberw.
Order: Erythrobasidiales R. Bauer, Begerow, J.P. Samp., M. Weiss, & Oberw.
Family: Erythrobasidiaceae Denchev
Erythrobasidiaceae represents a small but distinct family of basidiomycetous yeasts that are primarily characterized by a pink to red color, which is reflected in the name 'Erythro' (meaning red). Cells are ellipsoidal and cylindrical. They are primarily of the genus type found in the phyllosphere[65,86]. Currently, two genera are listed in this family, including Bannoa and Erythrobasidium[41]. In this study, Erythrobasidium primogenitum (one strain) is presented (Supplementary File 1).
Families Incertae sedis
Family: Symmetrosporaceae Q.M. Wang, F.Y. Bai, M. Groenew, & Boekhout
Symmetrospora species have been isolated from soil environments and leaf surfaces and other habitats, indicating broad ecological adaptation across terrestrial ecosystems[87]. The colonies produce entire margins and colony color varies from pink to brick-red, budding cells present. Currently, one genus is Symmetrospora listed in this family[41]. In this study, Sy. chiangmaiensis sp. nov. (three strains), and Sy. hydei sp. nov. (two strains) are presented (Fig. 26).
Symmetrospora chiangmaiensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 29)
MycoBank number: MB860193
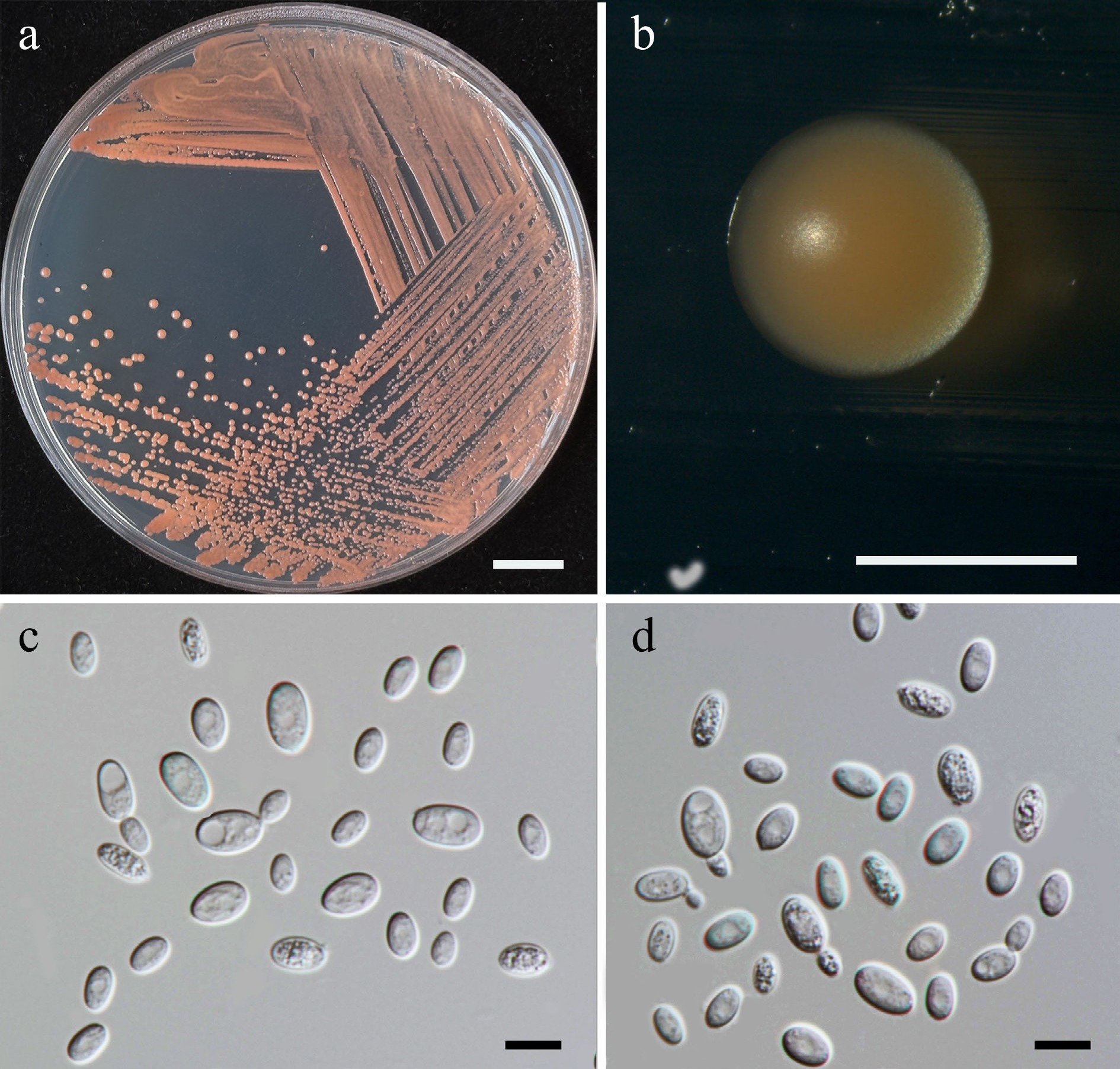
Figure 29.
Morphological characteristics of Symmetrospora chiangmaiensis (SDBR-CMU715, ex-type). (a) Culture, (b) single colony, and (c), (d) cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'chiangmaiensis' referring to Chiang Mai Province, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in marguerite daisy flower (Argyranthemum frutescens), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40114 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU715 = GMBCC2409 = TBRC21417. GenBank numbers PV834522 (D1/D2), PV834692 (ITS), PX622340 (SSU), PX582341 (tef1-α).
Description – Colonies on YMA after 5 d at 25 °C are circular form (0.8–1.2 mm in diameter), reddish orange, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroidal to ovoidal (3.43–6.72 × 6.17–11.48 μm, n = 50), occur singly and multilateral budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, sorbose, ribose, xylose, DL-arabinose, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, lactose, raffinose, melizitose, glycerol, ribitol, D-glucitol, mannitol, D-glucono-1,5-lactone (weak), D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, and xylitol are assimilated, but N-acetyl glucosamine, salicin, melibiose, inulin, soluble starch, erythritol, galactitol, myo-inositol, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine, and cadaverine (weak) are assimilated as sole nitrogen sources. No growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth at 10, 15, 25, and 30 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in watrakanu flower (Ruellia tuberosa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU570; Chang Phueak, in Persian gentian flower (Exacum affine), July 2024, P. Kodchasee, C. Senwanna, J. Kumla. and N. Suwannarach, living culture SDBR-CMU579. GenBank numbers SDBR-CMU570: PV834520 (D1/D2), PV834690 (ITS), PX622338 (SSU); SDBR-CMU579: PV834521 (D1/D2), PV834691 (ITS), PX622339 (SSU).
Notes – Symmetrospora chiangmaiensis, SDBR-CMU715, SDBR-CMU570 and SDBR-CMU579, formed a sister clade, with Sy. proteacearum, BRIP 45084, BRIP 763649a, and Sy. pseudomarina, CBS 14057 (Fig. 26). The D1/D2 sequences of these three strains were differed by 0.18%–0.34% nucleotide divergence (1–2 nt substitutions) and the ITS sequences of the three strains demonstrated divergence (7–9 nt substitutions) more than 1.23%–1.62%. Additionally, a distinct comparison of the phenotypic characteristics between Sy. chiangmaiensis and Sy. pseudomarina are shown in Table 8.
Table 8. Phenotypic characteristics differentiating Symmetrospora chiangmaiensis and Sy. hydei from closely related Symmetrospora species.
Characteristics 1 2 3 4 5 6 Carbon assimilation L-Sorbose + w l/w + – w D-Ribose + + l/w + + – D-Xylose + + + + + – D-Arabinose + + l/w + + – L-Rhamnose + – l/w + v – Maltose + + + + – – Methyl-α-D-glucoside + + l/w + + – Salicin – w w + nd – Melibiose – – – + – + Lactose + + – – + – Raffinose + + l/w + nd – Melizitose + + l/w + + – D-Glucuronate + – + + nd nd D-Galacturonic acid + – + – nd nd DL-Lactate + + l/w + nd – Citrate + – + + nd nd Xylitol + w nd nd nd nd Nitrogen assimilation Potassium nitrate + w – – v + Sodium nitrite + w nd nd nd – Ethylamine HCl + w – – nd – L-Lysine + w – – + + Cadaverine w w – – v – Growth characteristics Growth at 30 °C + – – + v – Growth at 35 °C – – – + – – Strains 1: Sy. chiangmaiensis sp. nov, 2: Sy. hydei sp. nov, 3: Sy. coprosmae, 4: Sy. oryzicola[88], 5: Sy. pseudomarina[87], and 6: Sy. rhododendri[63]. Symmetrospora hydei Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 30)
MycoBank number – MB860194
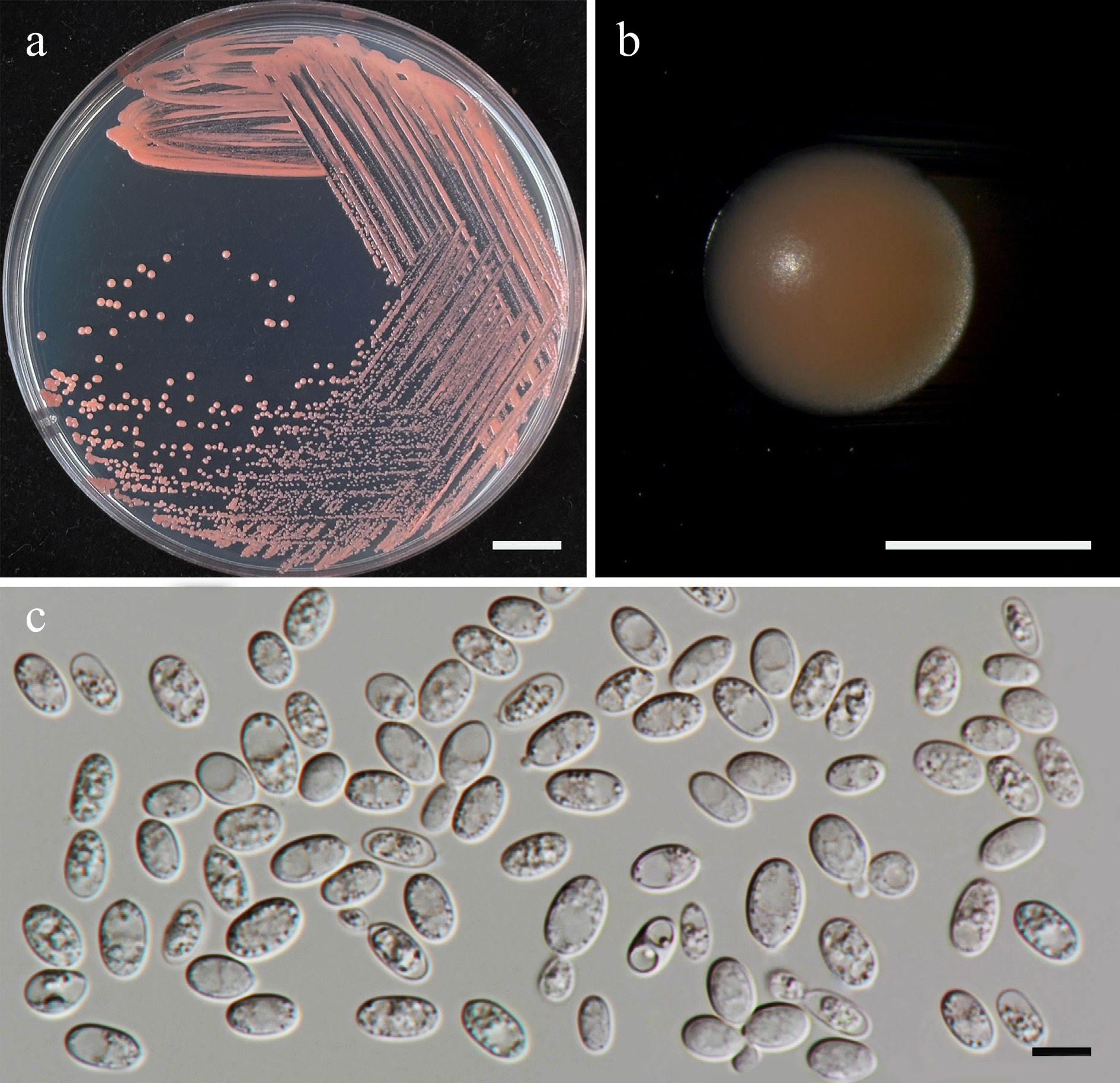
Figure 30.
Morphological characteristics of Symmetrospora hydei (SDBR-CMU620, ex-type). (a) Culture, (b) single colony, and (c) cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c) = 10 μm.
Etymology – 'hydei' referring to the name of Kevin D. Hyde in honor of his 70th birthday.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in Yellow-flowered waterhyssop (Mecardonia procumbens), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40100 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU620 = GMBCC2410 = TBRC21401. GenBank numbers PV834523 (D1/D2), PV834693 (ITS), PX622341 (SSU), PX582321 (rpb2), PX582342 (tef1-α).
Description – Colonies on YMA after 5 d at 25 °C are circular form (0.8–1.2 mm in diameter), orange red or yellowish red, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are spheroidal to ovoidal (6.02–9.31 × 7.01–13.35 μm, n = 50), occur singly and polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, D-galactose, sorbose (weak), ribose, xylose, DL-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin (weak), lactose, raffinose, melizitose, glycerol, ribitol, D-glucitol, mannitol, D-gluconate, DL-lactate, succinate, and xylitol (weak) are assimilated, but N-acetyl glucosamine, L-rhamnose, melibiose, inulin, soluble starch, erythritol, galactitol, myo-inositol, D-glucono-1,5-lactone D-glucuronate, D-galacturonic acid, citrate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate (weak), sodium nitrite (weak), ethylamine (weak), L-lysine (weak), and cadaverine (weak) are assimilated as sole nitrogen source. No growth occurs on media containing 50% glucose, 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, and 25 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in chili flower (Capsicum sp.), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU703. GenBank numbers PV834524 (D1/D2), PV834694 (ITS), PX622342 (SSU), PX582322 (rpb2), PX582343 (tef1-α).
Notes – Symmetrospora hydei formed a distinct lineage and was sister to Sy. coprosmae, Sy. oryzicola, and Sy. rhododendri with 100% BSML and 1 BYPP (Fig. 26). However, Sy. hydei differed from those three species by 0.64%, 0.79%, and 0.95% nucleotide divergence (4, 5, and 6 nt substitutions) in the D1/D2 domain, respectively, and by 0.87, 0.70, and 1.91% nucleotide divergence (5, 4, and 11 nt substitutions), respectively, in the ITS region. Additionally, the present strain is different from Sy. coprosmae and Sy. oryzicola by 4.77%–6.69% and 4.05%–5.72% nucleotide divergence in rpb2 and tef1-α, respectively. Moreover, distinguishing characteristics between Sy. hydei and Sy. coprosmae, Sy. oryzicola, and Sy. rhododendri are given in Table 8.
Class: Microbotryomycetes R. Bauer, Begerow, J.P. Samp., M. Weiss & Oberw.
Order incertae sedis
Families incertae sedis
Currently, 17 genera are listed in this Microbotryomycetes genera incertae sedis, including Atractocolax, Baiomyces, Curvibasidium, Fengyania, Litoriozyma, Nakaseozyma, Oberwinklerozyma, Pseudohyphozyma, Pseudoleucosporidium, Reniforma, Sampaiozyma, Slooffia, Spencerozyma, Trigonosporomyces, Udeniozyma, Vonarxula, and Yunzhangia[41,81]. In this study, Curvibasidium chiangmaiense sp. nov. (two strains) was presented.
Curvibasidium chiangmaiense Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 31)
MycoBank number: MB860195
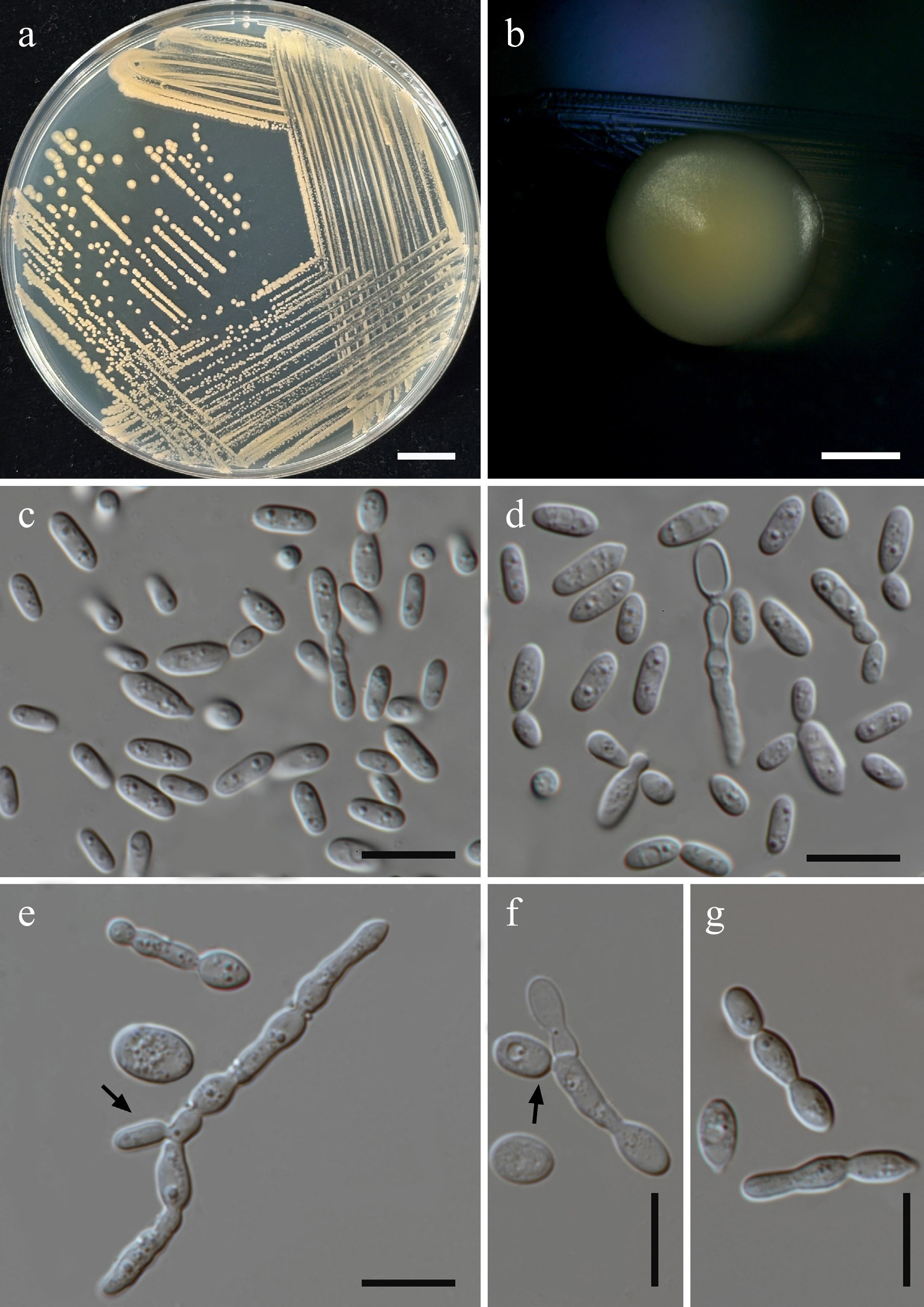
Figure 31.
Morphological characteristics of Curvibasidium chiangmaiense (SDBR-CMU574, ex-type). (a) Culture, (b) single colony, and (c), (d) cells on YMA after 5 d at 25 °C. (e), (f) Pseudohyphae and blastoconidia (indicated by arrows) on PDA after 2 weeks at 25 °C, and (g) elongate cells. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c)–(g) = 10 μm.
Etymology – The specific epithet 'chiangmaiense' referring to Chiang Mai Province, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in Siam tulip flower (Curcuma sessilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40089 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU574 = GMBCC2411 = TBRC21389. GenBank numbers PV834526 (D1/D2), PV834696 (ITS), PV819892 (SSU), PX582298 (rpb1), PX582323 (rpb2), PV947473 (tef1-α).
Description – Colonies on YMA after five days at 25 °C are circular form (1.8–2 mm in diameter), light orange, smooth surface, smooth, butyrous, with margin slightly fringed. The cells are spheroidal to ovoidal (1.82–3.59 × 4.9–8.89 μm, n = 50), occur singly and polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, L-arabinose, sucrose, maltose, α-α-trehalose, cellobiose, salicin, melizitose, ribitol, D-glucitol, mannitol, galactitol, D-glucono-1,5-lactone, D-gluconate, and succinate are assimilated, but xylose, N-acetyl glucosamine, D-arabinose, D-ribose, L-rhamnose, methyl-α-D-glucoside, melibiose, lactose, raffinose, inulin, soluble starch, glycerol, erythritol, myo-inositol, D-glucuronate, D-galacturonic acid, DL-lactate, citrate, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfates, ethylamine hydrochloride, and L-lysine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, and cadaverine are not assimilated. Growth on media containing 50% but not on media containing glucose, 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is positive. Growth on 10, 15, 25, 30, and 35 °C but absent at 37 and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in rose flower (Rosa sp.), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU578. GenBank numbers PV834527 (D1/D2), PV834697 (ITS), PV819893 (SSU), PX582299 (rpb1), PX582324 (rpb2), PV947474 (tef1-α).
Notes – Phylogenetic analyses of a concatenated D1/D2 domain, ITS, SSU, rpb1, rpb2, and tef1-α sequence dataset revealed that Curvibasidium chiangmaiense, SDBR-CMU574 and SDBR-CMU578, formed a separate clade, clustering with Cu. nothofaggi CBS 8166 and Cu. pallidicorallinum CBS9091 (Fig. 32). A comparison of D1/D2 domain and ITS sequences indicated that our strains are not significantly different from Cu. nothofaggi (2 nt substitutions in D1/D2 and 1 nt substitutions in ITS), and Cu. pallidicorallinum (5 nt substitutions, including 2 gaps in D1/D2 and 0 nt substitutions in ITS). However, Cu. chiangmaiense is different from Cu. nothofaggi by 7.86% nucleotide divergence (38 nt substitutions) in rpb1, 7.32% nucleotide divergence (78 nt substitutions) in rpb2, and 3.86% nucleotide divergence (55 nt substitutions) in tef1-α, respectively. Likewise, Cu. chiangmaiense is different from Cu. pallidicorallinum by 9.14.% nucleotide divergence (37 nt substitutions) in rpb1, 7.73% nucleotide divergence (84 nt substitutions) in rpb2, and 3.84% nucleotide divergence (64 nt substitutions) in tef1-α, respectively. Thus, the novel species, Cu. chiangmaiense, is introduced from Curcuma sessilis and Rosa sp. in Thailand. Additionally, distinguishing characteristics between Cu. chiangmaiense and another related species are given in Table 9.
