









-
Corticioid and hydnoid fungi are a group of large basidiomycetes that can utilize plant residues as substrates or in symbiotic relationships with live host plants, and these fungi are an important ecological functional group in terrestrial ecosystems, especially in forest ecosystems[1]. The confined definition of corticioid fungi refers to species with resupinate and coriaceous basidiomes with smooth hymenophoral surfaces, which are similar to those of Corticium Pers.[2], and the narrow definition of hydnoid fungi refers to species with toothed hymenophoral surfaces similar to those of Hydnum L.[3]. However, neither corticioid nor hydnoid fungi are established taxonomic units but rather constitute a group of macrofungi with similar hymenophoral morphological features[1,4].
With the extensive application of molecular biological methods, and the rapid development of fungal molecular systematics, accompanied by the constant efforts of taxonomists to develop an accurate natural classification system, there have been significant changes in the scope of defining the morphological characteristics of many genera of aphyllophoroid fungi[5−12]. Consequently, the taxa and morphological characteristics of corticioid and hydnoid fungi, in a broad sense, have also been greatly expanded. In terms of morphology, the corticioid and hydnoid fungi broadly include the species with smooth, tuberculate, grandinioid, odontioid, irpicoid, reticulatoid, merulioid to hydnoid hymenophoral surfaces, except for the poroid, in aphyllophoroid fungi (Fig. 1); the basidiomes can be resupinate, effuse-reflexed, pileate, stereoid, and stipitate hydnoid (such as the Hydnellum P. Karst. and Phellodon P. Karst.), or even thelephoroid (Thelephora Ehrh. ex Willd.); and the texture of basidiomes ranges from coriaceous, crustaceous, membranaceous, farinaceous, cottony, ceraceous, fleshy, or gelatinous.

Figure 1.
Variation in the hymenophoral surfaces and basidiomes of corticioid and hydnoid fungi. (a) Terana coerulea; (b) Gloeopeniophorella convolvens; (c) Tomentella olivacea; (d) Peniophorella rude; (e) Lopharia cinerascens; (f) Serpula himantioides; (g) Irpex lacteus; (h) Steccherinum laeticolor; (i) Mycoleptodonoides aitchisonii; (j) Thelephora ganbajun; (k) Cymatoderma elegans; (l) Hydnum albopallidum; (m) Hydnellum grosselepidotum.
Corticioid and hydnoid fungi have diverse nutritional modes, most of which are saprophytic, and some species can form ectomycorrhizae with woody plants. Saprophytic species are related to needle- and broad-leaved trees, such as living trees, dead trees, fallen trunks, branches, twigs, forest litter, and more, which can cause white- or brown-rot of wood. Some species prefer to grow on soil or produce basidiomes on other macrofungi. Many species in this group of fungi have obvious selectivity for substrates, such as coniferous or broad-leaved trees, different tree species, living or dead trees, coarse fallen trunks or fine twigs, fallen trunks at different stages of decay, etc[13,14]. These species occupy different ecological niches, jointly degrade lignocellulose, and play critical roles in energy flow and material cycling in forest ecosystems[15]. Ectomycorrhizal corticioid and hydnoid species (such as Tomentella Pers. ex Pat. and Thelephora) form symbiotic relationships with many host tree families, including Betulaceae, Dipterocarpaceae, Fagaceae, Orchidaceae, Pinaceae, Tiliaceae, etc.[16], thus obtaining carbohydrates from host plants while providing these host plants with mineral elements such as nitrogen and phosphorus absorbed from the soil. This group of fungi extends their mycelial tissue (rhizomorphs) out of the ground and attaches to fallen trunks, branches, litter, soil, stones, living grasses, ferns, and mosses as substrates, and then, these fungi produce basidiomes.
There are many edible and medicinal fungi among the corticioid and hydnoid fungi that have important economic value[17,18]. Wild ectomycorrhizal and edible fungi, such as Thelephora ganbajun M. Zang (干巴菌) and Sarcodon imbricatus (L.) P. Karst (black tiger palm, 黑虎掌), which currently cannot be artificially cultivated, as well as Hericium erinaceus (Bull.) Pers. (Hedgehog fungus, 猴头菌) and Sparassis latifolia Y.C. Dai & Zheng Wang (Hydrangea fungus, 绣球菌)[19], which have successfully been artificially cultivated, have a delicious flavour and are well-known edible fungi species in East Asia. Many species of corticioid and hydnoid fungi can produce various bioactive substances, such as phellodonic acid, which is produced by Phellodon melaleucus and has strong antibacterial and antitumour activities[20]; polyozellin, which is produced by Polyozellus multiplex (Underw.) Murrill and has therapeutic effects on inflammation, cancer, and oxidative stress-related diseases[21]; various types of scabronins, which are produced by Sarcodon scabrosus (Fr.) P. Karst., and can stimulate the synthesis of nerve growth factors[22]; and polysaccharides, which are extracted from the fermentation broth of Irpex lacteus (Fr.) Fr., and are used to treat chronic glomerulonephritis[23]. Therefore, this group of fungi is also an important source for the discovery of novel drugs.
Numerous saprophytic species of corticioid and hydnoid fungi can produce hydrolytic enzymes, such as laccase and manganese peroxidase (MnP). These enzymes degrade not only lignocellulose but also various environmental pollutants, including but not limited to polycyclic aromatic hydrocarbons (PAHs), synthetic dyes, insecticides, herbicides, fungicides, 2,4,6-trinitrooleene (TNT), benzo[a]pyrene[24,25], dibenzo-p-dioxin (DD), and recalcitrant macromolecular materials, such as polyethylene[26]. Therefore, this group also has broad application potential in environmental remediation. Moreover, these fungi also demonstrate significant industrial application potential in the biopulping of wood chips[27], bioethanol production[28], and other areas.
There are also many cases in which corticioid and hydnoid fungi are used for biological control in agriculture and forestry. For example, Phlebiopsis gigantea (Fr.) Jülich is used to prevent and control tree butt-rot caused by Heterobasidion annosum[29]; Phlebia uda (Fr.) Nakasone can inhibit the spore germination of Fusarium graminearum[30]; Climacodon septentrionalis (Fr.) P. Karst. can immobilize and kill a mycophagous nematode[31]; and Chondrostereum purpureum (Pers.) Pouzar is useful for inhibiting the sprouting and regrowth of freshly cut stems of weedy deciduous trees[32]. Moreover, Sebacina vermifera Oberw. can stimulate seed germination and increase plant growth[33,34].
In the past 15 years, with the support of molecular biological technology, the taxonomy and phylogeny of woody-inhabiting fungi in China have developed rapidly[35−41], and many new taxa of corticioid and hydnoid fungi have been reported, greatly enriching the known fungal species diversity in China[1,42−47]. This paper systematically summarizes the current status of the exploitation and utilization of these species, with the aim of providing guidance for in-depth research, excavation, conservation, and sustainable utilization of these fungal resources.
-
Species data were collected from monographs and papers published as of April 20, 2025. The checklist of Dai[5] was used as a foundation, and we appended the species information of utilization values in medicine, environment, agriculture, and industry. All the taxon names and authorities were verified according to the records updated as of April 20, 2025, in mycological databases, namely, Index Fungorum (
www.indexfungorum.org ), and the MycoBank Database (www.mycobank.org ). The reference DNA sequence accession numbers of the internal transcribed spacer (nrITS) or the large subunit of the nuclear ribosomal RNA (nLSU) gene were retrieved from published papers or GenBank. A classification hierarchy tree file was generated with Lifemap (https://lifemap-ncbi.univ-lyon1.fr/ , 05 May, 2025) using the checklist of collected species, and a diagram was generated with ITOL Version 6.9.1 (https://itol.embl.de/ ) with abundance data.The fresh fruiting bodies of the fungi were dried using an electronic dryer set at temperatures of 40–50 °C. After drying, they were labeled and stored in plastic bags and envelopes. The dried materials studied in this paper are deposited in the herbarium of the Institute of Applied Ecology, Chinese Academy of Sciences (IFP). Macro-morphological features of the fungal specimens were observed either directly or using a stereomicroscope (Nikon SMZ 1000, Tokyo, Japan). Color terms were defined based on the standards provided by Rayner[48] and Munsell[49] Micro-morphological data were obtained from dried specimens and observed under a light microscope (Nikon Eclipse E600 microscope, Tokyo, Japan) following the methods described by Mu et al.[50]. Samples for microscopic examination were individually mounted in Cotton Blue, Melzer's reagent, and 5% potassium hydroxide (KOH). The range of basidiospores provided in the study excludes the smallest and largest 5% of measurements, denoted in parentheses. The following abbreviations are used in this study: KOH = 5% potassium hydroxide water solution, CB = cotton blue, CB+ = cyanophilous, CB– = acyanophilous, IKI = Melzer's reagent, IKI– = both inamyloid and indextrinoid, L = mean spore length, W = mean spore width, Q = the range of length-to-width ratios of the basidiospores studied, and n (a/b) for the total number of basidiospores (a) measured from a specified number of specimens (b).
The newly generated sequences have been deposited in the GenBank database, and were used to establish phylogenetic relationships with other sequences retrieved from GenBank (
www.ncbi.nlm.nih.gov ). BioEdit sequence alignment editor[51]and MEGA 7[52] were used for sequence alignment. Phylogenetic analyses were performed by Maximum Likelihood[53] (RAxML v.1.5b2) with 1,000 rapid bootstrap replicates. Bayesian analysis was carried out using MrBayes 3.2.4[54] implementing the Markov chain Monte Carlo (MCMC) technique and parameters predetermined with the nucleotide substitution models as estimated by MrModelTest v. 2.3[55]. Four simultaneous Markov chains were run starting from random trees and keeping one tree every 100th generation until the average standard deviation of split frequencies was below 0.01. The value of burn-in was set to discard 25% of trees when calculating the posterior probabilities. The phylogenetic trees were visualized using FigTree v1.4.3[56]. Branches that received bootstrap support for ML (ML-BS) ≥ 70% and BI (BPP) ≥ 0.95 were considered significantly supported, respectively. -
A total of 1,400 species of corticioid and hydnoid fungi in China, which belong to 245 genera, 69 families, and 32 uncertain family-level taxa (incertae sedis), as well as 21 orders, and three uncertain order-level taxa in two classes (Agaricomycetes and Dacrymycetes), were compiled (Supplementary Table S1 and Fig. 2). Among the 21 orders, Polyporales has the highest number of species, with a total of 378, followed by Hymenochaetales, Russulales, and Thelephorales, with species numbers of 327, 218, and 166, respectively. At the family level, Hymenochaetaceae (belonged to Hymenochaetales) has the greatest number of species (142), following by Phanerochaetaceae (129 species), Schizoporaceae (127 species), and Thelephoraceae (113 species). At the genus level, Hymenochaete has the greatest number of species (115), followed by Phanerochaete, Tomentella, Xylodon, Hyphoderma, Hydnellum, Lyomyces, and Trechispora, with 66, 64, 53, 51, 44, 38, and 36 species, respectively. However, there are a relatively large number of single-species genera, with 149 genera consisting of 1–2 species, accounting for 60.8% of the total genera.
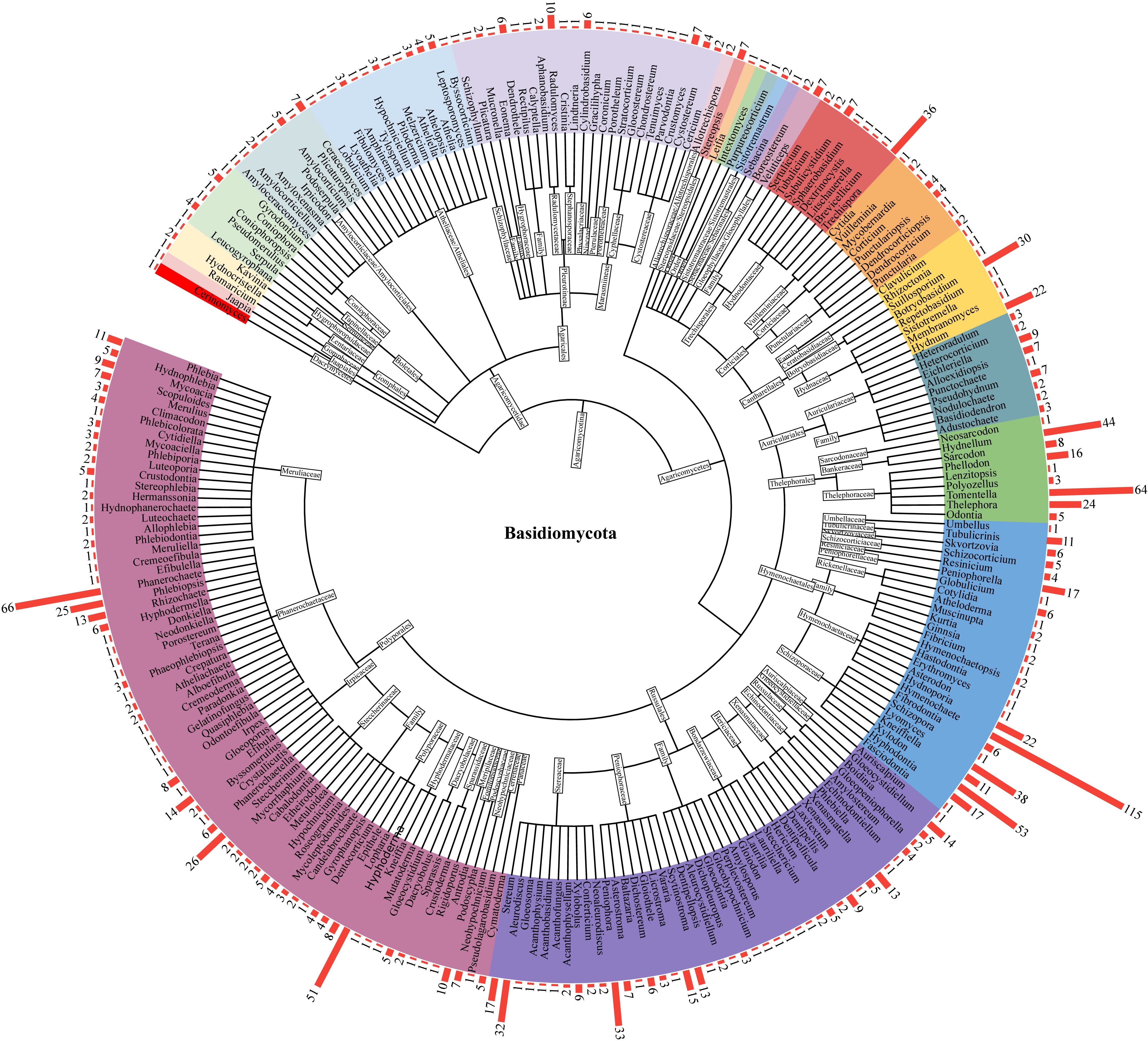
Figure 2.
