









-
Originating in the African continent, the Coffea genus includes 103 species belonging to the Rubiaceae family[1]. Among these species, C. arabica L. and C. canephora Pierre ex Froehner are commercially important, representing 60% and 40% of the world's coffee production in 2018/19, respectively[2]. Coffee is one of the most popular commodities worldwide, with world exports reaching around 10.5 billion kg of fruit in 2020, generating more than US$ 26 billion[2]. However, recent studies suggest that coffee production will be highly affected by warming in future predicted scenarios, especially in Brazil, the world's biggest producer[3,4]. From a biological point of view, the coffee genus presents peculiar characteristics, such as a two-year cycle and uneven flowering and ripening processes, that have not been extensively explored molecularly, and could be useful in understanding the flowering evolution in crops. Although recent advances regarding the environmental and endogenous factors affecting coffee production have been reported[5], these aspects were less explored during the flowering process and a general overview to clarify and direct future studies is essential for improvements in this important crop.
Coffee plants present a phenological cycle divided into two years, with the vegetative and reproductive phases occurring simultaneously after the first year of flowering[6], and the reproductive development can be divided as follows: development of branches and floral buds (FB) induction, FB development, anthesis, and fruit development[6, 7]. This organization was based on field observations and generalized to different coffee genotypes that present phenotypic differences. However, information at the molecular level of the coffee phenological cycle is scarce, and such information could help to unravel the biocycle and intraspecific differences[8]. For instance, there are genotypes considered to be early and others as late in their flowering pattern[9]. In addition, this process is reported as being strictly related to a drought period and reduced temperatures (winter season) in the main Brazilian coffee producing regions, with anthesis taking place as the rainy season starts[10]. However, the interplay between reproductive development and its connection with environmental cues finds no molecular evidence supporting such an influence or discriminating its relationship with the metabolic and energetic status that could trigger developmental processes.
Flowering is one of the most important developmental processes in the plant life cycle, and inappropriate environmental conditions can negatively affect it, decreasing the number of progenies produced, and thereby, interfering in the plant performance in natural environments for yield and quality[11]. Coffee flowering is an asynchronous process, with this sequential flowering resulting from the asynchronous development of buds, an intrinsic characteristic of the species. It often leads to uneven fruit maturation, which can greatly affect the final product and coffee cup quality[12,13].
In C. arabica producing areas around the equator, a higher number of flowering events are observed due to the absence of marked periods of water deficit, whereas in areas relatively far from it, dry periods are longer and allow coffee trees to display one main flowering event[10,14]. Water deficit and rehydration are the most influential factors for coffee flowering[15−17]. Although water influences coffee anthesis, other external and internal factors are also involved, such as modifications in temperature and humidity[18], photoperiod[19,20], plant nutrition status[21], production system (shade or full sun)[7,22], altitude[23], water deficit[17,24−26] and phytohormones[27−31]. To facilitate comprehension of these processes, we summarized the main changes occurring throughout the year, with the genes possibly related to them, and proposed a new classification of buds based on their position in the node, which demonstrates the previously mentioned uneven floral bud development.
Even though many studies have been developed to better understand the effect of environmental factors affecting coffee flowering, less attention has been directed to the molecular aspects connecting all these processes. Based on transcription profiles, our knowledge until now, is that different transcriptional factors seem to be involved in coffee flower development[13], and in silico analysis and expression profiles show that the C. arabica FLOWERING LOCUS C (CaFLC) or FRIGIDA homologs might be involved in coffee flowering regulation[32, 33]. However, due to the difficulties in obtaining coffee mutant plants, the function of these genes and other genes, such as FLOWERING LOCUS T (FT), LEAFY (LFY), SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1 (SOC1), CONSTANS (CO) have not yet been studied. In addition, different hormones seem to be involved in controlling coffee flowering, especially abscisic acid (ABA), ethylene, and gibberellin. Recent findings on the expression patterns of ethylene biosynthesis and signaling genes, as well as on ethylene production levels under different environmental conditions, suggest that this hormone is directly associated with the control of C. arabica flower opening time, and from the crosstalk with ABA and gibberellin, competence for responding to environmental cues and restarting their development is obtained by coffee flower buds, allowing anthesis to take place (Fig. 1). Nevertheless, further studies are necessary to link all of these elements. Here, we review the recent advances in the coffee flowering process and connect them to the historical knowledge about this crop to provide a better understanding of the coffee phenological cycle and perspectives to its control in different and challenging environmental scenarios.
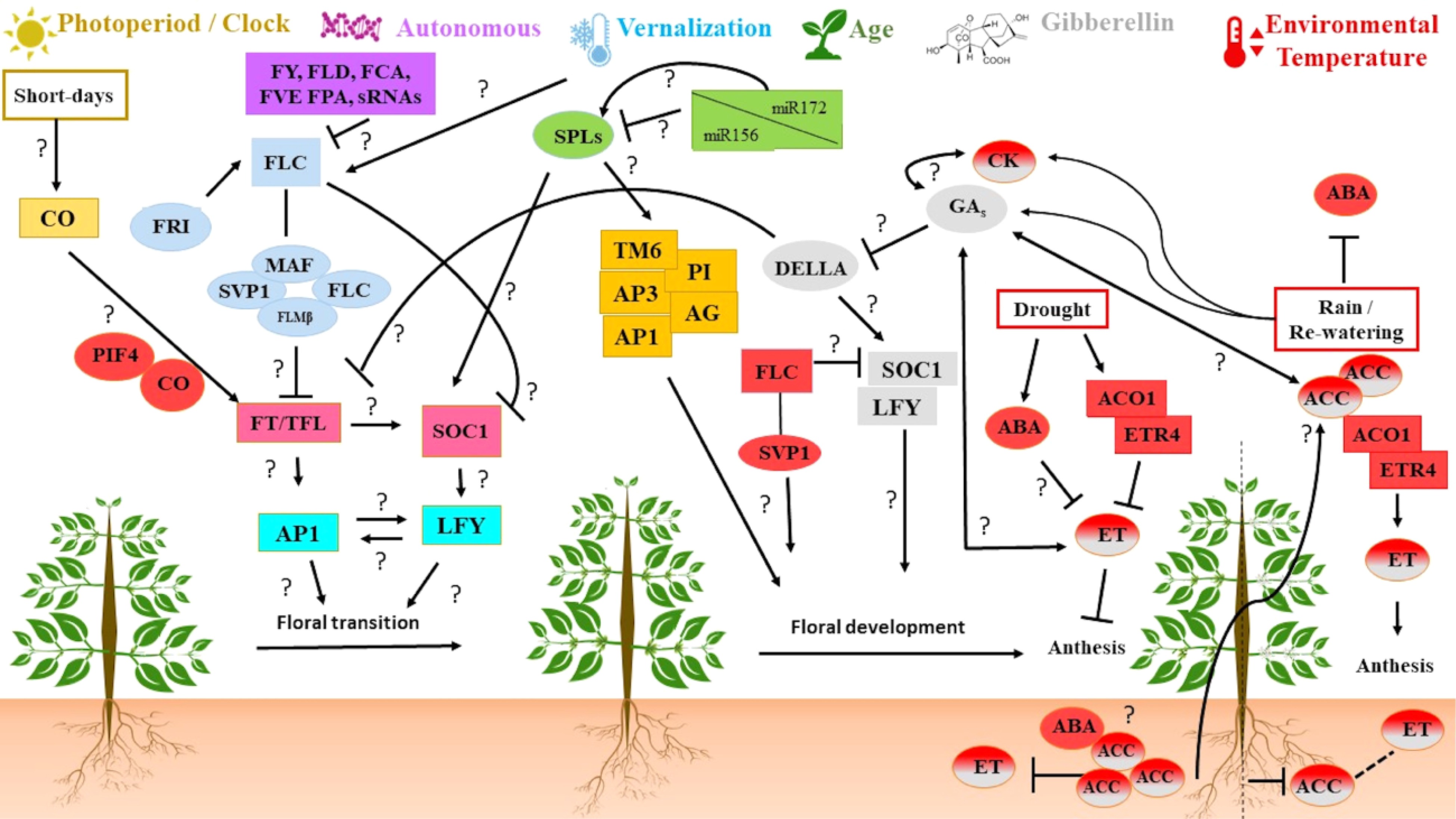