Table 9. Phenotypic characteristics differentiating Curvibasidium chiangmaiense and closely related species.
Characteristics 1 2 3 4 5 6 7 8 9 Carbon assimilation D-Galactose + + + s + – – – – L-Sorbose + + + + + + + – – D-Ribose – + – w – – – + w D-Xylose – + + s + – + + – L-Arabinose + + – s + + – + + D-Arabinose – + – w s – – + w Sucrose + – – – – + + + + Maltose + – – – – – + + + α-α-Trehalose + – – – – + v – + Methyl-α-D-glucoside – – – – – + + w w Salicin + + + + + + – + + Melibiose – – – – – + – nd – Raffinose – – – + – – – – – Melizitose + – – – nd + + nd + Inulin – – – – nd + + nd + Soluble starch – – – – – + w nd + Glycerol – – – s nd w + – – D-Glucitol + + + + + + – – – Galactitol + + + + + – – – – DL-Lactate – – – s – + + + + Succinate + + + + + + + + + Citrate – + + s – + – – – Ethanol – + + + + + + + + Nitrogen assimilation Potassium nitrate – – + – – + + + + Sodium nitrite – – – – nd + w – w L-Lysine + + + + + – + + + Cadaverine – + – – – + + w + Growth characteristics Growth at 15 °C + nd nd nd nd 17 °C nd nd 17 °C Growth at 25 °C + + + + + nd nd + nd Growth at 30 °C + – – – – – nd – – Growth at 35 °C + – nd – nd nd nd nd nd Growth in 0.01% Cycloheximide – nd + + – nd nd nd nd Strains 1: Cu. chiangmaiense sp. nov, 2: Cu. cygneicollum, 3: Cu. nothofagi, 4: Cu. pallidicollinum[89], 5: Cu. rogersii[90], 6: Baiomyces sejilaensis, 7: Fengyania pteridophytorum, 8: Nakaseozyma junci, and 9: N. lonicerae[81]. Thailandicolales Kodchasee, Senwanna, J. Kumla, & N. Suwannar., ord. nov.
MycoBank number: MB860196
Etymology – 'Thailandicolales' reference to the name of the host country Thailand.
Description – Member of Microtryomycetes. The diagnosis of the order Thailandicolales is based on the description of the genus Thailandicola. The nomenclature of the order is based on the genus Thailandicola.
Type family – Thailandicolaceae Kodchasee, Senwanna, J. Kumla, & N. Suwannar.
Thailandicolaceae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., fam. nov.
MycoBank number: MB860197
Etymology – 'Thailandicolaceae' refers to the name of the host country Thailand.
Description – Member of Thailandicolales (Microtryomycetes). The diagnosis of the family Thailandicolaceae is based on the description of the genus Thailandicola. The nomenclature of the family is based on the genus Thailandicola.
Type genus – Thailandicola Kodchasee, Senwanna, J. Kumla & N. Suwannar., gen nov.
Thailandicola Kodchasee, Senwanna, J. Kumla, & N. Suwannar., gen. nov.
MycoBank number: MB860198
Etymology – 'Thailandicola' refers to the name of the host country Thailand.
Type species – Thailandicola limtongiae Kodchasee, Senwanna, J. Kumla, & N. Suwannar.
Description – The colonies are circular pale yellow, and smooth surface. The cells are ellipsoidal, budding is polar. Pseudohyphae are produced. Basidiospores have not been observed in individual or in mixed cultures. Fermentation ability is absent. Nitrate and nitrite are not assimilated.
Notes – Thailandicola is introduced as a new genus based on morphology and multi-locus phylogenetic support. A BLASTn search using the D1/D2 sequence of SDBR-CMU552, SDBR-CMU561, SDBR-CMU635, and SDBR-CMU662 had a less than 90% similarity with other taxa in the Microbotryomycetes and 80%–90% (60%–100% coverage) in the ITS region. These differences indicate that the strains represent a new member of the Microbotryomycetes. Based on the combined multi-locus dataset (D1/D2, ITS, SSU, rpb1, rpb2, and tef1-α; Fig. 32), demonstrated that Thailandicola formed an independent clade basal to Microbotryales, Kriegeriales, Heterogastridiales, and Leucosporidiales with 100% MLBS and 1 BYPP support values.
Thailandicola hydei Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 33)
MycoBank number: MB860199
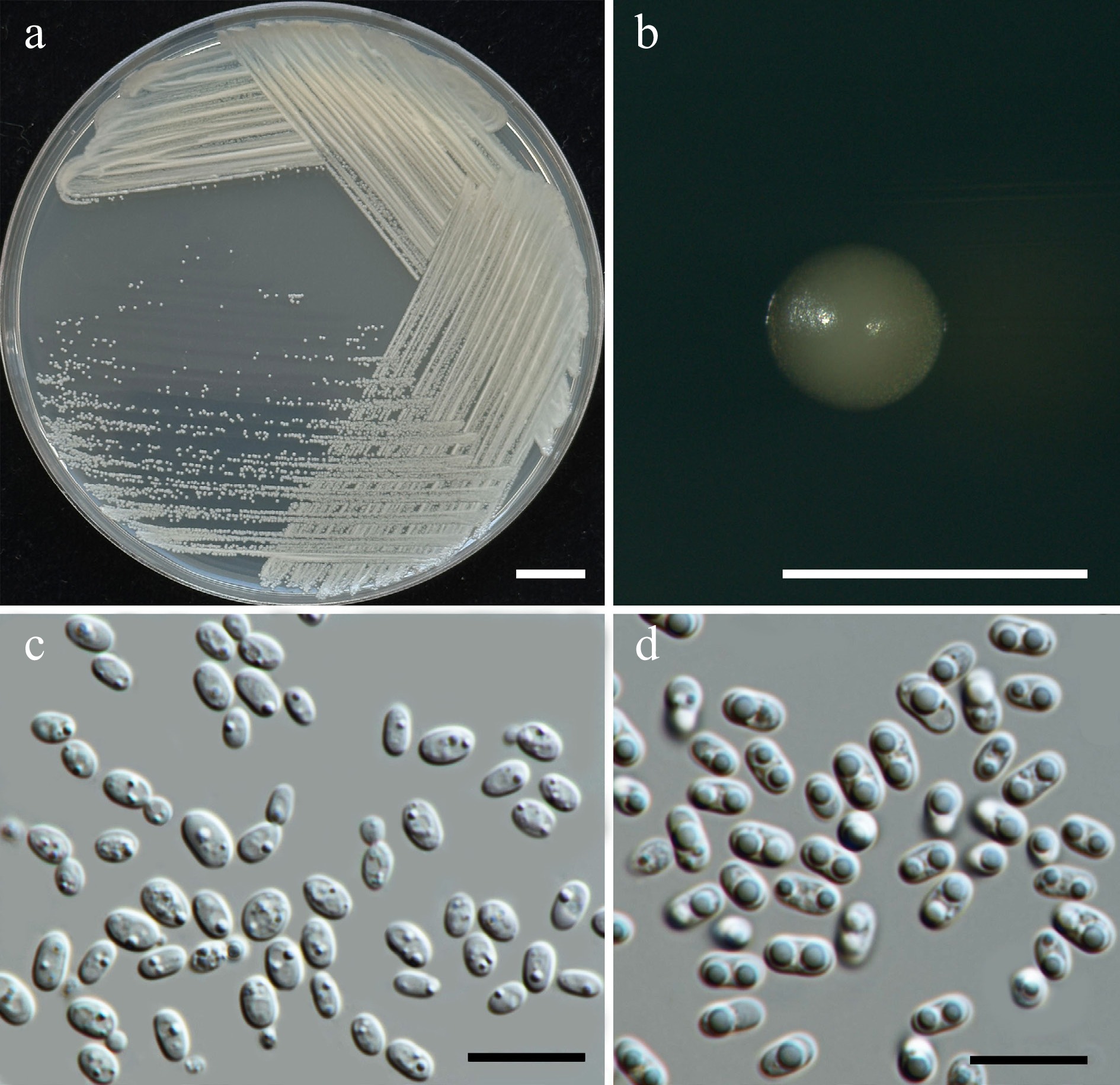
Figure 33.
Morphological characteristics of Thailandicola hydei (SDBR-CMU662, ex-type). (a) Culture, (b) single colony, (c), (d) cells and budding cells on YMA after 5 d at 25 °C. (d) Vegetative cells with one or two large globules on PDA at 25 °C for 2 weeks. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'hydei' referring to the name of Kevin D. Hyde in honor of his 70th birthday.
Holotype – THAILAND, Phayao Province, Mueang District, in vinca flower (Catharanthus roseus), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40105 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU662 = GMBCC2412 = TBRC21407. GenBank numbers PV834529 (D1/D2), PV834699 (ITS), PV819895 (SSU), PV941862 (rpb1), PV947463 (rpb2), PV947480 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.0 mm in diameter), pale yellow, smooth surface, glistening apperance, entire margin, and convex elevation. The cells are ellipsoidal (2.41–4.18 × 3.61–5.3 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced pseudohypae, and true hyphae were observed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, N-acetyl glucosamine (or weak), sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, raffinose, melizitose, D-glucono-1,5-lactone, and DL-lactate (or weak or slow) are assimilated, but D-galactose, sorbose, ribose, xylose, L-arabinose, D-arabinose, L-rhamnose, melibiose, lactose, inulin, soluble starch, glycerol, erythritol, ribitol, D-glucitol, mannitol, galactitol, myo-inositol, D-gluconate, citrate, methanol, ethanol, and xylitol are not assimilated. Assimilation is variable for salicin, D-glucuronate, D-galacturonic acid, and succinate. Ammonium sulfate and lysine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, ethylamine, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose. No growth occurs on media containing 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in Chinese ixora flower (Ixora chinensis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU561. GenBank numbers PV834528 (D1/D2), PV834698 (ITS), PV819894 (SSU), PV941861 (rpb1), PV947462 (rpb2), PV947479 (tef1-α).
Notes – In a BLASTn search revealed that the D1/D2 sequence of the Th. hydei SDBR-CMU662 and SDBR-CMU561 had 90% similarity with Oberwinklerozyma yarrowii CBS 7417, type strain whereas the ITS sequences were closely related to Fellozyma inositophila CBS 7310. Based on multi-locus phylogenetic analyses (Fig. 32), Th. hydei formed a sister clade to Th. limtongiae. Comparing the phenotypic characteristics of Th. hydei, Th. limtongiae, and another related specie are given in Table 10.
Table 10. Phenotypic characteristics differentiating Thailandicola hydei, Th. limtongiae and closely related species in order Heterogastridiales Kriegeriales, Leucosporidiales, and Microbotryales.
Characteristics/order Thailandicolales Heterogastridiales Kriegeriales Leucosporidiales Leucosporidiales Incertae sedis 1 2 3 4 5 6 7 8 9 10 11 12 13 Carbon assimilation D-Galactose s/w – – + w – + + – – – + – L-Sorbose – – – + w + + s w s – + + N-Acetyl glucosamine + w/+ nd nd nd nd nd nd – nd nd nd nd D-Ribose – – – + + + +/l s – s/w – – + D-Xylose – – – + w + +/l + – + – + + L-Arabinose – – – + w + – + – – – +/– – D-Arabinose – – – v + + +/l – – – – – + L-Rhamnose + – – + w + – – – – – – – Sucrose + + – + + + – + + + + – + Maltose + + – + + + – + + + +/l – + α-α-Trehalose + + – + + + +/l + + + +/l + + Methyl-α-D-glucoside + + nd + + nd – – + + – – + Cellobiose w/+ + v + + + +/l – + + – – + Salicin + v nd + + + – – – + – – + Melibiose – – – – – + – – – – – – – Lactose – – – – – + – – – + – – + Raffinose – + – – – + – – +/l – – – + Melizitose + + – + + + – + + + + – + Inulin – – nd – – + – + + v v – – Soluble starch – – – – – + – – w – – – + Glycerol w – – + + + + + w + + + + Erythritol – – – – nd + – – – – – – – Ribitol – – – + w + +/l + +/l s/w nd – + D-Glucitol – – nd + – + + + – + nd + + D-Mannitol + – + + + + + + + + + + + Galactitol – – nd + – + – – – – nd – – myo-Inositol – – – – – + – – – – nd – – D-Glucono-1,5-lactone + + nd nd + + +/l + nd nd nd +/l + D-Gluconate w – nd + nd + nd + nd – + nd + D-Glucuronate + v nd + nd + nd + – nd nd nd + D-Galacturonic acid s v nd nd nd nd nd nd nd nd nd nd + DL-Lactate s s/w nd + + + – s w – – nd + Succinate + v nd + – + + + + s/w – + + Citrate s – + + – + + + + – – – + Methanol – – nd – – – nd nd – – + – – Ethanol – – nd + w + + + – – nd nd – Xylitol – – nd v w nd nd – nd nd nd nd + Nitrogen assimilation Ammonium sulfate + + nd nd nd nd + nd + + nd nd nd Potassium nitrate – – – + – + – + + + – + + Sodium nitrite – – nd + – + – – + – nd nd + Ethylamine HCl – – nd nd + + + + + + nd nd – L-Lysine + + nd nd w + l + + – nd nd + Cadaverine – – nd nd nd + + + + – nd nd – Growth characteristics Growth at 25 °C + + nd + + + + + + + + + + Growth at 30 °C + + nd – + nd nd + + + + + + Strains 1: Th. hydei sp. nov, 2: Th. Limtongiae sp. nov., 3: Reniforma strues[93], 4: Kriegeria eriophori, 5: Meredithblackwellia eburnean[94], 6: Leucosporidium fellii[95], 7: Leu. Intermedium[96], 8: Leu. Krtinense[97], 9: Bauerozyma artemisiae[81], 10: Microbotryozyma swertiae[63], 11: Yunzhangia auriculariae, 12: Yu. sonckii, and 13: Sampaiozyma vanillic[65]. Thailandicola limtongiae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 34)
MycoBank number: MB860200
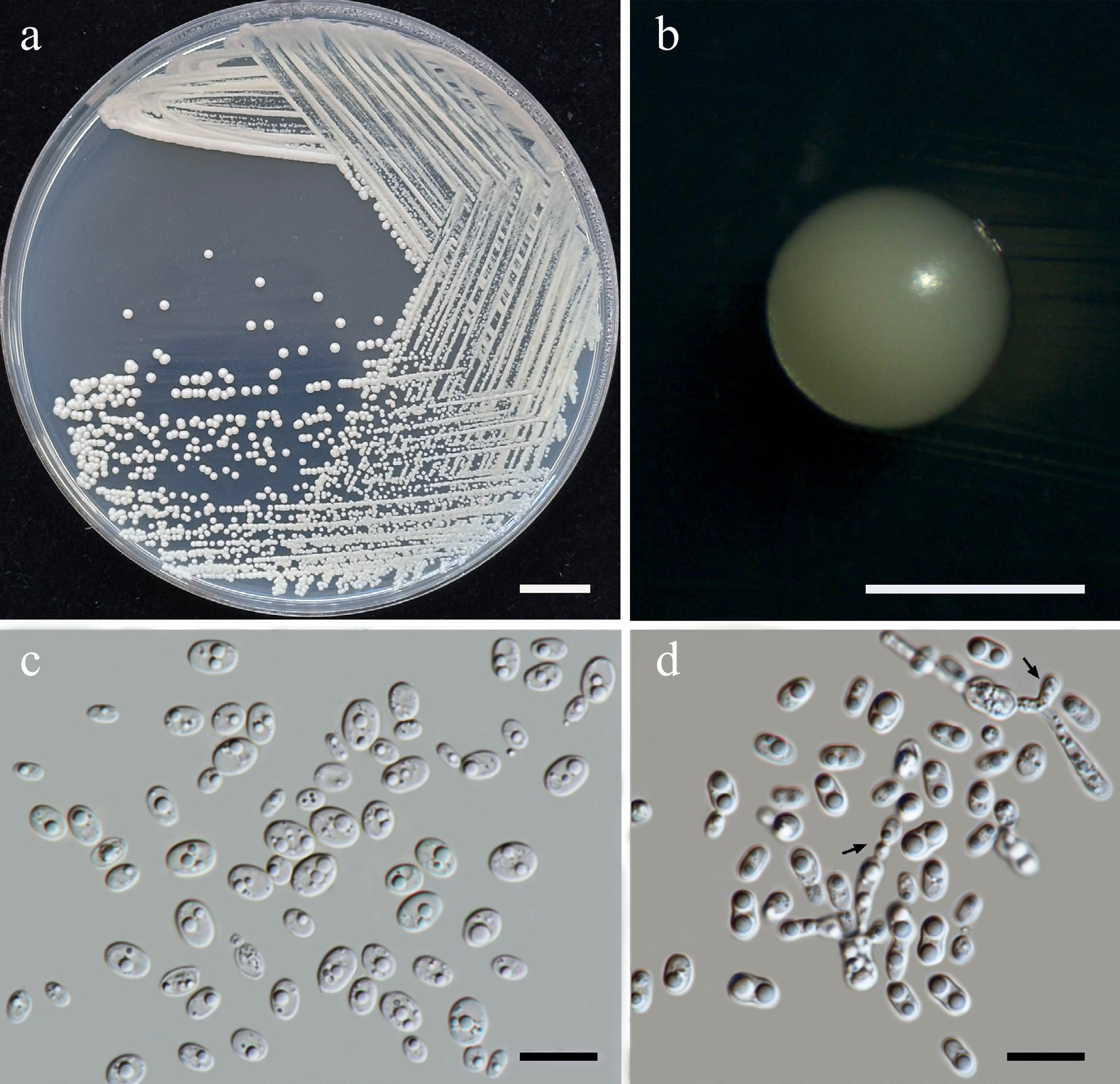
Figure 34.
Morphological characteristics of Thailandicola limtongiae sp. nov. (SDBR-CMU635, ex-type). (a) Culture, (b) single colony, (c) cells and budding cells on YMA after 5 d at 25 °C. (d) Vegetative cells with one or two large globules and pseudohyphae (indicated by arrows) on PDA at 25 °C for 2 weeks. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'limtongiae' named in honor of mycologist Savitree Limtong, for her contributions to yeast systematics.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep, in American cassia flower (Senna spectabilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40104 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU635 = GMBCC2413 = TBRC21405. GenBank numbers PV834531 (D1/D2), PV834701 (ITS), PV819897 (SSU), PV941864 (rpb1), PV947465 (rpb2), PV947482 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.0 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ellipsoidal and ovoid (1.98–4.29 × 2.73–5.98 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. Pseudohyphae were observed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose (or weak or slow), N-acetyl glucosamine, L-rhamnose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose (or weak), salicin, melizitose, glycerol (or weak), mannitol, D-glucono-1,5-lactone, D-gluconate (or weak), D-glucuronate, D-galacturonic acid (or slow), DL-lactate (or slow), succinate, and citrate (or slow) are assimilated, but sorbose, ribose, xylose, L-arabinose, D-arabinose, melibiose, lactose, raffinose, inulin, soluble starch, erythritol, ribitol, D-glucitol, galactitol, myo-inositol, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfate and lysine are assimilated as sole nitrogen source, but potassium nitrate, sodium nitrite, ethylamine, and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in West Indian jasmine (Ixora sp.), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU552. GenBank numbers PV834530 (D1/D2), PV834700 (ITS), PV819896 (SSU), PV941863 (rpb1), PV947464 (rpb2), PV947481 (tef1-α).
Notes – In a BLASTn search revealed that the D1/D2 sequence of the Th. limtongiae SDBR- SDBR-CMU635 and SDBR-CMU552 had 90% similarity with Rhodotorula mucilaginosa CBS 316, type strain whereas the ITS sequences were closely related to Meredithblackwellia eburnea CBS 12589. Based on phylogenetic analysis of a multi-locus dataset (Fig. 32), Th. limtongiae from a sister clade to Th. hydei. Likewise, the phenotypic differences of Th. limtongiae can be distinguished from Th. hydei (Table 10).
Order: Sporidiobolales Doweld
Family: Sporidiobolaceae R.T. Moore
Sporidiobolacae species occur worldwide and have been isolated from a wide variety of substrates, including air, tree leaves, and organic matter. Members of this family are known as decomposers and for their ability to thrive in diverse environmental conditions. Additionally, some species in this family are economically important due to their role as sources of biotechnological products. Their ability to utilize waste materials in the synthesis of carotenoids, lipids, and enzymes makes them valuable for applications in the food, pharmaceutical, and industrial applications[91,92]. Sporidiobolacae represents exhibit spherical to elongate, reproduces by budding, some species form pseudohyphae and produce basidiospores. The colony present salmon-pink to red[82]. Currently, three genera are listed in this family including Rhodosporidiobolus, Rhodotorula, and Sporobolomyces[41]. In this study, Rhodosporidiobolus fluvialis (three strains), R. ruineniae (three strains), Rhodotorula paludigena (one strain), Rh. toruloides (one strain), Rh. thailandensis sp. nov. (three strains), and Sporobolomyces thailandensis sp. nov. (three strains) were presented (Fig. 35).
