










-
Tea plant [Camellia sinensis (L.) O. Kuntze] is an important perennial evergreen economic crop, widely cultivated worldwide, which provides raw material for the production of the non-alcoholic beverage 'tea'[1]. Tea contains a large number of secondary metabolites beneficial to human health, such as catechins, theanine and polysaccharides[2]. Lignin is an aromatic polymer mainly deposited in secondary thickened cells[3]. Lignin provides the necessary mechanical strength for plant cells and tissues, contributes to the transport of water and nutrients in plants, and responds to various biotic or abiotic stresses[4, 5]. Excessive lignin accumulation was negatively correlated with tenderness. The tender age of tea leaves is an important basis for judging the quality of tea. The degree of lignification reflects the tenderness of tea leaves and determines their quality[6]. Lignin is one of the main products of the phenylalanine metabolic pathway. The genetics of lignin biosynthesis has been extensively studied in many plants. Regulatory genes encoding transcription factors (TFs) control the transcription of structural genes encoding enzymes involved in lignin biosynthesis[7, 8].
WRKY is one of the most important transcription factors, regulating the biological processes of plant stress defense, development and metabolism[8−12]. WRKY proteins have a WRKY domain with about 60 amino acids, which consists of the conserved amino acid sequence WRKYGQK and a CX4-5CX22-23HXH zinc finger-like binding motif. WRKY specifically binds to the DNA sequence motif TTTGACT/C, called W-box[13].
Existing evidence indicates that WRKY transcription factors play key roles in the regulation of lignin, cell wall biosynthesis, and in the adaptation of plants to biotic and abiotic stresses[14]. Overexpression of rice WRKY89 gene promotes lignification of stems[14]. Four Medicago WRKY transcription factors were overexpressed in tobacco plants, resulting in an increase of phenolic compounds and lignin levels in transformed leaves. The results indicated that these WRKY transcription factors were involved in the lignification process[15]. The ectopic expression of VvWRKY2 in tobacco plants caused drastic modifications in the lignin composition, especially the S/G ratio, and altered the organization of vascular structure[16]. The mutation of WRKY13 leads to a weaker stem phenotype in Arabidopsis thaliana, indicating that WRKY13 has a positive regulation effect on stem development[8]. Although WRKY transcription factors have been isolated and characterized in many plants, research on WRKY transcription factors in the lignin biosynthesis of tea plants is currently limited.
In this study, a WRKY member gene CsWRKY13 was isolated from tea plant 'Longjing 43', which is one of the most widely planted tea cultivars in China and has many excellent characteristics, such as cold resistance, early germination, and good taste[17]. Expression analysis showed that the CsWRKY13 gene was highly expressed in the stem. Subcellular localization analysis, transcriptional activity analysis, morphological and physiological characterization were performed. Overexpression of CsWRKY13 in Arabidopsis was carried out to further identify its function. These results will further deepen the understanding of CsWRKY13 on the regulation of biosynthesis of lignin, and provide a basis for future researchers on the pathway of lignin in tea plant.
-
Two-year-old tea plant 'Longjing 43' cut seedlings were grown under natural conditions. Arabidopsis were grown in a chamber at the Tea Science Research Institute, College of Horticulture, Nanjing Agricultural University (Nanjing, China). The conditions are as follows: 12-h light (200 μmol·m−2 s−1)/12-h darkness, 22/18 °C day/night, and 70% relative humidity. To analyze the expression of CsWRKY13 in different tissues, the first leaf, the third leaf, the fifth leaf, the immature (green) stem, mature (red) stem, and the flowers were sampled, respectively. The collected samples were immediately immersed in liquid nitrogen and stored at −80 °C for subsequent experiments.
RNA extraction and cDNA synthesis
-
The extraction of total RNA from tea plant samples was completed using the RNA Isolation Kit (Huayueyang, Beijing, China), as per the manufacturer's instructions. The purity and quality of total RNA were detected by NanoDrop ND 2000 Spectrophotometer (Thermo Scientific, USA) and denaturing agarose gel. PrimeScript RT reagent Kit (TaKaRa, Dalian, China) was used to reverse transcribe the RNA to cDNA.
Cloning and bioinformatics analysis of CsWRKY13 gene in tea plant
-
The amino acid sequence of Arabidopsis AtWRKY13 was used as the query in tea plant genome to retrieve CsWRKY13 (ID: CSA007816.1)[18]. Gene specific primers (forward: 5’-ATGCTCAACCAGGGGCTGTTTGA-3’ and reverse: 5’-CTACCAGAAGAAATTATTAAGTT-3’) were designed using Primer Premier 6 software to amplify CsWRKY13 for RT-PCR. CsWRKY13 was cloned from the leaves of 'Longjing 43' using Ex Taq Mix (TaKaRa, Dalian, China). The PCR products were connected to the pMD19-T vector and transformed into Escherichia coli DH5α. Finally, positive clones were selected and sequenced in Genscript (Nanjing, China).
Homologous search and conserved domain prediction were performed by BLASTp program from the NCBI website (http://blast.ncbi.nlm.nih.gov/Blast.cgi). Sequence multiple alignments were performed by DNAMAN. MEGA 6.0 software was used to construct the phylogenetic tree[19].
Subcellular localization of CsWRKY13
-
The CsWRKY13 gene lacking the termination codon was amplified by PCR using specific primers (forward: 5’-CACCATCACCATCACGCCATGATGCTCAACCAGGGGCTGTTTGA-3’ and reverse: 5’-CACTAGTACGTCGACCATGGCCCAGAAGAAATTATTAAGTT-3’). The target fragment was recovered and inserted into the expression vector pA7 containing green fluorescent protein (GFP) to construct a CaMV 35S: CsWRKY13-GFP fusion expression vector. Subsequently, the recombinant plasmid (CsWRKY13-GFP) and control vector (pA7-GFP) were transferred into onion epidermal cells using the particle bombardment (PDS-1000, Bio-Rad) method. After bombardment, onion epidermal cells were cultured on Murashige and Skooog (MS) medium for 16 h in the dark at 25 °C. The expression of CsWRKY13 in onion epidermal cells was observed using confocal laser confocal scanning microscopy (Zeiss LSM 780, Germany).