Figure 1. Summary of molecular pathways possibly involved in the flowering time and developmental processes in C. arabica: photoperiod/clock (yellow), autonomous (purple), vernalization (sky blue), age (green), gibberellin (gray), and environmental temperature (red). Pink boxes represent the main floral integrator genes. Light blue boxes represent the inflorescence meristem identity genes. Orange boxes represent the homeotic genes. Squared boxes represent genes having a pivotal role in the specific pathway. Circles represent several proteins or complexes. Solid lines indicate induction, dashed lines indicate no significant change in the relative amount and, the T end to the arrows indicate decreases. Interrogations indicate connections proposed in this review based on previous studies found in the literature for other species, but not confirmed for coffee. Relationships involving environmental changes associated with water (drought, rain, irrigation, re-watering) are connected in the environmental temperature pathway. The red and gray gradient in the circles indicates a possible interaction between the pathways. ABA: abscisic acid, ACC: 1-aminocyclopropane-1-carboxylic acid, ACO: 1-aminocyclopropane-1-carboxylic acid oxidase AG: AGAMOUS, AP1: APETALA 1, AP3: APETALA 3, CK: CYTOKININ, CO: CONSTANS, ET: ETHYLENE, ETR4: ETHYLENE RECEPTOR 4, FCA: FLOWERING LOCUS CA, FLC: FLOWERING LOCUS C, FLD: FLOWERING LOCUS D, FLM: FLOWERING LOCUS M, FPA: FLOWERING LOCUS PA, FRI: FRIGIDA, FVE: FLOWERING LOCUS VE, FY: FLOWERING LOCUS Y, GA: GIBBERELLIN, LFY: LEAFY, MAF: MADS AFFECTING FLOWERING PI: PISTILLATA, PIF4: PHYTOCHROME INTERACTING FACTOR 4, SOC1: SUPPRESSOR OF OVEREXPRESSION OF CONSTANS 1, SPL: SQUAMOSA PROMOTER-BINDING PROTEIN, sRNAs: SMALL RNAs, SVP: SHORT VEGETATIVE PHASE, TFL: TERMINAL FLOWER, TM6: TOMATO MADS BOX GENE 6.
-
Coffee cultivation is carried out under two production systems, under full sun or under agroforestry systems, depending on the geographic location and the type of agricultural exploitation desired in terms of costs and sustainability[34]. The photosynthesis rates are different in these two systems, with the reduced light received under the shade system directly affecting reproductive organs, drastically reducing flowering rates[35]. The solar radiation intercepted by coffee plants during their growth has a direct effect on the development of tissues, storage of photoassimilates, photosynthesis rate, flowering and, above all, the distribution of the energy needed for the flowering time and fruit set[36]. In a plantation under full sun conditions, anthesis events are more intense, which has a positive impact on ripening uniformity, although trees present a biennial production pattern[7,22]. On the other hand, shaded coffee trees tend to flower and produce fruits with the same intensity every year[34], but reductions of up to 20% have been reported in coffee plantations under shade production systems when compared to full sun conditions[37]. The decrease in flowering intensity is due to the lower carbon assimilation rates present under shade conditions, a greater growth stimulus of vegetative organs than the reproductive ones, and a lower number of nodes per branch and buds per node[38]. However, and curiously, larger bean size and a better cup coffee quality has been observed in this shade condition[39]. Physiologically, shading causes changes in the microclimate, especially in temperature and, due to the genetic plasticity of coffee trees, it can affect their anatomy[40] and physiology[41], and influence the process of floral induction, differentiation, and anthesis. The impacts depend on the type, density, duration, and shading time, as well as the prevailing climatic conditions.
The strength of flowering is a function of the monthly light intensity around flowering[35, 42]. Light quality and intensity, photoperiod, or day-length affect the plant growth, directly influencing flower development and many other traits[43]. In coffee, studies related to the photoperiod influence on flowering have found different results. In young and old coffee trees, floral initiation is stimulated by short days and delayed by long ones[27] and, together with phenological observations, coffee has been classified as a short-day plant[20]. However, other researchers have concluded that in equatorial regions, where day-length variation is small, coffee plants are not influenced by day-length and do not have a specific flowering season, as observed in Colombia, Costa Rica, and Central America[44, 45]. Thus, the inductive stimulus of the coffee reproductive cycle is still not clear and may be affected by the interplay of different environmental factors, in which the molecular aspects are poorly explored (Fig. 1).
With regard to Brazil, the main coffee-producing regions of the country are characterized by marked seasons. After the March equinox, day length reduces to less than 13 light hours and this reduction acts as a signal for FB development, the transition from vegetative axillary buds to reproductive buds[20]. The perception of this stimulus constitutes the stage of floral evocation (Fig. 2a). Then, morphophysiological changes trigger floral development, enabling the phenotypic distinction between vegetative buds and FBs creating a range of different stages characterized from G1 until G4[6,7,16]. During FB development, consecutive transformations over two months (Fig. 2a) promote their growth until the G4 stage (> 4 mm) (Fig. 2b), the FB developmental stage responsive to the water stimulus that triggers anthesis (Fig. 2b & 2c). This coincides with the winter season in the main coffee-producing areas of Brazil (Fig. 2a), where days are shorter and are accompanied by a lack of rain events, with G4 floral buds entering a latent state. Then, after an increase in the leaf water potential due to rain or irrigation, anthesis are triggered[6, 7, 12].