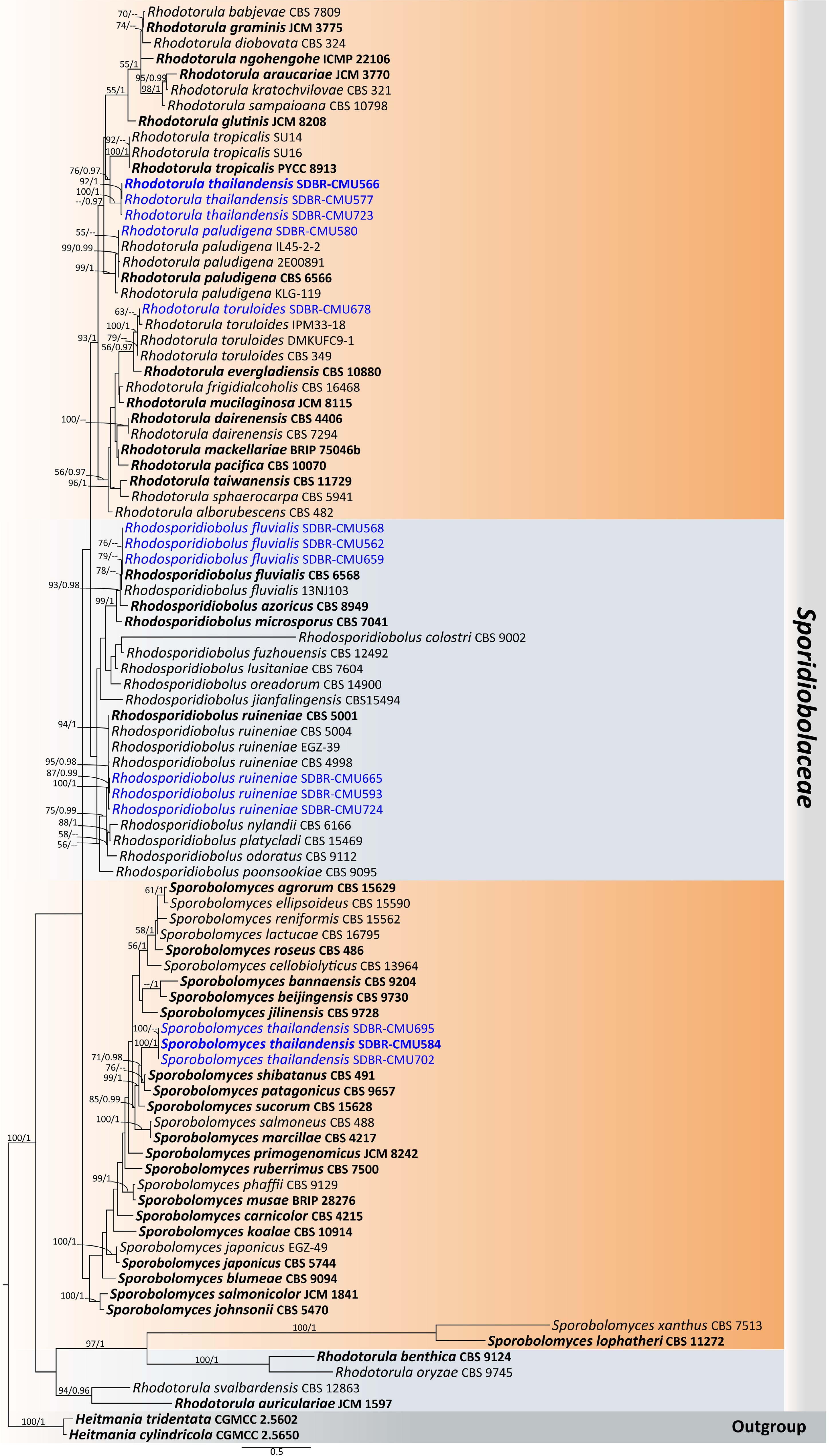
Figure 35.
Phylogram generated by maximum likelihood analysis of the combined D1/D2 domain of LSU, ITS, SSU, rpb1, rpb2, and tef1-α sequence data representing Sporidiobolaceae. The tree is rooted to Heitmania cylindrica (CGMCC 2.5650) and H. tridentata (CGMCC 2.5602). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Ninety-two strains are included in the combined sequence analysis, which comprise 7,114 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Rhodotorula thailandensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 36)
MycoBank number: MB 860201
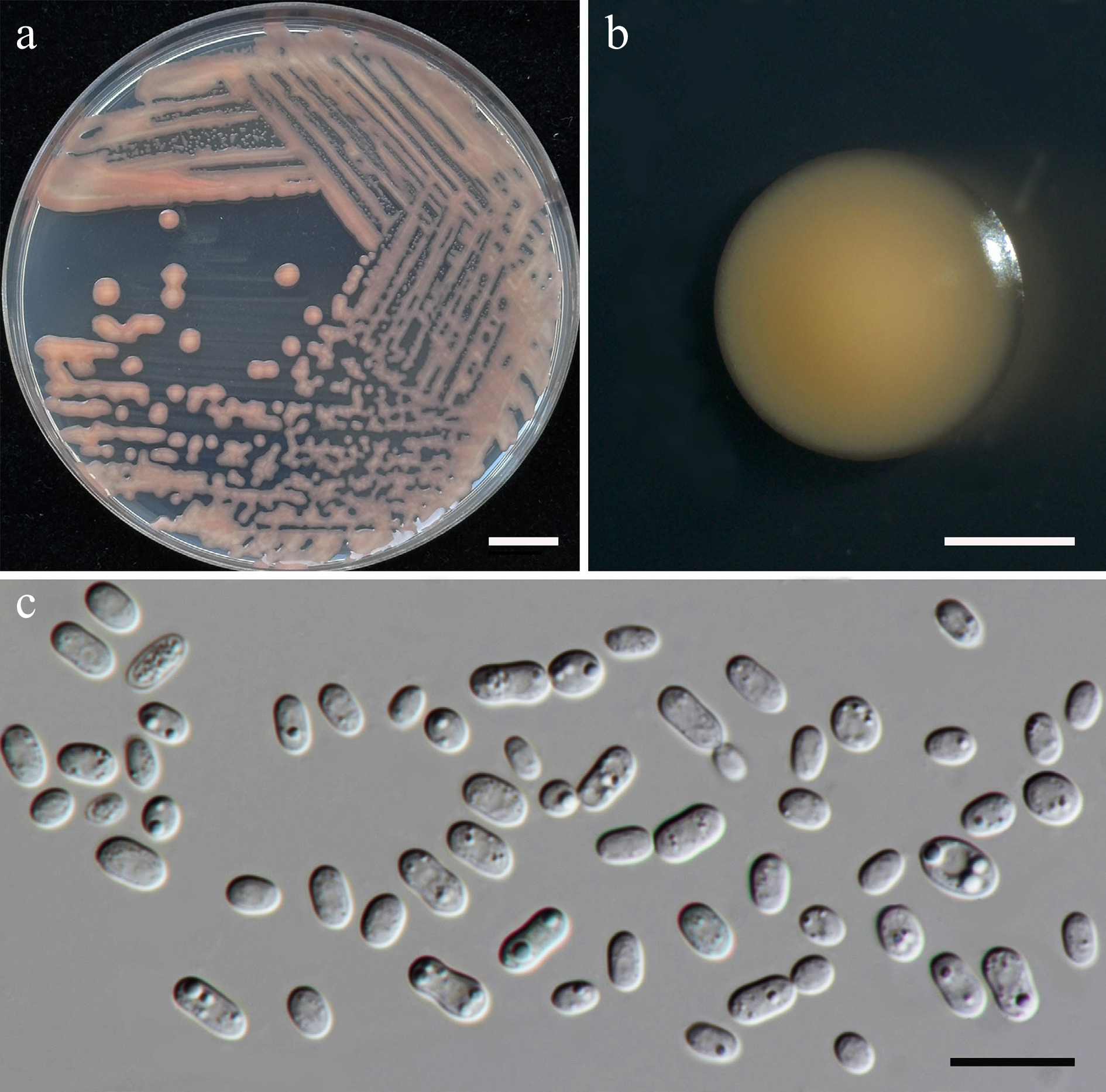
Figure 36.
Morphological characteristics of Rhodotorula thailandensis (SDBR-CMU566, ex-type). (a) Culture, (b) single colony, (c) cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c) = 10 μm.
Etymology – The species name 'thailandensis' refers to Thailand, the country where the type strain was isolated.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep in Fukien tea flower (Carmona retusa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40087 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU566 = GMBCC2414 = TBRC21387. GenBank numbers PV834539 (D1/D2), PV834709 (ITS), PX622343 (SSU), PV941865 (rpb1), PV947466 (rpb2), PV947483 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.0–2.0 mm in diameter), light orange or orange, white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ellipsoidal and cylindrical (1.66–3.09 × 2.75–5.90 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, ribose, xylose, L-arabinose (or slow), L-rhamnose (or weak), sucrose, α-α-trehalose, cellobiose, salicin, raffinose, glycerol, ribitol, D-glucitol (or slow), mannitol (or slow), galactitol (or slow), D-glucono-1,5-lactone, succinate, citrate, and xylitol are assimilated, but D-galactose, sorbose, N-acetyl glucosamine, melibiose, melizitose, inulin, soluble starch, erythritol, myo-Inositol, D-gluconate, D-glucuronate, D-galacturonic acid, methanol, and ethanol are not assimilated. Assimilation is variable for DL-lactate, D-arabinose, maltose, methyl-α-D-glucoside, and lactose. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, and L-lysine are assimilated as sole nitrogen sources, but cadaverine is not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Mae Hia, in Siam tulip flower (Curcuma sessilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU577; Doi Saket District, Samran Rat, in Chinese honey suckle (Combretum indicum) flower, August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU723. GenBank numbers SDBR-CMU577: PV834540 (D1/D2), PV834710 (ITS), PX622344 (SSU), PV941866 (rpb1), PV947467 (rpb2), PV947484 (tef1-α); SDBR-CMU723: PV834541 (D1/D2), PV834711 (ITS), PX622345 (SSU), PV941867 (rpb1), PV947468 (rpb2), PV947485 (tef1-α).
Notes – Rhodotorula thailandensis SDBR-CMU566, SDBR-CMU577 and SDBR-CMU723 were placed in the Rhodotorula clade (Fig. 35), and were closely related to Rh. tropicalis PYCC 8913 by 4.32% nucleotide divergence (26 nt substitutions) in D1/D2 regions and 5.92% nucleotide divergence (35 nt substitutions and 11 gaps) in the ITS region, which indicated that they represent a new species of Rhodotorula. Physiologically, Rh. thailandensis differs from Rh. tropicalis in its inability to galactose and sorbose. Additionally, Rh. thailandensis was able to grow on 50% and 60% glucose, while Rh. tropicalis could not grow at that medium[98].
Sporobolomyces thailandensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 37)
MycoBank number: MB860203
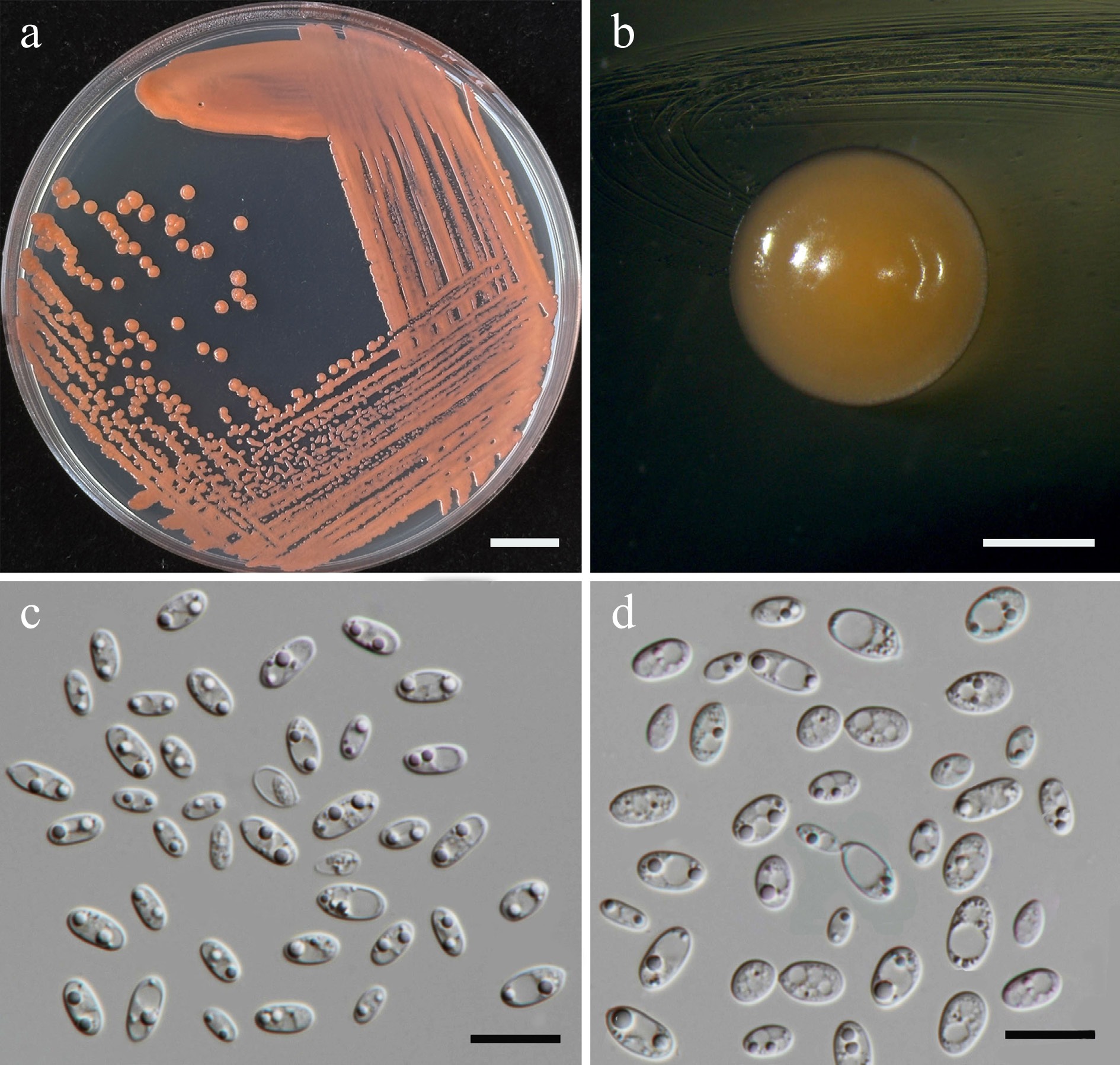
Figure 37.
Morphological characteristics of Sporobolomyces thailandensis (SDBR-CMU584, ex-type). (a) Culture, (b) single colony, (c) cells, and (d) budding cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The species name 'thailandensis' refers to Thailand, the country where the type strain was isolated.
Holotype – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in Oncidium flower (Oncidium sp.), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40091 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU584 = GMBCC2415 = TBRC21391. GenBank numbers PV834543 (D1/D2), PV834713 (ITS), PX622346 (SSU), PX582300 (rpb1), PX582325 (rpb2), PX582344 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.8–2.0 mm in diameter), reddish orange, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ellipsoidal and cylindrical (2.38–4.55 × 4.4–8.66 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, sorbose, xylose, sucrose, maltose, α-α-trehalose, raffinose, melizitose, glycerol (weak), ribitol, D-glucitol, mannitol, D-glucono-1,5-lactone, succinate, and citrate are assimilated, but D-galactose, N-acetyl glucosamine, ribose, D-arabinose, L-arabinose, L-rhamnose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose, inulin, soluble starch, erythritol, galactitol, myo-Inositol, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfate, potassium nitrate (weak), sodium nitrite (weak), ethylamine, and L-lysine are assimilated as sole nitrogen source, but cadaverine is not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is positive. Growth on 10, 15, 25, 30, and 35 °C but not at 37 and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in banana flower (Musa sapientum), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU695; Mueang District, Chang Phueak, in khae na flower (Dolichandrone serrulate), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU702. GenBank numbers SDBR-CMU695: PV834544 (D1/D2), PV834714 (ITS), PX622347 (SSU), PX582301 (rpb1), PX582326 (rpb2), PX582345 (tef1-α); SDBR-CMU702: PV834545 (D1/D2), PV834715 (ITS), PX622348 (SSU), PX582302 (rpb1).
Notes – Based on phylogenetic analyses (Fig. 35), Sporobolomyces thailandensis SDBR-CMU584, SDBR-CMU695, and SDBR-CMU702 found to be closely related to Sp. sucorum CBS 15628, Sp. shibatanus CBS 491, Sp. patagonicus CBS 9657 Sp. sucorum CBS 15628, and Sp. salmoneus CBS 488. These species are differed by 0.37%, 0.68%, 0.85%, and 1.23% nucleotide divergence (2, 4, 5, and 7 nt substitutions), respectively, in the D1/D2 domain. Meanwhile the ITS region revealed differences of 1.41%, 1.04%, 1.08%, and 2.48% (7, 6, 6, and 14 nt substitutions), respectively. Additionally, Sp. thailandensis is different from Sp. patagonicus and Sp. salmoneus by 12.92%–13.14% and 4.10%–7.38% nucleotide divergence in rpb2 and tef1-α, respectively. The phenotypic characteristics of Sp. thailandensis can be distinguished from Sp. patagonicus, Sp. salmoneus, Sp. shibatanus, and Sp. sucorum are provided in Table 11.
Table 11. Phenotypic characteristics differentiating Sporobolomyces thailandensis from closely related Sporobolomyces species.
Characteristics 1 2 3 4 5 Carbon assimilation D-Galactose – s v + w D-Ribose – + v +/s – Sucrose + + + + – Maltose + v + + – α-α-Trehalose + v + + – Methyl-α-D-glucoside – v + + nd Cellobiose – – + + – Salicin – – + + nd Melibiose – – + – – Raffinose + + + + – Melizitose + + + + – Soluble starch – + l + – Glycerol w s – + w Ribitol + – – v nd D-Glucitol + v + – nd D-Mannitol + s + + – D-Gluconate – + + + nd Citrate + – v + – Ethanol – – – + + Nitrogen assimilation Potassium nitrate w + v – + Sodium nitrite w – v – – Ethylamine HCl + + v nd w L-Lysine + + v nd – Growth characteristics Growth at 30 °C + + v + + Growth at 35 °C – – – – + Growth at 37 °C – – – nd w Growth in 0.1% Cycloheximide v – nd – + Strains 1: Sp. thailandensis sp. nov. 2: Sp. patagonicus[99], 3: Sp. salmoneus 4: Sp. shibatanus[88,100], and 5: Sp. sucorum[101]. Subphylum: Ustilaginomycotina Doweld
Class: Exobasidiomycetes Begerow, M. Stoll, & R. Bauer
Order: Exobasidiales Henn.
Family: Brachybasidiaceae Gäum.
Brachybasidiaceae represents a group of plant-parasitic basidiomycetes that decay dead organic matter, including wood and leaves. The vegetative morphology presents dimorphism, producing a saprobic haploid and a parasitic dikaryotic form. The cellular structure exhibits septate hyphae and specialized reproductive structures[102−104]. Currently, nine genera are listed in this family including Brachybasidium, Dicellomyces, Eidernor, Kordyana, Lelum, Marantokordyana, Meira, Proliferobasidium, and Yunzhangomyces[41]. In this study, Meira argovae (three strains), Mei. plantarum (one strain), Mei. chiangmaiensis sp. nov. (three strains), Mei. limtongiae sp. nov. (three strains), and Mei. pollinicola sp. nov. (three strains) are presented. (Fig. 38).
Figure 38.
Phylogram generated by maximum likelihood analysis of the combined D1/D2 domain of LSU, ITS, SSU, rpb1, rpb2, and tef1-α sequence data representing Meira and Laurobasidium in Exobasidiales. The tree is rooted to Rhamphospora nymphaeae (CBS 172.38). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Sixty-two strains are included in the combined sequence analysis, which comprise 5,633 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Meira chiangmaiensis Kodchasee, Senwanna, J. Kumla. & N. Suwannar., sp. nov. (Fig. 39)
MycoBank number: MB860204
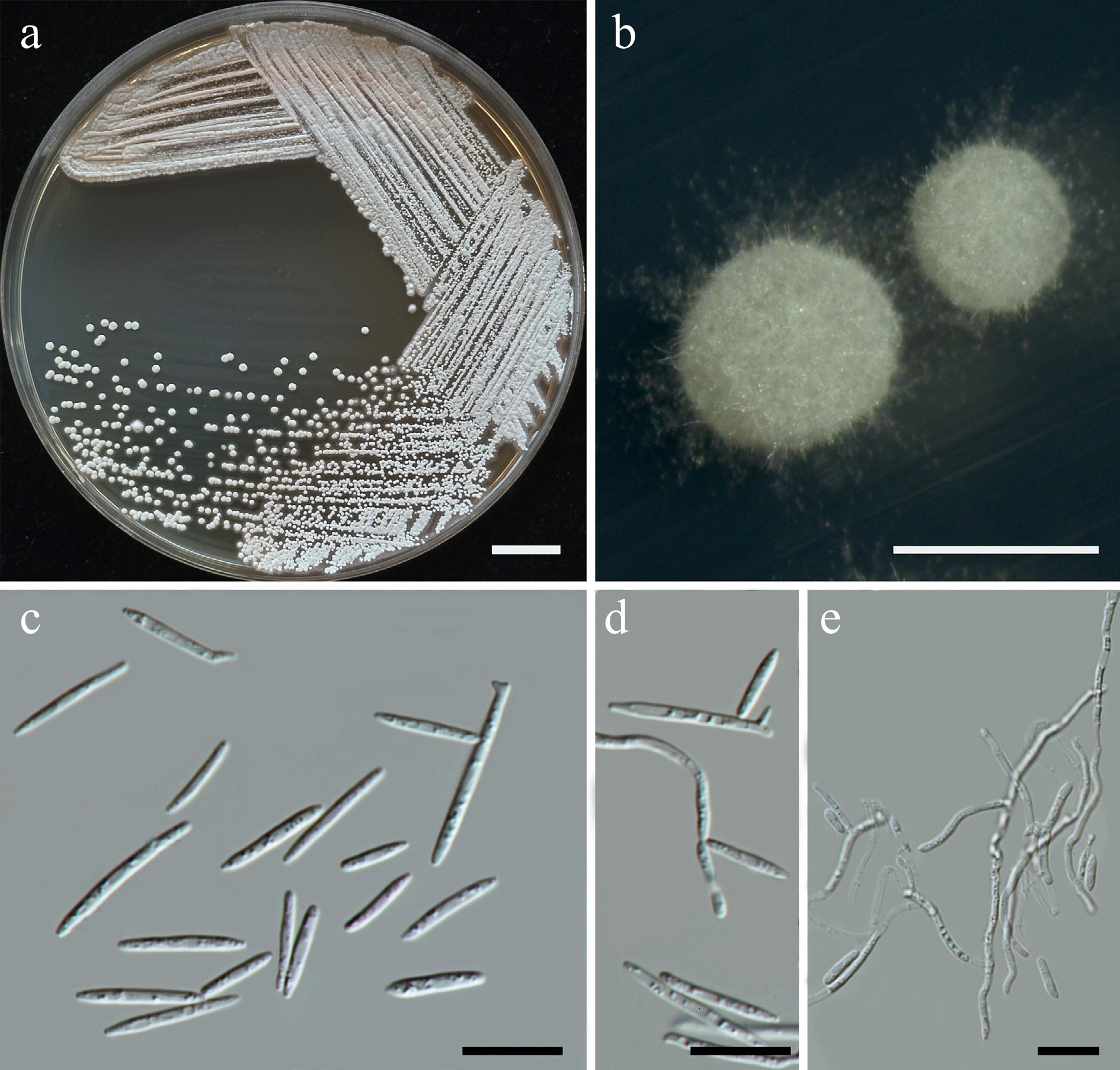
Figure 39.