Transcriptional activation analysis of CsWRKY13
-
CsWRKY13 was amplified using the specific primers containing EcoR I and Sal I recognition site (forward: 5’-ATGGCCATGGAGGCCGAATTCATGCTCAACCAGGGGCTGTTTGA-3’ and reverse: 5’-ATGCGGCCGCTGCAGGTCGACCTACCAGAAGAAATTATTAAGTT-3’) and then inserted into the yeast expression vector pGBKT7 (as the negative control). The vector pCL1 (containing full length copy of GAL4) was used as a positive control. The constructs were introduced into yeast strain Y2H. The pCL1 transformants were selected on a SD/-Leu medium, while the pGBKT7 and pGBKT7-CsWRKY13 were incubated on a SD/-Trp medium for three days at 30 °C and then the transgenic cell lines were transferred to a SD/-His-Ade medium containing the presence or absence of X-α-Gal.
Construction of overexpression vector and transformation of A. thaliana
-
Specific primers (forward: 5’-TTTACAATTACCATGGGATCCATGCTCAACCAGGGGCTGTTTGA-3’ and reverse: 5’-ACCGATGATACGAACGAGCTCCTACCAGAAGAAATTATTAAGTT-3’) was used to amplify CsWRKY13 gene. The PCR product was inserted into the pCAMBIA1301 vector containing the CaMV 35S promoter, and then the recombinant vector was introduced into Agrobacterium tumefaciens strain GV3101 using the electroporation transformation method. Floral dip method was performed for transforming into Arabidopsis[20]. The obtained seeds were sterilized and screened on MS medium containing 50 mg/L hygromycin. The selected Arabidopsis plants were stained with GUS staining and amplified with CsWRKY13 gene specific primers[21]. Three independent transgenic Arabidopsis lines were randomly selected for subsequent experiments.
Extraction and determination of lignin
-
The leaves and stems of the transgenic and wild-type (WT) Arabidopsis plants grown for 5 and 7 weeks were sampled, quickly placed in liquid nitrogen, and stored at −80 °C for RNA extraction and lignin content determination. Lignin was extracted and determined by previous methods[22, 23]. Each experiment had three biological replicates.
Histochemical staining and UV microscopy
-
Histochemical staining with safranin-O was performed to analyze the lignin distribution in WT and in transgenic Arabidopsis leaves and stems[24]. Safranin-O stained the lignified sections red; fast green stained the cellulosic tissues green. Lignin accumulated in plant tissues can spontaneously fluoresce under ultraviolet irradiation (UV). The structures of leaves and stems were observed under a fluorescence microscopy emitting UV. The fluorescence pictures were produced with a charge coupled device (CCD) camera.
Gene expression assays
-
The RT-qPCR was used to detect the expression level of CsWRKY13 in different tissues of tea plants. The quantitative primer for CsWRKY13 is forward: 5’-GCTCTGGCTGTCATCATCAATC-3’ and reverse: 5’-GGACCTCCCAAATGTGTAGTGA-3’. The CsGAPDH gene was used as an internal reference gene[2]. The transcription levels of lignin and flavonoid-related structural genes and CsWRKY13 were detected in leaves of wild-type and transgenic Arabidopsis. AtActin2 was used as an internal reference gene to normalize the expression of Arabidopsis related genes. Primer used in the experiments refer to Raes et al. and Xu et al. as shown in Supplemental Table S1[25, 26].
The RT-qPCR was performed using a Bio-Rad CFX96TM fluorescent quantitative PCR instrument by SYBR Premix Ex Taq. The amplification procedure was: 95 °C for 30 s, followed by 40 cycles at 95 °C for 10 s, 60 °C for 30 s, and melting curve analysis at 65 °C for 10 s. Three biological replicates were set up for each experimental sample and the relative gene transcription levels were analyzed using the
$2^{-\Delta \Delta} $ Cᴛ method[27].Statistical analysis
-
Excel 2010 and IBM SPSS Statistics 22.0 software were used for data collation and statistical analysis. Significant differences between values were determined with Duncan’s multiple range test (p < 0.05) and one-way ANOVA based on student’s t-test, and are indicated by asterisks (*, p < 0.05; **, p < 0.01; ***, p < 0.001).
-
The full-length cDNA sequence of the CsWRKY13 was isolated from tea plants (C. sinensis cv. Longjing 43). The results showed that the open reading frame of CsWRKY13 was 708 bp, encoding 235 amino acids. The sequence alignment showed that CsWRKY13 of tea plants had a high similarity with the WRKY13 protein sequence of other plants (Fig. 1). These WRKY13 proteins contains a conserved domain composed of about 60 amino acids at the C-terminal, which contains a 'WRKYGQK' motif and a 'CX4CX23HXH' C2H2 zinc finger structure. In addition, the C terminal of these proteins also contains a nuclear localization signal (NLS) domain.

Figure 1. Multiple alignment of the amino acid sequences of CsWRKY13 and other WRKY13 proteins of different species. The conserved DNA-binding domain is indicated by a black line. The comparator species were: Populus euphratica (PeWRKY13, XP_011025576.1), Theobroma cacao (TcWRKY13, XP_007048165.1), Gossypium hirsutum (GhWRKY13, NP_001314117.1), Arabidopsis thaliana (AtWRKY13, NP_195651), Vitis vinifera (VvWRKY13, RVW55274.1).
A phylogenetic tree of CsWRKY13 and WRKY proteins from other plants was constructed using MEGA 6.0 software with the neighbor-joining method. According to the classification method previously reported, all WRKY proteins used were derived from group I and group II[28−29]. Among them, AtWRKY12 and AtWRKY13 belong to the group II, and AtWRKY3, 4 and 25 belong to group I. Members of groups I and II all contain a conserved WRKY domain at the C-terminal end along with a C2-H2 zinc-finger-like motif, which are the unique zinc ligands. This study showed that CsWRKY13 was closely related to WRKY13 proteins of group II of other species, and was most closely related to PeWRKY13 of Populus euphratica (Fig. 2).