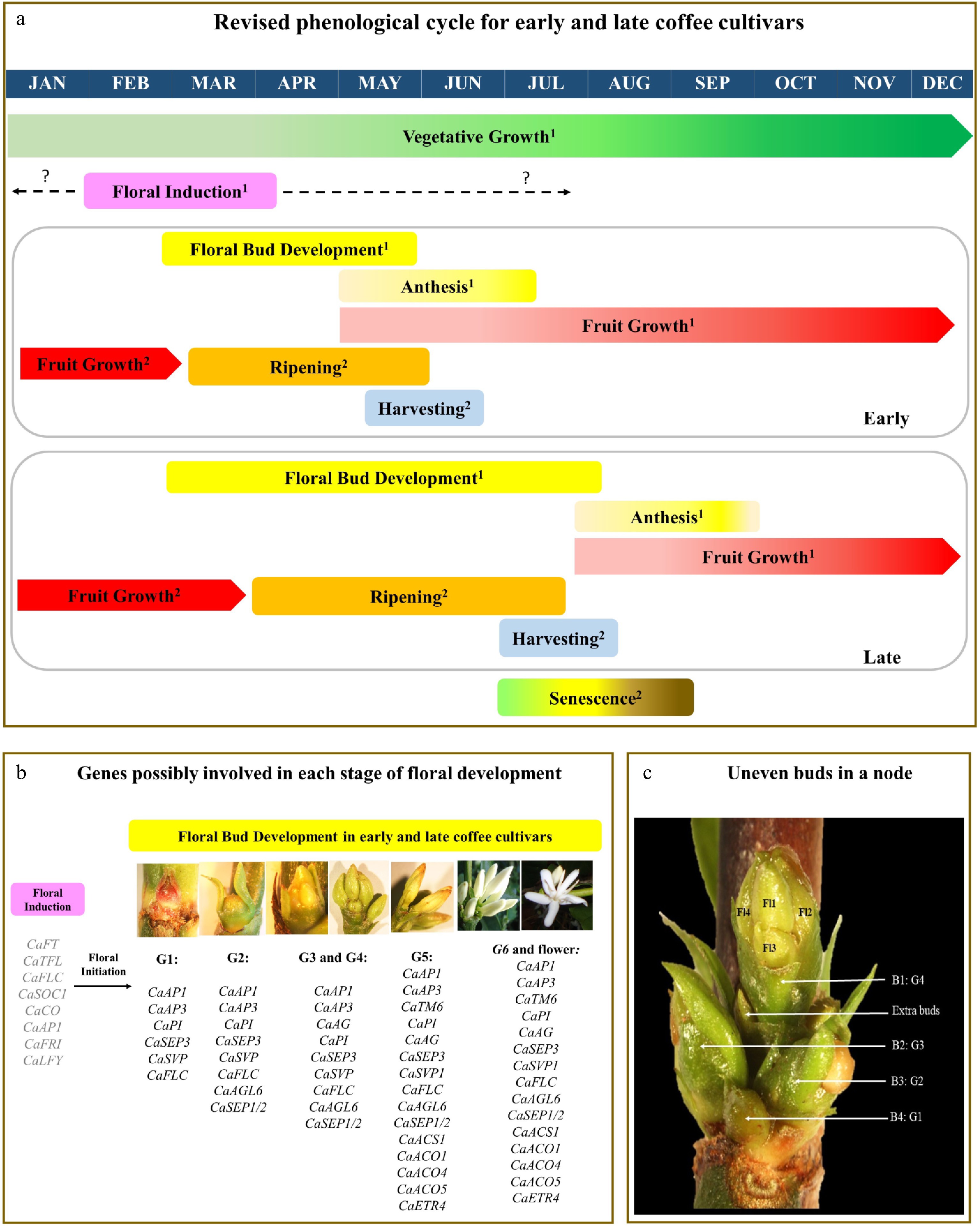
Figure 2. Proposed model for the induction and floral C. arabica development under Brazilian environmental conditions. (a) Revised model for vegetative and reproductive development in C. arabica cultivars regarding flowering time: early and late cultivars. The intensity of vegetative growth (leaf bud formation) for the two cultivars is shown by the gradient in the color of the green box. Floral induction (pink box) described in the literature between February and April is proposed to start in January, extending until July for early and late cultivars. The period of floral development (yellow box) is contrasting between cultivars, lasting 3 months in the early cultivar and 5 months in the late cultivar. Shortening in floral development stages in the early cultivar promotes anticipation of anthesis (the yellow box with gradient: light yellow indicates less intense flowering and dark yellow indicates greater or main flowering). While the late cultivar starts the lesser intensity flowering, the early cultivar advances in fruit growth (red box). Fruit development stages (pinhead, expansion, graining) for both cultivars end at the beginning of the second phenological year, followed by fruit ripening and harvesting. Self-pruning is shown as the progress from senescence to death of tertiary and quaternary branches (box with green to black color gradient). A dashed arrow suggests change at the beginning and end of the respective phase. Interrogation indicates an unproven relationship. Superscript numbers indicate the first (1) or second (2) year in the coffee tree's phenological cycle. (b) Genes are possibly involved in each stage of floral development in C. arabica. The floral induction process involves the activation of genes by environmental and endogenous pathways related to the differentiation of the Stem Apical Meristem (SAM) reproductive meristem. Then, flower initiation is initiated by combining flower organ identity genes related to the specification of whorls in floral buds. In the G5 and G6 stages, genes of the ethylene pathway (CaACS1, CaACO1, CaACO4, CaACO5) are related to opening flower progress. Genes in gray represent the connection suggested in this review and genes in black are proposed based on the quantitative and qualitative gene expressions found in the literature[13,32,33]. (c) Photo describing the uneven development of floral buds in a node located on the branch of a coffee plant. Four floral buds (B1, B2, B3, B4) are shown in ascending order of emergence with the respective identification of developmental stages (G4, G3, G2, G1[7]). In the floral bud B1 are indicated the four flowers (Fl) that finish its development with the anthesis. Thus, each node can potentially produce 16 flowers (four flowers from each bud[13]), but B3 and B4 commonly remain latent and, in some cases, extra floral buds can be developed, as identified in this photo.
The day length is detected by several components of the circadian clock that transform the inductive photoperiodic signal into chemical and molecular signaling directly associated with gene expression regulation related to flowering[46,47]. Many studies show that FT promotes the conversion of the Shoot Apical Meristem (SAM) into inflorescence meristems, where floral meristems are originated[48−50]. The FT is expressed in the leaves, and its expression is under the control of the CO, whose protein accumulates during the day. FT protein is translocated to the SAM through the phloem, forming a floral activation complex (FAC) from its interaction with a zip protein FLOWER LOCUS D (FD) and the 14-3-3 protein[51]. This protein complex is responsible for acting as a transcription factor to activate flowering identity genes, such as the SOC1, APETALA1 (AP1), and FRUIT FULL (FUL), which are directly associated with flower organ formation[52,53].
Morphology changes related to the reproductive development in the SAM and Axillary meristem (AM) are preceded by changes in the transcriptional level and the protein abundance of around 300 genes[54]. TERMINAL FLOWER 1 (TFL1) protein plays an antagonistic role with FT protein in the SAM, where it also interacts with the FD to form a floral repressor complex (FRC), responsible for keeping the SAM and AM in a vegetative state[55]. These two proteins, and the balance between them, had not yet been described in coffee, but FT and TFL1, known as flowering inducer and flowering repressor respectively, are directly associated with the inflorescence and branch architecture in annual and perennial species[55−57].
