










-
The leaf organ is the basic vegetative organ of plants, the main place for photosynthesis, and plays an essential role in the life activities of plants. The establishment of leaf polarity is the core of leaf development. The establishment of leaf polarity directly determines the development of leaf spreading and affects the physiological functions of photosynthesis, transpiration and stress resistance of plants. The transcription factors which regulate leaf polarity and leaf growth such as HD-ZIP genes AtPHABULOSA (AtPHB), AtREVOLUTA (AtREV) and AtPHAVOLUTA (AtPHV)[1], AtAS2 (ASYMMETRIC LEAVES2)[2], KANADI[3], AtYABBY and small non-coding RNAs[4] have been identified in Arabidopsis. Among them, the YABBY family is a unique regulator in seed plants[5].
The YABBY transcription factors contain two conserved domains, a C2C2 zinc finger domain located at the N-terminus and a helix-loop-helix named the YABBY domain which is located at the C-terminus of the protein[6]. In most dicotyledonous angiosperms, there are few YABBY family members which can usually be divided into five groups, CRABS CLAW (CRC), YABBY2 (YAB2), FILAMENTOUS FLOWER (FIL)/YABBY3 (YAB3), YABBY5 (YAB5), INNER NO OUTER (INO)[7−9]. YABBY genes have been reported that regulate plant growth and development, such as leaf development[10], establishment of anterior and posterior axiality of the blade[11], development of SAM[5] and flower development regulation[12]. In Arabidopsis, FIL/YAB3 and YAB2 are restrictedly expressed in abaxial regions of lateral organs including leaves and flower. AtCRC is restrictedly expressed in the abaxial domains of carpels and AtINO is only expressed in the outer integument of ovules[11]. In rice, OsYAB3 is orthologous to Arabidopsis FIL. No phenotypic changes were observed in its overexpressed plants, while OsYAB3 RNAi plants produced twisted and knotted leaves with no reduction in leaf width sheaths and leaves. This suggests that rice OsYAB3 is involved in leaf growth and differentiation[13]. OsDL in rice is orthologous to the Arabidopsis CRABS CLAW (CRC) gene. Ectopically expressed DL plants produce curled leaves, forming a cylindrical structure[14].
Camellia sinensis (L.) O. Kuntze (C. sinensis) is a perennial evergreen woody plant, which belongs to the Camellia genus. Fresh leaves are the ingredients for tea processing. China is a major tea-producing country. China's tea product market has many well-known categories of green tea. Raw materials of the most famous Chinese green teas, such as Xihu Longjing, Biluochun, Huangshan Maofeng, Lu'an Guapian, Xinyang Maojian, and Taiping Houkui, are all limited by the degree of leaf development or leaf shape (Supplemental Fig. S1). Abiotic stress, especially drought, will seriously affect the growth and yield of tea. Tea plants showed slowed growth of newly grown leaves which are small and of poor quality under drought conditions, which are not suitable for tea processing[15]. On the other hand, dehydration of fresh leaves in vitro is an essential step for processing of some Chinese teas. Withering is a critical process for white tea processing, and plays a vital role for the unique chemical form of white tea. During withering, the moisture content in the leaf is reduced by 70%–78%[16]. In the present study, the CsYABs were screened and phylogenetically analyzed. The functions of CsYABBYs were characterized by heterotopic expression in Arabidopsis. The correlation between CsYABs gene expression and leaf development in the leaves on the picked shoots and under drought conditions were studied.
-
The tissues of the tea plant (C. sinensis 'Duokangxiang') were collected from Nongcuiyuan Tea Plant Cultivar & Germplasm Resource Garden of Anhui Agricultural University at the end of May. A. thaliana (Col-0 ecotype) were used and grown in a green house. The methods used for seed germination and plant growth for A. thaliana were the same as those provided in our previous work[17]. Seeds were sterilized using 1.5% sodium hypochlorite (10% Clorox) for 8 min, washed three times with sterile distilled water, treated with 70% alcohol for 1 min, washed three times again with sterile distilled water and then germinated on an agar-solidified MS medium supplemented with 3% sucrose. The temperature, light intensity, and photoperiod were 22 ± 2 °C, 50 µmol m−2 s−1, and 16/8 h (light/dark), respectively.