Figure 2. Phylogenetic tree analysis of CsWRKY13 protein and other WRKY proteins of different species. Other plant WRKY proteins used for multiple sequence alignment and evolutionary analysis include: A. thaliana (AtWRKY3, At2g03340; AtWRKY4, AF425835; AtWRKY12, At2g44745; AtWRKY13, AF421153; AtWRKY25, At2G30250); P. euphratica (PeWRKY13, XP_011025576.1); T. cacao (TcWRKY13, XP_007048165.1); G. hirsutum (GhWRKY13, NP_001314117.1); V. vinifera (VvWRKY2, AY596466; VvWRKY13, RVW55274.1); Medicago truncatula (MtSTP, AEJ09956.1); Oryza sativa (OsWRKY24, AY676925; OsWRKY78, BK005212).
Expression analysis of CsWRKY13 gene in different tissues of tea plant
-
The expression levels of CsWRKY13 gene in different tissues of tea plants were detected and analyzed by RT-qPCR. The results showed that the CsWRKY13 gene was expressed in all test tissues of tea plants, and it was highly expressed in mature stems, followed by immature stems (Fig. 3). The expression level of the CsWRKY13 gene was low in tea leaves, and decreased gradually with the increase of tea leaf maturity.
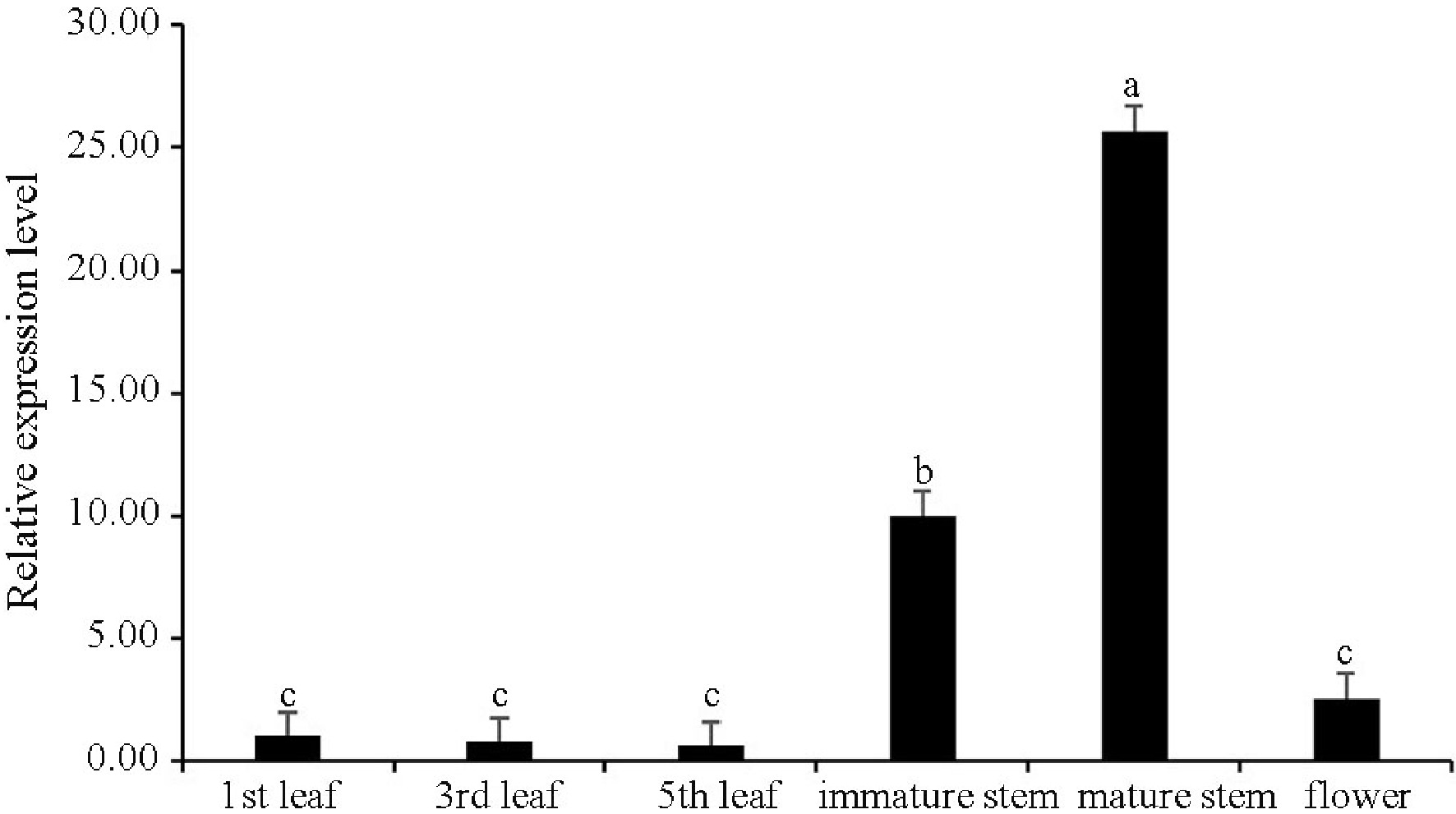
Figure 3. Expression profiles of CsWRKY13 in different tissues of tea plants. The value in the first leaf was set to 1. Expression levels of CsWRKY13 were detected by RT-qPCR, and the data were analyzed using the
$2^{-\Delta \Delta} $ Cᴛ method. Different letters represent significant difference at the 0.05 level.Subcellular localization of CsWRKY13 protein
-
To study the subcellular localization of CsWRKY13 protein, the recombinant plasmid CsWRKY13-GFP and control pA7-GFP were transferred into onion epidermal cells by particle bombardment. The GFP fluorescence signal showed that the control plasmid was expressed in the entire cell, while recombinant plasmid CsWRKY13-GFP was only observed in the nucleus of onion cells (Fig. 4). The above results indicate that CsWRKY13 is localized in the nucleus.