This balance is linked with the LEAFY (LFY) and AP1 regulation as shown that AP1 and LFY control TFL1 expression, while TFL1 is suppressed by AP1 and promoted by LFY[58]. This behavior might provide the basis for a better understanding of the determination of vegetative buds into reproductive ones in coffee (Fig. 1), as well as into the maintenance of vegetative branch growing since the asynchronous flowering and the development of new vegetative branches in coffee seems to be associated with a balance between vegetative and reproductive identity. LFY and AP1 play a central role in the determination of the SAM from its vegetative to its reproductive form[58], and is directly associated with the transcription regulation of key genes involved in floral identity, such as TFL1 and FD[51, 59]. Although the closest homolog of AP1 was already described in coffee plants, named CaAP1, and its expression pattern agrees with its described function in other species[13] the FT/TFL1, FD, LFY, CO, and other homologs related to the circadian clock have not been yet described in coffee. Studies involving these active hubs during the floral transition are necessary to a comprehensive overview regarding the control of flowering in coffee (Fig. 1) and helping to correlate to flowering control in other tropical perennial species
-
Several studies have shown the effect of temperature on coffee development and the overall impact on the flowering time and yield[8, 18, 60−62], including a negative warming scenario predicted for the next decades[63]. In coffee plants, the climate impacts are even more complex to evaluate because an 'optimum' temperature also depends on the species, phenological and developmental stage of the plant[64]. The adequate mean annual temperature for C. arabica cultivars is usually in the range of 18−23 °C[20] whereas for C. canephora is from 22−26 °C[65, 66] being explored in both species the effects of gradual increasing temperature or extreme heat stress[18, 67]. Minimal impact on photosynthetic-related parameters occurs in leaves when plants were exposed to temperatures up to 37 °C, but severe damage occurs at 42 °C[68]. However, this impact depends on tissue and species since elevated temperatures seem to cause, in C. arabica, abnormal reproductive structures[18] or loss of flowers and fruits when subjected to 45 °C for 1−1.5 h[69], but such effects were not described for C. canephora. Accordingly, in C. arabica, high temperatures associated with intense water deficit levels during the beginning of the anthesis period causes pollen tube dehydration and floral atrophy, leading to flower abortion due to their premature opening, with the petals remaining small and stiff, acquiring a starlet shape[6, 69, 70]. Although the impact of temperature in leaves and bean production has been established for coffee[64], such effects on flowering have not yet been quantified and the related molecular pathways not yet described.
Associated with the circadian clock genes and photoperiodic pathways, temperature is also a major stimulus affecting flowering control, being demonstrated that CONSTANS is responsive to high and cold temperatures to regulate FT transcription and protein accumulation (Fig. 1)[68, 71−73]. Alternatively, there are CO-independent pathways responsive to temperatures that directly regulate the FT expression. For example, at warmer temperatures PHYTOCHROME INTERACTION FACTOR 4 (PIF4) is activated and binding on the FT promoter activating expression[74, 75]. Opposite to warming, the flowering initiation and timing is also affected by exposure to cold periods (or vernalization) through the action of the FLOWERING LOCUS C (FLC; Fig. 1), an important process to guarantee plant reproductive success after winter periods[76]. Since the long process of coffee flowering faces different temperatures in Brazil, that coincides with developmental changes, for example floral meristem activation at summer and latent states of floral G4 buds in winter (Fig. 2), it is possible a direct influence occurs as suggested in some research[12, 25, 60, 64, 66]. However, although the relationship between thermosensory pathways and flowering mediated by PIF4 and FLC is described for plants, in coffee this molecular link has not been established (Fig. 1).
In contrast to PIF4, which has no sequences reported yet in coffee, an FLC homolog named CaFLC was described and its expression profile determined at some tissues and growth conditions providing cues about its role[13, 32, 33]. FLC is better understood in Arabidopsis being described as MADS-box transcription factor that form a protein complex with SHORT VEGETATIVE PHASE (SVP) to suppress the transcription of a set of genes related to the floral meristem identity, including FT, SQUAMOSA PROMOTER BINDING PROTEIN-LIKE 15 (SPL15), SOC1 and SEPALLATAs (SEP)[77−81]. FLC is repressed by vernalization, thus, its diversification is associated with control of the flowering time synchronizing the reproductive period with warmer temperatures during spring[77, 82, 83]. In coffee, CaFLC is significantly expressed in vegetative tissues, as roots, leaf and apical meristems, but also in reproductive ones, which include all floral developmental stages and fruiting[13]. This is a very similar pattern to the SVP homolog CaSVP-1, in contrast with the paralogs CaSVP-2 and CaSVP-3 only being expressed in vegetative tissues[13]. This coexpression is in agreement with the reported complex FLC-SVP[79], however the variable expression of CaFLC throughout the year during floral development (Fig. 2) suggests other regulatory roles than the repression of flowering, which was also reported in Arabidopsis[84]. Supporting this hypothesis, the SVP subclade diversification in coffee, represented by the three paralogs, could indicate a specialization of the combinatorial protein complexes possibly regulating different targets and developmental processes depending on the tissue.
Furthermore, recent data support the regulation of CaFLC by a cold-independent pathway coordinated by FRIGIDA (
CaFLR-like)[33]. In A. thaliana, FRIGIDA (AtFRI) regulates the floral transition, assembled with other proteins in a complex that binds to the FLC promoter triggering its expression and, consequently, repressing flowering[85]. Vernalization has no effect on AtFRI expression but, similar to AtFLC, the allelic variation is associated with flowering-time diversification[86, 87]. Other functions have been attributed to the interaction between AtFRI and AtFLC being related to ovule development in flowers of non vernalized plants[88] and the repression of SOC1, a MADS-box gene related to the floral meristem transition, necessary in development of the late stages of flowering[89]. Accordingly, in C. arabica, CaFLC were detected at late floral stages, G4 to anthesis, showing an expression pattern very similar to CaFRL-like transcripts[33], which confirmed previous analysis showing CaFLC is mainly expressed at the final stages of coffee floral development[13]. Thus, these results suggest different roles for CaFLC in coffee and are probably not related to cold, although this hypothesis has yet to be addressed, since the control of flowering time is a major concern for the sustainability of crop production. As far as we know, no functional or deeper transcriptional analyses of PIF4 and FLC homologs were made in coffee considering different temperature conditions and/or along the Brazilian seasons to associate with the FT regulation. Interestingly, PIF4 and FLC regulation in response to temperature changes involve epigenetic control of chromatin activity[82, 90, 91], which is an aspect completely unexplored in coffee despite being considered a promising field for flowering control and crop breeding[92]. Another difficulty to relate cold with the coffee floral development and CaFLC, is that it coincides with the drought period during the Brazilian winter, which impacts coffee physiology and production[64]. Expressed sequence tags (ESTs) referred to CaFLC were found in libraries of water-stressed plants cultivated in the field (pool of tissues)[93] and, in agreement, it was upregulated in leaves under drought[32]. Thus, it is possible that CaFLC is drought-sensitive, participating in floral development, especially from G4 to anthesis, and/or synchronizing the flowering time. However, drought seems to be a non-essential regulatory signal since constantly irrigated coffee trees flourish normally[15]. Alternatively, it seems more plausible to interpret the cold and dry period before anthesis as an adverse environmental condition, that delays the floral development as a consequence of the impact on energy metabolism (sugars), instead of having a direct effect on regulatory flowering. Supporting this, coffee trees that faced severe drought showed decreased leaf hydraulic conductance and carbohydrate content[94], together with stomatal function alterations mediated by aquaporins[95]. Based on this, in our opinion, these could explain the observed synchronization of flowering associated with drought during cold periods, since a decreased energy status could support the development of floral buds at the initial stages (lower energy demand) but delays at advanced stages (higher energy demand). Similarly, the water supply is in higher demand during the advanced stages of floral development to support cell expansion and rapid growth of the already determined organs, in contrast to the initial stages[13], thus, its absence also could contribute to floral bud synchronization. The water deficit relationship with coffee flowering and the involvement of hormones in such developmental process is detailed below.