Morphological characteristics of Meira chiangmaiensis (SDBR-CMU624, ex-type). Culture (a), single colony (b) and cells (c, d) on YMA after five days at 25 °C. Pseudohyphae on PDA after 2 weeks at 25 °C (d, e). Scale bars a = 10 mm, b = 1 mm, c and d = 10 μm.
Etymology – 'chiangmaiensis' referring to Chiang Mai Province, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in jasmine flower (Jasminum sambac), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40101 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU624 = GMBCC2416 = TBRC21402. GenBank numbers PV834549 (D1/D2), PV834719 (ITS), PX622349 (SSU), PV941868 (rpb1), PV947460 (rpb2), PV947488 (tef1-α).
Description – Colonies on YMA after 3 d at 25 °C is circular form (1–2 mm in diameter) pale yellow smooth, after 5 d rigid, wrinkled, warty, velvety, and presence of thin aerial mycelia. After growth on YMA at 25 °C for 3 d, the cells are ellipsoidal to fusiform (0.95–1.84 × 5.42–18.1 μm, n = 50), occur polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, N-acetyl glucosamine (or weak), ribose, xylose, L-arabinose, D-arabinose (or weak), sucrose, maltose, α-α-trehalose, cellobiose, salicin (or weak), melibiose, raffinose, melizitose, erythritol, ribitol (or weak), D-glucitol, mannitol, myo-inositol (or weak), D-glucono-1,5-lactone, succinate, citrate (or weak), and xylitol (or weak) are assimilated, but sorbose, L-rhamnose, methyl-α-D-glucoside, lactose, inulin, soluble starch, glycerol (or weak), galactitol, methanol, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, and ethanol are not assimilated. Ammonium sulfates, potassium nitrate (or weak), and ethylamine hydrochloride are assimilated as sole nitrogen sources, L-lysine are variable, but sodium nitrite and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (or weak). Not growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Suthep, in bougainvillea (Bougainvillea hybrid), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU638; White cheesewood (Alstonia scholaris) flower, September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU731. GenBank numbers SDBR-CMU638: PV834550 (D1/D2), PV834720 (ITS), PX622350 (SSU), PV941869 (rpb1), PV947489 (tef1-α); SDBR-CMU731: PV834551 (D1/D2), PV834721 (ITS), PX622351 (SSU).
Notes – Meira chiangmaiensis SDBR-CMU624, SDBR-CMU638 and SDBR-CMU731 were related to Mei. pollinicola by 0.98% nucleotide divergence (6 nt substitutions) in the D1/D2 domain and 3.08% (24 nt substitutions, including 19 gaps) in the ITS region. In addition, Mei. chiangmaiensis and Mei. pollinicola differed from Mei. argovae CBS 110053 by 0.5% and 0.67% nucleotide divergence (3 nt substitutions and 4 nt substitutions, including 1 gap, respectively,) in the D1/D2 regions and 2.3% and 2.79% (14 nt substitutions and 17 nt substitutions including 20 gaps, respectively) in the ITS region. Moreover, Mei. chiangmaiensis is different from Mei. argovae by 3.67% nucleotide divergence (27 substitutions and 1 gap) in rpb1, 4.56% nucleotide divergence (49 substitutions and 1 gap) in rpb2 and 3.52% nucleotide divergence (31 substitutions and 1 gap) in tef1-α (Fig. 38). Based on physiological tests, Mei. chiangmaiensis can distinguish it from Mei. argovae[105] and Mei. pollinicola, which are shown in Table 12.
Table 12. Phenotypic characteristics differentiating Meira chiangmaiensis sp. nov., Mei. pollinicola sp. nov., and Mei. limtongiae sp. nov. from closely related Meira species.
Characteristics 1 2 3 4 5 6 7 8 9 10 Carbon assimilation L-Sorbose – – – – – w/l – – – – D-Arabinose w/– – w + + – +/w + – w L-Rhamnose – – – – – – –/w nd – – Methyl-α-D-glucoside – + – – – – w nd – – Cellobiose + + + + w l + + + + Salicin w/+ + + l l w/l w l l/w w Melibiose + – – +/– + l + + + + Melizitose + + + – – + + + + + Inulin – – – w l l –/w – – + Soluble starch – – – +/l l w/l –/w + – – Glycerol – w – w/l l – –/w – – w Ribitol w + w +/l +/l – +/w – – w D-Glucitol + + + + + + –/w + + w Galactitol – – – +/l l – + – v – myo-Inositol w/– – – – – – –/w – – – D-Glucono-1,5-lactone + + – + l nd + – nd nd D-Gluconate – w – – – nd w – nd nd DL-Lactate – + w w/l l l v w – – Succinate + + w + + l +/w + + w/l Citrate w/– + – + l w/l w – – – Ethanol – – – l l + w + – w Xylitol w/– + – +/d +/d nd +/w + nd nd Nitrogen assimilation Potassium nitrate w/– + + + – l + + + + Sodium nitrite – + + + – l + + + + L-Lysine v w w – + l + nd l/w + Cadaverine – – – + + + + nd – + Growth characteristics Growth at 25 °C + + + + + nd + + + + Growth at 30 °C + + + + + + + nd nd nd Growth at 35 °C – – – – + nd nd nd nd nd Growth at 37 °C – – – – + nd – – nd nd Growth in 0.01% cycloheximide – – – – + nd – nd nd nd Growth in 0.1% cycloheximide – – – – + nd – nd nd nd Growth on 50% glucose + + + – – + + nd – – Strians 1: Mei. chiangmaiensis sp. nov., 2: Mei. limtongiae sp. nov, 3: Mei. pollinicola sp. nov., 4: Mei. argovae, 5: Mei. geulakonigii[105], 6: Mei. marina[81], 7: Mei. miltonrushii[103], 8: Mei. nashicola[106], 9: Mei. plantarum, and 10: Mei. pileae[107]. Meira limtongiae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 40)
MycoBank number: MB860178
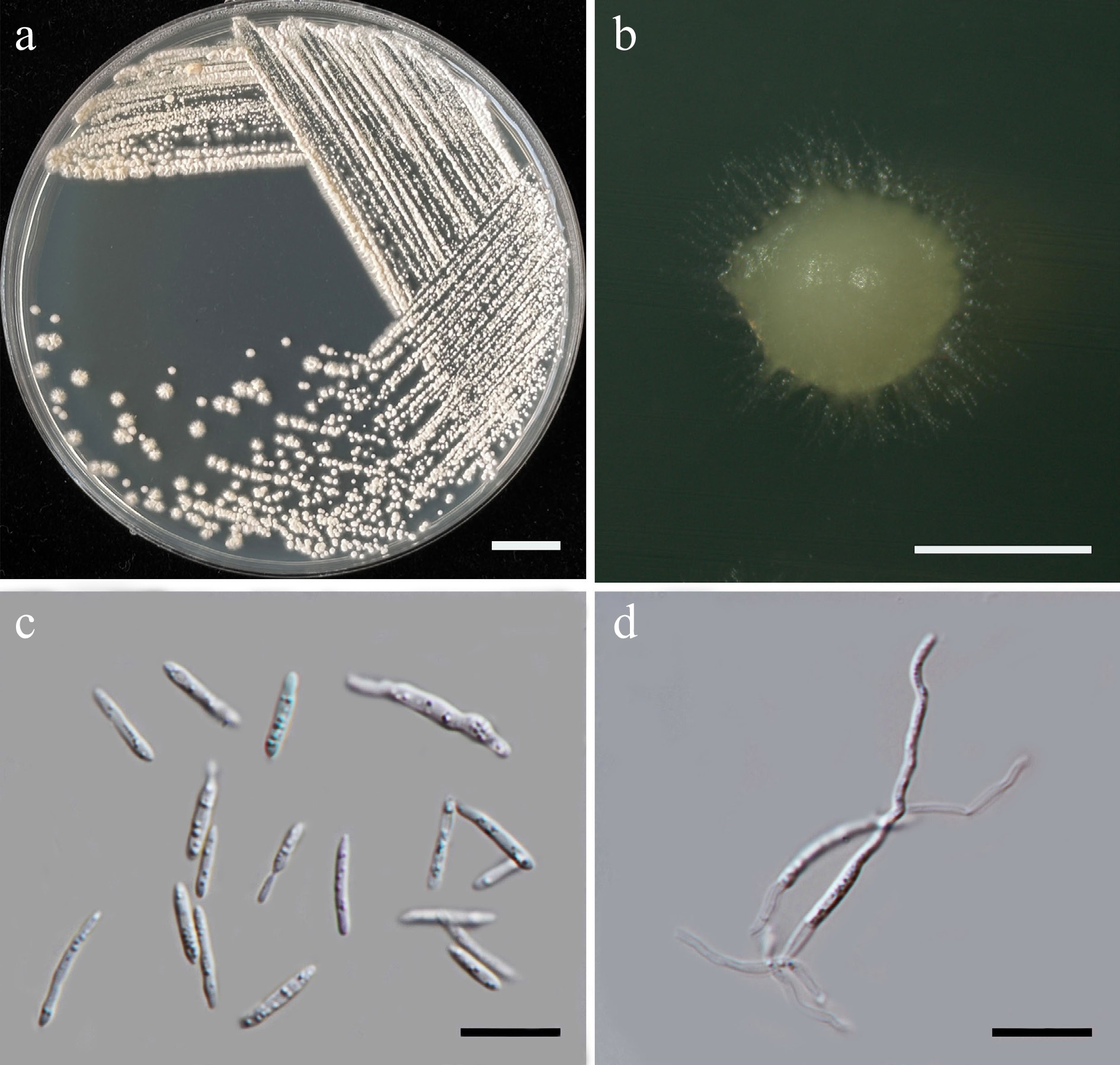
Figure 40.
Morphological characteristics of Meira limtongiae (SDBR-CMU696, ex-type). (a) Culture, (b) single colony, and (c) cells on YMA after 5 d at 25 °C. (d) Pseudohyphae on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'limtongiae' named in honor of mycologist Savitree Limtong, for her contributions to yeast systematics.
Holotype: THAILAND, Phayao Province, Mueang District, in blue ginger flower (Dichorisandra thyrsiflora), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40110 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU696 = GMBCC2417 = TBRC21413. GenBank numbers PV834554 (D1/D2), PV834724 (ITS), PX622354 (SSU), PX570013 (rpb2).
Description – Colonies on YMA after 3 d at 25 °C are circular form (1–2 mm in diameter) pale yellow smooth, after 5 d rigid, wrinkled, warty, velvety, and presence of thin aerial mycelia. After growth on YMA at 25 °C for 3 d, the cells are ellipsoidal to fusiform (1.08–2.24 × 4.01–16.92 μm, n = 50), occur polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, ribose, xylose, L-arabinose, D-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, raffinose, melizitose, glycerol (weak), erythritol, ribitol, D-glucitol, mannitol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, DL-lactate, succinate, citrate, and xylitol are assimilated, but sorbose, N-acetyl glucosamine L-rhamnose, melibiose, lactose, inulin, soluble starch, galactitol, myo-inositol, methanol, D-galacturonic acid, and ethanol are not assimilated. Ammonium sulfates, potassium nitrate, sodium nitrite, ethylamine hydrochloride (weak), and L-lysine (weak) are assimilated as sole nitrogen source, but cadaverine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose (weak). Not growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in adenium flower (Adenium obesum), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU572; Phrao District, Nam Phrae, in dancing ladies ginger (Globba winitii) flower, July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU600. GenBank numbers SDBR-CMU572: PV834552 (D1/D2), PV834723 (ITS), PX622352 (SSU), PX570011 (rpb2); SDBR-CMU600: PV834553 (D1/D2), PV834715 (ITS), PX622353 (SSU), PX570012 (rpb2).
Notes – Meira limtongiae formed a distinct lineage and was sister to Mei. argrovae, Mei chiangmaiensis, Mei. pollinicola, Mei. miltonrushii and, Mei. geulakonigae (Fig. 38). Meira limtongiae differed from these species by 0.83%–1.85% nucleotide divergence (5–10 substitutions, including gaps) in the D1/D2 domain, 4.64%–6.46% nucleotide divergence (28–39 substitutions, including gaps) in the ITS region, and 13.84%–14.69% nucleotide divergence (148–157 substitutions, including gaps) in rpb2. Additionally, distinguishing physiological characteristics between Mei. limtongiae and other Maira species are provided in Table 12.
Meira pollinicola Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 41)
MycoBank number: MB860206
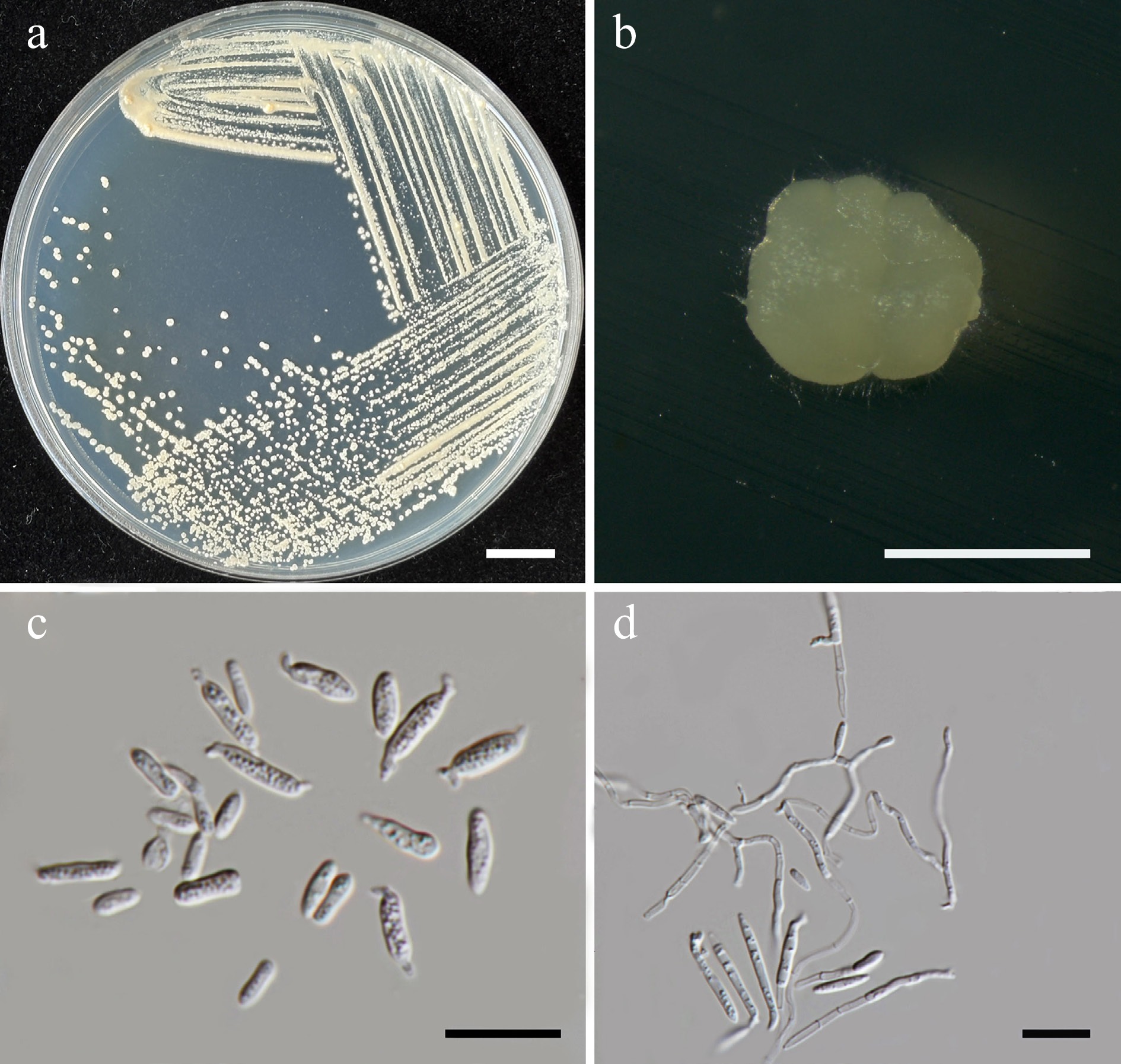
Figure 41.
Morphological characteristics of Meira pollinicola (SDBR-CMU680, ex-type). (a) Culture, (b) single colony, and (c) cells on YMA after 5 d at 25 °C. (d) Pseudohyphae on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The specific epithet 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Phayao Province, Mueang District, in chaya spinach flower (Cnidoscolus aconitifolius), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype CMUB40108 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU680 = GMBCC2418 = TBRC21410. GenBank numbers PV834557 (D1/D2), PV834727 (ITS), PX622356 (SSU), PX582304 (rpb1), PX582328 (rpb2).
Description – Colonis on YMA after 3 d at 25 °C is circular form (1–2 mm in diameter) yellow/orange smooth, after 5 d rigid, wrinkled, warty, velvety, and presence of thin aerial mycelia. After growth on YMA at 25 °C for 5 d, the cells are ellipsoidal to fusiform (1.16–2.84 × 3.81–12.00 μm, n = 50), occur polar budding. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, ribose, xylose, L-arabinose, D-arabinose (weak), sucrose, maltose, α-α-trehalose, cellobiose, raffinose, melizitose, erythritol, ribitol (or weak), D-glucitol, mannitol, DL-lactate (or weak), and succinate, (or weak) are assimilated, but sorbose, N-acetyl glucosamine L-rhamnose, methyl-α-D-glucoside, salicin, melibiose, lactose, inulin, soluble starch, glycerol, galactitol, myo-inositol, D-glucono-1,5-lactone, citrate, methanol, D-gluconate, D-glucuronate, D-galacturonic acid, ethanol, and xylitol are not assimilated. Ammonium sulfates, potassium nitrate, sodium nitrite, ethylamine hydrochloride, and L-lysine (or weak) are assimilated as sole nitrogen source, but cadaverine are not assimilated. Growth occurs on media containing 50% glucose and 60% glucose (weak). Not growth occurs on media containing 10% NaCl/5% glucose 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth on 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in frangipani flower (Phyllanthus pulcher), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU550; Mueang District, Chang Phueak, in purslane flower (Portulaca grandiflora), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU700. GenBank numbers SDBR-CMU550: PV834556 (D1/D2), PV834714 (ITS), PX622355 (SSU), PX582303 (rpb1), PX582327 (rpb2); SDBR-CMU700: PV834558 (D1/D2), PV834728 (ITS), PX622357 (SSU), PX582305 (rpb1), PX582329 (rpb2).
Notes – Meira pollinicola SDBR-CMU680, SDBR-CMU550 and SDBR-CMU700 were related to Mei. argovae CBS110053 and Mei. chiangmaiensis SDBR-CMU624 (Fig. 38). Meira pollinicola differed from the to Mei. argovae by 0.67% nucleotide divergence (4 nt substitutions and 1 gap) in D1/D2 regions and 2.79% (17 nt substitutions and 20 gaps) in ITS region. Likewise, Mei. pollinicola differed from Mei. chiangmaiensis with 0.98% nucleotide divergence (6 nt substitutions) in D1/D2 regions and 3.08% (19 nt substitutions and 24 gaps) in ITS region. Moreover, Mei. pollinicola is different from Mei. argovae and Mei. chiangmaiensis by 2.47%–4.26% and 3.09% to 4.22% nucleotide divergence in rpb1 and rpb2, respectively. Based on physiological test, Mei. pollinicola can be distinguished from Mei. argovae[105] and Mei. chiangmaiensis as shown in Table 12.
Family: Laurobasidiaceae Pinruan, Sommai, Suetrong, Somrith, & E.B.G. Jones
Laurobasidiaceae morphologically possesses basidial sterigmata, thin-walled basidiospores with an abaxial hilum, and sporulates on the surface of the host tissues that cause to causing aerial-root-like galls on the host plant. Sexual morph present elongated basidium holobasidiate, clavate to cylindrical, with sterigmata; basidiospores ellipsoidal, oblong, or cylindrical, septate, thin-walled, with an abaxial hilum, while asexual morph present thin- and smooth-walled, fusiform, clavate to narrow ellipsoid of conidia hyaline[108]. Currently, one genus is listed in this family, including Laurobasidium[41]. Laurobasidium hachijoense (two strains) was isolated and identified in this study (Fig. 38, Supplementary File 1).
Order: Microstromatales R. Bauer & Oberw.
Families incertae sedis
Currently, five genera are listed under Microstromatales genera incertae sedis, including Baueromyces, Jaminaea, Parajaminaea, Pseudomicrostroma, and Sympodiomycopsis [41]. In this study, seven yeast species were isolated and presented, including Jaminaea lantanae (one strain), Parajaminaea hydei sp. nov. (two strains), Sympodiomycopsis europaea (two strains), Sym. paphiopedili (two strains), Sym. hydei sp. nov. (four strains), Sym. limtongiae sp. nov. (two strains) and Sym. saisamorniae sp. nov. (two strains).