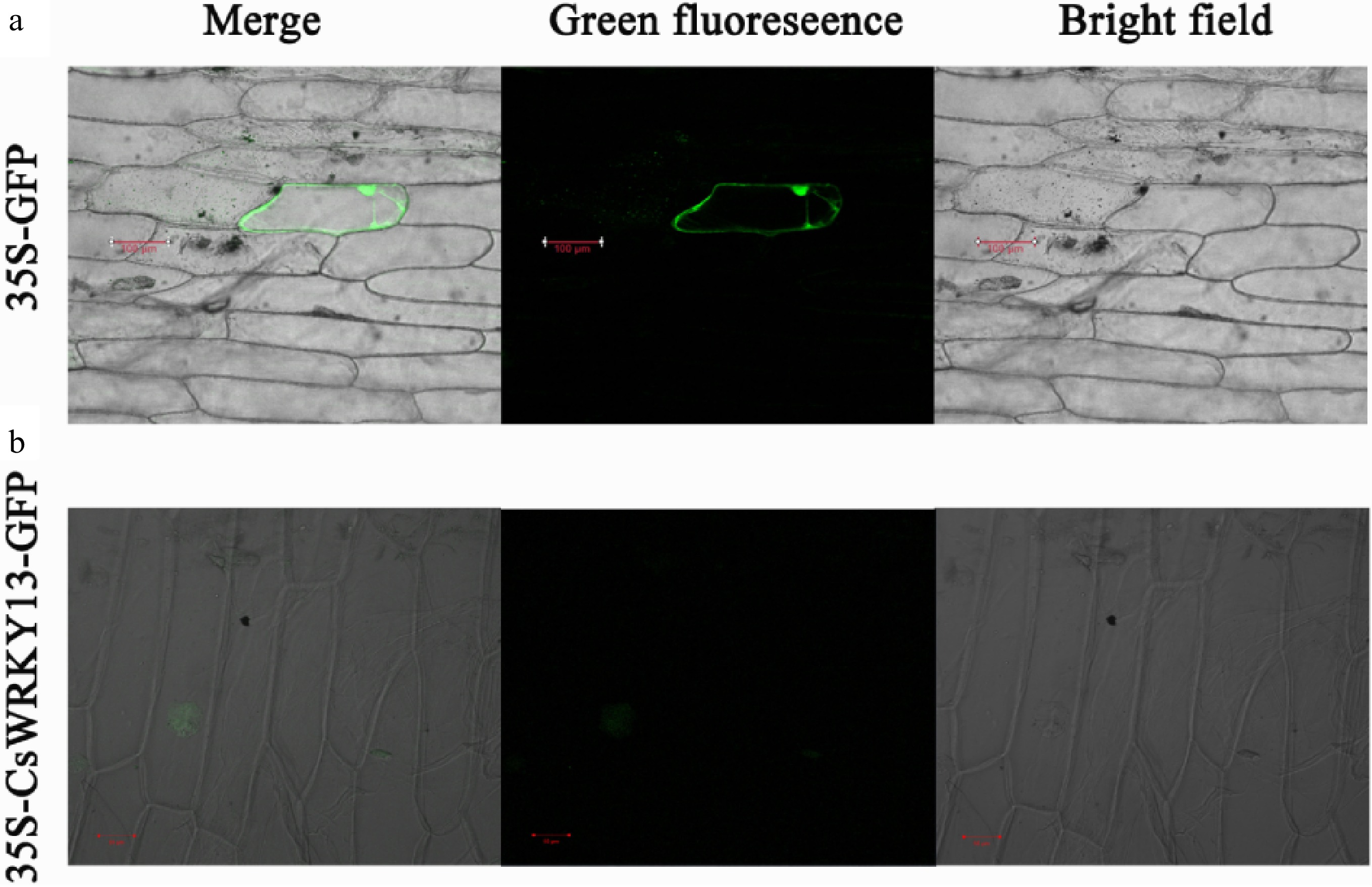
Figure 4. Subcellular location of CsWRKY13 proteins in onion epidermal cells.
Transcriptional activation of CsWRKY13
-
In order to determine the trans-activation capacity of CsWRKY13, CsWRKY13 was fused to the GAL4-binding domain and expressed in yeast strain Y2H. The yeast strain with a complete GAL4 domain (pCL1) and pGBKT7-CsWRKY13 were able to grow on SD/-His-Ade medium; in contrast, the negative control strain with the pGBKT7 vector was able to grow on SD/-Trp medium, but not on the SD/-His-Ade medium. As shown in Fig. 5, the pigmentation induced when the medium contained X-α-Gal. The result showed that CsWRKY13 had no transcriptional activation activity in yeast cells in vitro.
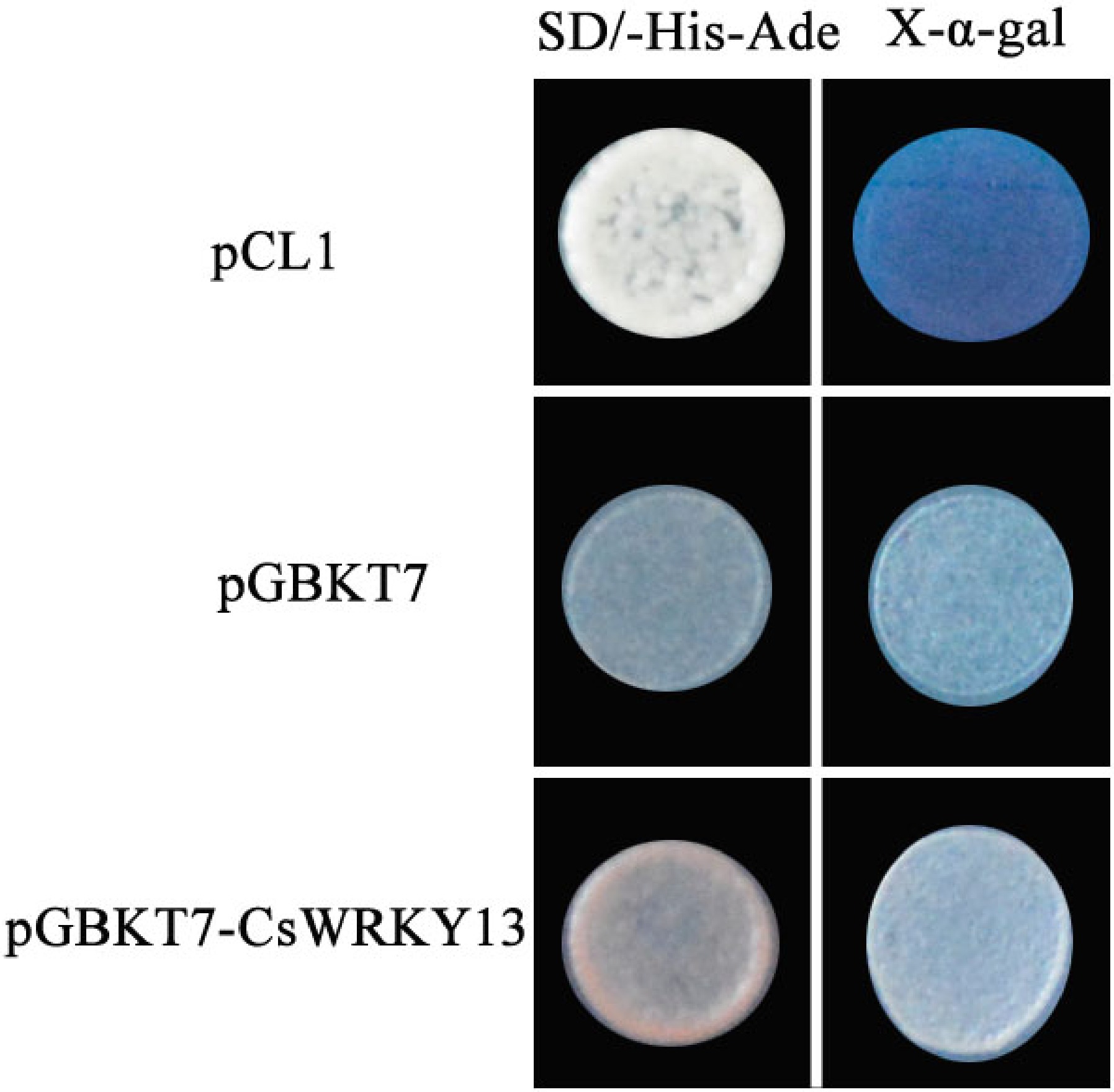
Figure 5. Trans-activation activity analysis of CsWRKY13 in yeast.