-
Coffee plantations are exposed to different climatic conditions around the world[14], with this wide range of conditions affecting their growth, development, and production[96]. Water is one of the most limiting factors that coffee plants face during their development[96], and it can directly influence coffee developmental processes, such as the time and number of flowering events[14]. An adequate rainfall distribution throughout the year leads to a higher number of flowering events in coffee plantations close to the equator, whereas in areas relatively distant from this region, dry periods are longer and allow coffee trees to display one main flowering event[14]. During the dry period, coffee flower buds are thought to become sensitive to respond to the stimulus for regrowth[16, 97−99], and only buds at the G4 stage seem to be able to respond to this stimulus, possibly due to the fact that only from these buds, on a well-defined vascular cylinder, containing secondary xylem, is present on their pedicels[15].
Different studies have focused on identifying adequate levels of leaf water potential required for floral buds to restart their growth after the dry period in an uniform and synchronized manner, without affecting other physiological functions, such as photosynthesis, cell expansion, and leaf growth[15, 24, 28, 100, 101]. However, the intensity of the water stress required for making coffee flower buds competent to restart their growth is still controversial. It was shown that coffee anthesis can be induced with a relatively short period of a severe water deficit, as long as predawn leaf water potential (Ψpd) is below −0.8 MPa[15]. In this same study, it was shown that more negative leaf water potential did not promote an enhancement in flowering stimulation, and a similar flowering stimulation could be induced with less severe, but more prolonged, water deficit. It was found that the critical value of Ψpd for triggering flower opening was approximately −1.2 MPa, and it was shown that flower bud dormancy was broken only when Ψpd reached −2.6 MPa[28, 102]. These previously mentioned studies also have shown that if these threshold leaf water potentials are not reached, flower opening does not take place, with flower buds reaching the G4 stage, and eventually declining in number due to bud senescence. However, most of these studies were conducted under greenhouse conditions and bud senescence is not usually seen under field conditions. Some authors attribute this to the fact that the cumulative exposure to short water deficit periods under field conditions, during hot days, would be enough to make flower buds competent to flower[103]. From a practical point of view, the longer period of dormancy imposed by the dry period allows that flower buds at different developmental stages reach stage G4, enabling a more uniform flowering upon soil rewetting[104]. Importantly, although most of these studies mention that coffee floral buds enter into a dormancy period, which is broken by rain or irrigation, once they reach the G4 stage[7], there are no studies showing the physiological basis of this type of dormancy, and we suggest that the correct term to be used should be 'dormant or latent state'.
Rainfall during the latter stages of flower development can negatively affect the flowering process causing an increase in the number of blossoming times, which consequently affects ripening uniformity, however, this is not always the case. It was found that coffee trees continuously irrigated or exposed to some period of water deficit, showed the same number of flowering events, emitting flowers to a similar extent[105, 106]. In terms of productivity, non-irrigated plantations usually show a decrease in productivity, however a less severe dry period (irrigation suspension for up to 60 days) can show similar, or even higher, productivity levels compared to well-watered plants, with the advantage of displaying a higher number of fruits at the ideal ripening stage for harvest, the cherry stage[26, 101, 105, 106]. Accordingly and more recently, it was shown that the rate of opened flowers increased with the reduction of Ψpd, and though the leaf gas exchange was strongly affected by water deficit, coffee yield was not affected and the percentage of cherry fruits increased slightly with the reduction of Ψpd[17]. In addition, this study showed that moderate water deficit levels (Ψpd of approximately −1.15 MPa) are adequate to cause a high percentage of anthesis in flower buds, and the increments in anthesis decreased with more severe water deficit levels, as observed in other studies[16]. However, climatic conditions among the different coffee production regions vary greatly, and for this reason the timing and extent of water deficit implementation in not causing any negative effect on the productivity levels may also vary.
Thus, although coffee flowering under well-watered conditions may show contradictory patterns in some cases, and the intensity of water stress required to make flower buds competent to respond the stimulus for regrowth is still not defined, a period of water deficit seems to be essential, not only to permit a synchronization of flower bud development, but also for making flower buds competent to flower[15, 17, 26]. However, how water deficit turns coffee flower buds competent to flower and the mechanism through which flower opening is triggered, are still poorly understood processes in coffee flowering.
-
Although there is a scarcity of studies relating the involvement of plant hormones in coffee flowering as a whole, they are certainly involved in coffee flower development, especially gibberellin, ABA, and more recently, ethylene. Gibberellin-mediated flowering control is a hormonal regulation pathway with effective participation of ethylene. In Arabidopsis, increased levels of ethylene induces the GAs degradation by the 26S proteasome, promoting the accumulation of DELLA protein and thus delaying flowering[107]. DELLA proteins have been shown to act as negative flowering regulators and their increased levels are associated with floral opening delay[108−110]. The DELLA-GA interaction is inversely proportional, and in the absence of GA, these proteins repress the LFY and SOC1 transcription involved in the signaling to activate floral identity genes[107,111]. Gibberellin (GA3) application induced anthesis of flower buds larger than 4 mm in length in the absence of precipitation[27]. This effect in anthesis promotion was present only in plants exposed to some level of water deficit, and under relatively mild water stress conditions, exogenous GA application was mandatory for triggering anthesis[28]. Gibberellin concentration appears to increase upon rewatering before buds gain fresh weight, and since GA biosynthesis inhibitors did not prevent the release from the dormant or latent state of the floral buds, being supposed that GA is converted from a bound form to a free, active form at the time of release from dormancy[112]. More recently, studies show that exogenous GA does not increase the number of coffee floral buds[113], but it may anticipate anthesis when GA3 is applied in G4 floral buds promoting flowering and fruit ripening synchronization[30]. Accordingly, after rain or irrigation, GA levels increase rapidly in the buds and gradually decrease with the expansion of the floral organs, while its levels in the xylem remain stable[113]. Similarly, after rehydration, cytokinin levels increase in floral buds and xylem, suggesting a synergistic effect with GA in floral development and anthesis[112]. Therefore, GAs can regulate advanced stages of floral development in coffee, although the flowering-related gibberellin pathway has not yet been explored. Although coffee flower bud dormancy is suggested to be maintained by a balance between GA and ABA, ABA levels don't seem to be different between dormant and non-dormant buds[112]. Relatively high ABA levels are present in coffee floral buds during the drought period that precedes anthesis[35,114], and recently it has been suggested that a crosstalk between ABA and ethylene may be involved in the regulation of coffee anthesis[31].
In relation to flowering, ethylene has been shown to inhibit[107], or promote[115, 116] flowering depending on the species, and it regulates different aspects of flower development, including pollen and ovule development[117, 118], flower opening[119, 120], and flower senescence[121].
Ethylene's involvement in coffee flowering was first suggested considering a series of events, water deficit followed by plant rehydration, that is known to produce a burst in ethylene production in woody species[122], due to the accumulation in roots of the ethylene precursor ACC during water deficit and its transport to the shoot once plants are rehydrated. In addition, floral bud competence and regrowth are probably controlled by a root-sourced signal[15].