Escherichia coli DH5α and BL21, Agrobacterium tumefaciens GV3101 (TransGenBiotech, Beijing, China) were used as the host strain and expression strain for the vector construction. pDOR207 and pCB2004 were kindly provided by Professor Chengbing Xiang from the University of Science and Technology of China.
Cloning of CsFILa, CsFILb, and CsYAB2
-
Total RNA was extracted from the buds of tea plants using RNAiso mate and RNAiso Plus (Takara, DaLian, China) according to the manufacturer's instructions. The reverse transcription reaction was carried out using the PrimeScript RT Reagent Kit (Takara). The full open reading frame (ORF) was amplified by high-fidelity PCR using cDNA from the total RNA according to the CDS sequence provided at Tea Plant Information Archive (TPIA;
http://tpia.teaplant.org ).The primers of YABBY are provided in Supplemental Table S1. The PCR program was performed as follows: Starting with 98 °C for 30 s, followed by the cycle program including 98 °C for 10 s, 60 °C for 30 s and 72 °C for 45 s. This cycle program is repeated 30 times. A final extension is performed at 72 °C for 10 min. The PCR amplification products were purified using a standard 1% Agarose Gel following the manufacturers instructions (Aidlab, Beijing, China). The purified DNA were ligated into an Easy-Blunt vector (TransGen Biotech, Beijing, China) and then transformed into DH5α competent cells for sequencing.
Sequence alignment and phylogenetic analysis
-
Deduced amino acid sequences of CsFILa, CsFILb, CsYAB2, CsYAB5, CsINO and CsCRC were used for BLAST searches of the TPIA site (
http://tpia.teaplant.org/index.html ). The obtained sequences were used for alignment using the DNAMAN. Those homologous sequences were further used to construct phylogenetic trees using a neighbor-joining method with bootstrap analysis of 1,000 replicates, which is included in the MEGA 5 software. The resulting phylogenetic tree clustered CsYABBYs together with their closest homologs.Conserved motif analysis
-
The conserved motifs of CsYABBY proteins were used for identification via the online MEME software (v. 4.9.1) (
https://meme-suite.org/meme/tools/meme )[18]. The motif discovery mode selected was Classic Mode. The parameter of the motif number was set to less than 20 motifs, with a maximum width of 200 amino acids.Quantitative realtime PCR
-
On a 96-well plate, each reaction system is 20 μ, including 100 ng/μL cDNA template, 10 μL SYBR Green PCR Master Mix (Takara), and 200 nmol L−1 gene-specific primers. The PCR procedure was as follows: 95 °C for 30 s and 40 cycles of 95 °C for 5 s, 30 s at 60 °C, and 30 s at 72 °C, followed by melting curve analysis from 55 °C to 95 °C. GAPDH in tea plant was selected as a reference gene sample for various tissues. Three biological replicates and three technical replicates per biological replicate were performed for each experiment. The specificity of amplification was verified through melting curve analysis (55 °C to 95 °C). The gene glyceraldehyde-3-phosphate dehydrogenase (GAPDH, Accession No. GE651107) in tea plants was used as control genes in qPCR analysis. Their gene primers are listed in Supplemental Table S1. The relative expression level was calculated using a previously described method[19].
Agrobacterium mediated transformation of Arabidopsis
-
The ORFs of the CsYABBYs were cloned into the vector pDONR207 using the Gateway BP Enzyme mix according to the manufacturer's instructions (Invitrogen, Carlsbad, CA, USA). The primers are listed in Supplemental Table S1. The entry vectors were recombined into the plant transformation destination vector pCB2004 using the LR Enzyme mix (Invitrogen). The recombinant pCB2004-YABBYs were electroporated into Agrobacterium tumefaciens Gv3101. The Agrobacterium tumefaciens infused with pCB2004-YABBYs was confirmed by PCR. The PCR procedure was performed at 98 °C for 10 min, 94 °C for 2 min, 30 cycles at 94 °C for 30 s, 60 °C for 30 s and 72 °C for 45 s, followed by a final extension at 72 °C for 10 min.