Overexpression of CsWRKY13 in Arabidopsis
-
Three CsWRKY13 transgenic Arabidopsis lines (OE1, OE2 and OE3) were screened to investigate the function of the CsWRKY13 gene. Wild-type and transgenic plants of Arabidopsis were cultured on medium for a week and stained with GUS dye solution. As shown in Fig. 6a, three transgenic Arabidopsis plants hosting CsWRKY13 gene were stained blue. The transgenic plants were further confirmed by PCR amplification and RT-qPCR (Fig. 6b & 6c).
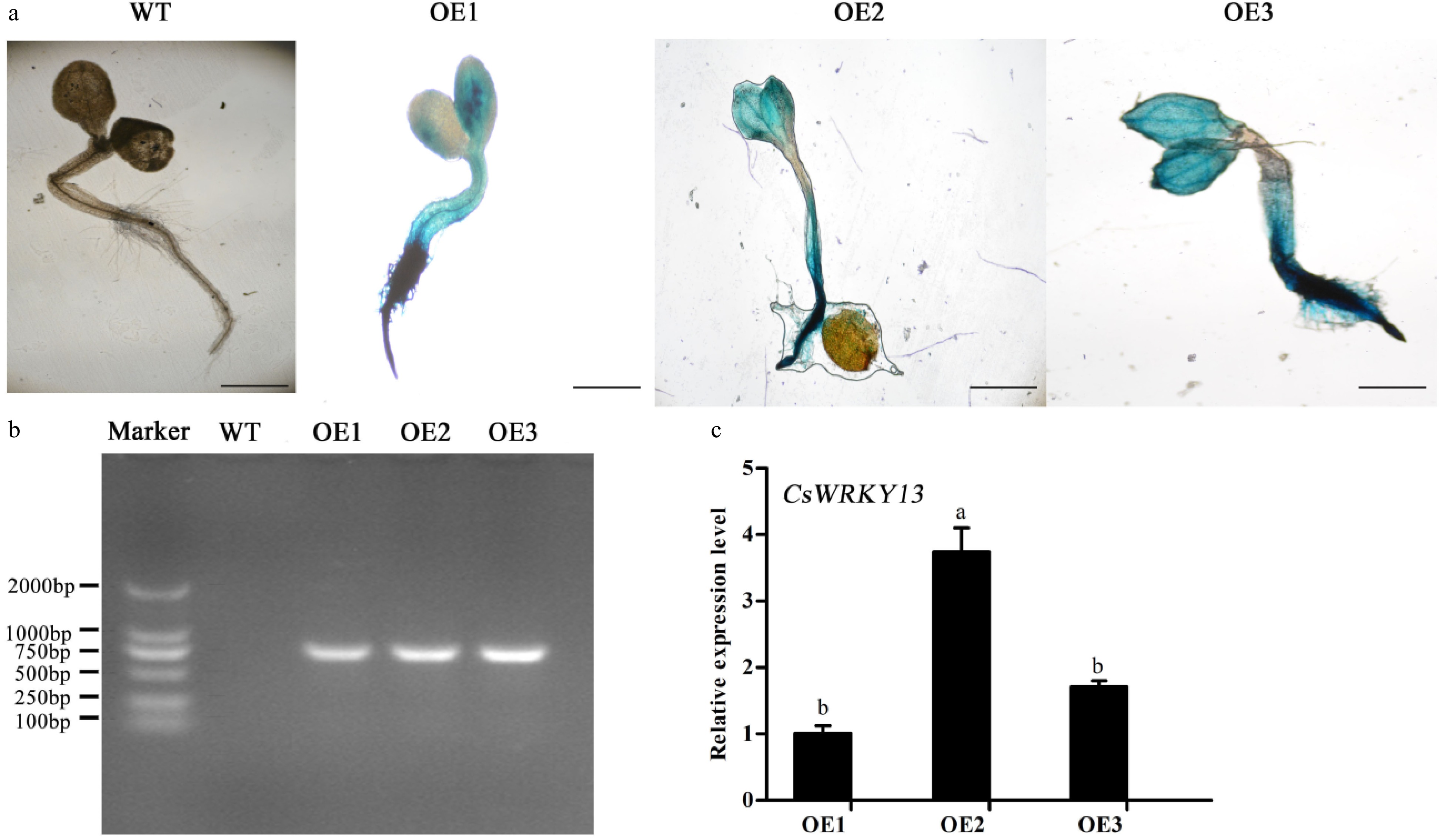
Figure 6. Confirmation of transgenic Arabidopsis carrying the CsWRKY13 gene by GUS staining and PCR assays. (a) GUS assays of WT and transgenic Arabidopsis. Bar: 1 mm. (b) PCR analysis of CsWRKY13 of WT and transgenic Arabidopsis. (c) The relative expression levels of the CsWRKY13 gene.