Recently, an association has been shown between ethylene levels and the expression of ethylene biosynthesis and signaling genes with anthesis induction triggered by rain, as well as by growth regulators[31]. Water deficit downregulates the expression of coffee ethylene biosynthesis genes, which are characterized by multigene families[123], and decreases coffee ethylene production in shoots. Decreased ethylene levels in response to water deficit has been shown to occur in other species, such as rose and other herbaceous species[124], and even in coffee itself, as observed in floral buds[28]. This regulation pattern may be associated to ABA, which has been shown to accumulate during the dry season in response to leaf water deficit[125] and that may limit expression of ACO genes and the activity of the enzymes encoded by these genes[126,127]. On the other hand, root ethylene evolution does not change in response to water deficit, but ACC, the ethylene precursor, seems to accumulate in this tissue considering the downregulation of ACO genes under this condition[31]. The accumulation of ACC under water deficit conditions occurs in other woody species as previously mentioned, and plant re-watering stimulates ACC transport to the shoot, inducing an ethylene burst[122], which may be involved in anthesis induction in coffee. Indeed, re-watering coffee trees after a period of water deficit increase shoot ethylene production[31], similar to what has been found in other species such as wheat (Triticum aestivum)[128] and tomato (Solanum lycopersicum)[129], possibly through the transport of the accumulated ACC in the roots[129,130]. Ethylene, or even ACC, which recently has been shown to act as a signaling molecule independently of ethylene signaling[131], could than help in inducing coffee anthesis by maybe inducting a set of rehydration-responsive genes, as observed for ethylene in rose (Rosa hybrida)[132]. However, increased levels of ethylene seem insufficient to promote coffee anthesis, and modifications on ethylene sensitivity are also necessary[31]. Ethylene exerts its action via the ethylene signalling pathway, where it is perceived by a family of receptors and its signal is mediated downstream by members of different gene families[133]. Among these members, ethylene receptors mediate ethylene sensitivity, acting as negative regulators[134], meaning that decreased amounts of ethylene receptors enhance ethylene sensitivity[135−137]. Interestingly, 1-MCP (1-Methycyclopropene), an ethylene action inhibitor[138, 139] application seems to mimic plant rehydration by up-regulating ethylene biosynthesis genes, possibly due to the loss of the negative feedback regulation of ethylene biosynthesis[140, 141], and inducing ethylene production, triggering coffee anthesis and thus overcoming the requirement of the series of events composed by soil drying and re-watering for anthesis induction[31]. In addition to modifying ethylene evolution, 1-MCP application potentially increases ethylene sensitivity, considering the elevated ethylene levels and the reduction in the expression levels of the ethylene receptor analyzed CaETR4-like[31], which seem to be essential for inducing coffee anthesis, considering that the surfactant co-applied with 1-MCP also promoted the induction of ethylene biosynthesis genes, but it did not modify CaETR4-like expression as observed in response to 1-MCP treatment, not being able to induce coffee anthesis[31]. Thus, the study conducted by Lima et al. concluded that during the dry season, ethylene evolution decreases in the shoots of coffee trees, possibly due to the elevated levels of ABA[126, 127, 142], decreasing ethylene sensitivity, since expression of the ethylene receptor CaETR4-like did not change during this period. Under these conditions, flower buds are maintained in a dormant state, preventing their progression from the G4 developmental stage to anthesis. Upon rewatering, the possibly lower ABA levels, and the ACC accumulated in the roots during the dry season is transported to the shoot, enable the elevation of ethylene levels in the shoot, increasing the sensitivity to ethylene, allowing coffee anthesis to take place by possibly ensuring rehydration recovery in flowers, as observed in rose[132].
Studies involving the effects of 1-MCP on coffee receptors and the ACC as a signaling molecule in an independent pathway from ethylene on the metabolic network of coffee flowering are ongoing. Together, these data will be helpful to elucidate the hormonal crosstalk and connect flowering mechanisms in coffee trees, possibly contributing for a better understanding of these processes in other woody species.
-
The transition of meristems to a reproductive state in coffee is regulated by a complex regulatory network, in which transcription factors, especially Flowering Locus T (FT), interact with environmental and endogenous cues to determine the coffee flowering time (Fig. 1 & 2). After this transition, the floral meristems arise and the floral organogenesis is initiated being coordinated by other transcription factors, such as, APETALA1 and LEAFY[143, 144]. The core regulatory mechanisms are at some level conserved in angiosperms, but it has also diversified throughout gene duplications and functionalization changing phenotypic characteristics and expanding competitive strategies among species[145]. For example, the coffee flower is anatomically complete with four distinct whorls that develop centripetally, first sepals, followed by petals, stamens and carpels. This pattern of development is similar to model plants, such as Arabidopsis and tomato, but phenotypical differences are also observed as a two-year developmental cycle, epipetalous stamens, and secretory structures named colleters[8]. Several studies were dedicated to understanding floral diversification[146−148], however, this was poorly explored in the Coffea genus (Rubiaceae), which would clarify many biological aspects, such as: i) floral development in perennial crops; ii) evolutionary changes related to new organ formation; iii) how to control uneven bud development and flowering; iv) how to provide male sterility for breeding; v) how to decrease early opening of flowers and the high level of abortion after anthesis.
Key genes related to the development of the floral organs were described through mutant analysis, especially in A. thaliana, which gave rise to the classic ABC model that explains flower architecture[147, 149−151]. In this model, homeotic genes are grouped as functional factors that act in combinatorial protein-protein interactions to form tetrameric complexes that specify the identity of the four floral whorls. In detail, the A-function are encoded by APETALA1 (AP1) and APETALA 2 (AP2), the B-function by APETALA3 (AP3) and PISTILLATA (PI), and the C-function by AGAMOUS (AG), and their single or combined action in each whorl results in the specification of floral organs: A in the first whorl specify sepals, whereas A+B in the second whorl determines the petals, B+C in the third whorl leads to the formation of the stamens and, finally, C in the fourth and most internal whorl specify the carpels. Subsequently, it was demonstrated that the SEPALLATAs (SEP1, SEP2, SEP3, and SEP4) act as connectors joining the ABC proteins throughout the K-box domain[152], adding a new factor named E-function and expanding the model to ABCE[153].
These cited genes are master regulators of transcriptional cascades that coordinate whorls development[154, 155] and, except for AP2, all of them belong to the MADS-box family of transcription factors, which is characterized by a MADS domain of 59 highly conserved amino acids[156]. This gene family displays a variable number of members in different species[156], which revealed an important evolutionary aspect of diversification, specifically studied for ABC-function homologs[153, 157]. Based on that, the diversification of the MADS-box family, represented by changes in the gene sequence, expression profile, and functions, could explain the morphological variability of flowers, which has been associated with the relative quick adaptation of Angiosperms to contrasting environments[158]. Despite its importance and the availability of coffee genomes[159, 160], few studies explored the MADS-box genes existing in Coffea sp. and their expression during development[13, 32, 33].