Diagram showing the classification hierarchy and species abundance of corticioid and hydnoid fungi in China. The red bar on the outer circle represents the number of species in the corresponding genus.
Among the 245 genera of corticioid and hydnoid fungi included in this study, 60 genera exhibit diverse biological activities and have extensive application and development potential in medicine, environment, agriculture, and industry. The majority of the 60 genera have known medicinal effects, and those that have been extensively studied include Dentipellis, Gloeostereum, Hericium, Hydnellum, Hydnum, Irpex, Mycoleptodonoides, Polyozellus, Sarcodon, Sparassis, and Thelephora. The genera related to the degradation of environmental pollutants and industrial applications include Crustoderma, Cystostereum, Peniophora, Phlebia, Porostereum, and especially Phanerochaete, which has been extensively studied as a model for studying the degradation of environmental pollutants. The genera related to agriculture and biological control include Boidinia, Cymatoderma, Lopharia, Phlebia, Phlebiopsis, Pseudolagarobasidium, Sebacina, etc. However, many genera, including Aleurodiscus, Chondrostereum, Climacodon, Coniophora, Hymenochaete, Irpex, Resinicium, Steccherinum, and Stereum, exhibit multiple activities and potential applications in medicine, the environment, and industry.
Taxonomy
-
Botryobasidium naviculisporum L.J. Zhou & H.S. Yuan, sp. nov. Figs 3, 4
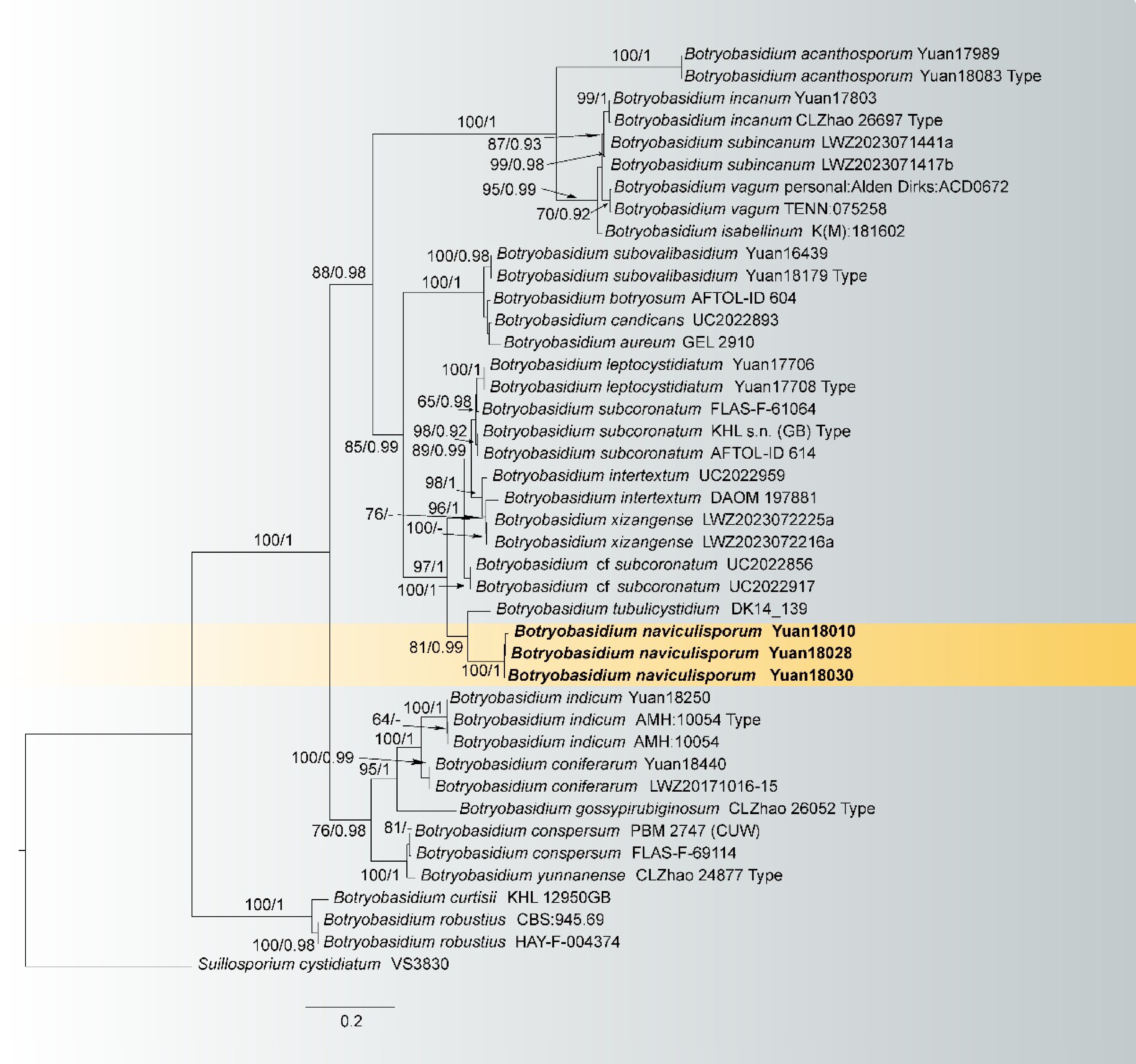
Figure 3.
Maximum likelihood tree illustrating the phylogeny of Botryobasidium naviculisporum related taxa based on ITS + LSU nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).

Figure 4.
Botryobasidium naviculisporum (holotype Yuan 18030). (a) A basidiocarp. Microscopic structures: (b) section through basidiocarp; (c) Basidiospores; (d) Basidioles; (e) Basidia; (f) Cystidia.
Fungal Names number: FN 572327
Etymology: Naviculisporum (Lat.), referring to the naviculiform basidiospores.
Holotype: China, Yunnan Province, Diqing Prefecture, Baimaxueshan National Nature Reserve, 28°20'4.56" N, 99°9'49.9" E, Alt. 4,125 m, on stump of Picea, 13 August, 2023, Yuan 18030 (IFP 019989, ITS: PQ878083, LSU: PV700909).
Diagnosis: Basidiocarps annual, resupinate, pellicular, smooth; hyphal system monomitic, generative hyphae thin-walled, with clamp connections; cystidia present; basidiospores navicular, foliaceous to fusiform, thick-walled, smooth, 12–16 × 6–8 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, adnate, pellicular, effuse, fluffy, adherent to the substrate and separable, ca. 100 µm thick. Hymenophoral surface smooth, yellowish (1A2–4A2) to pale yellow (1A3–4A3) when fresh, and unchanged when drying; margin not differentiated, distinct.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thin- to thick-walled, colorless; cyanophilous, inamyloid, tissues dark yellow in KOH.
Subiculum: subicular hyphae thick-walled, smooth, thick-walled, long-celled, vertical at the branch, irregularly and loosely interwoven, 7–9 μm in diam; cyanophilous, inamyloid. Subhymenial hyphae thin- to slightly thick-walled, frequently branched, irregularly interwoven, 7–11 μm in diam; cyanophilous, inamyloid.
Cystidia: cylindrical to clavate, thin-walled, smooth, colorless, the apex obtuse, inflated at the base, 26–55 × 6–9 µm; acyanophilous, inamyloid.
Basidia: cylindrical, thin-walled, smooth, colorless, with four sterigmata, clamped at the base, 14–24 × 6.5–10 µm; acyanophilous, inamyloid.
Basidiospores: navicular, foliaceous to fusiform, thick-walled, smooth, colorless, with oily contents, 12–16(–17) × (5.5–)6–8 µm, L = 14.2 µm, W = 6.85 µm, Q = 1.86–2.43 (n = 60/2); cyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimens examined: China. Yunnan Province, Diqing Prefecture, Baimaxueshan National Nature Reserve, 28°20'4.56" N, 99°9'49.9" E, Alt. 4,125 m, 13 August, 2023, on the stump of Picea, Yuan 18028 (IFP 019988, ITS: PQ878082); on the stump of Picea, Yuan 18010 (IFP 019987, ITS: PQ878081).
Notes: Botryobasidium naviculisporum and B. tubulicystidium clustered in a branch in the phylogenetic tree (Fig. 3). In morphology, the new species differs from B. naviculisporum by having smooth hymenophore, generative hyphae with clamped, distinctly bigger spores (12–16 × 6–8 µm vs 7.5–8.5 × 3–4 µm), and shorter cystidia (26–55 µm vs 60–110 µm)[2,57]. Moreover, B. naviculisporum is similar to B. leptocystidiatum in having greyish white to yellowish basidiocarps and clamped generative hyphae. However, the new species differs from the latter by having thick-walled and bigger basidiospores (12–16 × 6–8 µm vs 6.5–7.8 × 2.9–3.7 µm), shorter cystidia (26–55 µm vs 21.5–77 µm) and wider subhymenial hyphae (7–11 µm vs 4–7 μm)[58].
Ceraceomyces submicrosporus L.J. Zhou & H.S. Yuan, sp. nov. Figs 5, 6
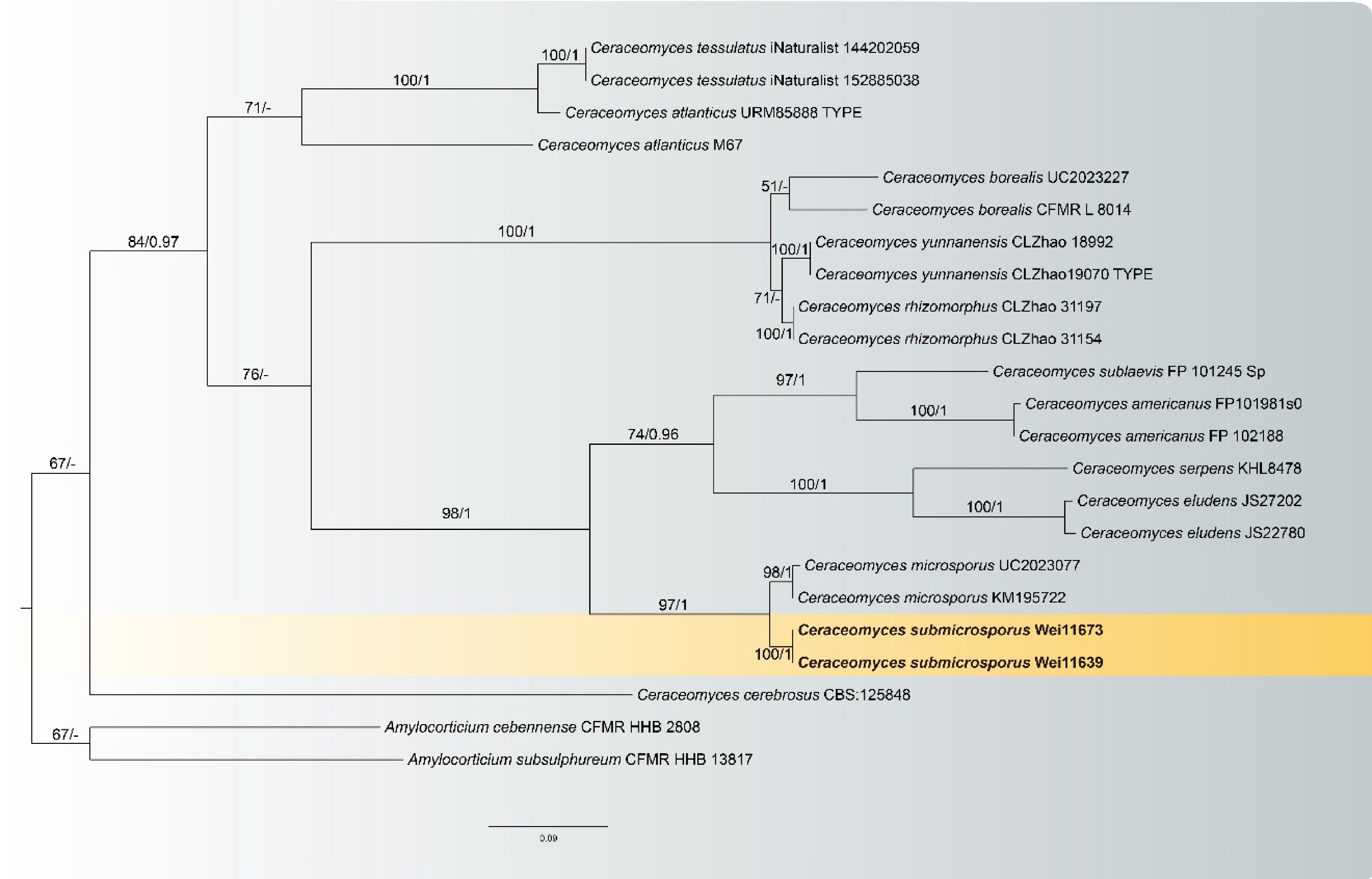
Figure 5.