Parajaminaea hydei Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 42)
MycoBank number: MB860207
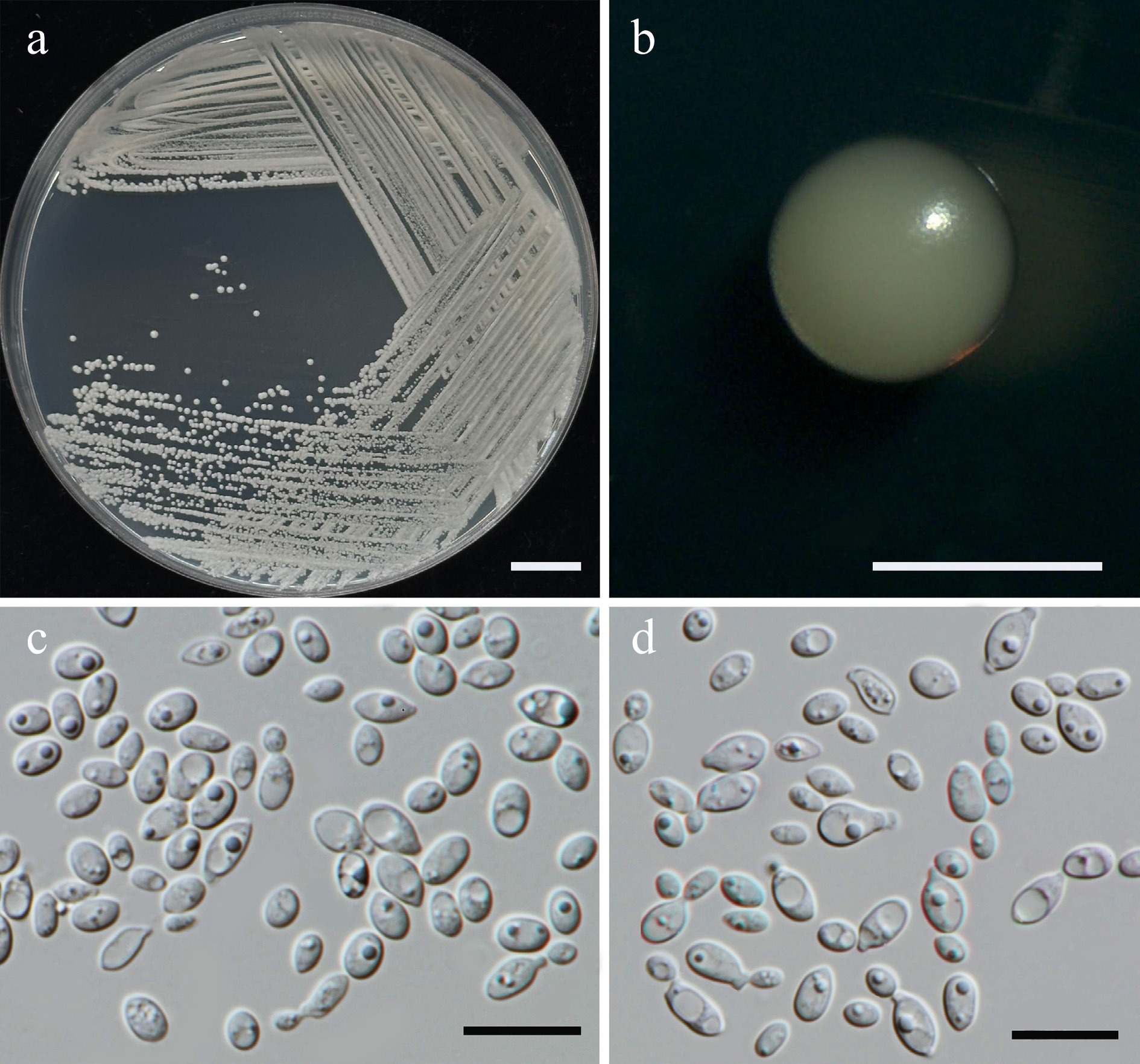
Figure 42.
Morphological characteristics of Parajaminaea hydei (SDBR-CMU601, ex-type). (a) Culture, (b) single colony, and (c), (d) cells on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'hydei' referring to the name of mycologist Kevin D. Hyde in honor of his 70th birthday.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in ground cherry flower (Physalis minima), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40096 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU601 = GMBCC2419 = TBRC21395. GenBank numbers PV834562 (D1/D2), PV834732 (ITS), PX622358 (SSU), PV947453 (rpb2), PV947477 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.0 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ellipsoidal and ovoid (2.25–3.63 × 3.06–6.8 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, neither pseudohyphae nor true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, ribose (or latent or slow), L-arabinose, sucrose, maltose (or slow), methyl-α-D-glucoside (or latent or slow), raffinose, melizitose, glycerol (or slow), erythritol (or weak), mannitol (or latent), and ethanol are assimilated, but D-galactose, sorbose, xylose, D-arabinose, L-rhamnose, α-α-trehalose, salicin, melibiose, lactose, inulin, soluble starch, ribitol, myo-inositol, D-glucono-1,5-lactone, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, methanol, and xylitol are not assimilated. Assimilation is variable for N-acetyl glucosamine, cellobiose, D-glucitol, galactitol, and D-gluconate. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Phayao Province, Mueang District, in bridal bouquet flower (Plumeria pudica), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU685. GenBank numbers SDBR-CMU685: PV834563 (D1/D2), PV834733 (ITS), PX622359 (SSU), PV947454 (rpb2), PV947478 (tef1-α).
Notes – Parajaminaea hydei SDBR-CMU601 and SDBR-CMU685 were identified as members of Parajaminaea and related to Par. phylloscopi CBS14087 and Par. albiziae CMW 36935 (Fig. 43). The D1/D2 sequences differed by 3.5% nucleotide divergence (18 nt substitutions including 2 gaps), and 8.43% nucleotide divergence (50 nt substitutions including 16 gaps) in the ITS region with Par. phylloscopi, while differed from Par. albiziae by 3.22% nucleotide divergence (19 nt substitutions including 1 gap) and 8.13% (53 nt substitutions including 17 gaps) in D1/D2 and ITS region, respectively, which indicated that they represent a novel species in Parajaminaea. Additionally, Par. hydei can be distinguished from Par. phylloscopi by its inability to assimilate α-α-trehalose, inulin, succinate and its lack of growth at 35 and 42 °C[109].
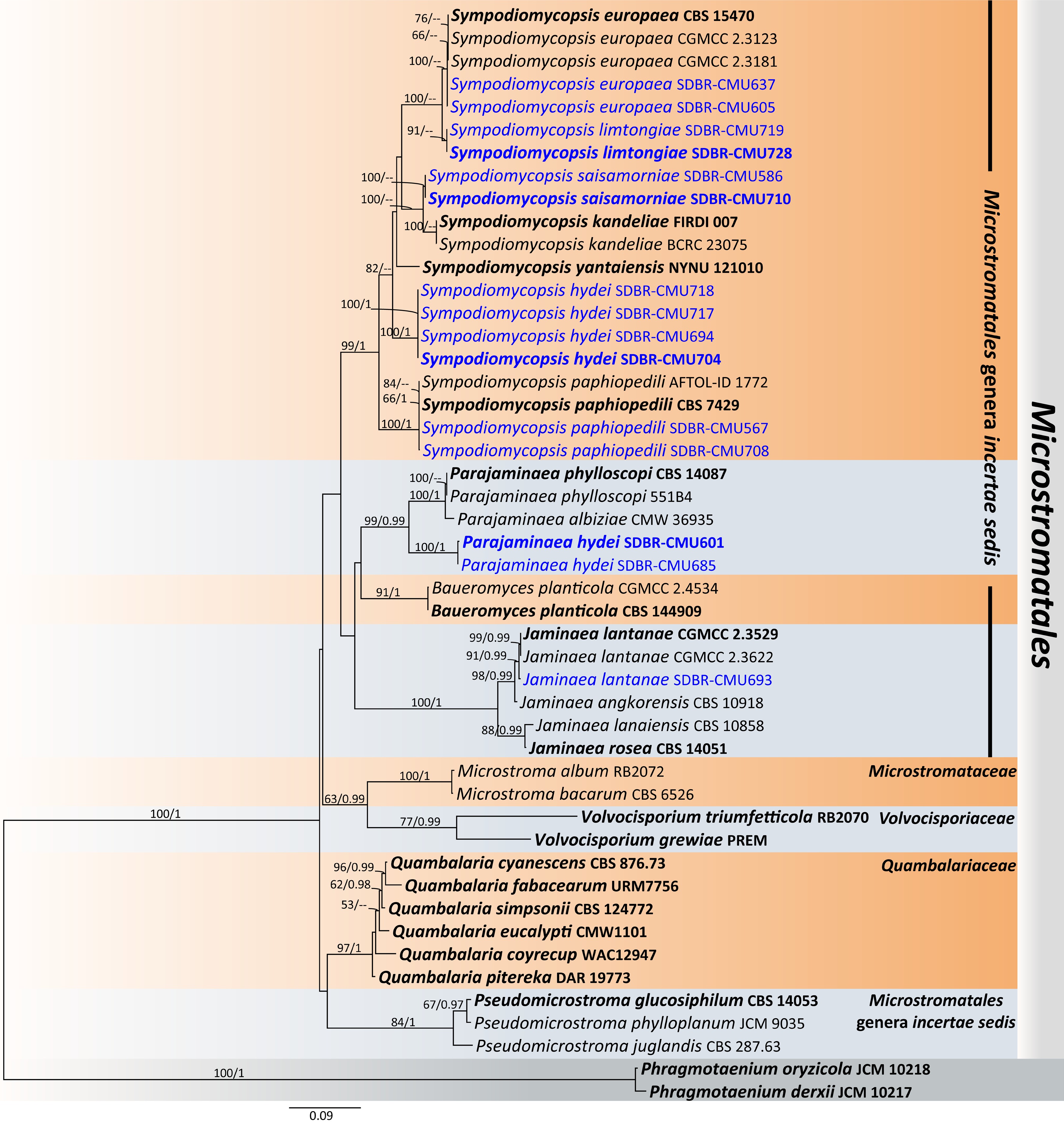
Figure 43.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Microstromatales. The tree is rooted to Phragmotaenium derxii (JCM 10217) and P. oryzicola (JCM 10218). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fourty-eight strains are included in the combined sequence analysis, which comprise 1,326 characters with gaps. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.95 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.95 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Sympodiomycopsis hydei Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 44)
MycoBank number: MB860208
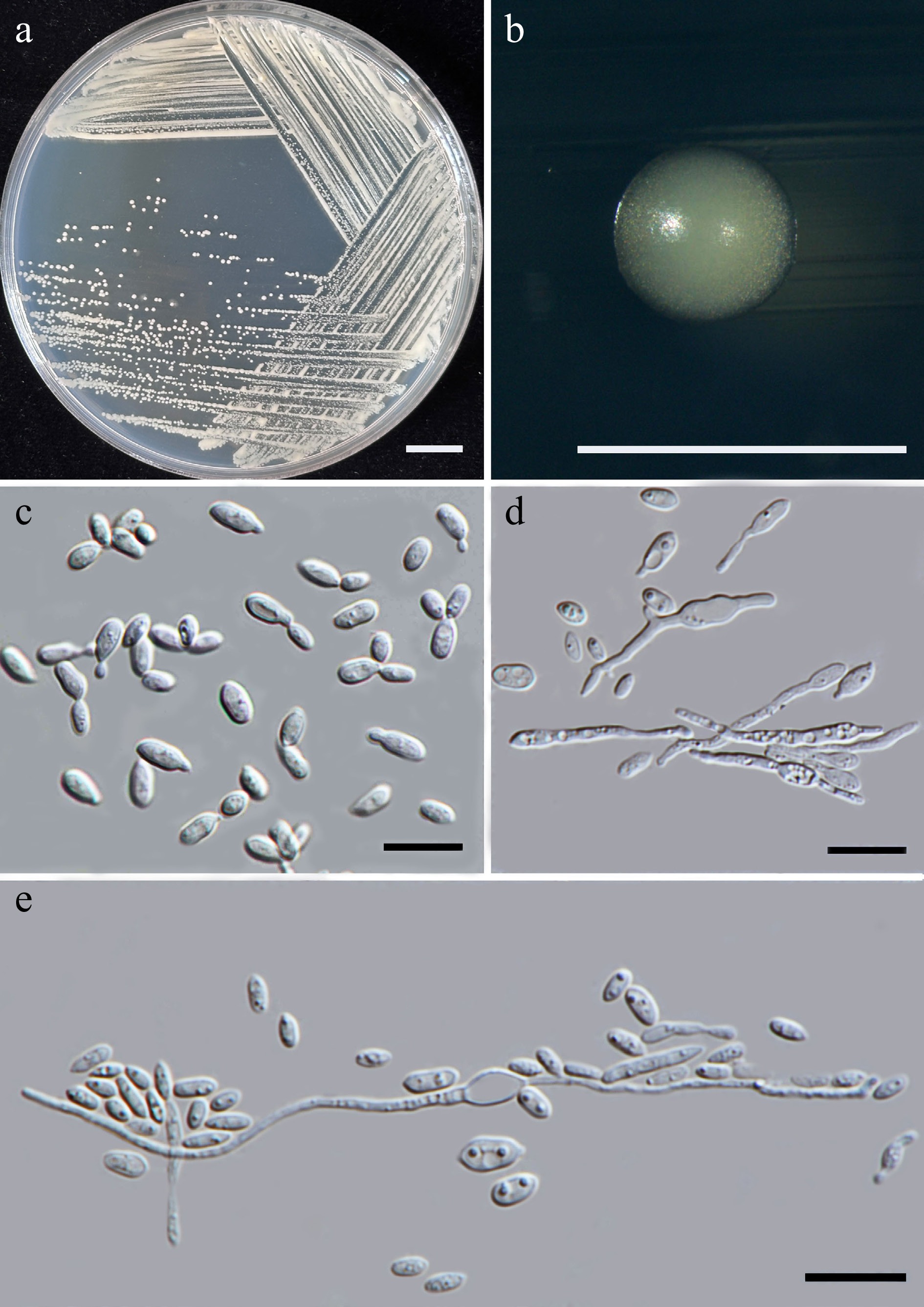
Figure 44.
Morphological characteristics of Sympodiomycopsis hydei (SDBR-CMU704, ex-type). (a) Culture, (b) single colony, and (c) cells on YMA at 25 °C for 5 d. (d), (e) Pseudohyphae on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c)–(e) = 10 μm.
Etymology – 'hydei' referring to the name of Kevin D. Hyde in honor of his 70th birthday.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak in cape leadwort flower (Plumbago auriculata), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40112 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU704 = GMBCC2420 = TBRC21415. GenBank numbers PV834567 (D1/D2), PV834737 (ITS), PV941870 (rpb1), PV947456 (rpb2).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.5–0.8 mm in diameter), yellowish white, smooth surface, glistening appearance, entire margin, and convex elevation. The cells are ovoid to obclavate (2.04–3.52 × 3.4–6.68 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, ribose (or weak), L-arabinose, sucrose, maltose (or weak), methyl-α-D-glucoside, melibiose (or slow), raffinose, melizitose, glycerol, erythritol (or slow), ribitol, D-glucitol, mannitol, and xylitol (or weak) are assimilated, but inulin, soluble starch, lactose, cellobiose, salicin, N-acetyl glucosamine, xylose, D-arabinose, L-rhamnose, α-α-trehalose, galactitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, and L-lysine are assimilated as sole nitrogen source, but ethylamine and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose (weak). No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in ylang-ylang tree flower (Cananga odorata), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU694; Mueang District, Chang Phueak, in marguerite daisy flower (Argyranthemum frutescens), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU717; San Kamphaeng District, Ton Pao, in flosreginae flower (Lagerstroemia speciosa), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU718. GenBank numbers SDBR-CMU694: PV834566 (D1/D2), PV834736 (ITS); SDBR-CMU717: PV834568 (D1/D2), PV834738 (ITS); SDBR-CMU718: PV834569 (D1/D2), PV834739 (ITS).
Notes – Based on phylogenetic analysis (Fig. 43), Sym. hydei (SDBR-CMU704, SDBR-CMU694, SDBR-CMU717, and SDBR-CMU718) formed a separate lineage and clustered as sister to Sym. europaea, Sym. kandeliae, Sym. limtongiae, Sym. paphiopedili, and Sym. yantaiensis, However, Sym. hydei differed from those species by 0.67%–2.10% nucleotide divergence (4–13 nt substitutions including gaps) in the D1/D2 domain and 6.55%–7.91% mismatches (39–51 nt substitutions including 12–20 gaps) in the ITS regions. Furthermore, Sym. hydei can be distinguished from those species in its physiological characteristics (Table 13).
Table 13. Phenotypic characteristics differentiating Sym. hydei, Sym. limtongiae, and Sym. saisamorniae from closely related Sympodiomycopsis species.
Characteristics 1 2 3 4 5 6 7 Carbon assimilation D-Glucose + + + + + + w D-Galactose w + – + +/s + w L-Sorbose + s/+ + + + + – D-Arabinose – – – – +/s + – α-α-Trehalose + v + w + + + Cellobiose – – – – +/s + – Salicin – – – – w w – Melibiose w s – + v + – Lactose – v – + +/s + – Inulin – – – – w/– – – Soluble starch – – – + w/– – – Erythritol w w/+ w + + + + Ribitol - v – – v + – D-Glucitol w v – +/w + + w D-Mannitol + + w + + + + Galactitol – – – – w – – myo-Inositol – – – w w/– + – D-Gluconate v – – nd s/w nd + Succinate + v – nd w + + Citrate – – – nd w + + Ethanol – – – + s + + Xylitol – w/+ – nd v nd – Nitrogen assimilation Potassium nitrate + + + v w + + Sodium nitrite + + + – v + – Ethylamine HCl – – w – w + – L-Lysine + + w + w – – Cadaverine – – – + nd – – Growth characteristics Growth at 25 °C + + + + + + + Growth at 30 °C + + + – + + + Growth in 0.01% Cycloheximide – – – nd + nd + Growth on 50% Glucose + + + – nd + + Growth on 60% Glucose + + + nd nd nd – Growth on 10% NaCl + 5% glucose w + – nd nd nd + Strian 1: Sym. hydei sp. nov, 2: Sym. limtongiae sp. nov, 3: Sym. saisamorniae sp. nov, 4: Sym. europaea[92], 5: Sym. kandeliae[110], 6: Sym. paphiopedili[111], and 7: Sym. yantaiensis[112]. Sympodiomycopsis limtongiae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 45)
MycoBank number: MB860209
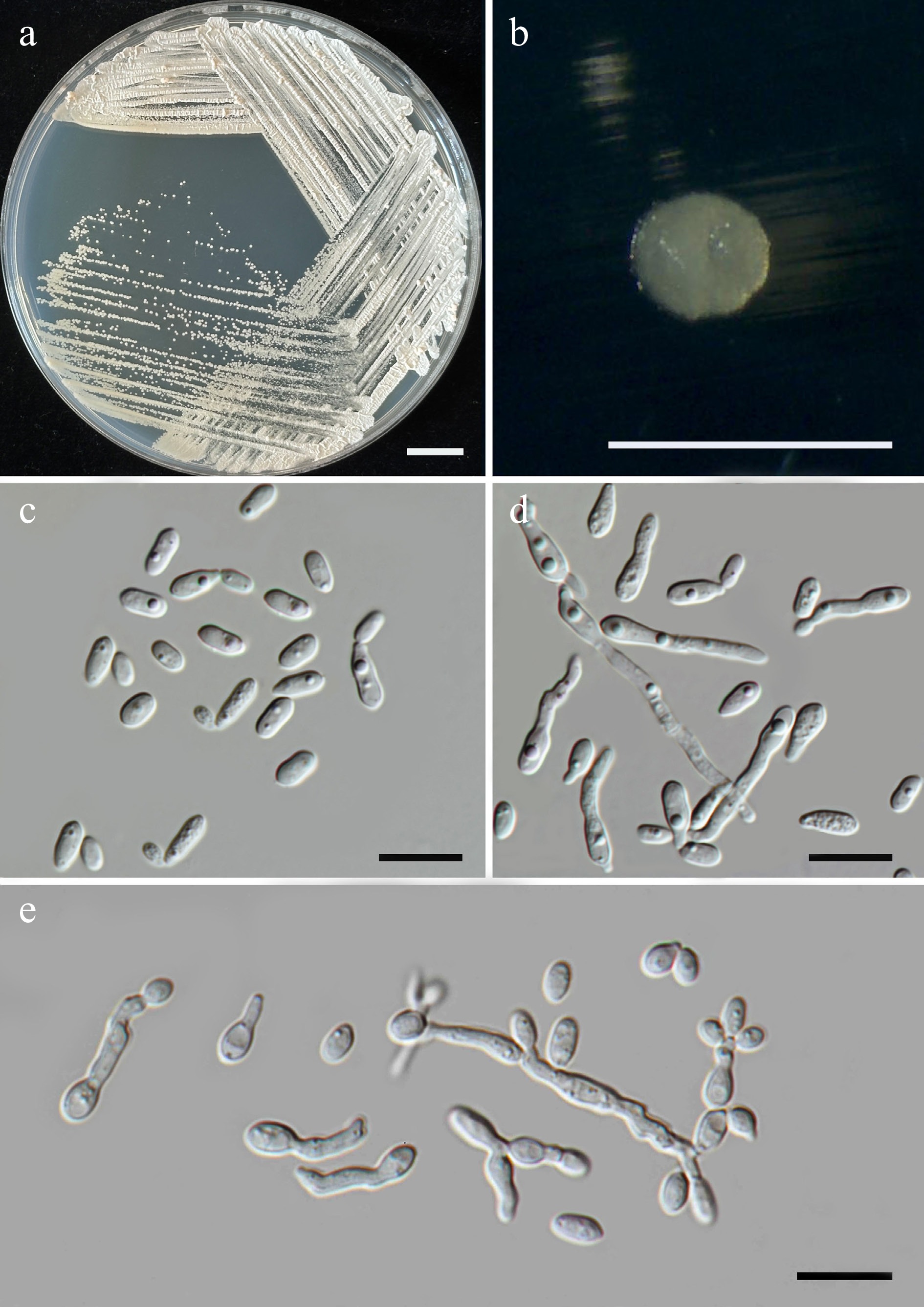
Figure 45.
Morphological characteristics of Sympodiomycopsis limtongiae (SDBR-CMU728, ex-type). (a) Culture, (b) single colony, and (c) cells on YMA at 25 °C for 5 d. (d), (e) Pseudohyphae on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, and (c)–(e) = 10 μm.