From a transcriptome database[161] followed by clone resequencing, 23 MADS-box full-length unigenes were identified in C. arabica by de Oliveira et al.[13] and classified into the described Arabidopsis subfamilies[162]. Interestingly, all MADS proteins found in coffee belong to type II (or MICK proteins) and the lack of type I members is suggested to be due to a low expression level or expression in very specific tissues and developmental stages[163]. The expression profile from 18 of these coffee MADS-box genes showed that the majority of these genes are expressed in both vegetative and reproductive tissues, with six genes being expressed exclusively in vegetative tissues[13]. In the same work, the closest relative homologs for the ABCE-function genes (CaAP1, CaAP3, CaPI, CaAG, CaSEP1/2, and CaSEP3) were also determined, along with their spatio-temporal expression profiles. For instance, CaAP1 (A-function) and CaAP3 (B-function) have similar expression patterns, with higher expression levels, which were restricted to sepals and only later in petals, in the first stages of floral development. In contrast, CaPI (B-function), CaAG (C-function), and CaSEP3 (E-function) are highly expressed during the late stages of floral development, with the expression of the first two genes being restricted to stamens and carpels.
The described expression patterns of ABC-function genes in Coffea sp. are following the conserved function of these genes, in which the B and C-function genes guarantee the internal reproductive primordia cells [153, 164] and compete with A-function genes that, in turn, preserve the external layer of cells to promote infertile organ formation of the perianth[147]. However, there are notable differences from the ABC model described in other species, suggesting functionalization of homeotic genes and/or participation of other genes to discriminate the perianth and reproductive organs in coffee flowers. Based on that, we reviewed the published expression profiles of coffee MADS-box genes to suggest their roles during reproductive development (Fig. 1) and propose an ABC model in Coffea sp. comparing it to Arabidopsis and a relatively closer species to coffee, tomato (Solanum lycopersicum) (Fig. 3). RNA and proteins are mobile signals that can act in different places than where they were produced[165], which could be a criticism of our proposal, however, it was already described that the expression of ABC genes follows the localization of their corresponding protein function[166].
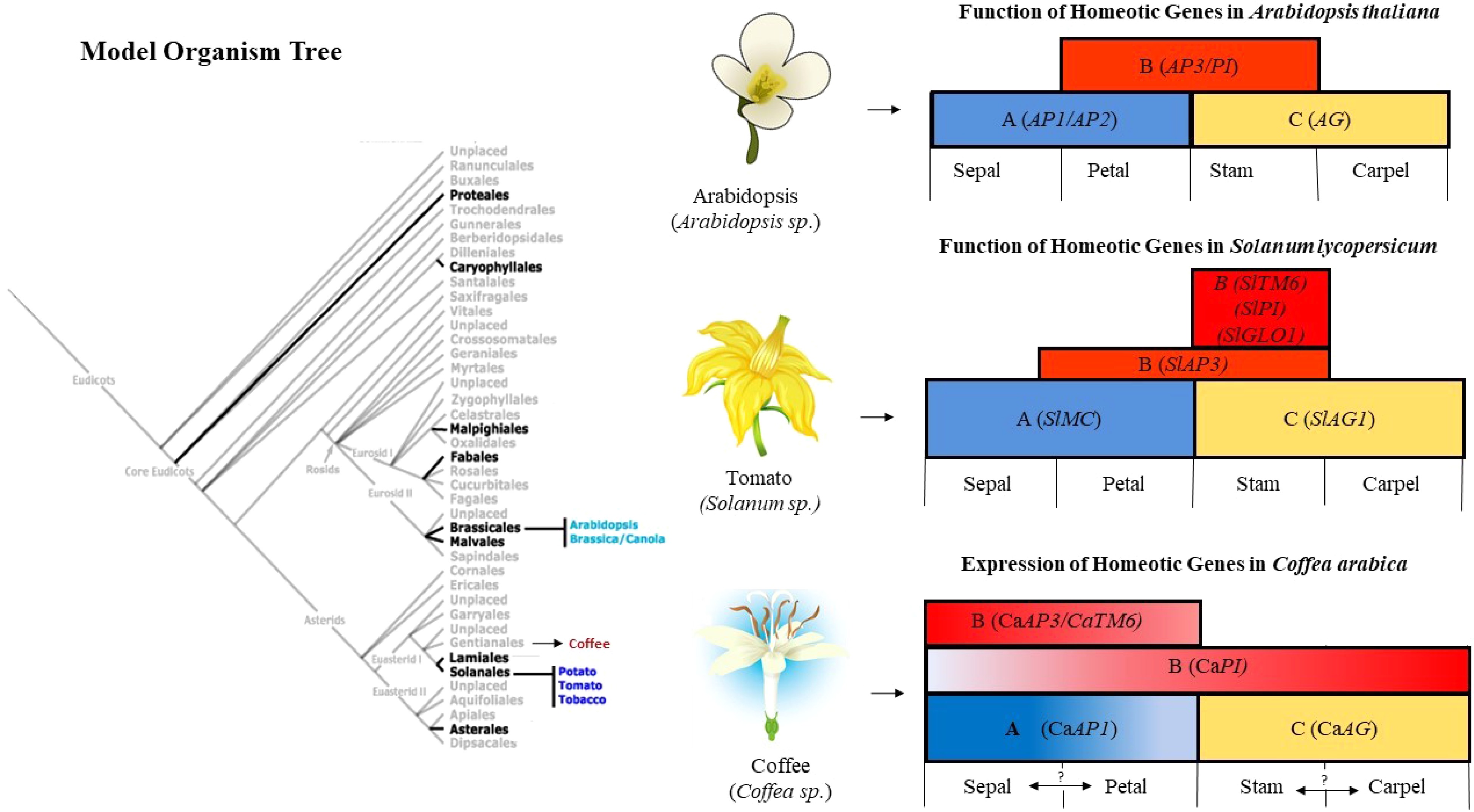
Figure 3. Proposed ABC model for floral organ identity in C. arabica. Left, phylogenetic classification of eudicots orders highlighting the model organisms (adapted from Geuten & Irish[167], which shows the relationship between the three species compared: Arabidopsis thaliana, C. arabica, and Solanum lycopersicum. A schematic representation of the respective flowers of the compared species is shown in the center. Right, variations of the ABC model explaining floral organ differentiation are represented. The ABC model of Arabidopsis thaliana guides the investigation of orthologous homeotic genes in flowering species, in which the variation in the ABC model may occur due to cumulative differences along evolutionary time in sequences, expression patterns, and/or combination of these genes for the specification of the four floral whorls. The ABC model for Arabidopsis thaliana and Solanum lycopersicum was built considering the functional analysis in mutant plants for specification floral organs[168−171]. For C. arabica, the ABC model is proposed based on the quantitative and qualitative gene expression[162]. The gradient color indicates the highest, lowest, or absent (white) expression of class A, B, and C genes in the initial, intermediate and final developmental phases of each floral whorl.