Maximum likelihood tree illustrating the phylogeny of Ceraceomyces submicrosporus related taxa based on ITS + LSU nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).
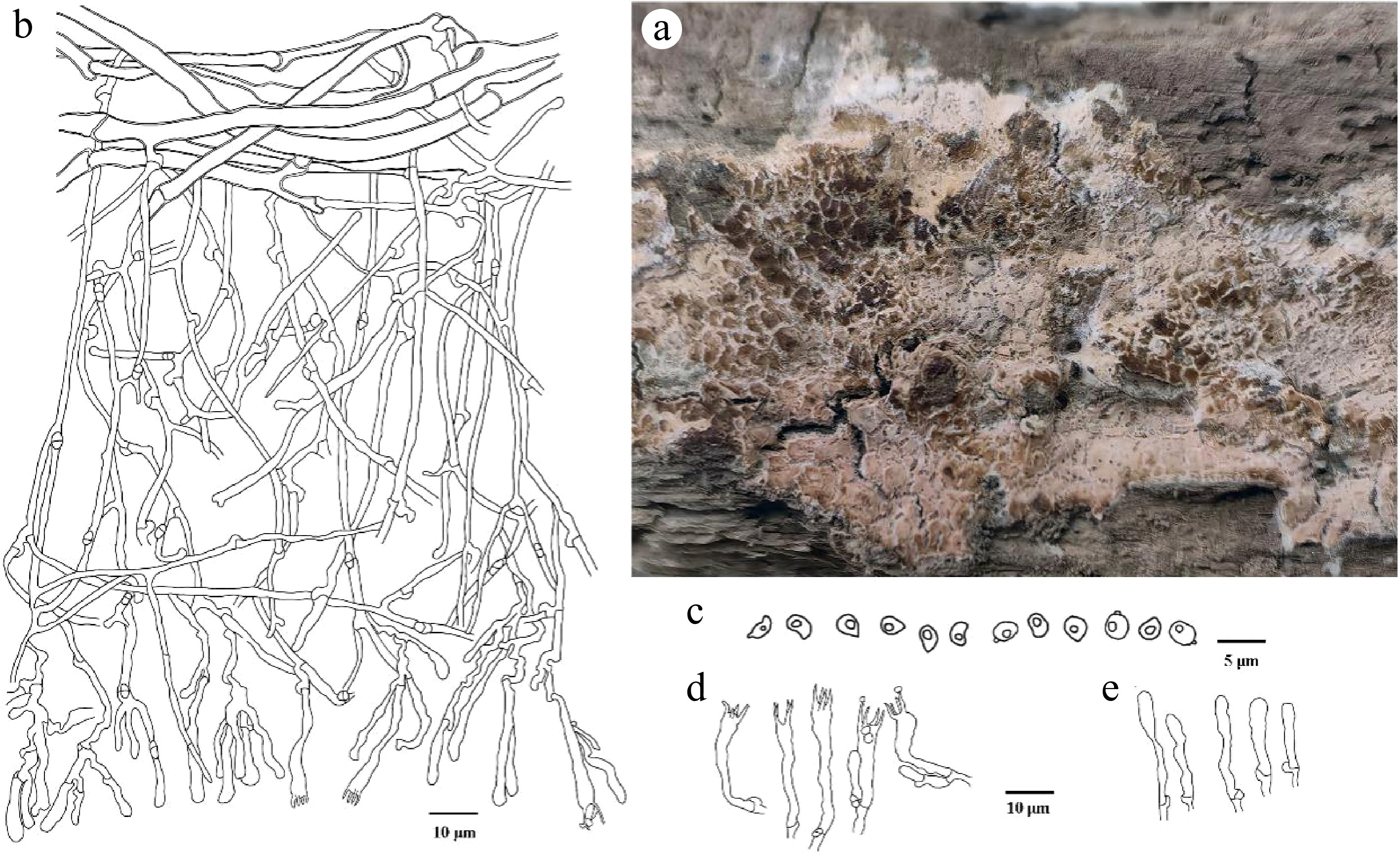
Figure 6.
Ceraceomyces submicrosporus (holotype Wei 11639). (a) Basidiocarp. Microscopic structures: (b) section through basidiocarp; (c) Basidiospores; (d) Basidia; (e) Basidioles.
Fungal Names number: FN 572326
Etymology: Submicrosporus (Lat.): refers to the similarity to C. microsporus.
Holotype: China, Zhejiang Province, Lishui City, Qingyuan County, Baishanzu National Nature Reserve, 27°45'33.38" N, 119°12'52.75" E, Alt. 1,458 m, on fallen branch of Pinus, 20 July, 2023, Wei 11639 (IFP 019985, ITS: PQ878079, LSU: PV700907).
Diagnosis: Basidiocarps annual, resupinate, smooth to tuberculate, cracked; hyphal system monomitic, generative hyphae thin-walled, frequently branched, with clamp connections; cystidia absent; basidiospores reniform, ovoid to globose, thin-walled, smooth, 2–3 × 1.5–2 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, separable, smooth, tuberculate, and cracked when fresh, cracked and slightly fragrant odor when drying, up to 15 cm long, 3 cm wide, 200 µm thick. Hymenophoral surface smooth, wax-like, beige (4C3) to orange red (8A6–8A8) when fresh, reddish white (7A2–13A2) to orange red (8A6–8A8) when drying. Margin sterile white.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thin- to slightly thick-walled; cyanophilous, inamyloid, tissues reddish brown in KOH.
Subiculum: subicular hyphae slightly thick-walled, smooth, colorless, moderately branched, irregularly and loosely interwoven, 2–5 μm in diam; cyanophilous, inamyloid. Subhymenial hyphae thin-walled, smooth, colorless, frequently branched, more or less flexuous, long-celled, unequal septate, irregularly interwoven, 1–2 μm in diam; acyanophilous, inamyloid.
Cystidia: absent.
Basidia: narrowly clavate to clavate, thin-walled, smooth, colorless, with four sterigmata, sterigmata up to 6 µm long, clamped at the base, 17–28 × 3–4 µm; acyanophilous, inamyloid.
Basidiospores: reniform, ovoid to globose, thin-walled, smooth, colorless, with oily contents, 2–3 × 1.5–2 µm, L = 2.4 µm, W = 2.0 µm, Q = 1.0–1.46 (n = 60/2); acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China. Zhejiang Province, Lishui City, Qingyuan County, Baishanzu National Nature Reserve, 27°45'33.38" N, 119°12'52.75" E, Alt. 1,458 m, on fallen branch of Pinus, 20 July 2023, Wei 11673 (IFP 019986, ITS: PQ878080, LSU: PV700908).
Notes: Ceraceomyces submicrosporus and Ce. microsporus clustered in a branch in the phylogenetic tree (Fig. 5). In morphology, they share nearly orange basidiocarps, subhymenial hyphae of similar wide (ca. 2 µm), and the spores with oily contents. However, the new species differs from Ce. microsporus by having distinctly shorter basidia (17–28 µm vs 22–42 µm), smaller spores (2–3 µm long vs 2.7–3.7 µm), and wider subicular hyphae (2–5 µm vs 2.2 µm)[59]. Moreover, Ce. submicrosporus is similar to Ce. bizonatus in having cracked basidiocarps when drying, similarly basidia (17–28 × 3–4 µm vs 16.5–24 × 3.1–5.5 µm), and spores (2–3 ×1.5–2 µm vs 2.5–3.3 × 1.8–2.2 µm). However, Ce. bizonatus differs from Ce. submicrosporus in having yellowish-brown basidiocarps and encrusted subhymenium hyphae[60].
Cylindrobasidium bambusinum C.L. Zhao, sp. nov. Figs 7, 8
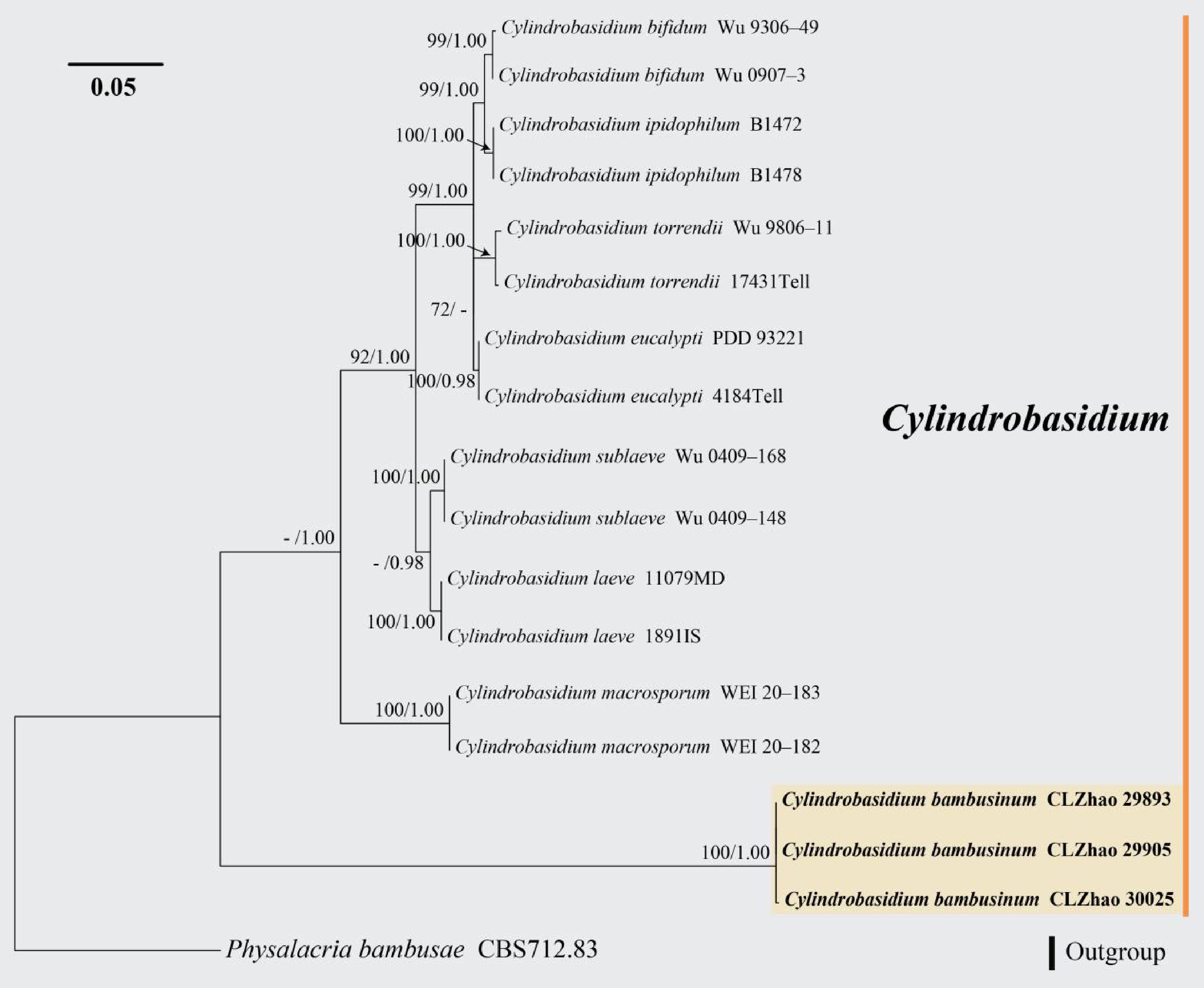
Figure 7.
Maximum likelihood tree illustrating the phylogeny of Cylindrobasidium bambusinum related taxa based on ITS nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).

Figure 8.
Cylindrobasidium bambusinum (holotype CLZhao 29905). (a), (b) Basidiocarp. Microscopic structures: (c) Basidiospores; (d) Basidia and basidioles; (e) Cystidioles; (f) Section through basidiocarp. Scale bars: (a) = 1 cm; (b) = 1 mm; (c)–(f) = 10 µm.
Index Fungorum number: IF 859190
Etymology: Bambusinum (Lat.): referring to growing on bamboo of the type specimen.
Holotype: China, Yunnan Province, Dehong Dai and Jingpo autonomous prefecture, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°12" N, 97°33" E, Alt. 3,404.6 m, on the dead bamboo, 18 July, 2023, CLZhao 29905 (SWFC00029905, ITS: PV705576, LSU: PV705584).