Etymology – 'limtongiae' named in honor of mycologist Savitree Limtong, for her contributions to yeast systematics.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep, in white cheese wood flower (Alstonia scholaris), July 2024, P. Kodchasee, C. Senwanna, J. Kumla and N. Suwannarach, holotype, CMUB40115 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU728 = GMBCC2421 = TBRC21418. GenBank numbers PV834570 (D1/D2), PV834740 (ITS), PV941872 (rpb1), PV947455 (rpb2), PV947476 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.7–1.2 mm in diameter), white glistening appearance, and eroded margins. The cells are ovoid to obclavate (2.05–3.42 × 3.69–6.43 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose (slow), ribose, xylose, L-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, melibiose (slow), lactose (slow), raffinose, melizitose, glycerol, erythritol, D-mannitol, D-glucono-1,5-lactone (weak), DL-lactate, succinate, and xylitol are assimilated, but N-acetyl glucosamine, D-arabinose, L-rhamnose, cellobiose, salicin, inulin, soluble starch, ribitol, D-glucitol, galactitol, myo-inositol, D-gluconate, D-glucuronate, D-galacturonic acid, citrate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, and L-lysine are assimilated as sole nitrogen source, but ethylamine and cadaverine are not assimilated. Growth occurs on media containing 50% glucose, 60% glucose, and 10% NaCl/5% glucose. No growth occurs on media containing 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth on 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, San Kamphaeng District, Ton Pao, in flosreginae flower (Lagerstroemia speciosa), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU719. GenBank numbers PV834571 (D1/D2), PV834741 (ITS).
Notes – Sympodiomycopsis limtongiae SDBR-CMU728 and SDBR-CMU719 were identified as members of Sympodiomycopsis and related to Sym. europaea CBS 15470, CGMCC 2.3123, and CGMCC 2.3181 (Fig. 43). A comparison of the D1/D2 and ITS sequences revealed that Sympodiomycopsis limtongiae differed from the latter by 0.48% nucleotide divergence (2 nt substitutions) and 1.96% (13 nt substitutions and 1 gap), respectively. In addition, Sym. limtongiae is different from Sym. europaea by 7.74%, 6.11%, and 2.08% nucleotide divergence (58, 66, and 18 nt substitutions) in rpb1, rpb2, and tef1-α, respectively. Likewise, distinguishing characteristics between Sym. limtongiae and Sym. europaea[92] are shown in Table 12.
Sympodiomycopsis saisamorniae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 46)
MycoBank number: MB860210
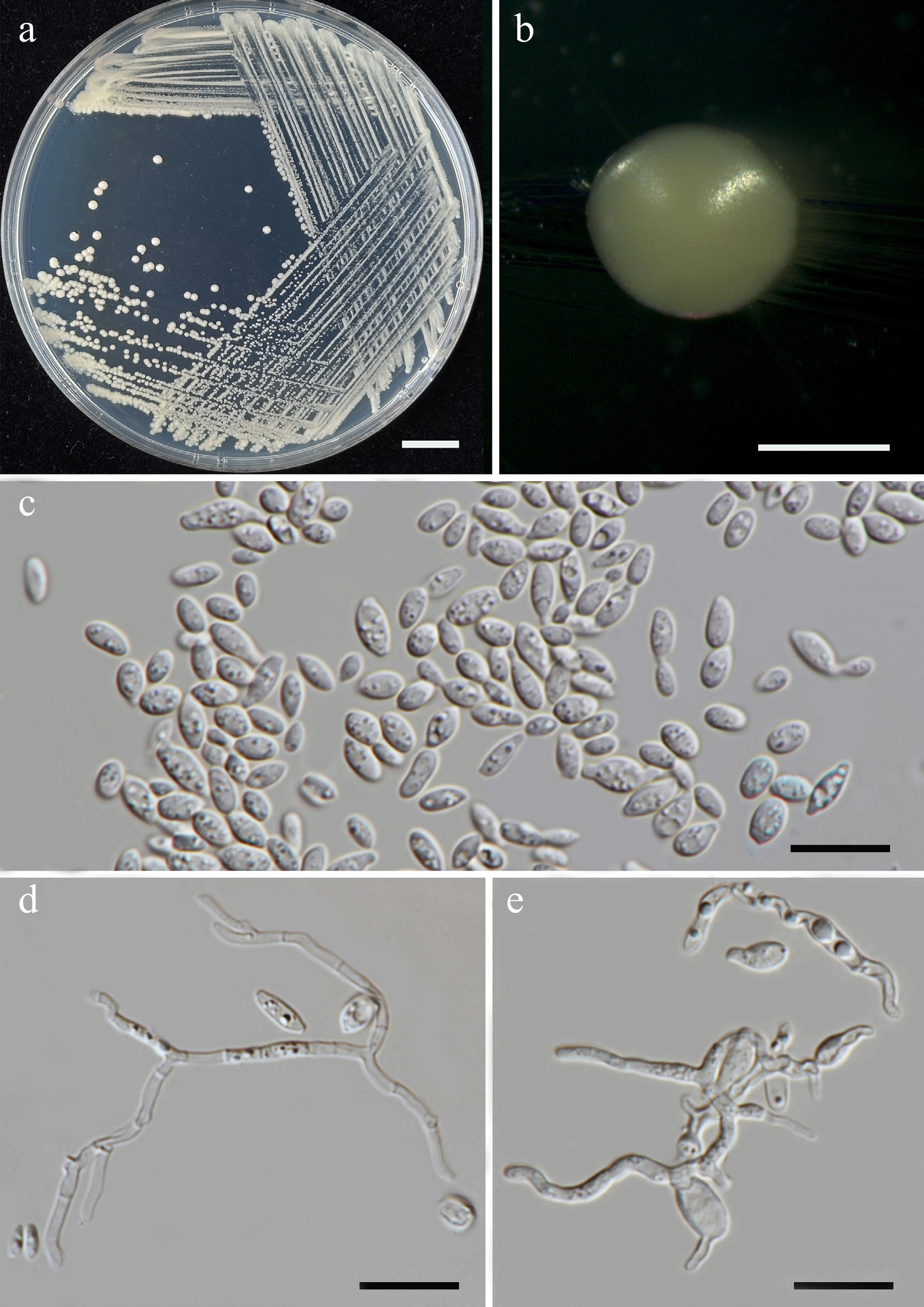
Figure 46.
Morphological characteristics of Sympodiomycopsis saisamorniae (SDBR-CMU710, ex-type). (a) Culture, (b) single colony, (c) cells and budding cells on YMA at 25 °C for 5 d. (d), (e) True hyphae and pseudohyphae on PDA after 2 weeks at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c)–(e) = 10 μm.
Etymology – 'saisamorniae' referring to the name of mycologist Saisamorm Lumyong, in honor of her 75th birthday.
Holotype – THAILAND, Chiang Mai Province, San Kamphaeng District, Ton Pao, in Indian mulberry flower (Morinda citrifolia), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40113 (preserved in metabolically inactive state), ex-type living SDBR-CMU710 = GMBCC2422 = TBRC21416. GenBank numbers PV834574 (D1/D2), PV834744 (ITS), PX622360 (SSU), PX582306 (rpb1).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.2 mm in diameter), pale orange, smooth surface, glistening appearance, circular shape, entire margin, and convex elevation. The cells are ovoid to obclavate (2.01–3.78 × 3.61–7.22 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, sorbose, ribose (weak), xylose (weak), L-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, raffinose, melizitose, glycerol, erythritol (weak), and D-mannitol (weak) are assimilated, but galactose, N-acetyl glucosamine, D-arabinose, L-rhamnose, cellobiose, salicin, melibiose inulin, lactose, soluble starch, ribitol, D-glucitol, galactitol, myo-inositol, D-glucono-1,5-lactone (weak), D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, methanol, ethanol, and xylitol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine (weak), and L-lysine (weak) are assimilated as sole nitrogen source but and cadaverine are not assimilated. Growth occurs on media containing 50% glucose 60% glucose. No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Urease reaction is positive. Acid formation is negative. Growth was observed at 10, 20, 25, 30 °C but not at 35, 37, 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Phrao District, Nam Phrae, in kaempfer flower (Boesenbergia rotunda), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU586. GenBank numbers PV834575 (D1/D2), PV834745 (ITS), PX622361 (SSU), PX582307 (rpb1).
Notes – Two strains of Sym. saisamorniae (SDBR-CMU710 and SDBR-CMU586) introduced as a new member of Sympodiomycopsis, formed a distinct lineage, closely related to Sym. kandeliae (Fig. 43). Sympodiomycopsis saisamorniae different from Sym. kandeliae by 0.5% nucleotide divergence (3 nt substitutions) in D1/D2, 2% (13 nt substitutions) in the ITS regions, and 11.40% nucleotide divergence (80 nt substitutions and 21 gaps) in rpb1. Moreover, Sym. saisamorniae can be distinguished from Sym. kandeliae in its physiological tests (Table 13).
Class: Ustilaginomycetes Warm.
Order: Ustilaginales Bek.
Family: Ustilaginaceae Tul. & C. Tul.
Members of Ustilaginaceae are facultative parasites that have a yeast-like or filamentous saprophytic phase, which causes smut in corn, wheat, and grass crops. In biotechnology, Ustilaginaceae have the potential applications in the food, pharmaceutical, and chemical industry with substances glycolipids, organic acids, and polyols[113−115]. Almost all Ustilaginaceae species have a dimorphic life cycle that includes an asexual, saprophytic yeast-like stage and a filamentous sexual stage, which is required to parasitize a host. The aerial mycelium consists of ramifying, acropetal chains of fusiform conidia. Chlamydospores may be present, but ballistoconidia are absent[92,116]. Currently, 25 genera are listed in this family including Ahmadiago, Aizoago, Anomalomyces, Anthracocystis, Bambusiomyces, Centrolepidosporium, Dirkmeia, Eriocaulago, Eriosporium, Franzpetrakia, Kalmanozyma, Langdonia, Macalpinomyces, Melanopsichium, Moesziomyces, Parvulago, Pattersoniomyces, Pseudozyma, Shivasia, Sporisorium, Stollia, Tranzscheliella, Triodiomyces, Ustilago, and Yunchangia[41]. In this study, Anthracocystis heteropogonicola (two strains), Moesziomyces antarcticus (four strains), Mo. bullatus (five strains), Mo. parantarcticus (four strains), Pseudozyma chiangmaiensis sp. nov. (seven strains), Ps. lannaensis sp. nov. (five strains), Ps. limtongiae sp. nov. (four strains), Ps. pollinicola sp. nov. (four strains), and Ps. saisamorniae sp. nov. (eight strains) are presented (Fig. 48, Supplementary File 1)
Pseudozyma chiangmaiensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 47)
MycoBank number: MB860211
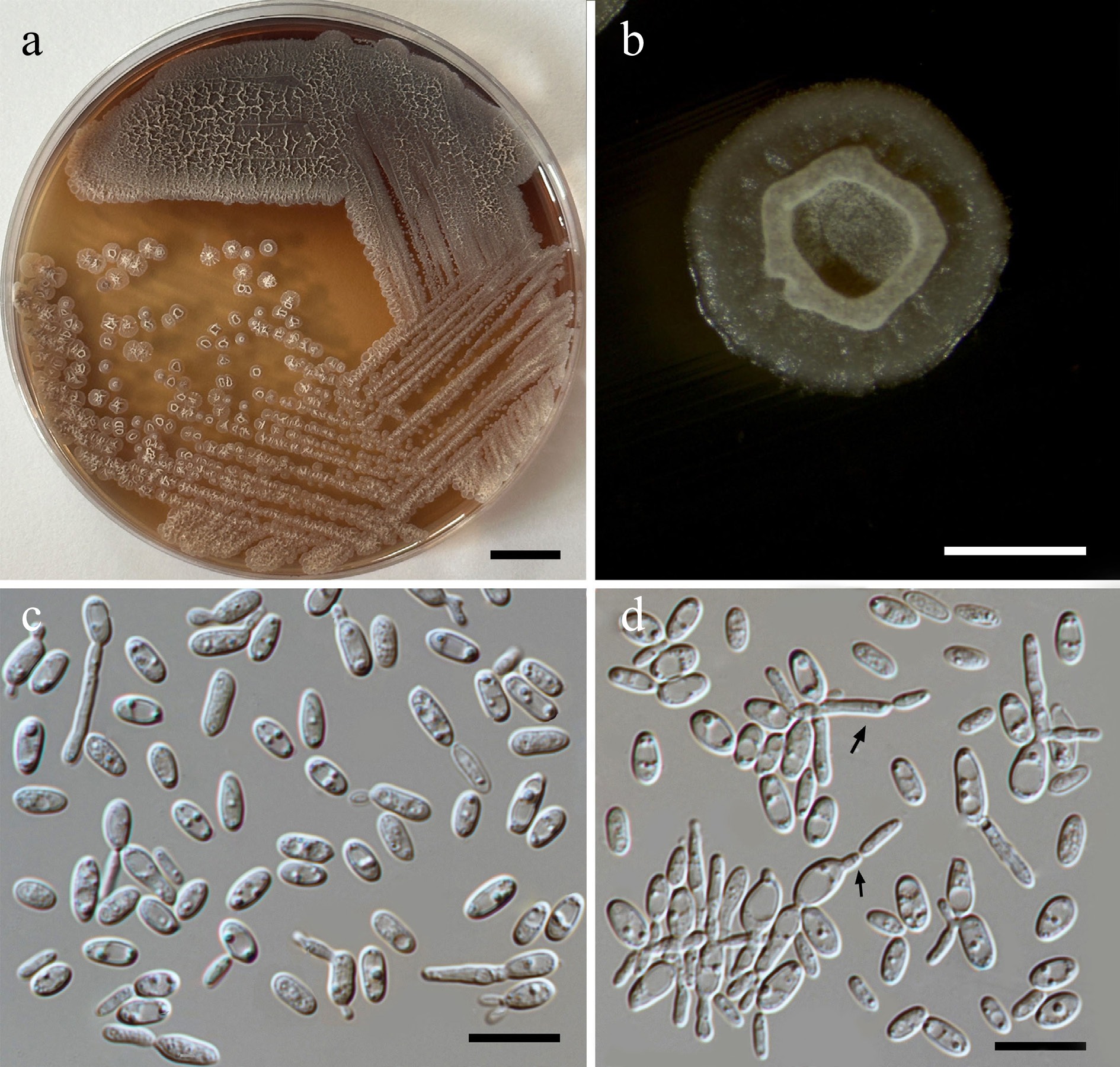
Figure 47.
Morphological characteristics of Pseudozyma chiangmaiensis (SDBR-CMU611, ex-type). (a) Culture, (b) single colony, (c), (d) cells rudimentary pseudohyphae (indicated by arrows) on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'chiangmaiensis' referring to Chiang Mai Province, where the new species was found.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Suthep, in Fukien tea flower (Carmona retusa), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40082 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU611 = GMBCC2423 = TBRC21397. GenBank numbers PV834594 (D1/D2), PV834764 (ITS), PX622365 (SSU), PX582308 (rpb1), PX582346 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (1.8–3.0 mm in diameter), yellowish white, umbonate, zonate towards the margin, and with the margin fringed and producing brown pigment into agar. The cells are ovoid to ellipsoidal (2.04–3.25 × 4.31–8.15 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strain and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose (weak), melibiose, lactose, raffinose, melizitose, glycerol, erythritol, ribitol, D-glucitol, mannitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-glucuronate, D-galacturonic acid, DL-lactate (weak), succinate (weak), citrate, and xylitol are assimilated, but L-rhamnose, salicin, inulin, soluble starch, galactitol, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine, and cadaverine are all assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth is present at 10, 15, 25, and 30 °C, but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in Persian gentian flower (Exacum affine), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU581; confederate vine flower (Antigonon leptopus), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU596; Indian cork flower (Millingtonia hortensis), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU725; Suthep, in Ixora flower (Ixora chinensis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU587; Phrao District, Nam Phrae, in ylang-ylang tree flower (Cananga odorata), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU648; canna lily flower (Canna indica), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU682. GenBank numbers SDBR-CMU581: PV834591 (D1/D2), PV834761 (ITS), PX622362 (SSU); SDBR-CMU587: PV834592 (D1/D2), PV834762 (ITS), PX622363 (SSU); SDBR-CMU596: PV834593 (D1/D2), PV834763 (ITS), PX622364 (SSU); SDBR-CMU648: PV834595 (D1/D2), PV834765 (ITS), PX622366 (SSU); SDBR-CMU682: PV834596 (D1/D2), PV834766 (ITS), PX622367 (SSU); SDBR-CMU725: PV834597 (D1/D2), PV834767 (ITS), PX622368 (SSU).
Notes – Pseudozyma was introduced by Bandoni, based on the type species, Pseudozyma prolifica[117]. Currently the genus comprises 13 epithets; however, only eight epithets are supported by molecular data[47]. Due to their uncertain phylogenetic positions, the taxonomic placement of some Pseudozyma species remains unresolved. Thus, the term 'pro tempore (pro tem.)' was purposed to denote single-species lineages that are temporarily maintained, viz. Ps. alboarmeniaca pro tem., Ps. thailandica pro tem., Ps. hubeiensis pro tem., and Ps. pruni pro tem.[65]. In a BLASTn search of NCBI GenBank, the closest match of the D1/D2 domain and ITS sequences of Pseudozyma strains isolated in this study was P. hubeiensis with 98%–100% and 96%–100% similarity, respectively. Based on a concatenated D1/D2 domain and ITS sequence dataset, these strains clearly formed five distinct subclades, each corresponding to a separate species (Fig. 48, Supplementary File 1). A comparison of D1/D2 and ITS sequences among the species indicates that each species differs from the others (Table 14). Likewise, they can also be distinguished by colony characteristics, cells, and pseudohypha characters (Figs 47, 49−52). Although many strains on various substrates worldwide have been identified as Ps. hubeiensis, molecular evidence suggests that certain strains (i.e., AD_L20, AD_L22, DMic 154889, PYCC 9351, and Py1C; Fig. 48) may represent separate species. These strains are tentatively named as Ps. hubeiensis until more collections and sequence data clarify their taxonomic status. Additionally, phenotypic differences between Ps. chiangmaiensis, Ps. lannaensis, Ps. limtongiae, Ps. pollinicola, Ps. saisamorniae, and Ps. hubeiensis are shown in Table 15.
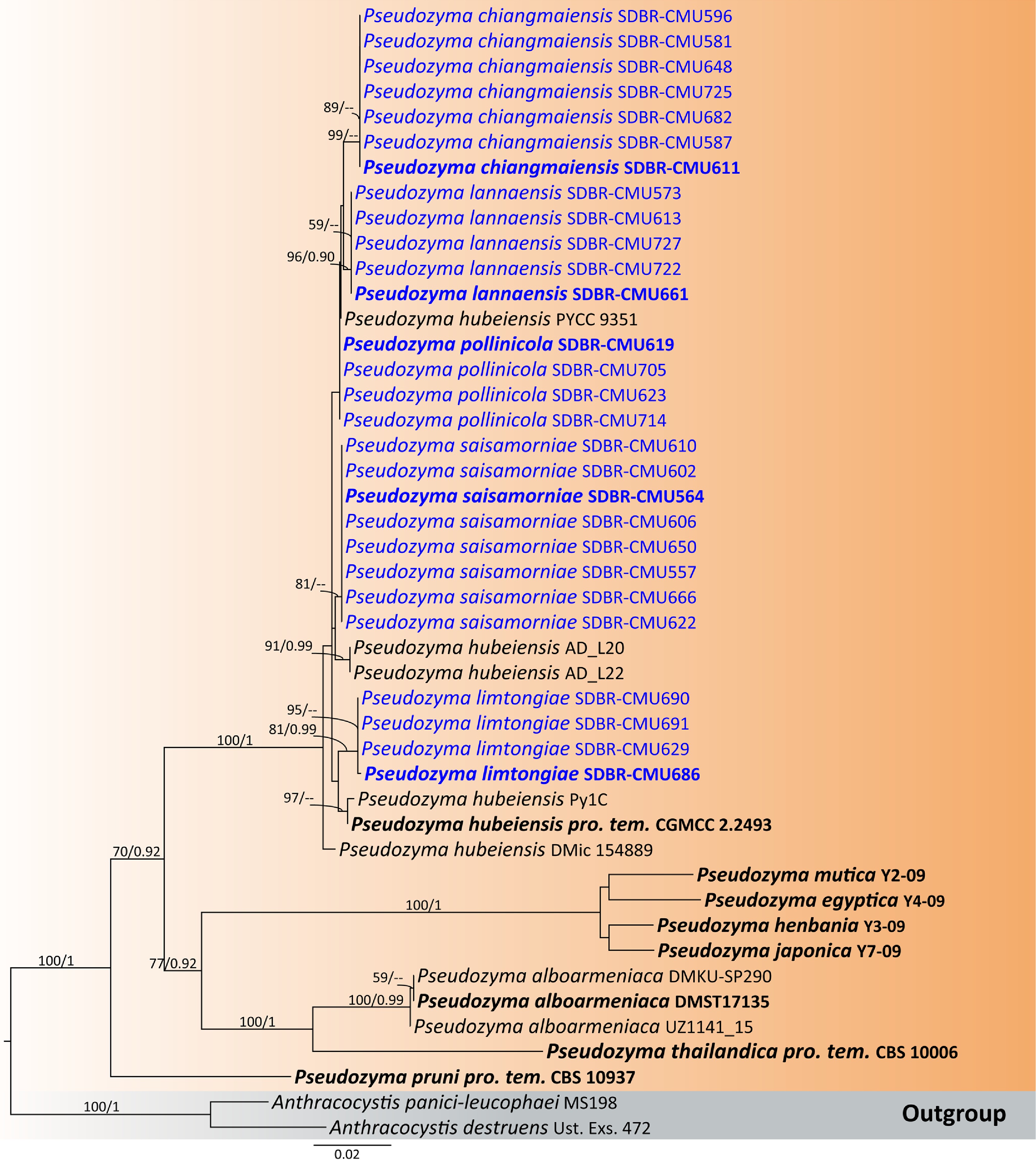
Figure 48.
Phylogenetic tree generated by maximum likelihood analysis of the combined D1/D2 domain of LSU and ITS sequence data representing Pseudozyma. The tree is rooted to Anthracocystis destruens (Ust. Exs. 472) and A. panici-leucophaei (MS198). Single-locus analyses were also performed, and topology and clade stability were compared from combined gene analyses. Fourty-five strains are included in the combined sequence analysis, which comprise 1,704 characters with gaps. The average standard deviation of the split frequencies of the BI analysis was 0.004541. Bootstrap support values for maximum likelihood ≥ 50% (ML, left) and Bayesian posterior probabilities ≥ 0.90 (PP, right) are indicated above the node. Double dashes (--) represent support values less than 50% ML/0.90 PP. The scale bar represents the expected number of nucleotide substitutions per site. The ex-type strains are in bold, and the newly generated sequences in this study are in blue.