Overexpression of CsWRKY13 reduced lignin accumulation in transgenic Arabidopsis leaves
-
There was no significant phenotypic difference between transgenic and WT Arabidopsis at 5 weeks (Fig. 7a). Compared with WT, the lignin content in leaves of OE1, OE2 and OE3 transgenic Arabidopsis lines decreased by 5.2%, 32%, and 27%, respectively (Fig. 7b). Histochemical staining showed that lignin was mainly deposited in the xylem of leaves of Arabidopsis plants, but there was no significant difference in the lignin degree of transgenic and WT Arabidopsis plants (Fig. 7c). The UV-excited fluorescence in the leave of transgenic and WT Arabidopsis plants are shown in Fig. 7d. Lignin autofluorescence was detected in the xylem, which is consistent with the finding in Fig. 7c.
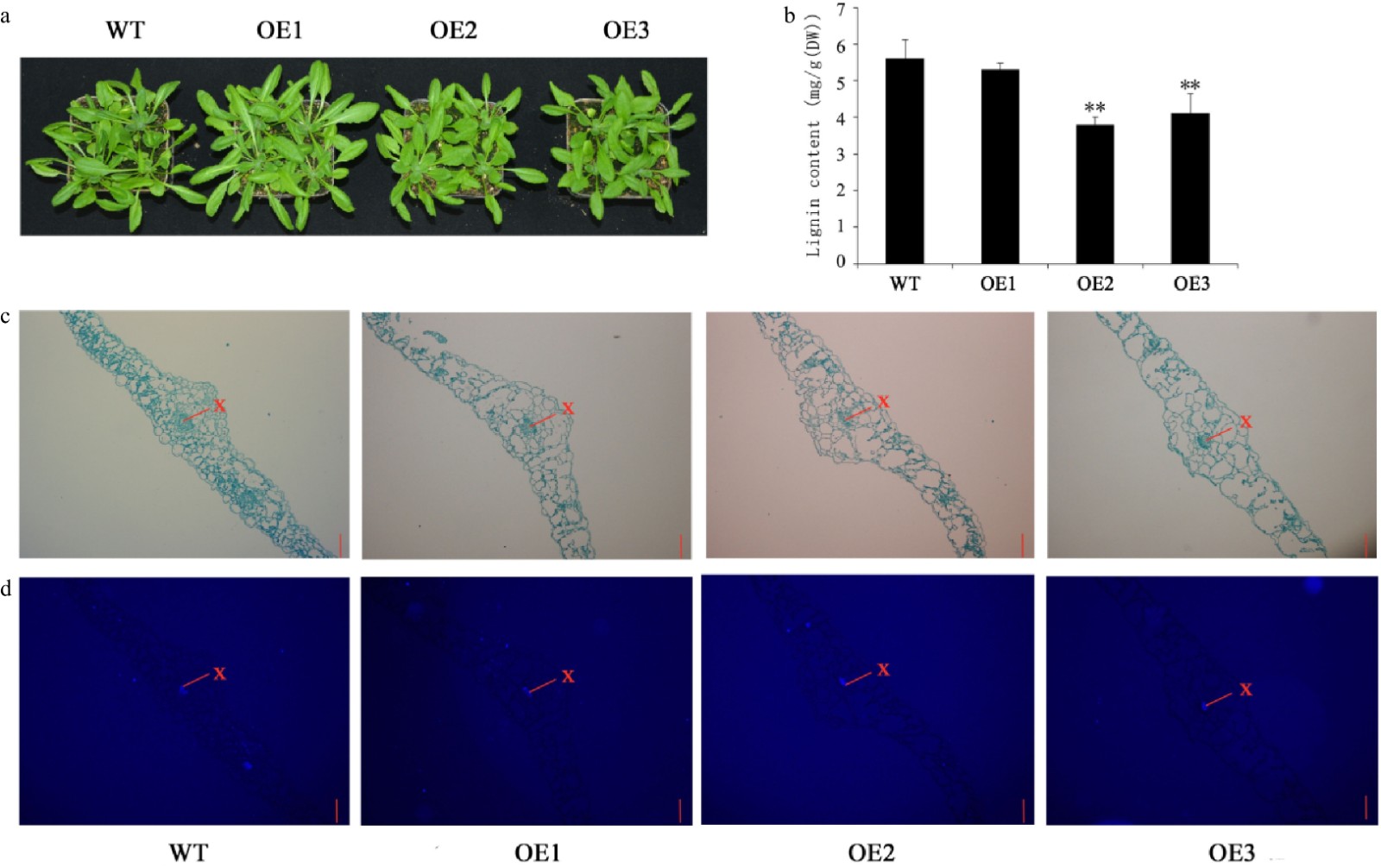
Figure 7. Growth status and lignin content of 5-week-old Arabidopsis. (a) Growth status of WT and transgenic Arabidopsis. (b) Lignin content of leaves. (c) Safranin-O staining. (d) Fluorescence micrographs. Scale bars = 50 μm. The red lines indicate the xylem. Error bars represent standard deviation (SD). (**) indicates the significance of the difference (p < 0.01).
Overexpression of CsWRKY13 reduced lignin accumulation in transgenic
Arabidopsis stems -
The results showed that OE1, OE2 and OE3 displayed a weaker stem phenotype at 7 weeks (Fig. 8a). Compared with WT, the lignin content of OE1, OE2 and OE3 transgenic Arabidopsis stems decreased by 3.1%, 2% and 7.1%, respectively (Fig. 8b). Histochemical staining showed that lignin was mainly deposited in the secondary cell wall of the xylem, and there was no significant difference in the degree of stem lignification between the transgenic plants and WT plants (Fig. 8c). UV-excited autofluorescence showed similar results (Fig. 8d).