Diagnosis: Basidiocarps annual, resupinate, closely adnate, farinaceous; hyphal system monomitic, generative hyphae thick-walled, occasionally branched, with clamp connections; cystidia present; basidiospores drop-like to broadly ellipsoid, thin-walled, smooth, 4.5–6 × 3–4 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, thin, closely adnate, without odor or taste when fresh, up to 6 cm long, 1.3 cm wide, and 120 µm thick. Hymenophoral surface farinaceous, white (–A1) when fresh and unchanged upon drying. Sterile margin narrow, white, up to 1 mm.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thick-walled, colorless, occasionally branched, interwoven, 3.4–3.6 µm in diam; cyanophilous, inamyloid, tissues unchanged in KOH.
Cystidia: subclavate, thin-walled, smooth, colorless, abundant, 22–24 × 4–5.5 µm; acyanophilous, inamyloid.
Basidia: cylindrical to subcylindrical, thin-walled, smooth, colorless, with four sterigmata, clamped at the base, 12.5–15.5 × 3.5–5.5 µm; basidioles in shape similar to basidia, but slightly smaller; acyanophilous, inamyloid.
Basidiospores: drop-shaped to broadly ellipsoid, thin-walled, smooth, colorless, with several guttulae, (4–)4.5–6 × 3–4 µm, L = 5.17 µm, W = 3.34 µm, Q = 1.48–1.54 (n = 60/2); acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimens examined: China. Yunnan Province, Dehong Dai and Jingpo Autonomous Prefecture, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°12" N, 97°33" E, Alt. 3,404.6 m, on the dead bamboo, 18 July, 2023, CLZhao 29893 (SWFC00029893, ITS: PV705575, LSU: PV705583), CLZhao 30025 (SWFC00030025, ITS: PV705577, LSU: PV705585).
Notes: Morphologically, Cylindrobasidium bambusinum resembles Cy. bifidum and Cy. macrosporum sharing the clamped generative hyphae. However, Cy. bifidum can be distinguished from Cy. bambusinum by having cream hymenial surface and pyriform basidiospores; Cy. macrosporum differs from Cy. bambusinum by hacing smooth to tuberculate hymenial surface, longer basidia (25–30 × 5–6 µm vs 12.5–15.5 × 3.5–5.5 µm), and larger basidiospores (10–11 × 4.7–5.2 µm)[61].
Hyphoderma nitidum X.J. Zhang & C.L. Zhao, sp. nov. Figs 9, 10
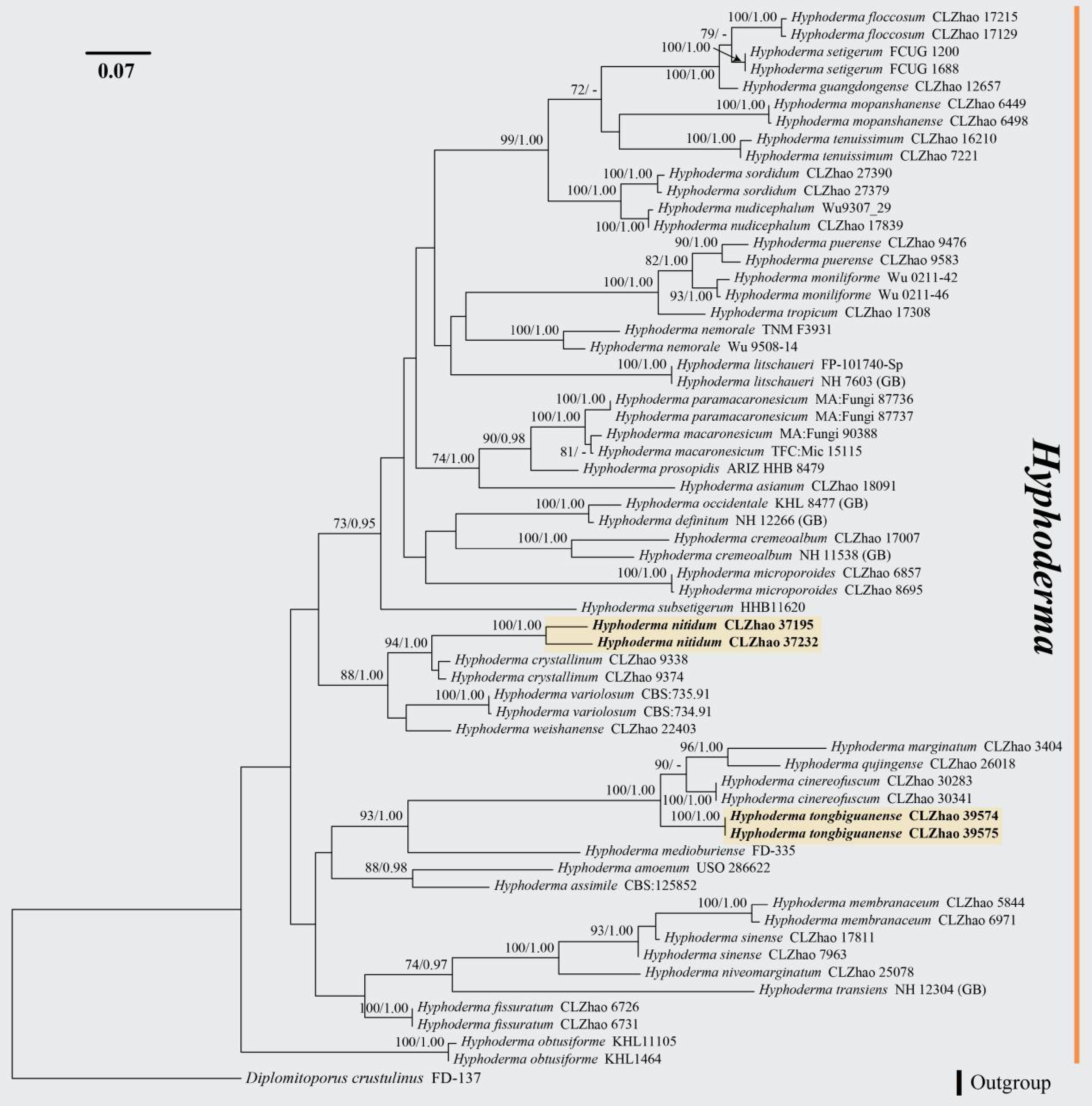
Figure 9.
Maximum likelihood tree illustrating the phylogeny of Hyphoderma nitidum and H. tongbiguanense related taxa based on ITS nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).

Figure 10.
Hyphoderma nitidum (holotype CLZhao 37195). (a), (b) Basidiocarp. Microscopic structures: (c) Basidiospores; (d) Basidia and basidioles; (e) Tubular cystidia; (f) Pseudocystidia; (g) Section through basidiocarp. Scale bars: (a) = 1 cm; (b) = 1 mm; (c)–(g) = 10 µm.
Index Fungorum number: IF 859193
Etymology: Nitidum (Lat.): refers to the glossy hymenophore of the type specimen.
Holotype: China, Yunnan Province, Dehong Dai and Jingpo autonomous prefecture, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°28" N, 97°31" E, Alt. 1,125 m, on the fallen angiosperm branch, 02 July, 2024, CLZhao 37195 (SWFC00037195, ITS: PV470533).
Diagnosis: Basidiocarps annual, resupinate, farinaceous to membranous, smooth; hyphal system monomitic, generative hyphae thin-walled, with clamp connections; cystidia present; basidiospores cylindrical, thin-walled, smooth, 10–13 × 3–4.5 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, farinaceous when fresh, turn to membranous upon drying, without odor or taste, up to 6.5 cm long, 2 cm wide, 50–150 µm thick. Hymenophoral surface smooth, white (–A1) when fresh, white (–A1) to yellowish white (1A2–4A2) upon drying. Sterile margin white, up to 1 mm.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thin-walled, colorless, branched, interwoven, 2.8–4.3 µm in diam; acyanophilous, inamyloid, tissues unchanged in KOH.
Cystidia of two types: (1) pseudocystidia thin-walled, colorless, larger, with presence of abundant encrustations, 30–32.4 × 4.4–5.4 µm; (2) tubular cystidia, thin-walled, colorless, 34.5–42 × 6.2–8.8 µm, cystidioles absent; acyanophilous, inamyloid.
Basidia: clavate, thin-walled, smooth, colorless, with 4 sterigmata, clamped at the base, with oil drops, 16.5–18 × 4.5–6.4 µm; basidioles dominant, similar to basidia in shape, but slightly smaller; acyanophilous, inamyloid.
Basidiospores: cylindrical, thin-walled, smooth, colorless, with oil drops, (9–)10–13(–13.5) × (2.8–)3–4.5(–5) µm, L = 11.36 µm, W = 3.62 µm, Q = 3.14 (n = 60/2); acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China. Yunnan Province, Dehong Dai and Jingpo autonomous prefecture, Yingjiang County, Tongbiguan Provincial Nature Reserve, 24°28" N, 97°31" E, Alt. 1,125 m, on the fallen angiosperm branch, 02 July, 2024, CLZhao 37232 (SWFC00037232, ITS: PV470534).
Notes: Morphologically, Hyphoderma nitidum is similar to H. cinereofuscum and H. guangdongense in having cylindrical basidiospores. However, H. cinereofuscum is distinguished from H. nitidum by having cinereous and fuscous hymenial surface and longer basidia (23.5–26 × 5.3–6.5 µm vs 16.5–18 × 4.5–6.4 µm)[62]. H. guangdongense differs from H. nitidum by having longer tubular cystidia (55–63 × 6.5–10.3 µm vs 34.5–42 × 6.2–8.8 µm), and shorter basidiospores (7.4–9.0 × 3.2–4.0 µm vs 10–13 × 3–4.5 µm)[63].
Hyphoderma tongbiguanense X.J. Zhang & C.L. Zhao, sp. nov. Figs 9, 11

Figure 11.
Hyphoderma tongbiguanense (holotype CLZhao 39575). (a), (b) Basidiocarp. Microscopic structures: (c) Basidiospores; (d) Basidia and basidioles; (e) Section through basidiocarp. Scale bars: (a) = 1 cm; (b) = 1 mm; (c)–(e) = 10 µm.
Index Fungorum number: IF 859194
Etymology: Tongbiguanense (Lat.): refers to the locality (Tongbiguan Provincial Nature Reserve) of the type specimen.
Holotype: China, Yunnan Province, Dehong Dai and Jingpo autonomous prefecture, Mang city, Tongbiguan Provincial Nature Reserve, 24°28" N, 97°31" E, Alt. 1,125 m, on the dead bamboo, 08 July, 2024, CLZhao 39575 (SWFC00039575, ITS: PV470532).
Diagnosis: Basidiocarps annual, resupinate, coriaceous, smooth; hyphal system monomitic, generative hyphae thin-walled, frequently branched, with simple septate; cystidia absent; basidiospores ellipsoid, thin-walled, smooth, 8–11 × 4–6 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, coriaceous, without odor or taste, up to 10 cm long, 3 cm wide, 50–150 µm thick. Hymenophoral surface smooth, white (–A1) when fresh, white (–A1) to pale yellow (1A3–4A3) upon drying. Sterile margin white, up to 1 mm.
Hyphal structure: hyphal system monomitic; generative hyphae with simple septate, colorless, thin-walled, frequently branched, interwoven, 3.6–5.2 µm in diam; acyanophilous, inamyloid, tissues unchanged in KOH.
Cystidia: absent.
Basidia: clavata, thin-walled, smooth, colorless, with four sterigmata, simple septate at the base, with oil drops, 17.9–33.6 × 7.9–11.3 µm; basidioles dominant, similar to basidia in shape, but slightly smaller; acyanophilous, inamyloid.
Basidiospores: ellipsoid, thin-walled, smooth, colorless, with oil drops, (7.5–)8–11 × 4–6(–6.5) µm, L = 9.42 µm, W = 5.50 µm, Q = 1.4–1.71 (n = 60/2); acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China. Yunnan Province, Dehong Dai and Jingpo autonomous prefecture, Mang city, Tongbiguan Provincial Nature Reserve, 24°28" N, 97°31" E, Alt. 1,125 m., on the dead bamboo, 08 July, 2024, CLZhao 39574 (SWFC00039574, ITS: PV470531).
Notes: Morphologically, Hyphoderma tongbiguanense is similar to H. puerense and H. tropicum in having ellipsoid basidiospores. However, H. puerense can be distinguished from H. tongbiguanense by having a cream to slightly buff hymenial surface and shorter basidiospores (6–7.5 × 3–4.5 µm vs 8–11 × 4–6 µm)[64]. H. tropicum differs from H. tongbiguanense by having clamped generative hyphae and narrower basidia (15.5–20 × 3.5–4.5 µm vs 17.9–33.6 × 7.91–11.3 µm)[65].