The floral development program is widely studied in model plants such as A. thaliana and tomato, a model for fleshy fruit plants[168, 170−173]. Class A and C homeotic genes are highly conserved among these organisms, also sharing an antagonism in their functions to specify whorls (Fig. 3). MACROCALYX (MC) of tomato is the closest homolog of AP1[172] showing a similar function, with their mutants completely converting the development of the sepals into leaf-like structures and petals not being specified[172, 173]. Whereas, it has been observed for C-function redundant expression patterns and functions among their representatives SHATTERPROOF1 (SHP1) and AG in Arabidopsis[174, 175], which is similarly reported for the respective tomato closest homologs SlAGL1 and SlAG1[174, 176]. Comparing to coffee plants, the CaAP1 (A-function) is expressed in the perianth, far from CaAG (C-function), whose expression is restricted to the primordia of stamens and carpels[13]. This is consistent with the conserved expression of C-function genes and its ancestral function related to the internal fertile organ development in other species, including gymnosperms[164, 177−179]. This spatial separation in the expression of these genes is in agreement with their antagonistic action, and competition for layers between A- and C-function genes, indicating that the A and C-factors in Coffea sp.
keep functional similarities to Arabidopsis and tomato homologs (Fig. 3). Nevertheless, an interesting characteristic of CaAP1 expression is its expression only at the late stages of petals development (gradient color in Fig. 3), after the emergence and separation of stamen primordia cells, suggesting a late A-function to provide fertile epipetalous stamens at the final stage of the flower developmental process[13]. Thus, because CaAP1 is expressed in both the primordia of sepals and petals, another homeotic gene seems to provide identity for coffee petals. FRUITFULL (FUL) is a close homolog of AP1, classified in the same SQUA subclade and known to display a redundant function, since it is involved in the floral development activation and perianth formation[162, 180]. The putative ortholog of FUL was described in C. arabica (CaFUL) and its expression is higher in flowers than in fruits[13], suggesting that it could be involved in petal identity. Other candidate genes for coffee petal specification are CaAGL6 and CaSEPs that make part of the conserved superclade SEP/AGL6/AP1 and may perform similar functions to AP1[181−183]. CaAGL6 and CaSEP3 are exclusively expressed in flowers and fruits, whereas CaSEP1/2 is expressed also in leaves and SAM[13], thus, it is possible that some functionalization has occurred in Coffea sp. Overall, these data suggest functional diversification of the SEP/AGL6/AP1 subfamily during coffee evolution and, accordingly, the A-function is very diverse and arguably so in angiosperms[184]. Functional studies of these genes is a promising field since it could be used to change the flowering time or to enhance floral activation of latent meristems providing more fruits, as well as, to explain the abnormal formation of petals and the premature opening, the 'starlet flowers' phenomenon[185], that appears to be induced by warming conditions[6, 70, 186].
B-function genes, represented by APETALA3 (AP3) and PISTILLATA (PI) in Arabidopsis, are related to the formation of petals and stamens and, accordingly, it is reported that AP3 and PI interact with each other forming functional heterodimers in the second and third whorls[187]. Surprisingly, in coffee plants, CaAP3 was expressed in sepals but not in stamens, with expression levels increasing progressively throughout floral development. On the other hand, CaPI expression was found in petals and stamens[13], in agreement with its conserved function and expression in Arabidopsis and tomato (Fig. 3), but also in colleters. These results do not explain how stamens and carpels are differentiated in coffee plants since these organs show similar expression patterns of CaPI and CaAG (Fig. 3; de Oliveira et al.[13]). Duplication events are reported for the DEF/AP3 subclade, which originated the divergent paralogous lineage TOMATO MADS BOX GENE6 (TM6)[188]; related to functional diversification out of Brassicaceae[187, 189]. For example, euAP3 and PI of Arabidopsis and its respective orthologs in tomato, SlGLO1[190] and SlPI[190], control the specification of petals and stamens in both species[190]. However, the silencing of these two genes in tomatoes does not affect the identity of the petals[168] and the complete differentiation of stamens requires the action of TM6, a sub-functionalization that is not present in Arabidopsis[172]. Similarly, this duplication event and the closest CaTM6 homolog was reported in C. arabica with its expression being found exclusively in reproductive tissues and at higher levels during the final stages of floral development[13]. However, CaTM6 expression was detected in the perianth, a similar pattern of CaAP1[13], suggesting a more complex regulation for coffee whorls differentiation and epipetalous stamens (Fig. 3).
Based on that, compensatory changes in B-function genes may have occurred in the Rubiaceae family with the participation of other players, non-described MADS-box genes and/or miRNAs, as suggested for Solanaceae[168], which could result in functional evolution and the novel structures found in coffee flowers.
-
Considering the ever-growing demand for higher quality coffees and the threat of climate change, efforts should be urgently taken to better understand the coffee flowering process at the molecular level, since it can directly affect coffee yield and cup quality. As shown in this review, advances in the hormonal regulation of coffee flowering, particularly on ACC and ethylene's involvement during anthesis, reveal a new avenue for better comprehending the flowering process, as well as to manipulate it through the use of growing regulators. With the availability of coffee genomes and RNAseq global transcriptomics, which allows comparisons between different tissues, developmental stages, and growth conditions, a better picture of the molecular regulation of the coffee phenological cycle and floral development also starts to take place. Future studies should be dedicated to identify all coffee MADS-box genes, as well as other possible players involved in floral development and whorls specification, along with their functional analysis using model species, easier to be transformed and manipulated. These approaches will certainly improve the knowledge towards the flowering process in such important species, enabling the manipulation of processes such as flowering induction and anthesis and potentially contributing to a better coffee cup quality.
- We want to thank the "Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)", Honduran Foundation for Agricultural Research (FHIA), and "Programa de becas Honduras 2020" for the financial support of the first author. We also thank the "Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)", the "Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)", and the "Instituto Brasileiro de Ciência e Tecnologia do Café (INCT/Café)", under FAPEMIG grant (CAG APQ 03605/17) for financially supporting the experiments.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
López ME, Santos IS, de Oliveira RR, Lima AA, Cardon CH, et al. 2021. An overview of the endogenous and environmental factors related to the Coffea arabica flowering process. Beverage Plant Research 1: 13 doi: 10.48130/BPR-2021-0013

An overview of the endogenous and environmental factors related to the Coffea arabica flowering process
- Received: 03 August 2021
- Accepted: 25 October 2021
- Published online: 18 November 2021
Abstract: The transition of vegetative meristems to a reproductive state followed by floral development and flowering is a complex process regulated by many regulatory pathways that integrate (epi) genetic and environmental factors to guarantee the reproductive success in angiosperms. In woody and perennial species, such as coffee trees, the extended life cycle reflects adaptive strategies, i.e., two-year cycle, sequential flowering and bud latency during the winter, that ensure reproductive success in a variable environment. The dynamics of coffee flowering integrate multiple stimuli to induce morpho-physiological changes, especially during the reproductive phase. We discussed diverse stimuli which are directly and indirectly associated with molecular pathways related to the reproductive meristem transition and floral development until anthesis. Here, we propose to redefine some concepts regarding coffee floral development, such as: i) a summary of genes possibly involved in the flowering pathways; ii) a new classification of buds by position in the node (B1 - B4) to avoid misunderstandings with the uneven developmental stages; iii) comparative ABC model to Arabidopsis and tomato to allow evolutionary discussions of whorls development in Coffea sp. From this, we expect to collaborate to drive future studies towards coffee breeding and production, for example, improving floral bud activation, controlling the flowering time, anthesis synchronization to produce more homogeneous ripening fruits, reducing abortion or early flower openings, and increasing fruitification. Moreover, we think that the explored relationship between physiological and molecular approaches during the coffee flowering may be extended to understand other tropical perennial species, especially those with asynchronous flowering behavior, and to understand evolutive aspects in Rubiaceae.