Figure 8. Growth status and accumulation of lignin in 7-week-old WT and CsWRKY13 transgenic A. thaliana. (a) Growth status of WT and transgenic Arabidopsis. (b) Lignin content of stems. (c) Safranin-O staining. (d) Fluorescence micrographs. Scale bars = 50 μm. The red lines indicate the xylem. Error bars represent standard deviation (SD). (*) indicate that the value is significantly different from that of the WT at the same time point (* p < 0.05; ** p < 0.01; *** p < 0.001).
Gene expression of lignin, flavonoids metabolism pathway
-
The effect of the CsWRKY13 gene on the transcription level of transgenic Arabidopsis lignin synthesis gene was detected by RT-qPCR. Due to substrate competition between lignin pathway and flavonoid pathway, we also examined whether the CsWRKY13 gene affected the transcription of flavonoid structural genes in transgenic Arabidopsis hosting CsWRKY13 gene. The results showed that the transcription levels of three phenylpropanoid pathway genes AtPAL, AtC4H and At4CL were significantly decreased in three transgenic Arabidopsis lines (Fig. 9a). The transcriptional levels of lignin-specific pathway structural genes AtHCT, AtCCoAOMT, AtCOMT, AtCCR, AtF5H, AtCAD4 and AtCAD6 also decreased, among which AtCAD4 and AtCAD6, two cinnamyl alcohol dehydrogenase genes, showed the largest decrease (Fig. 9b). The transcription levels of most flavonoid structural genes AtF3H, AtF3'H, AtDFR, AtLDOX and AtUGT78D2 were up-regulated in OE2 and OE3 transgenic lines (Fig. 9c). The expression in transgenic OE1 was not completely consistent with that of OE2 and OE3. In addition to F3’H, DFR, LDOX, lignin synthesis gene AtCAD6 was reduced only about five times in OE1, but nearly 100 times in OE2 and OE3. The reasons may be complicated and need further study.
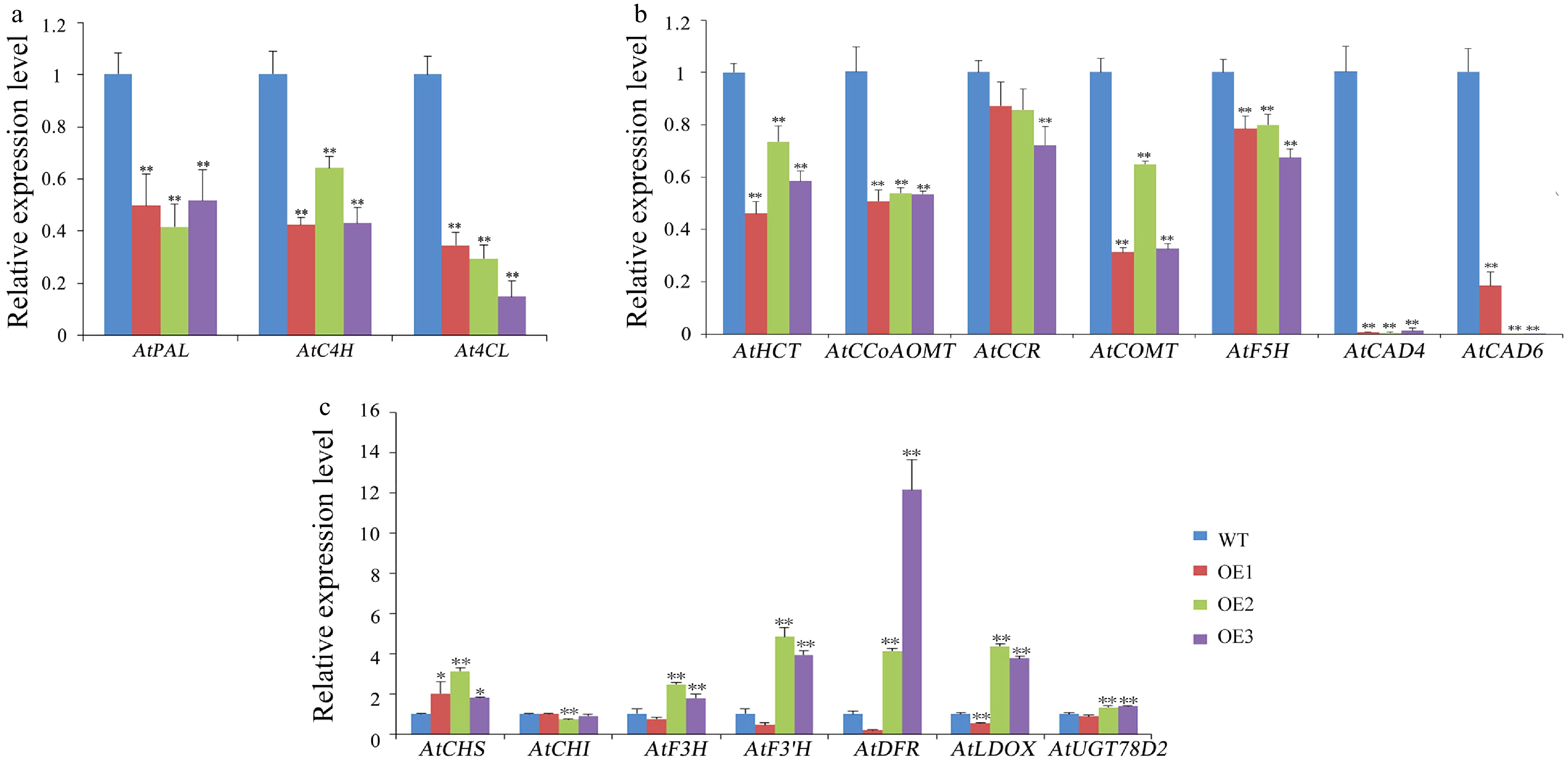
Figure 9. Expression levels of lignin and flavonoids pathway genes and secondary-wall-related genes in WT and transgenic Arabidopsis. (a,b) Expression levels of lignin pathway genes in WT and transgenic Arabidopsis. (c) Expression levels of flavonoids pathway genes in WT and transgenic Arabidopsis. Error bars represent standard deviation (SD). (*) indicate that the value is significantly different from that of the WT at the same time point (* p < 0.05; ** p < 0.01; *** p < 0.001).