Schizocorticium microsporum L.J. Zhou & H.S. Yuan, sp. nov. Figs 12, 13
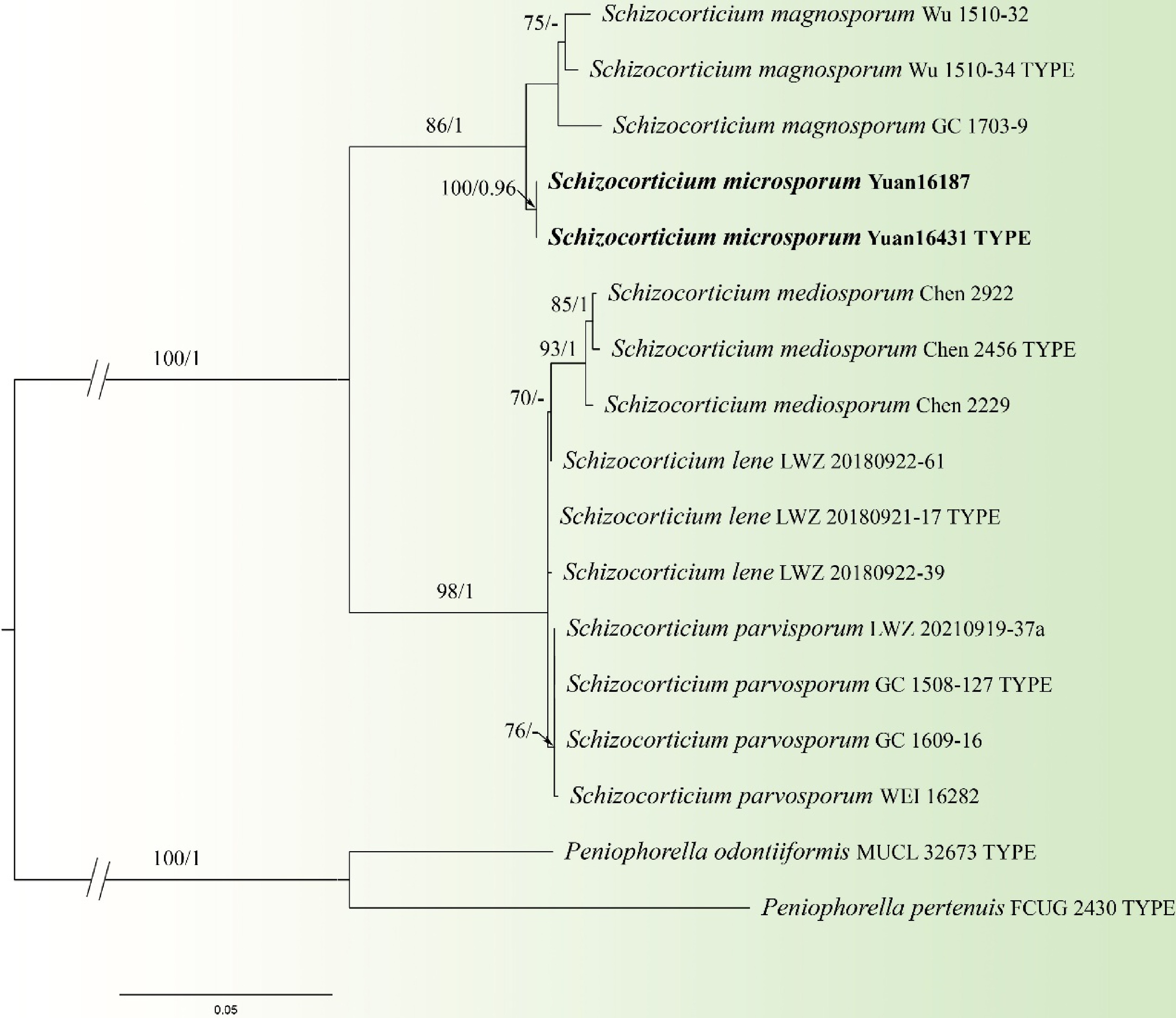
Figure 12.
Maximum likelihood tree illustrating the phylogeny of Schizocorticium microsporum related taxa based on ITS + LSU nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).
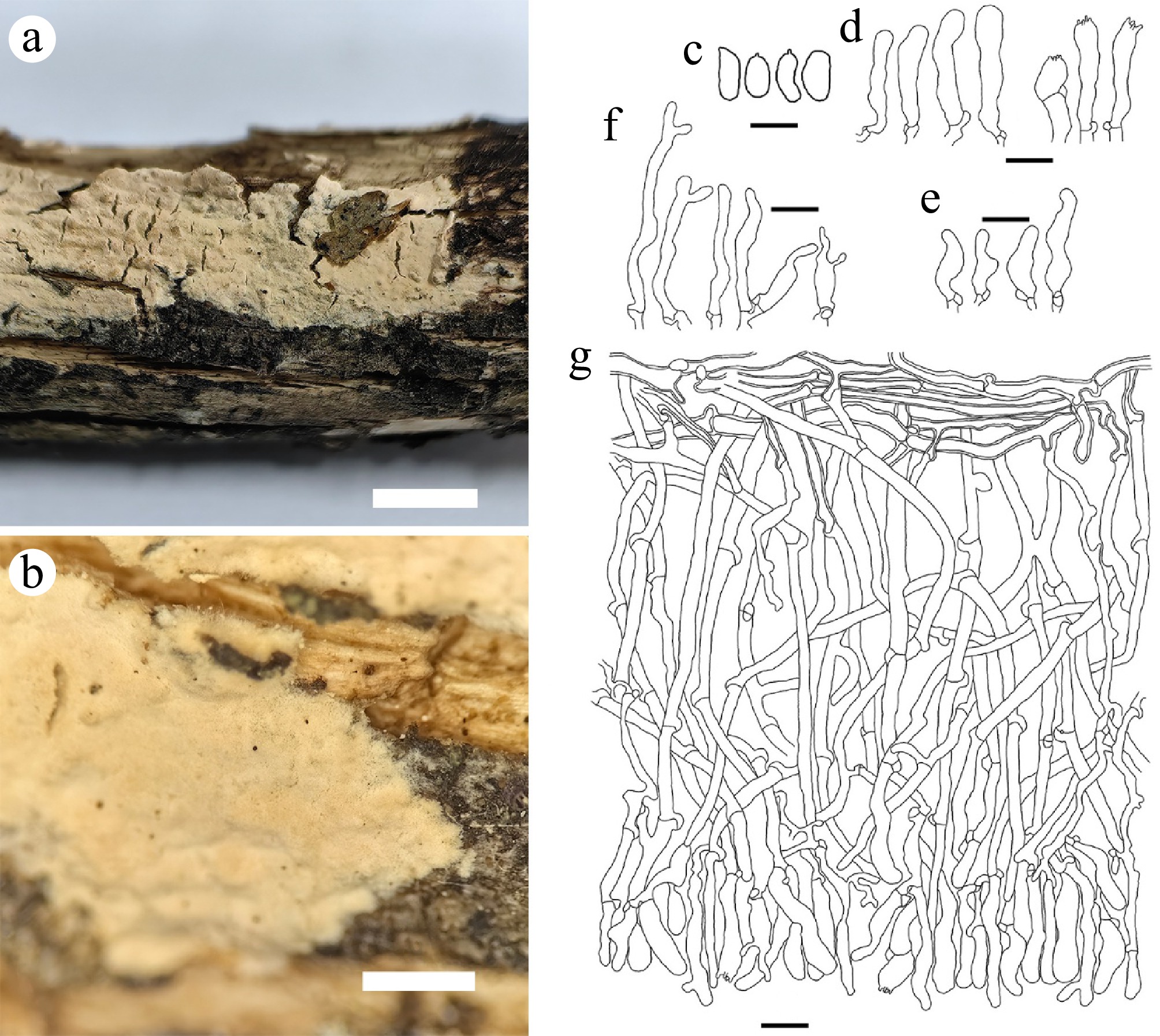
Figure 13.
Schizocorticium microsporum (holotype Yuan 16431). (a) Basidiocarp. Microscopic structures: (b) Section through basidiocarp; (c) Basidiospores; (d) Basidioles; (e) Basidia; (f) Cystidia. Scale bars: (a) = 1 cm; (b) = 1 mm; (c) = 5 µm; (d)–(g) = 10 µm.
Fungal Names number: FN 572729
Etymology: Microsporum (Lat.), referring to having smaller basidiospores.
Holotype: China, Xizang Autonomous Region, Linzhi City, Bomi County, 29°46'58.53" N, 95°42'0.94" E, Alt. 3,700 m, on fallen angiosperm branch, 26 October, 2021, Yuan 16431 (IFP 020068, ITS: PV702216, LSU: PV759768).
Diagnosis: Basidiocarps annual, resupinate, adnate, membranaceous to coriaceous, smooth to tuberculate; hyphal system monomitic, generative hyphae thin- to slightly thick-walled, moderately branched, with clamp connections; cystidia present; basidiospores narrowly ellipsoid to ellipsoid, thin-walled, smooth, 4.5–6 × 2–3 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, adnate, effused, membranaceous to coriaceous, more or less cracked, adherent to the substrate and uneasily separable, 100–200 μm thick. Hymenophoral surface smooth to tuberculate, orange white (5A2–6A2) to reddish white (7A2–13A2) when drying; sterile margin indistinct.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thin- to thick-walled; cyanophilous, inamyloid, tissues unchanged in KOH.
Subiculum: subicular hyphae thick-walled, smooth, colorless, moderately branched, parallel interwoven, sometimes flexuous, 2–4 μm in diam; cyanophilous, inamyloid. Subhymenial hyphae thin- to slightly thick-walled, smooth, colorless, moderately branched, sometimes flexuous, irregularly interwoven, 2–3.5 μm in diam; cyanophilous, inamyloid.
Cystidia: subfusiform to subulate, thin-walled, smooth, colorless, with an obtuse apex, clamped at the base, 15–33 × 4–6 μm; acyanophilous, inamyloid.
Hyphidia: cylindrical, thin-walled, smooth, colorless, occasionally branched at the apex and similar dendrohyphidia, abundant, 17–51 × 2–5 μm; acyanophilous, inamyloid.
Basidia: clavate, thin-walled, smooth, colorless, with four sterigmata, clamped at the base, sometimes flexuous, 9–30 × 4–6 µm; acyanophilous, inamyloid.
Basidiospores: narrowly ellipsoid to ellipsoid, thin-walled, smooth, colorless, (4–)4.5–6(–7) × 2.0–3.0 µm, L = 5.20 µm, W = 2.55 µm, Q = 1.67–2.86; acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China, Xizang Autonomous Region, Linzhi City, Milin County, Nanyigou Park, 29°4'3.64" N, 94°13'47.75" E, Alt. 3,250 m, on fallen angiosperm branch, 22 October, 2021, Yuan 16187 (IFP 020069, ITS: PV702215, LSU: PV759767).
Notes: Schizocorticium microsporum and S. magnosporum clustered in a branch in the phylogenetic tree (Fig. 12). In morphology, they share cracked basidiocarps, ellipsoid basidiospores, and hyphae of similar width (ca. 2–4 μm). However, the new species differs from S. magnosporum by having subulate cystidia, distinctly smaller basidia (9–30 × 4–6 µm vs 27–45 × 7–8.5 µm), smaller basidiospores (4.5–6 × 2.0–3.0 µm vs 9.8–11.5 × 4.2–5.5 µm), and longer hyphidia (17–51 µm vs 20–45 µm)[66]. Moreover, S. microsporum differs from S. lene by having hyphidia, subulate cystidia, and smaller basidiospores (4.5–6 × 2.0–3.0 µm vs 4.8–5.8 ×2.9–3.5 µm)[67]. S. microsporum differs from S. mediosporum, S. parvisporum by having cystidia and smaller basidiospores (4.5–6 × 2.0–3.0 µm vs 8–9.8 × 3.8–4.8 µm, 6–7.5 × 3.2–3.8 µm), and longer hyphidia (17–51 µm vs 22–47 µm, 20–45 µm)[66].
Thelephora chawalongensis H.S. Yuan & Y.Q. Zhu, sp. nov. Figs 14, 15
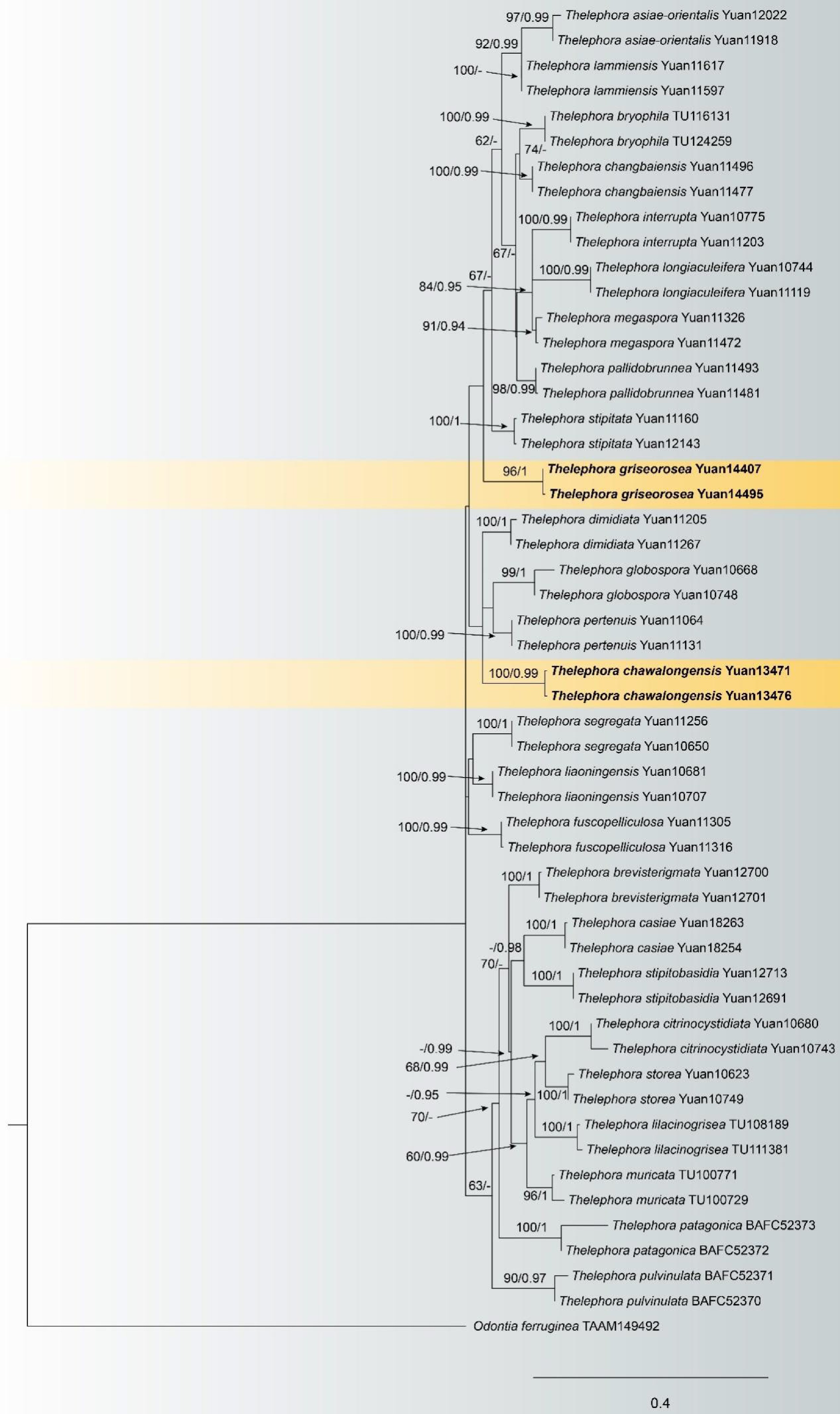
Figure 14.