Table 14. Percentage of nucleotide divergence in the ITS region among the species most closely related to Pseudozyma hubeiensis.
Taxa Ps. chiangmaiensis Ps. lannaensis Ps. limtongiae Ps. pollinicola Ps. saisamorniae Ps. hubeiensis Ps. chiangmaiensis – 1.20% (9 nt) 3.28% (25 nt) 1.07% (8 nt) 1.98% (15 nt) 2.73% (19 nt) Ps. lannaensis 1.20% (9 nt) – 3.27% (25 nt) 1.07% (8 nt) 1.19% (19 nt) 1.87% (13 nt) Ps. limtongiae 3.28% (25 nt) 3.13% (25 nt) – 2.62% (20 nt) 2.48% (19 nt) 2.97% (21 nt) Ps. pollinicola 1.07% (8 nt) 1.07% (8 nt) 2.62% (20 nt) – 1.72% (13 nt) 1.87% (13 nt) Ps. saisamorniae 1.98% (15 nt) 2.51% (19 nt) 2.48% (19 nt) 1.72% (13 nt) – 2.29% (16 nt) Ps. hubeiensis 2.73% (19 nt) 1.87% (13 nt) 3.03% (21 nt) 1.87% (13 nt) 2.29% (16 nt) – Each value indicates that nucleotide divergence (%) between pairs of species, the number of nucleotide substitutions (nt) between the indicated species pairs. Phylogenetic analysis (Fig. 48) shows that Ps. chiangmaiensis is sister to Ps. lannaensis. Pseudozyma chiangmaiensis is similar to Ps. lannaensis in having ovoid to ellipsoidal cells (Figs 47, 49). Budding cells of Ps. chiangmaiensis are polar, while Ps. lannaensis are polar on short stalk. Furthermore, Ps. chiangmaiensis forms colonies with umbonate elevation and produces a brown pigment diffusing into the agar. Whereas Ps. lannaensis forms colonies with umbonate elevation and filamentous margins. Physiologically, Ps. chiangmaiensis differs from Ps. lannaensis and Ps. saisamorniae in its ability to utilize gluconate. Compared to Ps. pollinicola, it shows a different ability to assimilate ribitol. Additionally, Ps. chiangmaiensis differs from Ps. hubeiensis in its ability to utilize sorbose, myo-inositol, and citrate, whereas Ps. hubeiensis cannot grow on these substrates. Moreover, Ps. chiangmaiensis is capable of growing in the presence of 50% glucose, while Ps. hubeiensis cannot grow[65] (Table 15).
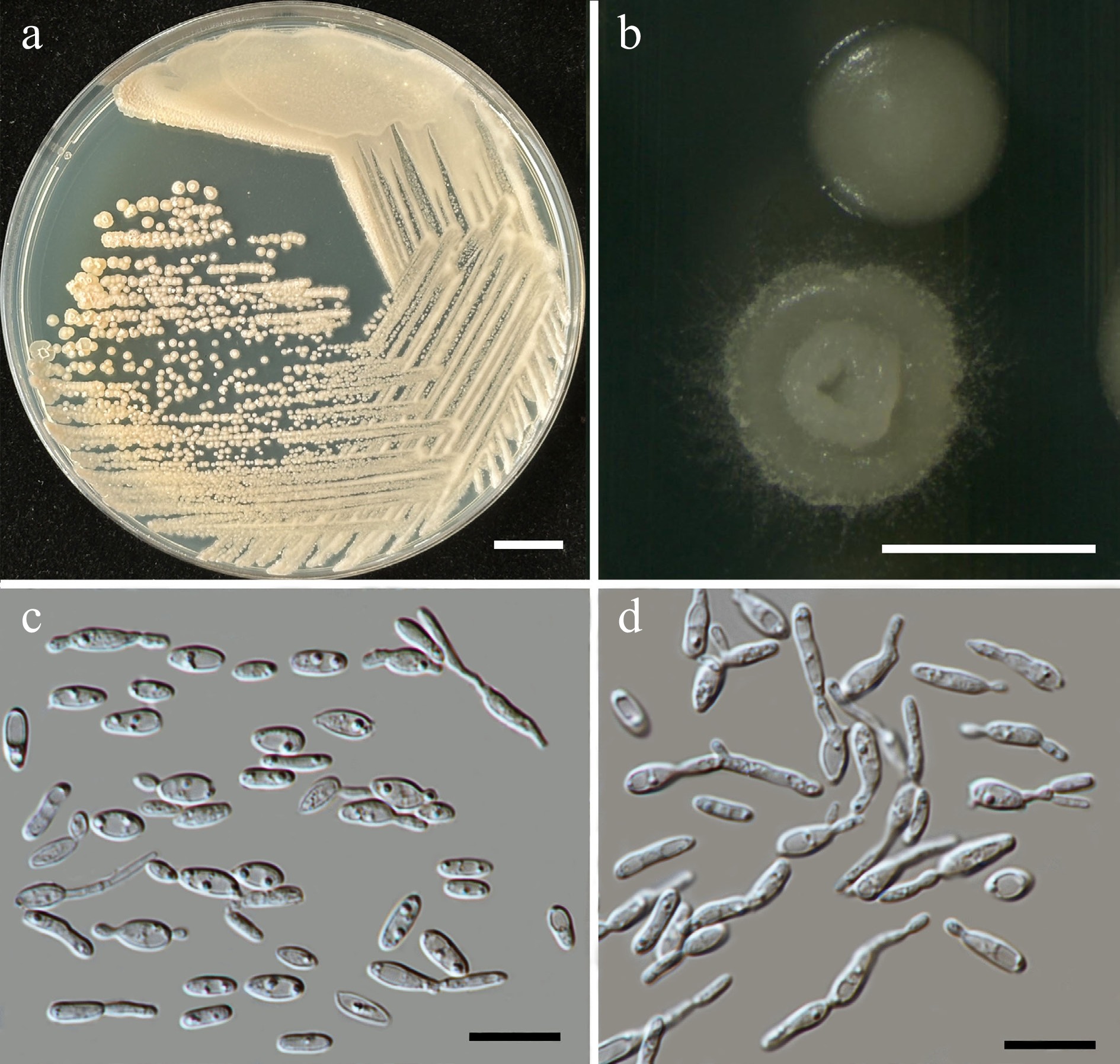
Figure 49.
Morphological characteristics of Pseudozyma lannaensis (SDBR-CMU661, ex-type). (a) Culture, (b) single colony, (c) cells, and (d) rudimentary pseudohyphae on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Table 15. Phenotypic characteristics differentiating Pseudozyma chiangmaiensis, Ps. lannaensis, Ps. pollinicola, Ps. limtongiae, and Ps. saisamorniae from closely related Pseudozyma species.
Characteristics 1 2 3 4 5 6 Carbon assimilation L-Sorbose + + + + + – N-Acetyl glucosamine + + + w + + L-Arabinose + w + + + + D-Arabinose + w + + + + Cellobiose + w + w w + Salicin w w + w – – Soluble starch – – – – – +/w Glycerol + w + + – + Erythritol + w + w – + Ribitol + w + – w + D-Glucitol + + + + w + D-Mannitol + + + + w + myo-Inositol + + + + + – D-Gluconate + – + w – nd D-Galacturonic acid + + w w + nd DL-Lactate w + + w + – Succinate w + + + + + Citrate + + + + + – Nitrogen assimilation Sodium nitrite + + + w + + Ethylamine HCl + + + w + + Growth characteristics Growth at 35 °C – w – – – nd Growth at 37 °C – w – – – + Growth on 50% Glucose + + + + w – Strains 1: Ps. chiangmaiensis sp. nov., 2: Ps. lannaensis, 3: Ps. limtongiae, 4: Ps. pollinicola, 5: Ps. saisamorniae, and 6. Ps. hubeiensis[65]. Pseudozyma lannaensis Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 49)
MycoBank number: MB860212
Etymology – 'lannaensis' referring to the Kingdom of Lanna, the historic name of northern Thailand, where the new species was found.
Holotype – THAILAND, Phayao Province, Mueang District, in vinca flower (Catharanthus roseus), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40084 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU661 = GMBCC2424 = TBRC21406. GenBank numbers PV834600 (D1/D2), PV834770 (ITS), PX622371 (SSU), PX582309 (rpb1), PX582330 (rpb2), PX582347 (tef1-α).
Description – The culture on YMA after 5 days at 25 °C, colonies are circular from (0.8–1.5 mm in diameter), pale brownish-yellow or brownish, initially dull, smooth with entire margin, becoming umbonate with undulate and filamentous margins. The cells are ovoid and ellipsoidal (2.02–3.55 × 4.65–9.12 μm, n = 50), occur singly or in pairs. Budding is polar on a short stalk. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose (weak), D-arabinose (weak), sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose (weak), salicin (weak), melibiose, lactose, raffinose, melizitose, glycerol (weak), erythritol (weak), ribitol (weak), D-glucitol, mannitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-galacturonic acid (weak), DL-lactate, succinate, citrate, and xylitol are assimilated, but L-rhamnose, inulin, soluble starch, galactitol, D-glucuronate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine, and cadaverine are all assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth is present at 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in ground cherry flower (Physalis minima), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU573; Suthep, in White cheesewood flower (Alstonia scholaris), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU727; Mae Hia, in yellow trumpetbush flower (Tecoma stans), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU613; San Kamphaeng District, Ton Pao, in Queen's flower (Lagerstroemia speciosa), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU722. GenBank numbers SDBR-CMU573: PV834598 (D1/D2), PV834768 (ITS), PX622369 (SSU); SDBR-CMU613: PV834599 (D1/D2), PV834769 (ITS), PX622370 (SSU); SDBR-CMU722: PV834601 (D1/D2), PV834771 (ITS), PX622372 (SSU); SDBR-CMU727: PV834602 (D1/D2), PV834772 (ITS), PX622373 (SSU).
Notes – The new species, Pseudozyma lannaensis is similar to Ps. saisamorniae in having ovoid and ellipsoidal cells that formed polar budding on short stalk. Pseudozyma lannaensis forms colonies with umbonate elevation and filamentous margins, while Ps. saisamorniae formed wrinkled colonies when mature. In addition, cells of Ps. lannaensis are shorter than the latter (2.02–3.55 × 4.65–9.12 μm vs 2.2–3.31 × 4.43–7.83 μm). Physiologically, Ps. lannaensis differs from Ps. hubeiensis in its ability to utilize sorbose, myo-inositol, lactate and citrate, whereas Ps. hubeiensis cannot grow on these substrates. Moreover, Ps. lannaensis was able to grow on 50% glucose, while Ps. hubeiensis could not grow at that medium[65]. Compared to Ps. chiangmaiensis, Ps. limtongiae, and Ps. pollinicola, them shows different ability to assimilate gluconate (Table 15).
Pseudozyma limtongiae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 50)
MycoBank number: MB860213
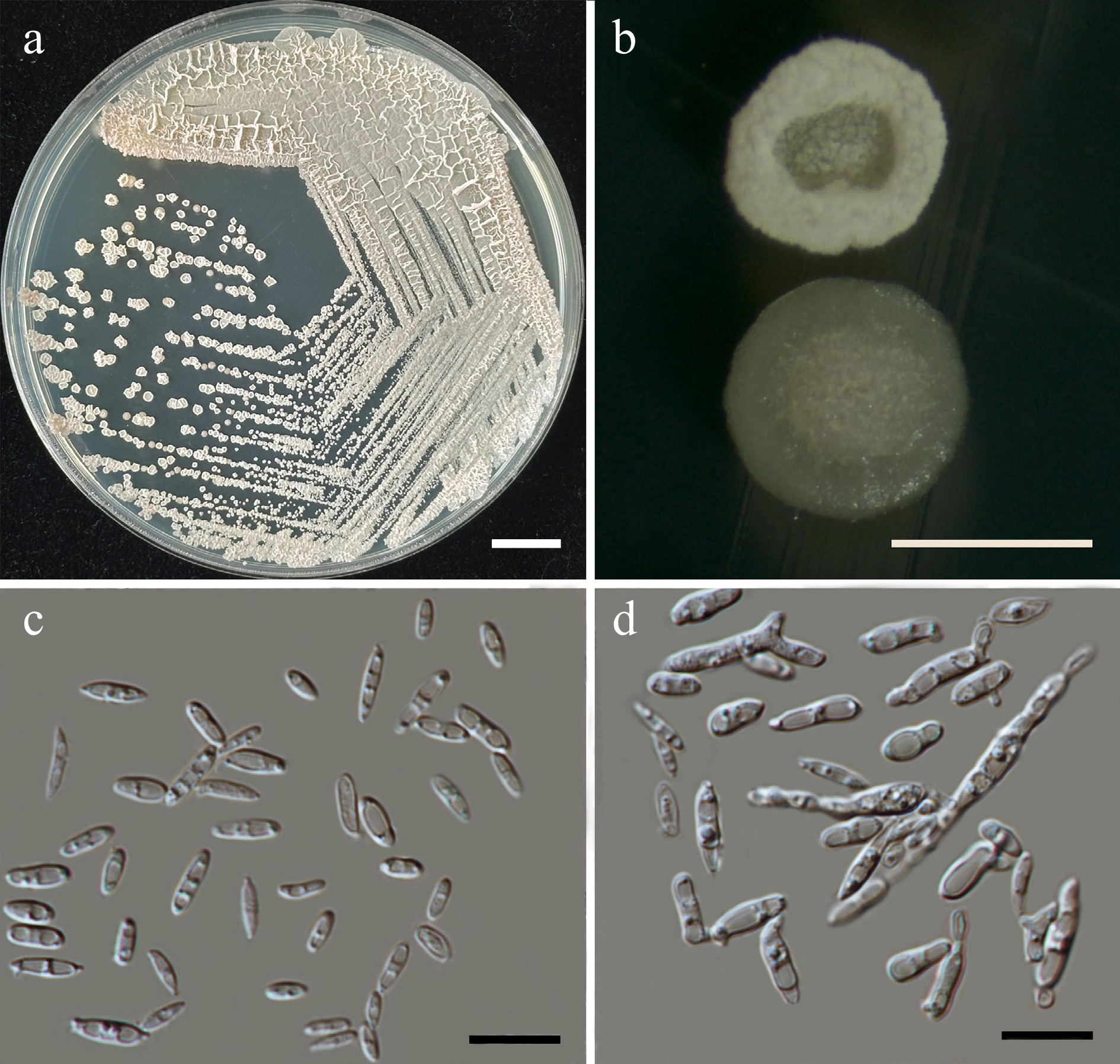
Figure 50.
Morphological characteristics of Pseudozyma limtongiae (SDBR-CMU686, ex-type). (a) Culture, (b) single colony, (c) cells, and (d) rudimentary pseudohyphae on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'limtongiae' named in honor of mycologist Savitree Limtong, for her contributions to yeast systematics.
Holotype – THAILAND, Phayao Province, Mueang District, in bridal bouquet flower (Plumeria pudica), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40085 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU686 = GMBCC2425 = TBRC21412. GenBank numbers PV834616 (D1/D2), PV834786 (ITS), PX622383 (SSU), PX582312 (rpb1), PX582333 (rpb2), PX582350 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, colonies are circular form (0.8–1.5 mm in diameter), yellowish white, initial smooth, glistening, convex with entire margin, becoming crateriform when mature. The cells are fusiform or cylindrical (1.32–2.87 × 4.33–9.15 μm, n = 50), occur singly or in pairs. Budding is polar on short stalks. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose, salicin, melibiose, lactose, raffinose, melizitose, glycerol, erythritol, ribitol, D-glucitol, mannitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate, D-galacturonic acid (weak), DL-lactate, succinate, citrate, and xylitol are assimilated, but L-rhamnose, inulin, soluble starch, galactitol, D-glucuronate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine and cadaverine are all assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide and 0.1% cycloheximide. Acid formation is negative. Growth is present at 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – THAILAND, Phayao Province, Mueang District, in bridal bouquet flower (Plumeria pudica), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU690 and SDBR-CMU691; Chiang Mai Province, Mueang District, Suthep, in teak flower (Tectona grandis), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU629. GenBank numbers SDBR-CMU629: PV834615 (D1/D2), PV834785 (ITS), PX622382 (SSU); SDBR-CMU690: PV834617 (D1/D2), PV834787 (ITS), PX622384 (SSU); SDBR-CMU691: PV834618 (D1/D2), PV834788 (ITS), PX622385 (SSU).
Notes – Pseudozyma limtongiae shares morphological similarity with Ps. hubeiensis (CGMCC 2.2493) in having cylindrical with polar budding on a short stalk[118]. However, they can be distinguished by their phylogenetic relationships, colony morphology and phenotypic characteristics. Physiologically, Ps. limtongiae differs from Ps. hubeiensis in its ability to utilize sorbose, salicin, myo-inositol, lactate and citrate, whereas Ps. hubeiensis cannot assimilated on these substrates. Additionally, Ps. limtongiae was able to grow on 50% glucose, while Ps. hubeiensis could not grow at that medium[65]. Compared to Ps. saisamorniae, that Ps. limtongiae ability to utilize glycerol and erythritol but Ps. saisamorniae cannot assimilate (Table 15).
Pseudozyma pollinicola Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 51)
MycoBank number: MB860214
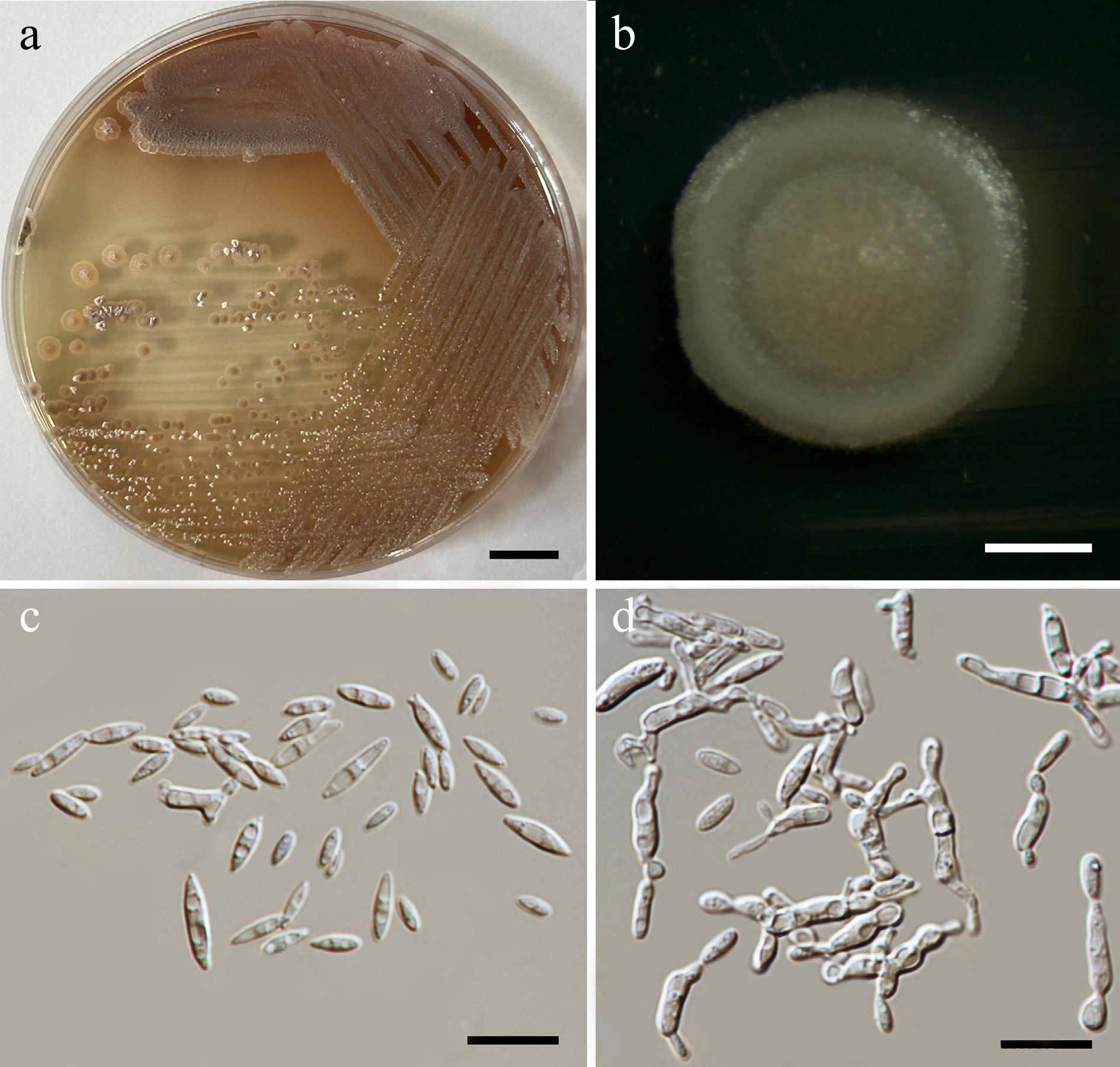
Figure 51.
Morphological characteristics of Pseudozyma pollinicola (SDBR-CMU619, ex-type). (a) Culture, (b) single colony, (c) cells, and (d) rudimentary pseudohyphae on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – The specific epithet 'pollinicola' refers to the substrate origin of the type strain, pollen structure.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in yellow-flowered waterhyssop (Mecardonia procumbens), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40083 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU619 = GMBCC2426 = TBRC21400. GenBank numbers PV834603 (D1/D2), PV834773 (ITS), PX582310 (rpb1), PX582331 (rpb2), PX582348 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, the colonies are circular form (0.8–1.2 mm in diameter), brownish-yellow or brownish, butyrous, dull and wrinkled, producing brown pigment in agar. The cells are ellipsoidal to fusoid (1.53–3.14 × 4.29–9.39 μm, n = 50), occur singly or in pairs. Budding is polar. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine (weak), ribose, xylose, L-arabinose, D-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose (weak), salicin (weak), melibiose, lactose, raffinose, melizitose, glycerol, erythritol (weak), D-glucitol, mannitol, myo-inositol, D-glucono-1,5-lactone, D-gluconate (weak), D-glucuronate, D-galacturonic acid (weak), DL-lactate (weak), succinate, citrate, and xylitol are assimilated, but L-rhamnose, inulin, soluble starch, ribitol, galactitol, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine, and cadaverine are all assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose and 60% glucose (weak). No growth occurs on media containing 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is negative. Growth is present at 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – Thailand, Chiang Mai Province, Mueang District, Chang Phueak, in jasmine flower (Jasminum sambac), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU623; cape leadwort (Plumbago auriculata) flower, August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU705; marguerite daisy flower (Argyranthemum frutescens), September 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU714. GenBank numbers SDBR-CMU623: PV834604 (D1/D2), PV834774 (ITS); SDBR-CMU705: PV834605 (D1/D2), PV834775 (ITS); SDBR-CMU714: PV834606 (D1/D2), PV834776 (ITS).