An Agrobacterium tumefaciens colony was transferred into Arabidopsis using the floral-dip method as previously described[20]. The transgenic plants were confirmed by PCR with transcript template using end-to-end primers. The PCR procedure was performed at 94 °C for 2 min, 30 cycles at 94 °C for 30 s, 60 °C for 30 s and 72 °C for 45 s, followed by a final extension at 72 °C for 2 min.
The inflorescence infestation method was used to screen the Glufosinate-ammonium -added medium (add 90 μL of 50 mg/mL Glufosinate-ammonium and 300 μL of 300 mg/mL Temetine per 300 mL of MSK medium) to obtain transgenic Arabidopsis. The transgenic plants were confirmed by PCR amplification with ORF primers of YABBY.
Calculating the leaf rolling index
-
The natural width of the Arabidopsis leaf (the width of the leaf in its natural state, Ln), and the flat width (the width when the leaf is flattened, Lw) were measured, and the leaf rolling index (LRI) was calculated according to the formula: LRI = (Lw − Ln) / Lw × 100%[21].
Drought treatment with PEG4000
-
The tea shoots containing four leaves and one bud were picked and kept in 500 mL tissue culture flask with 200 mL half concentration of Murashige and Skoog medium (1/2 MS) for 24 h. Then 1/2 MS containing 10% and 15% PEG4000 were used to simulate drought conditions. The leaf length and width of shoots culturing for 0 h, 24 h and 48 h were measured, respectively. Leaves developed from buds at the corresponding time were collected for extracting RNA and detecting the gene expression level.
Statistical analysis
-
Statistical significance of trends was evaluated with SPSS software (SPPS Inc., Chicago). Two-group comparative analyses were performed with Student's t test. Parameters were calculated using data from three biological replicates.
-
We searched available transcriptomes and genomes of C. sinensis and identified six CsYABBY transcription factors (Fig. 1). The phylogenetic analysis containing five Arabidopsis and 40 known YABBY proteins from Angiosperm was performed by a neighbor-joining (NJ) phylogenetic tree. Protein information of YABBYs in the phylogenetic tree is shown in Supplemental Table S2. Phylogenetic analysis showed two CsYABBYs were classified into FIL/YAB3 clusters, named CsFILa and CsFILb. The other four CsYABBYs were classified into YAB2, YAB5, INO and CRC clusters, respectively (Fig. 1), which were named CsYAB2, CsYAB5, CsINO and CsCRC.