Maximum likelihood tree illustrating the phylogeny of Thelephora chawalongensis and T. griseorosea related taxa based on ITS + LSU nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).
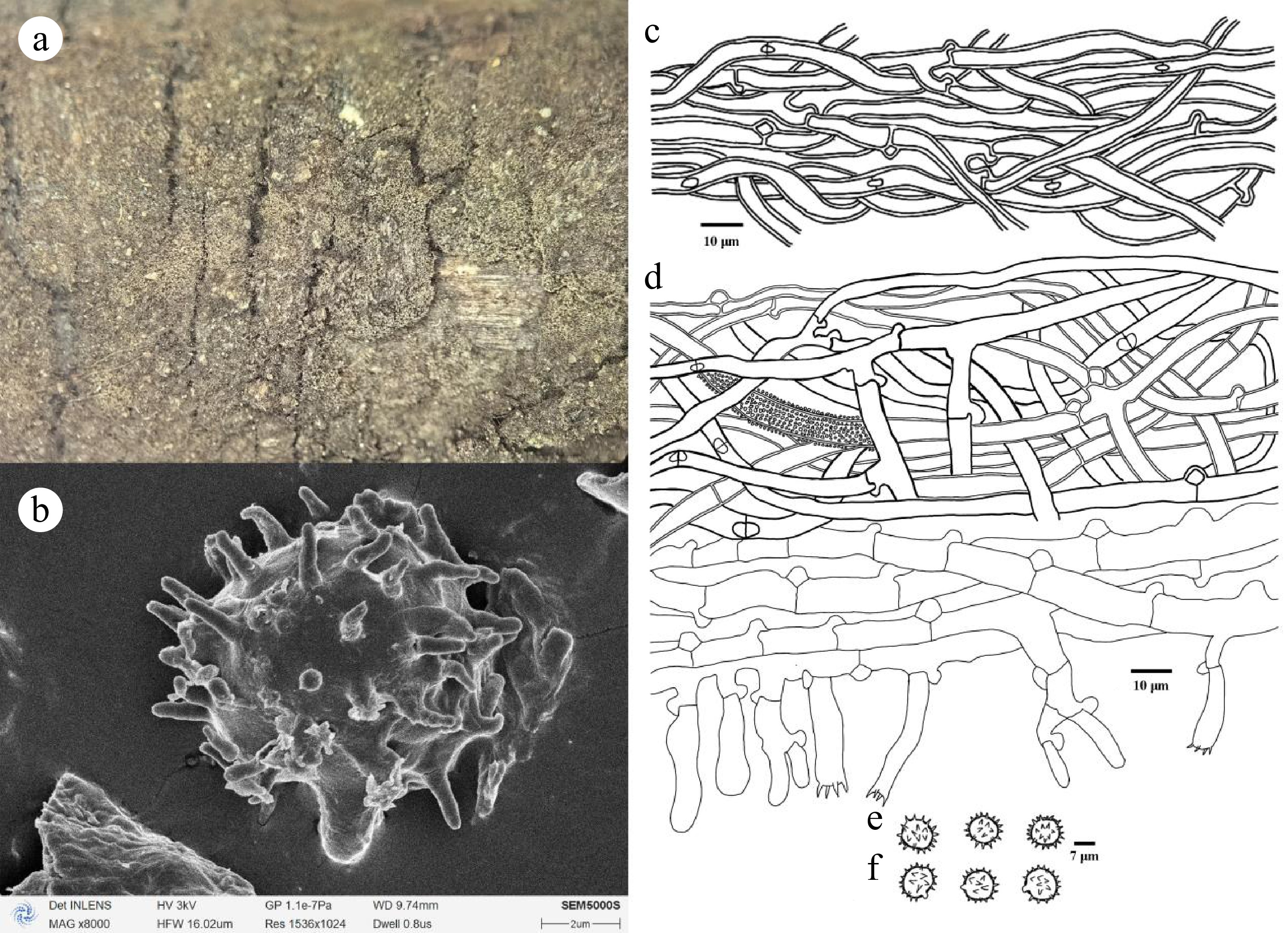
Figure 15.
Thelephora chawalongensis (holotype Yuan 13476). (a) Basidiocarp; (b) SEM of basidiospores. Microscopic structures: (c) Hyphae from a rhizomorph; (d) Section through basidiocarp; (e) Basidiospores in frontal view; (f) Basidiospores in lateral view.
Fungal Names number: FN 572323
Etymology: Chawalongensis (Lat.), referring to the location where the type specimen was collected.
Holotype: China, Xizang Autonomous Region, Linzhi City, Chayu County, Chawalong, 28°35'23" N, 98°20'36" E, Alt. 2,000 m, on fallen angiosperm branch, 13 July, 2019, Yuan 13476 (IFP 019981, ITS: PQ656822, LSU: PQ878075).
Diagnosis: Basidiocarps annual, resupinate, arachnoid, continuous, smooth; hyphal system monomitic, generative hyphae thin-walled, frequently branched, with clamps and rarely simple septate; cystidia absent; basidiospores subglobose to globose, slightly thick-walled, echinulate to aculeate, 8.2–10 × 7.5–9.2 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, adherent to the substrate, arachnoid, without odor or taste when fresh, 0.1–0.2 mm thick, continuous. Hymenophoral surface smooth, pale brown to brown (6E4–6F4) and concolorous with the subiculum. Sterile margin often indeterminate, arachnoid, concolorous with hymenophore.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps and rarely simple septate, slightly thick- to thick-walled; cyanophilous, inamyloid, pale brown in KOH and distilled water.
Rhizomorphs: present in subiculum and margins, 50–80 mm in diam; rhizomorphic surface more or less smooth, hyphae in rhizomorph monomitic, undifferentiated, of type A (according to Agerer, 1987–2008), loosely interwoven and of uniform single hyphae clamped, thick-walled, occasionally branched, 3–7 µm in diam, without encrustation, pale brown in KOH, cyanophilous, inamyloid.
Subiculum: subicular hyphae slightly thick- to thick-walled, encrustation, with clamped and rarely simple septate, frequently branched, 4–7 µm in diam; cyanophilous, inamyloid. Subhymenial hyphae thin-walled, without encrustation, cells short and inflated, 5–10 µm in diam; acyanophilous, inamyloid.
Cystidia: absent.
Basidia: clavate, thin-walled, smooth, colorless, not stalked, sinuous, without transverse septa, with four sterigmata, sterigmata 5–8 µm long, clamped at the base, 15–40 × 5–10 µm; acyanophilous, inamyloid.
Basidiospores: subglobose to globose in frontal and later face, slightly thick-walled, echinulate to aculeate, echinuli usually isolated, up to 2 µm long, (7.8–)8.2–10(–10.5) × (6.9–)7.5–9.2(–9.6) µm, L = 9 µm, W = 8.26 µm, Q = 1.01–1.31 (n = 60/2); cyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China. Xizang Autonomous Region, Linzhi City, Chayu County, Chawalong, 28°35'23" N, 98°20'36" E, Alt. 2,000 m, on fallen branch of Quercus, 13 July, 2019, Yuan 13471 (IFP 019982, ITS: PQ656821, LSU: PQ878076).
Notes: In the phylogenetic tree (Fig. 14), Thelephora chawalongensis clustered with Th. dimidiata, Th. globospora and Th. pertenuis from China, and the basidiocarps adherent to the substrate are their common characteristics[68]. In addition, the rhizomorphic smooth surface and presence of rhizomorphs for Th. dimidiata are also similar to Th. chawalongensis, and the type of rhizomorphs are the opposite of Th. Chawalongensis[68]. Th. globospora can be distinguished from Th. chawalongensis by the dark brown basidiocarps, the absence of rhizomorphs, and larger basidiospores; Th. pertenuis can set it apart from Th. chawalongensis by the absence of the rhizomorphs, the smaller basidiospores, and larger basidia[68].
Thelephora griseorosea H.S. Yuan & Y.Q. Zhu, sp. nov. Figs 14, 16
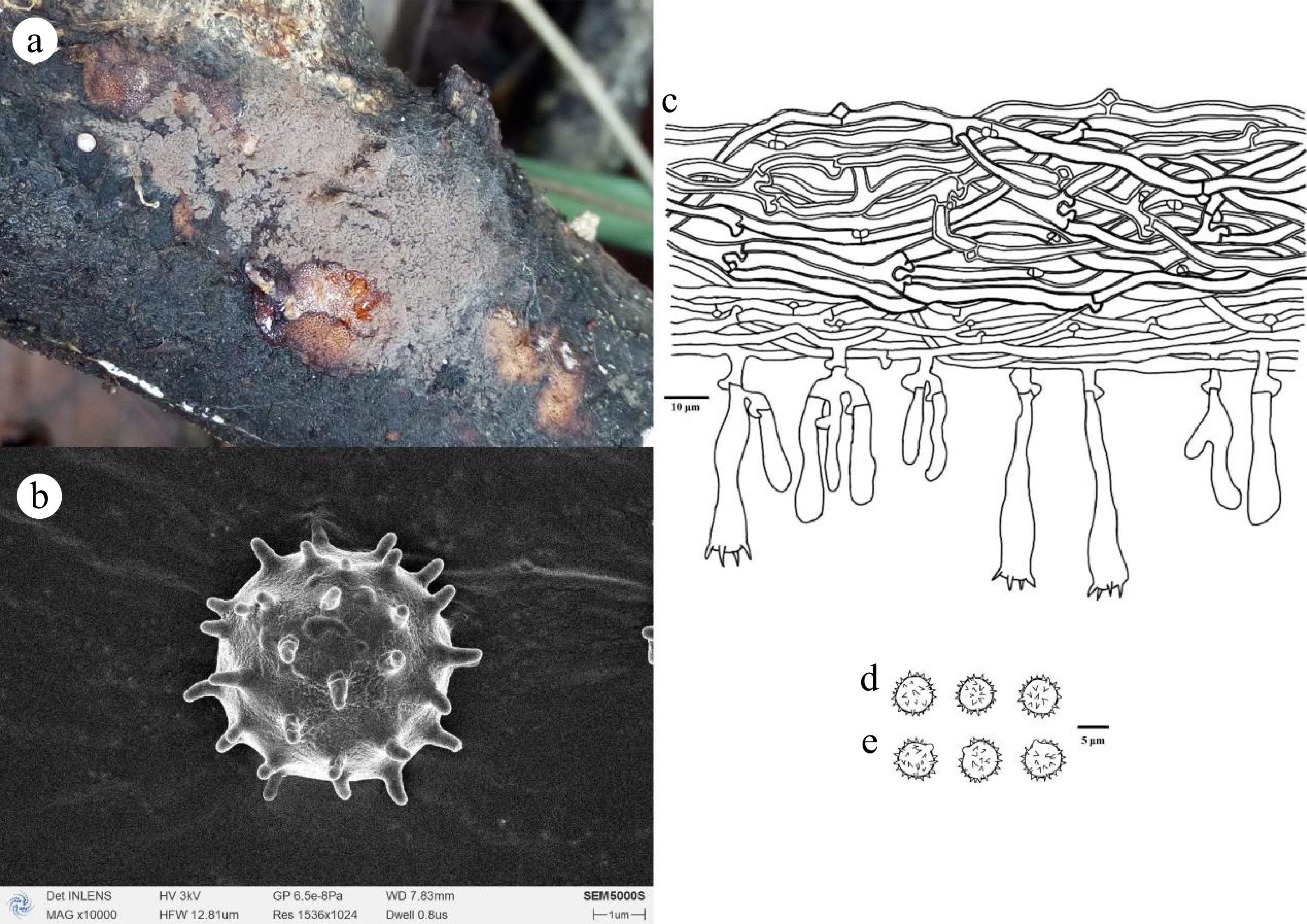
Figure 16.
Thelephora griseorosea (holotype Yuan 14495). (a) Basidiocarp; (b) SEM of basidiospores. Microscopic structures: (c) Section through basidiocarp; (d) Basidiospores in frontal view; (e) Basidiospores in lateral view.