Notes – Pseudozyma pollinicola produces a brown pigment diffusing into the agar similar with Ps. chiangmaiensis but can be distinguished by forming round with raise margin colonies and having ellipsoidal to fusoid cells. Moreover, Ps. pollinicola can be distinguished from Ps. hubeiensis by its ability to assimilate sorbose, myo-inositol and citrate as well as by its ability to grow on media containing 50%[65]. Additionally, Ps. pollinicola distinguished from Ps. saisamorniae by its ability to assimilate salicin and glycerol (Table 15).
Pseudozyma saisamorniae Kodchasee, Senwanna, J. Kumla, & N. Suwannar., sp. nov. (Fig. 52)
MycoBank number: MB860215
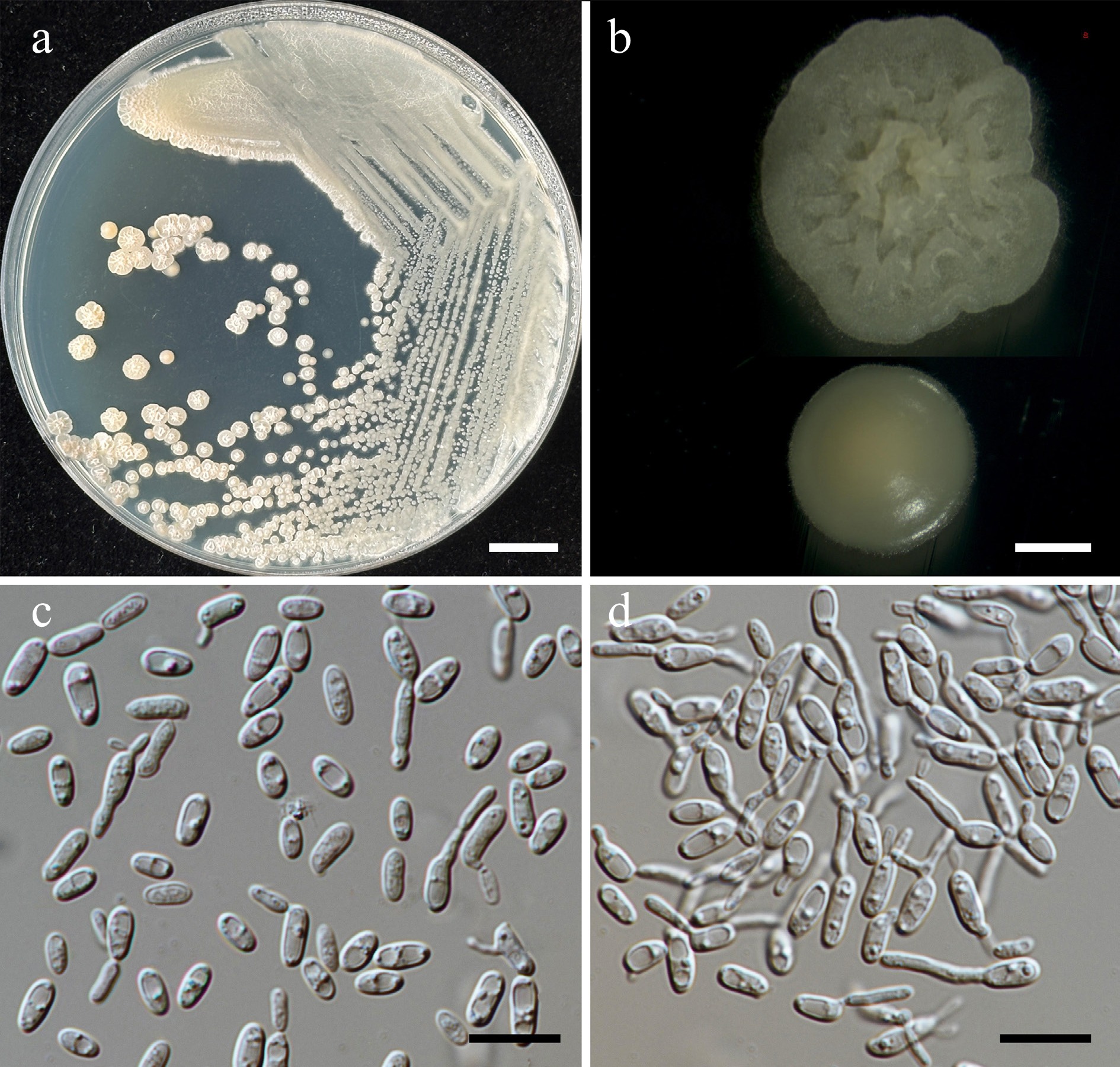
Figure 52.
Morphological characteristics of Pseudozyma saisamorniae (SDBR-CMU564, ex-type). (a) Culture, (b) single colony, (c) cells, and (d) rudimentary pseudohyphae on YMA after 5 d at 25 °C. Scale bars: (a) = 10 mm, (b) = 1 mm, (c), (d) = 10 μm.
Etymology – 'saisamorniae' referring to the name of mycologist Saisamorm Lumyong, in honor of her 75th birthday.
Holotype – THAILAND, Chiang Mai Province, Mueang District, Chang Phueak, in angelonia flower (Angelonia goyazensis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, holotype, CMUB40116 (preserved in metabolically inactive state), ex-type living culture SDBR-CMU564 = GMBCC2427 = TBRC21386. GenBank numbers PV834608 (D1/D2), PV834778 (ITS), PX622375 (SSU), PX582311 (rpb1), PX582332 (rpb2), PX582349 (tef1-α).
Description – The culture on YMA after 5 d at 25 °C, the colonies are circular form (1.2–3.15 mm in diameter), yellowish white, initially dull, smooth with entire margin, becoming wrinkled with undulate margins. The cells are ovoid and ellipsoidal (2.2–3.31 × 4.43–7.83 μm, n = 50), occur singly or in pairs. Budding is polar on a short stalk. Ballistoconidia were not produced. In Dalmau plates after 2 weeks on cornmeal agar and PDA at 25 °C, pseudohyphae and true hyphae are formed. Basidiospores were not obtained for individual strains and strain pairs on YMA, CMA, 5% MEA, PDA, and V8 agar after incubation at 25 °C for one month.
Fermentation of glucose is negative. D-Glucose, galactose, sorbose, N-acetyl glucosamine, ribose, xylose, L-arabinose, D-arabinose, sucrose, maltose, α-α-trehalose, methyl-α-D-glucoside, cellobiose (weak), melibiose, lactose, raffinose, melizitose, ribitol (weak), D-glucitol (weak), mannitol (weak), myo-inositol, D-glucono-1,5-lactone, D-glucuronate, D-galacturonic acid, DL-lactate, succinate, citrate, and xylitol are assimilated, but L-rhamnose, salicin, inulin, soluble starch, glycerol, erythritol, galactitol, D-gluconate, methanol, and ethanol are not assimilated. Ammonium sulfate, potassium nitrate, sodium nitrite, ethylamine, L-lysine, and cadaverine are all assimilated as sole nitrogen source. Growth occurs on media containing 50% glucose (weak). No growth occurs on media containing 60% glucose, 10% NaCl/5% glucose, 16% NaCl/5% glucose, 0.01% cycloheximide, and 0.1% cycloheximide. Acid formation is positive (weak). Growth is present at 10, 15, 25, and 30 °C but not at 35, 37, and 40 °C.
Additional strains examined – Thailand, Chiang Mai Province, Mueang District, Chang Phueak, in Orange jasmine flower (Murraya paniculata), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU557; rose (Rosa sp.), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU610; frangipani (Plumeria obtusa) flower, July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU602; mealycup sage flower (Salvia farinacea) flower, July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU606; Mae Hia, in Siam tulip flower (Curcuma sessilis), July 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU622; Phayao Province, Mueang District, in, in holy basil flower (Ocimum tenuiflorum), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU666; peregrina flower (Jatropha integerrima), August 2024, P. Kodchasee, C. Senwanna, J. Kumla, and N. Suwannarach, living culture SDBR-CMU650. GenBank numbers SDBR-CMU557: PV834607 (D1/D2), PV834777 (ITS), PX622374 (SSU); SDBR-CMU602: PV834609 (D1/D2), PV834779 (ITS), PX622376 (SSU); SDBR-CMU606: PV834610 (D1/D2), PV834780 (ITS), PX622377 (SSU); SDBR-CMU610: PV834611 (D1/D2), PV834781 (ITS), PX622378 (SSU); SDBR-CMU622: PV834612 (D1/D2), PV834782 (ITS) PX622379 (SSU); SDBR-CMU650: PV834613 (D1/D2), PV834783 (ITS), PX622380 (SSU); SDBR-CMU666: PV834614 (D1/D2), PV834784 (ITS), PX622381 (SSU).
Notes – Pseudozyma saisamorniae can be distinguished from the other new taxa described in this study based on colony characteristics, cell morphology, and phylogenetic analyses, as discussed above. In addition, Ps. saisamorniae can be distinguished from Ps. hubeiensis with its ability to assimilate sorbose, myo-inositol, lactate and citrate, but inability to assimilate soluble, starch glycerol and erythritol. Moreover, its ability to grow on media containing 50%[65]. Compared to Ps. chiangmaiensis, Ps. lannaensis, Ps. limtongiae, and Ps. pollinicola, which can utilize salicin, glycerol, and erythritol, Ps. saisamorniae cannot assimilate these compounds (Table 15).
-
Yeasts can be isolated from an exceptionally varied range of sources in ecosystem on Earth. Natural environments serve as primary reservoirs, including soil, freshwater and marine systems, air, and plants[23,119−122]. Animal-associated sources are equally important, encompassing the gastrointestinal tracts, skin, and oral cavities of mammals, birds, and insects, as well as various invertebrate hosts[4,123−126]. Traditional fermented foods and beverages represent another major isolation sources, including beer, bread, cheese, kimchi, wine, yogurt, and various cultured products from around the world[26,127−131] Clinical, medical, and industrial environments also offer valuable opportunities for yeast isolation[120,126,132−134]. Moreover, plant-based materials offer rich isolation opportunities from fruits, vegetables, flowers, grains, nectar, plant exudates, and decomposing organic matter[3,6,135−137]. This study, which investigated anthophilous yeasts from flowers in northern Thailand, revealed high diversity, with 187 yeast strains obtained and identified across nine classes, 17 orders, 22 families, 36 genera, and 73 species, including 33 species newly introduced to science. Their identification was based on polyphasic approaches, including morphology, physiology, biochemistry, and muti-locus phylogenetic analysis according to several previous studies[27,60,63,65,81,98]. In addition, some yeast species obtained in this study, including Curvibasidium chiangmaiensis, Cystobasidium thailandicum, and Symmetrospora hydei, could not be distinguished solely using D1/D2 and ITS sequences, therefore, physiological characteristics and additional sequences (SSU, rpb1, rpb2, and tef1-α) were used to clarify their taxonomy from its closet relative. This result was in agreement with previous studies[65,81,138,139] demonstrating that multi-locus sequence data provided a good resolution regarding relationships and resolve species boundaries. This study demonstrates that flowers represent a rich and diverse habitat for anthophilous yeasts, with the potential for the discovery of novel species. Some yeast species obtained in this study have been reported from flowers, such as Entelexis stigmatis, Cyberlindnera fabianii, Hannaella pagnoccae, Meyerozyma caribbica, Metschnikowia koreensis, Naganishia diffluens, Papiliotrema flavescens, Rhodosporidiobolus ruineniae, Starmerella etchellsii, and Sympodiomycopsis paphiopedi[3,4,8,9,14,45,140]. The present study reveals a dominance of anthophilous basidiomycetous yeasts (77.54%) over those belonging to Ascomycota (22.46%) with Pseudozyma the most frequent basidiomycetous yeast and Metschnikowia being the most frequent ascomycetous yeast. Similarly, Han et al.[8] studied the diversity of anthophilous yeasts from wildflowers around Jangseong Lake in Jeollanam-do, located in South Korea, and found that basidiomycetous yeasts were more abundant (78.33%) than ascomycetous yeasts (21.67%), with Pseudozyma and Metschnikowia being the most frequent basidiomycetous and ascomycetous yeasts, respectively. Canto et al.[3] reported a higher abundance of basidiomycetous yeasts compared to ascomycetous yeasts across a wide diversity of flowers in Mexico with Ustilago the most frequent basidiomycetous yeast and Metschnikowia being the most frequent ascomycetous yeast. Mittelbach et al.[2] studied the diversity of yeasts in flowers on the island of Tenerife in the eastern Anaga Mountains and found that ascomycetous yeasts were more abundant (58.64%) than basidiomycetous yeasts (41.36%), with Metschnikowia being the most frequent ascomycetous yeast and Cryptococcus the most frequent basidiomycetous yeast. Additionally, Hyun et al.[141] isolated anthophilous yeasts from wildflowers in Ulleungdo and Yokjido, Gyeongsangbuk-do, South Korea, and showed different ratios of yeast numbers between ascomycetous and basidiomycetous yeasts. In Ulleungdo, they found that ascomycetous yeasts were more abundant (62.50%) than basidiomycetous yeasts (37.50%), while in Yokjido, they found that basidiomycetous yeasts were more abundant (60%) than ascomycetous yeasts (40%). Rhodotorula is the most abundant basidiomycetous genus found in both Ulleungdo and Yokjido, whereas the most abundant ascomycetous genera found in Ulleungdo and Yokjido were Pichia and Metschnikowia, respectively. However, Pozo et al.[7] investigated the yeast communities in flower nectar from Cazorla-Segura-Las Villas Natural Park, southeastern Spain, and found that ascomycetous yeasts were more abundant (92.59%) than basidiomycetous yeasts (7.14%), with Metschnikowia being the most frequent ascomycetous yeast and Rhodotorula being the most frequent basidiomycetous yeast. Kanpiengjai et al.[9] isolated anthophilous yeasts from the flowers of Camellia sinensis var. assamica in northern Thailand and found that ascomycetous yeasts (82.93%) were significantly more abundant than basidiomycetous yeasts (17.07%), with Pseudozyma being the most frequent basidiomycetous yeast and Metschnikowia being the most frequent ascomycetous yeast.
The diversity of anthophilous yeasts is influenced by multiple interrelated factors. Canto et al.[3] suggest that the primary factor influencing yeast community composition is the specific interaction between yeasts and host plants, as different flowers offer unique habitats and resources such as varying nectar compositions and floral structures. Lachance[142] found that only ascomycetes from the order Metschnikowiaceae are related to anthophilous yeasts due to their ability to utilize sucrose, the main component of nectar. Mittelbach et al.[2] found that ascomycetous yeasts tend to inhabit sucrose-dominant nectars, whereas basidiomycetous yeasts are more commonly associated with nectars rich in monosaccharides, reflecting the variation in sugar composition among different flower species. Additionally, Peay et al.[143] has shown that nectar habitats stand out by high sugar concentrations, microaerophilic conditions, low nitrogen levels, and the widespread presence of anti-microbial compounds may regulate the colonization of anthophilous yeasts and influence their diversity. The diversity and behavior of pollinators and other insect visitors (such as bees, bumblebees, butterflies, and birds) significantly affect anthophilous yeast dispersal and colonization among flowers[2,4,6]. For example, Cystobasidium minutum, Meira argovae, and Rhodotorula toruloides are anthophilous yeasts that have been found in honeycombs and honeybees, with the pollinator Apis species potentially acting as a vector in their dispersal[3,144,145]. Mittelbach et al.[2] found that nectar sugar concentration, and the frequency of flower visitors (such as birds, bees, and bumblebees) influence the diversity of anthophilous yeasts on the Canary Islands. Environmental conditions, including temperature, humidity, rainfall, wind, and geographic location, further impact the survival and proliferation of anthophilous yeasts[120,146]. Therefore, the factors influencing the diversity of anthophilous yeasts in northern Thailand should be further explored in future studies, as understanding these determinants may provide valuable insights into yeast ecology, insect-plant-microbe interactions, and regional biodiversity. The composition of nectar (including sugars, amino acids, and lipids) of each flower species requires further study. Additionally, the relationship between the yeasts detected in this study, and their surrounding environment, warrants further investigation.
Northern Thailand represents one of the most significant global biodiversity hotspots, particularly for fungal diversity (filamentous fungi, mushrooms, and yeasts)[9,20,23,41,147−153]. This study highlights the diversity of anthophilous yeasts isolated from flowers within this region, which can also be considered an important habitat for yeast diversity, and may potentially include new and/or previously undocumented species. Therefore, this study provides valuable information and enhances the understanding of anthophilous yeast diversity. It may also serve as a catalyst, encouraging mycologists to further investigate the distribution, diversity, and ecological roles of anthophilous yeasts not only in Thailand but throughout Asia and globally. Furthermore, both previously known and novel yeast strains identified in this study represent valuable resources for future biotechnological innovation because these tropical yeasts exhibit unique characteristics (such as osmotolerance, thermal stability, and specialized metabolic pathways) that could support a wide range of industrial applications, including the synthesis of enzymes, bioethanol, natural flavor compounds, and biocontrol agents.
-
This study investigated anthophilous yeasts isolated from floral samples in northern Thailand. From 63 flower specimens, a total of 187 yeast strains were obtained, representing nine classes, 17 orders, 22 families, 36 genera, and 73 species. Molecular characterization revealed that the majority of isolated yeasts belonged to Basidiomycota (77.54%), with the remainder classified as Ascomycota (22.46%). Interestingly, one new order, one new family, 33 new species, and two validated species were introduced based on a polyphasic approach integrating morphological descriptions, physiological characteristics, and multi-locus phylogenetic analyses. Furthermore, this study documented 14 species with newly recognized geographical distributions, one novel habitat association, and 37 first host records. These findings significantly advance knowledge of the biodiversity and biogeography of anthophilous yeasts in tropical ecosystems, revealing a rich but previously underexplored diversity in Thailand and Southeast Asia.
-
Not applicable.
-
The authors confirm contribution to the paper as follows: conceptualization: Kodchasee P, Kumla J, Suwannarach N; formal analysis, validation, data curation: Kodchasee P, Khunnamwong P, Senwanna C, Kumla J, Suwannarach N; software: Kodchasee P, Senwanna C, Tiemsan S, Kumla J; resources, visualization: Kodchasee P, Senwanna C, Kumla J, Suwannarach N; writing − original draft: Kodchasee P, Khunnamwong P, Senwanna C, Kumla J, Suwannarach N; investigation, methodology, writing − review and editing: Kodchasee P, Khunnamwong P, Senwanna C, Duangkon N, Wongsa K, Tiemsan S, Kaewnunta A, Kumla J, Suwannarach N; supervision, project administration, funding acquisition: Suwannarach N. All authors have read and agreed to the published version of the manuscript.
-
The DNA sequence data obtained from this study have been deposited in GenBank under accession numbers: D1/D2 (PV834432 to PV834618), ITS (PV834619 to PV834788), rpb1 (PV941856 to PV941873, PX582293 to PX582312, PX570009, PX570010), rpb2 (PV947453 to PV947472, PX582313 to PX582333, and PX570011 to PX570013), tef1-α (PV947473 to PV947490, PV844824 to PV844827, PX582334 to PX582350, PX570014, and PX570015), and SSU (PV819892 to PV819897, and PX622318 to PX622385). All data analyzed during this study are included in this published article and its supplementary information.
-
This research was financially supported by Chiang Mai University, Thailand.
-
The authors declare that they have no conflict of interest.
- Supplementary File 1 Supplementary materials to this study.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Kodchasee P, Khunnamwong P, Senwanna C, Duangkon N, Wongsa K, et al. 2026. Anthophilous cryptic yeasts: an integrative polyphasic approach reveals novel taxa in northern Thailand. Mycosphere 17: e002 doi: 10.48130/mycosphere-0026-0002 

Anthophilous cryptic yeasts: an integrative polyphasic approach reveals novel taxa in northern Thailand
- Received: 05 August 2025
- Revised: 17 December 2025
- Accepted: 18 December 2025
- Published online: 11 March 2026
Abstract: The diversity of this cryptic yeast group is important for understanding the microbial diversity and ecology of tropical ecosystems. This study focuses on anthophilous cryptic yeasts isolated from flowers in northern Thailand. A total number of 187 yeast strains (73 species) were isolated from 63 flower samples. Analysis of the sequences of the D1/D2 domain of the large subunit (LSU), and the internal transcribed spacer (ITS) regions of ribosomal DNA gene of all yeast strains allowed classification into the phyla Ascomycota (22.46%), and Basidiomycota (77.54%). Additionally, new taxa higher than species level were proposed based on the multi-locus phylogenetic analyses including the D1/D2 domain, the ITS, the small subunit rDNA (SSU), the largest subunit of RNA polymerase II (rpb1), the second largest subunit of RNA polymerase II (rpb2), and translation elongation factor 1 alpha (tef1-α). This study provides morphological descriptions, physiological characteristics, and phylogenetic positions for one new order (Thailandicolales), one new family (Thailandicolaceae), one new genus (Thailandicola), and 33 new species, including three new species belonging to three genera (Priceomyces, Starmerella, and Wickerhamiella) in Ascomycota, and 30 new species occurring in 17 genera (Boekhoutia, Filobasidium, Curvibasidium, Cystobasidium, Halobasidium, Kwoniella, Meira, Papiliotrema, Parajaminaea, Pseudozyma, Rhodotorula, Saitozyma, Sporobolomyces, Symmetrospora, Sympodiomycopsis, Thailandicola, and Vishniacozyma) in Basidiomycota. Fourteen new geographical records, one new habitat, and 37 new records from flower species across 36 genera, 22 families, 17 orders and 9 classes were identified. Furthermore, the invalidly described Entelexis stigmatis and Starmerella orientalis are validated here as two additional new species. These findings significantly expand our knowledge of anthophilous yeast diversity and distribution in tropical flowers.
-
Key words:
- 33 new species /
- Anthophilous yeast /
- Multi-locus phylogeny /
- Polyphasic identification /
- Taxonomy /
- Tropical area