-
Lignin is a secondary metabolite with biological functions in plants, playing an important role in maintaining the normal growth and development of plants and responding to external stimuli. Plants can enhance their resistance to abiotic or biotic stress by increasing lignin content. For example, water stress can significantly improve the content of lignin in carrot roots[30]. Changes in lignin content in leaves and roots of Phoenix dactylifera were caused by brittle leaf disease[31]. Lignin is essential for plant growth and development, while excessive accumulation of lignin will have negative effects on crops. For example, increased lignin content will affect the taste and quality of carrot, okra and celery[32−37].
The biosynthesis of lignin involves many enzymes and is influenced by complex transcriptional regulation. WRKY transcription factors are widely involved in plant development and regulate plant stress resistance[38, 39]. Recent studies have shown that WRKY transcription factor can also regulate the accumulation of lignin by regulating the transcription level of lignin biosynthesis structural genes. As a cash crop for leaf use, the degree of lignification of fresh tea leaves will affect the tenderness and quality of tea. The screening and identification of WRKY transcription factors related to lignin accumulation in tea plants is of great significance to regulate lignin content, improve the tenderness of tea leaves, and thus improve tea quality.
Lignin content is usually higher in stem tissues with higher lignification. Previous studies have shown that the AtWRKY13 gene is involved in stem tissue development and regulates lignin biosynthesis[8]. Grape VvWRKY2 gene is highly expressed in young woody stems, and overexpression in tobacco changes the expression level of lignin biosynthesis related genes and affects cell wall formation[16]. Yang et al. identified a PtrWRKY19 gene from poplar, which is highly expressed in the stem tissue of poplar and acts as a transcription suppressor to negatively regulate lignin biosynthesis[40]. In this study, CsWRKY13 gene was highly expressed in the stem tissues of tea plants. Heterologous expression of CsWRKY13 inhibited lignin synthesis in Arabidopsis, suggesting that CsWRKY13 may act as a transcription suppressor to negatively regulate the lignin synthesis in transgenic Arabidopsis. This is inconsistent with the research on the WRKY13 gene in Arabidopsis. Li et al. found that the Arabidopsis wrky13 mutant inhibited transcription of genes related to lignin biosynxthesis[8]. Wang et al. found that interfering with AtWRKY12 transcription promotes lignin biosynthesis[41]. MlWRKY12, as a homologous gene of AtWRKY12, shows similar functions to AtWRKY12 and inhibits lignin biosynthesis[42]. In Arabidopsis, AtWRKY12 is a transcriptional repressor, whereas AtWRKY13 shows transactivation activity. Li et al. suggested that these two antagonistic genes may regulate different subregions of stems, and that antagonism may be a mechanism that restricts the allocation of wasted carbon to stem cells that are not necessary to support the plant to resist gravity. CsWRKY13, an ortholog of AtWRKY13, which has a similar function to AtWRKY12. This is worthy of further research, whether there are other homologs of CsWRKY13 that play vital roles in lignin synthesis.
There is potential substrate competition between lignin and flavonoid pathways, which share the phenylpropane metabolic pathway. In order to explore whether CsWRKY13 affects the flavonoid metabolism pathway, the expression patterns of flavonoid structural genes were also analyzed. The results showed that CsWRKY13 upregulated the transcription of most flavonoid pathway genes in Arabidopsis, suggesting that CsWRKY13 may also regulate the flavonoid pathway. Catechins are the main flavonoid metabolites of tea plants, accounting for 12%−24% of the dry weight of tea leaves. However, Arabidopsis lacks the key genes for catechin synthesis, resulting in almost no catechin synthesis. So far, transgenic tea plants have not yet matured. In the future, we should further verify the function of CsWRKY13 using tea plant transgenic system.
-
WRKY transcription factors play important roles in lignin biosynthesis. A CsWRKY13 gene was identified from tea plant cultivar 'Longjing 43'. CsWRKY13 was highly expressed in stems of tea plants. Overexpression of CsWRKY13 in Arabidopsis decreased the lignin content in transgenic Arabidopsis plants, and also caused downregulation of lignin biosynthesis related genes. Further research on the function and regulation mechanism of the CsWRKY13 gene will broaden our comprehension of WRKY transcription factors and provide a basis for lignin biosynthesis in tea plants.
- The research was supported by the Jiangsu Agricultural Science and Technology Innovation Fund (JASTIF, CX(20)3114), National Natural Science Foundation of China (31870681), and Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD).
- The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers used for qRT-PCR in this study.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Teng R, Wang Y, Lin S, Chen Y, Yang Y, et al. 2021. CsWRKY13, a novel WRKY transcription factor of Camellia sinensis, involved in lignin biosynthesis and accumulation. Beverage Plant Research 1:12 doi: 10.48130/BPR-2021-0012

CsWRKY13, a novel WRKY transcription factor of Camellia sinensis, involved in lignin biosynthesis and accumulation
- Received: 10 September 2021
- Accepted: 14 October 2021
- Published online: 09 November 2021
Abstract: Lignin is an aromatic polymer that provides the necessary mechanical strength for the transport of water and nutrients in higher plants. Lignin biosynthesis and accumulation affect growth and development of tea plants. The degree of lignification related to the tenderness of fresh tea leaves determines the quality of tea. WRKY transcription factors play central roles in plant development and physiological processes. However, the roles of WRKY transcription factors in lignin biosynthesis of tea plants remain unclear. In this study, a WRKY gene, CsWRKY13, was cloned from tea plant 'Longjing 43'. The open reading frame (ORF) of CsWRKY13 gene was 708 bp, encoding 235 amino acids. Sequence analysis showed that CsWRKY13 contained a conserved WRKYGQK amino acid sequence and a zinc-finger-like motif CX4CX23HXH. Subcellular localization showed that CsWRKY13 was localized in the nucleus. The yeast trans-activation assay showed that CsWRKY13 had no transcriptional activity. Expression analysis showed that the CsWRKY13 gene was highly expressed in the stem. Overexpression of CsWRKY13 in Arabidopsis thaliana reduced lignin content and the expression levels of genes related to lignin biosynthesis in transgenic plants. Most flavonoids pathway related genes were significantly up-regulated. This study shows that CsWRKY13 might function as a negative regulator in regulation of lignin synthesis.
-
Key words:
- Camellia sinensis /
- Lignin /
- CsWRKY13 /
- Biosynthesis /
- Yeast /
- trans-activation assay /
- Gene expression