Fungal Names number: FN 572324
Etymology: Griseorosea (Lat.), referring to the grey to pinkish grey hymenophoral surface.
Holotype: China, Yunnan Province, Chuxiong Yi Autonomous Prefecture, Wuding County, Shizi Mountain, 25°32'11" N, 102°22'24" E, Alt. 2,400 m, on rotten angiosperm debris, 27 September, 2019, Yuan 14495 (IFP 019983, ITS: PQ656824, LSU: PQ878078).
Diagnosis: Basidiocarps annual, resupinate, arachnoid, continuous, smooth; hyphal system monomitic, generative hyphae thin-walled, occasionally branched, with clamp connections; cystidia absent; basidiospores subglobose to globose, slightly thick-walled, echinulate, 5.2–7.2 × 4.8–6.2 µm, cyanophilous, inamyloid.
Basidiocarps: annual, resupinate, separable from the substrate, arachnoid, without odor or taste when fresh, 0.3–0.8 mm thick, continuous. Hymenophoral surface smooth, grey (–E1) to pinkish grey (11D3–12D3), and concolorous with the subiculum. Sterile margin often indeterminate, byssoid, whitish, paler than hymenophore.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, slightly thick- to thick-walled; cyanophilous, inamyloid, tissues pale brown in KOH.
Rhizomorphs: absent.
Subiculum: subicular hyphae slightly thick- to thick-walled, smooth, colorless, with clamped, 3–5 µm in diam; cyanophilous, inamyloid. Subhymenial hyphae thin-walled, colorless, occasionally branched, cells more or less uniform, 2–4 µm in diam; acyanophilous, inamyloid.
Cystidia: absent.
Basidia: utriform, thin-walled, smooth, colorless, not stalked, sinuous, without transverse septa, with four sterigmata, sterigmata up to 5 µm long, clamped at the base, 15–45 × 3–6 µm; acyanophilous, inamyloid.
Basidiospores: subglobose to globose in frontal and later face, slightly thick-walled, echinulate, echinuli usually isolated, up to 1 µm long, (4.9–)5.2–7.2(–7.5) × (4.2–)4.8–6.2(–6.5) µm, L = 6.18 µm, W = 5.32 µm, Q = 1.02–1.41 (n = 60/2); cyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Additional specimen examined: China, Yunnan Province, Dali Bai Autonomous Prefecture, Nanjian Yizu Autonomous County, Lingbao Mountain National Forest Park, 24°45'43" N, 100°30'10" E, Alt. 2,500 m, on stump of Pinus, 19 September, 2019, Yuan 14407 (IFP 019984, ITS: PQ656823, LSU: PQ878077).
Notes: In the phylogenetic tree (Fig. 14), two samples of Thelephora griseorosea formed an isolated clade, and clustered with the other nine species without support. In morphology, they share similar morphological characteristics: an indeterminate sterile margin and the absence of cystidia. However, Th. griseorosea possesses significant morphological differences in comparison with other species. For instance, Th. griseorosea can be distinctly differentiated from Th. stipitata by its arachnoid (vs mucedinoid) and grey (vs yellowish brown) basidiomes, narrower hyphal (2–4 µm vs 5–8 µm), smaller basidia (15–45 × 3–6 µm vs 20–50 × 6–9 µm), and smaller basidiospores (5.2–7.2 × 4.8–6.2 µm vs 7.5–8.5 × 6.5–8 µm)[68]. Th. griseorosea often has clamped hyphae and smaller basidiospores, which are distinctly different from Th. Lammiensis[69].
Tubulicrinis hongheensis C.L. Zhao, sp. nov. Figs 17, 18
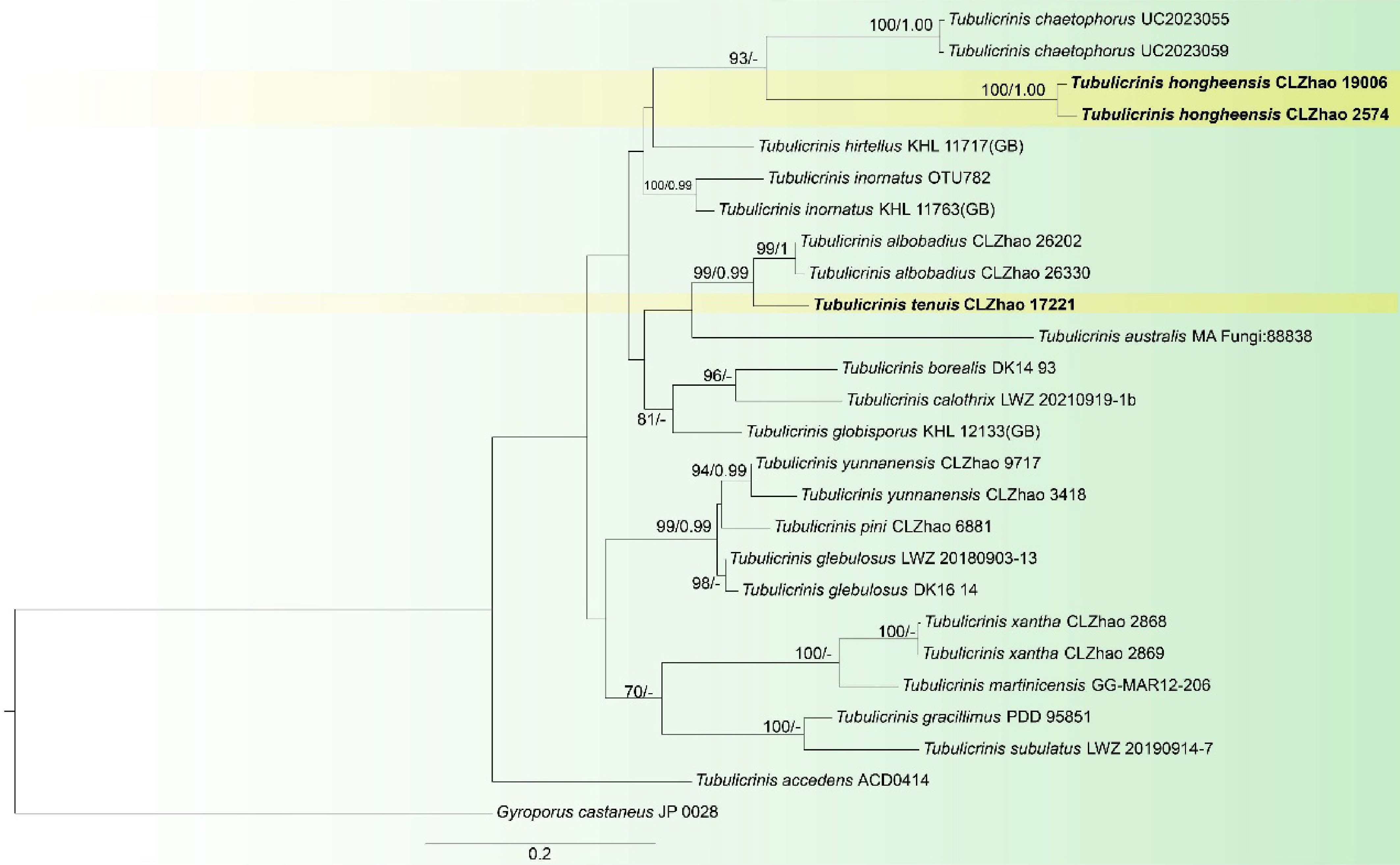
Figure 17.
Maximum likelihood tree illustrating the phylogeny of Tubulicrinis hongheensis and T. tenuis related taxa based on ITS nuclear rDNA sequences dataset. Branches are labeled with Maximum likelihood bootstrap equal to or higher than 60%, and Bayesian posterior probabilities equal to or higher than 0.9. Vouchers are indicated after the species names. New species in bold (black).
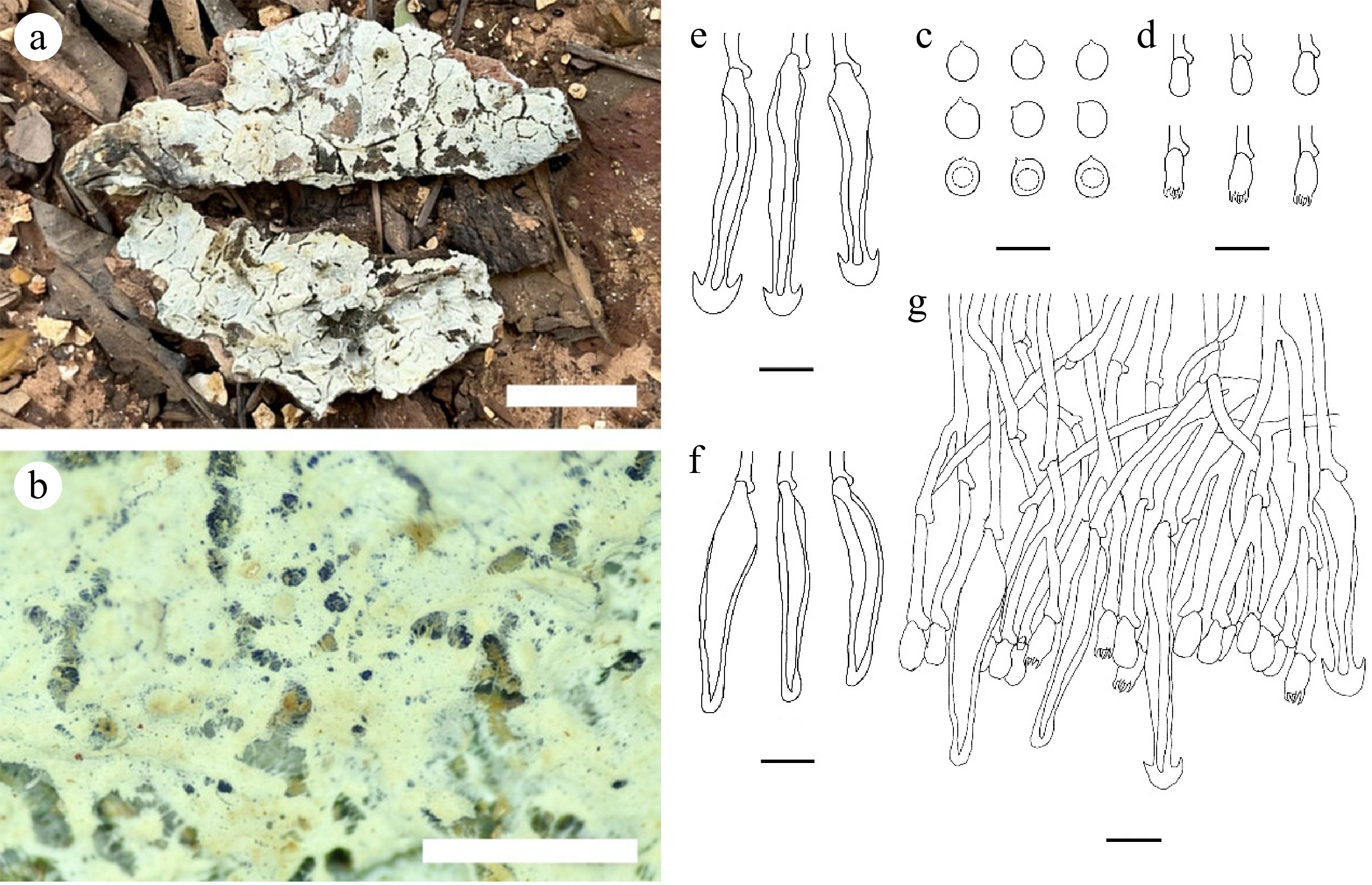
Figure 18.
Tubulicrinis hongheensis (holotype CLZhao 19006). (a), (b) Basidiocarp. Microscopic structures: (c) Basidiospores; (d) Basidia and basidioles; (e), (f) Cystidia; (g) Section through basidiocarp. Scale bars: (a) = 1 cm; (b) = 1 mm; (c) = 5 µm; (d)–(g) = 10 µm.
Index Fungorum number: IF 859192
Etymology: Hongheensis (Lat.): referring to the locality (Honghe Hani and Yi Autonomous Prefecture) of the type specimen.
Holotype: China, Yunnan Province, Honghe, Pingbian County, Daiweishan National Nature Reserve, 28°42" N, 114°11" E, Alt. 1,356 m, on fallen angiosperm trunk, 9 June, 2020, CLZhao 19006 (SWFC00019006, ITS: PV705579, LSU: PV705582).
Diagnosis: Basidiocarps annual, resupinate, coriaceous, cracked; hyphal system monomitic, generative hyphae thin- to thick-walled, occasionally branched, with clamp connections; cystidia present; basidiospores broadly ellipsoid, thin-walled, smooth, 3.6–4.1 × 3–3.5 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, adnate, coriaceous, without odor or taste when fresh, up to 2.6 cm long, 1.2 cm wide, and 50–70 µm thick. Hymenial surfaces cracking, white (–A1) to cream (4A3) when fresh, cream (4A3) to pale yellow (1A3–4A3) upon drying. Sterile margin thin, up to 0.1 mm wide.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, colorless, thin to thick-walled, occasionally branched, smooth, 2.0–4.0 µm in diam; cyanophilous, inamyloid, tissues unchanged in KOH.
Cystidia of two types: (1) Subfusiform cystidia, colorless, smooth, widest near the base, unevenly thick-walled with a narrow or wide lumen, gradually tapering, apically rounded to a insignificantly widened head, 33.5–71.5 × 5.5–10.5 µm; cyanophilous, inamyloid; (2) Cylindrical cystidia, smooth, unevenly thick-walled with a narrow or wide lumen, colorless, rooted at the base, gradually tapering, broadly rounded at the apex and covered by a large umbrella-shaped crystalline head, 32.0–85.5 × 3.5–11.0 µm; acyanophilous, inamyloid.
Basidia: clavate, thin-walled, smooth, colorless, occasionally sinuous, with four sterigmata and a clamp connection at the base, 7.5–14.5 × 4.0–7.0 µm; basidioles dominant, in shape similar to basidia, but slightly smaller; acyanophilous, inamyloid.
Basidiospores: broadly ellipsoid, thin-walled, smooth, colorless, (3.5–)3.6–4.1 × (2.8–)3.0–3.5(–3.8) µm, L = 3.88 µm, W = 3.21 µm, Q = 1.19–1.22 (n = 60/2); acyanophilous, inamyloid.
Additional specimen examined: China, Yunnan Province, Yuxi, Xinping County, Mopanshan National Forestry Park, 23°56" N, 101°29" E, Alt. 2,200 m, on a fallen angiosperm trunk. 20 August, 2017, CLZhao 2574 (SWFC00002574, ITS: PV705578).
Notes: Morphologically, Tubulicrinis hongheensis resembles Tu. xantha, Tu. yunnanensis sharing the cylindrical to allantoid basidiospores. However, Tu. xantha can be distinguished from Tu. hongheensis by its slightly cream to slightly buff basidiocarps, and wider basidia (8–14.5 × 2.5–3.5 µm vs 7.5–14.5 × 4.0–7.0 µm)[72]. Tu. yunnanensis can be distinguished from T. hongheensis by its slightly cream to slightly buff basidiocarps, and wider basidiospores (4.2–6.5 × 1.2–2 µm vs 3.6–4.1 × 3.0–3.5 µm)[72]. Tu. hongheensis differs from Tu. chaetophorus by its shorter cystidia, wider basidia (4–7 µm vs 3–3.5 µm), smaller basidiospores (3.6–4.1 × 3.0–3.5 µm vs 5.5–6.5 × 3–3.5 µm)[2].
Tubulicrinis tenuis C.L. Zhao, sp. nov. Figs 17, 19

Figure 19.
Tubulicrinis tenuis (holotype CLZhao 17221). (a), (b) Basidiocarp. Microscopic structures: (c) Basidiospores; (d) Basidia and basidioles; (e) Lyocystidia; (f) Section through basidiocarp. Scale bars: (a) = 1 cm; (b) = 1 mm; (c) = 5 µm; (d)–(f) = 10 µm.
Index Fungorum number: IF 859191
Etymology: Tenuis (Lat.): referring to the thin basidiocarps of the type specimen.
Holotype: China, Yunnan Province, Wenshan Zhuang and Miao Autonomous Prefecture, Pingba Town, Wenshan National Nature Reserve, 23°22" N, 104°43" E, Alt. 1,500 m, on the fallen branch of Picea, 28 July, 2019, CLZhao 17221 (SWFC00017221, ITS: PV705580, LSU: PV705581).
Diagnosis: Basidiocarps annual, resupinate, adnate, pruinose, smooth; hyphal system monomitic, generative hyphae thin to slightly thick-walled, with clamp connections; cystidia present; basidiospores cylindrical to allantoid, thin-walled, smooth, 4.5–5.5 × 1.5–2 µm, acyanophilous, inamyloid.
Basidiocarps: annual, resupinate, thin, adnate, pruinose, without odor or taste when fresh, up to 8 cm long, 3.5 cm wide, and 150 µm thick. Hymenophoral surface smooth, white (–A1) when fresh and became white (–A1) to pale yellow (1A3–4A3) when drying. Sterile margin narrow, white, up to 1 mm.
Hyphal structure: hyphal system monomitic; generative hyphae with clamps, thin to slightly thick-walled, colorless, branched, interwoven, 1.7–2.5 µm in diam; cyanophilous, inamyloid, tissues unchanged in KOH.
Lyocystidia: cylindrical to slightly sinuous, thick-walled, smooth, colorless, abundant, projecting, bi-rooted, capillary lumen ending gradually, 28.5–66.5 × 5.2–8.3 µm, projecting above the hymenium; cyanophilous, inamyloid.
Cystidia: absent.
Basidia: barreled, small, thin-walled, colorless, with four sterigmata, clamped at the base, 7.0–9.5 × 3.8–4.5 µm; basidioles in shape similar to basidia, but slightly smaller; acyanophilous, inamyloid.
Basidiospores: cylindrical to allantoid, thin-walled, smooth, colorless, with one or two guttate, (4.3–)4.5–5.5(–5.7) × 1.5–2.0(–2.3) µm, L = 4.97 µm, W = 1.81 µm, Q = 2.75 (n = 30/1); acyanophilous, inamyloid.
Chlamydospores and anamorph not seen.
Notes: Morphologically, Tubulicrinis tenuis resembles Tu. pini, Tu. xantha, Tu. yunnanensis sharing cylindrical to allantoid basidiospores. However, Tu. pini can be distinguished from Tu. tenuis by having slightly buff basidiocarps, and larger basidia (11–14.5 × 4.5–5.5 µm vs 7–9.5 × 3.8–4.5 µm)[70]. Tu. xantha differs from Tu. tenuis by having yellow to buff basidiocarps, and narrower basidia 8–14.5 × 2.5–3.5 µm vs 7–9.5 × 3.8–4.5 µm)[71].
-
The species diversity and taxonomy of corticioid and hydnoid fungi have been the focus of research on aphylloporoid fungi due to their rich diversity and multifunctionality. Research on Chinese corticioid and hydnoid fungi has made significant progress in the past 20 years; in particular, the number of species has increased from 506 recorded in 2010[72], to 1,400 currently recorded, with most of the increase resulting from the discovery of new taxa. The number of species present in other countries and/or continents has been documented in previous studies: Northern Europe has recorded 463 species[73]; Europe and adjacent areas have recorded 822 species[2]; the temperate northern hemisphere, viz. all parts of Europe, the former USSR, and North America, with the exception of Mexico, has recorded 774 species from 131 genera[74,75]; South America has recorded 363 species[76]; Africa has recorded 488 species[76]; Italy has recorded 453 species[2]; India has recorded 328 species[77]; Australasia has recorded 210 species[78]; and North America, with the highest number of species before this study, has recorded 1,163 species from 23 orders, and 56 families[79]. Compared with the available global data, the total number of corticioid and hydnoid fungi in China exceeds that in Europe, South America, Africa, Australasia, India, and Italy, and North America, and ranks first in the world. However, as mentioned by multiple previous studies, the delimitation of corticioid fungi differs among mycologists: 'comparison of quoted total numbers of corticioid taxa should be treated with caution'[2,79].
This study revealed that the genera Hymenochaete, Phanerochaete, Tomentella, Xylodon, Hyphoderma, Hydnellum, Lyomyces, and Trechispora in China have the greatest number of species, with 115, 66, 64, 53, 51, 44, 38, and 36, respectively. This reflects the high species richness of these genera in China, and on the other hand, it also reflects the significant progress that has been made in the study of corticioid and hydnoid fungi in the past decade. The abovementioned genera include both saprophytic and ectomycorrhizal fungi, which share the common characteristic that the fruiting bodies are fully resupinate or resupinate-reflexed, and lack significant morphological characteristics to delimit species. Consequently, distinguishing closely related species by macroscopic and microscopic morphology is challenging, which is an important reason why these groups of fungi have been previously understudied. With the help of molecular biology techniques, which allow for the comparison of nucleotide sequence differences and the construction of phylogenetic trees, it is possible to explore and elucidate morphological differences that may not have been noticed by mycologists in the past, or may have been treated as variations between different specimens within a species. Therefore, considering China's vast territory, complex geological conditions, diverse climate, and abundant vegetation types, it can be predicted that with the exploration of fungal diversity, the number of corticioid and hydnoid fungi in China will continue to increase in the future.
Although there may be many undescribed species, there is still a severe lack of research on some genera. For example, the ectomycorrhizal fungus Tomentella-Thelephora lineage has very high species diversity and abundance in forest ecosystems[80,81]. Approximately 450 species names of Tomentella were recorded in the Index Fungorum (31 October, 2025) and MycoBank databases, but only 200 were accepted species. However, in sharp contrast, 1,400 species hypothesis codes (species hypotheses, SHs) of this genus were identified in the UNITE database with a threshold value of 1.5%[82]. A large number of DNA sequences related to ecological studies were obtained from the environment (mainly soil and root tips) by high-throughput sequencing techniques, and have been deposited in GenBank (NCBI). Owing to the lack of taxonomic studies and the lack of species names with reference sequences, massive numbers of sequences can be identified only as families and genera, which seriously hinders the in-depth study of many scientific issues related to ecology. Promoting traditional specimen-based taxonomic research on Tomentella-Thelephora fungi, and continuously exploring and describing new species, will help solve these scientific problems.
The correct species name is the foundation for development, utilization, and conservation. The checklist of corticioid and hydnoid fungi in China lists 60 genera of fungal species with various biological activities, accounting for 25% of all genera. If counted on the basis of species, this number may be less than 5%, which means that the vast majority of these fungi have not yet been explored. However, during the review of the biological functions of these species, we noticed that the study of secondary metabolites of these fungi may be questioned due to the correctness of species identification. For example, the name of Thelephora vialis Schwein., which is a thelephoid fungus described from the United States, has been cited multiple times by Chinese and Japanese researchers[83−85], and was used to name several new secondary metabolites on the basis of 'vialis' (Vialinins A, B, and C). However, considering the extensive species collection and studies in recent years[86], we speculate that 'Thelephora vialis' may not be found in China (or even Japan). In addition, hydnoid fungi include many species that need to be protected, such as Sarcodon, Hydnellum, and Phellodon. Many species in these genera are listed on the Red List in European countries (
www.iucnredlist.org )[87], but their species diversity and ecological habitats in China are still unclear. Given the current context of global climate change and multiple disruptions caused by human activities, research on the taxonomy and diversity of these fungi will help clarify and evaluate the goals of related protection. Therefore, from the perspective of the demands of relevant disciplines, research on the diversity and taxonomy of corticioid and hydnoid fungi in China is urgently needed. -
The authors confirm contributions to the paper as follows: conceptualization, supervision, funding acquisition, and writing – review: Yuan HS; investigation: Yuan HS, Zhao CL, Wei YL, Zhao H, Zhou LJ, Zhu YQ, Zhang XJ; data collection: Yuan HS, Zhao CL, Zhao H, Zhou LJ; analysis and interpretation of results: Yuan HS, Zhao H; draft manuscript preparation: Yuan HS. All authors reviewed the results and approved the final version of the manuscript.
-
The data that support the findings of this study are available in the National Center for Biotechnology Information (NCBI) databases, Index Fungorum, and the Fungal Names repository.
-
This research was financed by the National Natural Science Foundation of China (Project Nos U2102220, 32570013, and U23A20142), and the Fund of CAS Key Laboratory of Forest Ecology and Silviculture, Institute of Applied Ecology, Chinese Academy of Sciences (No. KLFES-2028).
-
The authors declare that they have no competing interests.
-
accompanies this paper online at: https://doi.org/10.48130/mycosphere-0026-0004.
-
#Authors contributed equally: Hai-Sheng Yuan, Lin-Jiang Zhou
- Supplementary Table S1 A checklist of Chinese corticioid and hydnoid fungi with their reference DNA sequences and applications.
- Copyright: © 2026 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yuan HS, Zhou LJ, Zhu YQ, Wei YL, Zhang XJ, et al. 2026. Species diversity of corticioid and hydnoid fungi in China and their medicinal, environmental, agricultural, and industrial values. Mycosphere 17: e005 doi: 10.48130/mycosphere-0026-0004 

Species diversity of corticioid and hydnoid fungi in China and their medicinal, environmental, agricultural, and industrial values
- Received: 13 November 2025
- Revised: 01 January 2026
- Accepted: 29 January 2026
- Published online: 17 April 2026
Abstract: Corticioid and hydnoid fungi are a group of aphyllophoroid basidiomycetes that perform important ecological functions. These fungi exhibit high species diversity, play important roles in energy flow and material cycling through saprophytic, symbiotic, or parasitic nutrient acquisition, and are widely distributed in various forest ecosystems around the world. Many of these fungi have important economic value and potential for use in multiple applications, particularly as the most important sources of lead compounds in innovative drug discovery in the pharmaceutical field. In the past decade, research on corticioid and hydnoid fungi in China has increased rapidly, with many new species being described, the phylogenetic positions of many taxonomic units being elucidated, and numerous new potentially available fungal resources being recognized. This study systematically compiled a total of 1,400 corticioid and hydnoid fungal species in China, which belong to 245 genera, 69 families, and 32 uncertain family-level taxa (incertae sedis), as well as 21 orders and three uncertain order-level taxa in two classes (Agaricomycetes and Dacrymycetes). The number of recorded species has increased by 2.8 times compared with that from 2010, with the majority of increases being attributed to the discovery of new species, and this number exceeds that in North America, and ranks first in the world. In this study, a checklist of corticioid and hydnoid fungi in China was compiled, and the available accession numbers of reference nrITS sequences are provided. The value of these fungi for application in the medicinal, environmental, agricultural, and industrial fields was summarized at the genus level. Additionally, ten new species were illustrated and described. This study provides a foundation for taxonomy, resource assessment, conservation, and sustainable utilization of these fungal resources.