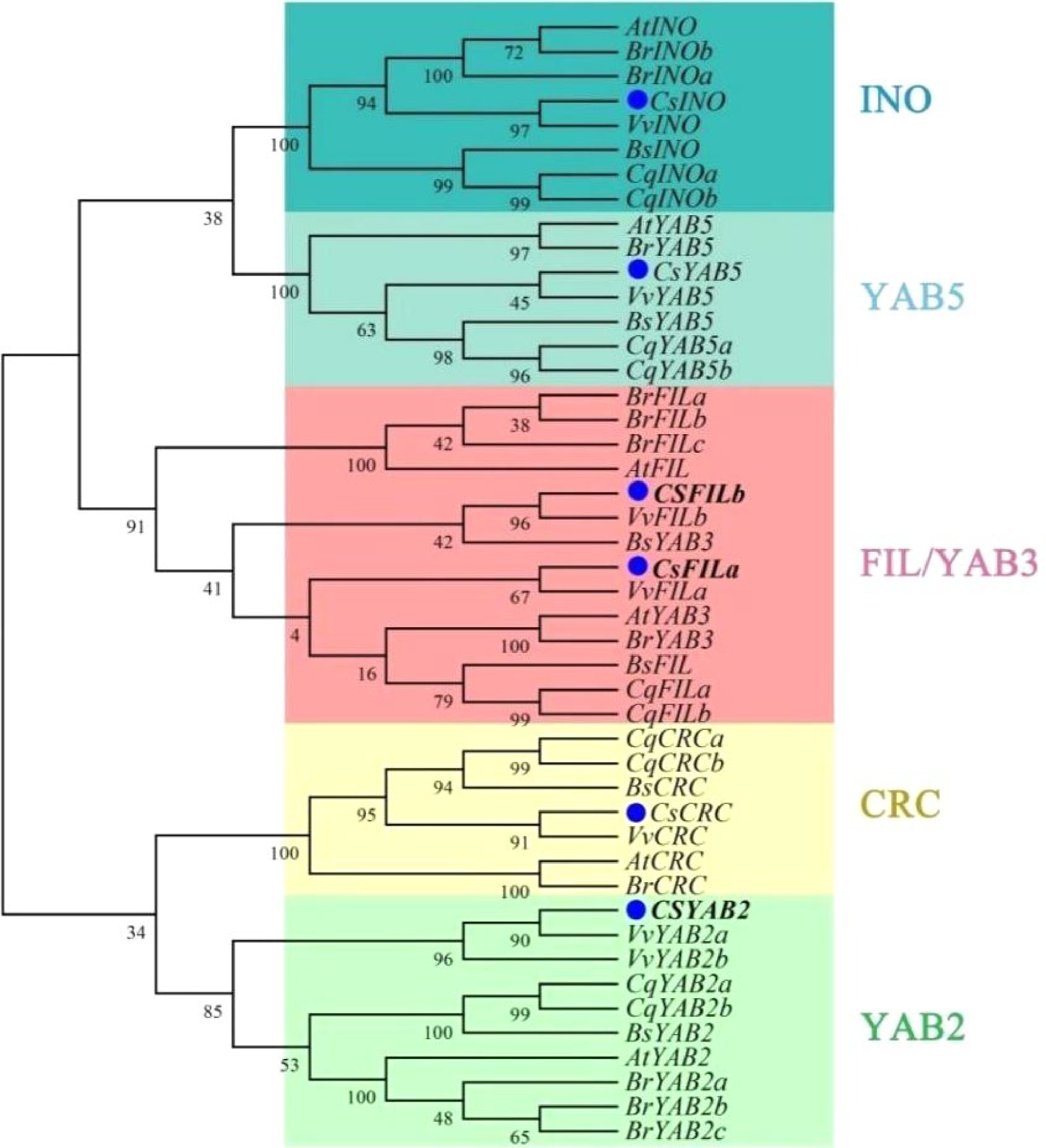
Figure 1.
Phylogenetic analysis of the YABBY transcript factors of C. sinensis. Cs, Camellia sinensis; Vv, Vitis vinefera; At, Arabidopsis thaliana, Br, Brassicarapa, Bs, Bienertia sinuspersici; Cq, Chenopodium quinoa. Protein information of YABBYs in the phylogenetic tree is shown in Supplemental Table S2. The phylogenetic tree was constructed by the p-distance method using MEGA X.
All YABBY transcription factors shared conserved YABBY (motif 2) and zinc finger domains (motif 1, Fig. 2). The orthologous proteins in different species are highly conserved and share multiple motifs. Among them, the CsYAB2 lack of motif 5 is compared to other YAB2 cluster proteins. CsINO lack of motif 18, motif 7, motif 4 is compared to other INO cluster proteins. It suggested that their functions might be different from other orthologous proteins. CsFILa and CsFILb shared almost the same motifs with AtYAB3 indicating these three proteins performed the same functions.

Figure 2.
Motifs of the YABBY family proteins from C. sinensis, V. vinefera, A. thaliana, B. rapa, B. sinuspersici, and C. quinoa. The motifs were identified using MEME Suite 5.1.0 server on line (
http://meme-suite.org/tools/meme ). The detailed protein information is listed in Supplemental Table S2.Expression of CsYABBY in different tissues of Camellia sinensis
-
In order to screen CsYABBYs which are involved in leaf development, the gene expression profiles of CsYABBY in different tissues were acquired from the Tea Plant Information Archive (TPIA;
http://tpia.teaplant.org ) established by our laboratory[22]. The CsFILa was highly expressed in buds while being lower expressed in other organs or tissues (Fig. 3a). There is no expression of CsFILa in roots. The expression of CsFILb gradually decreased with development of leaves.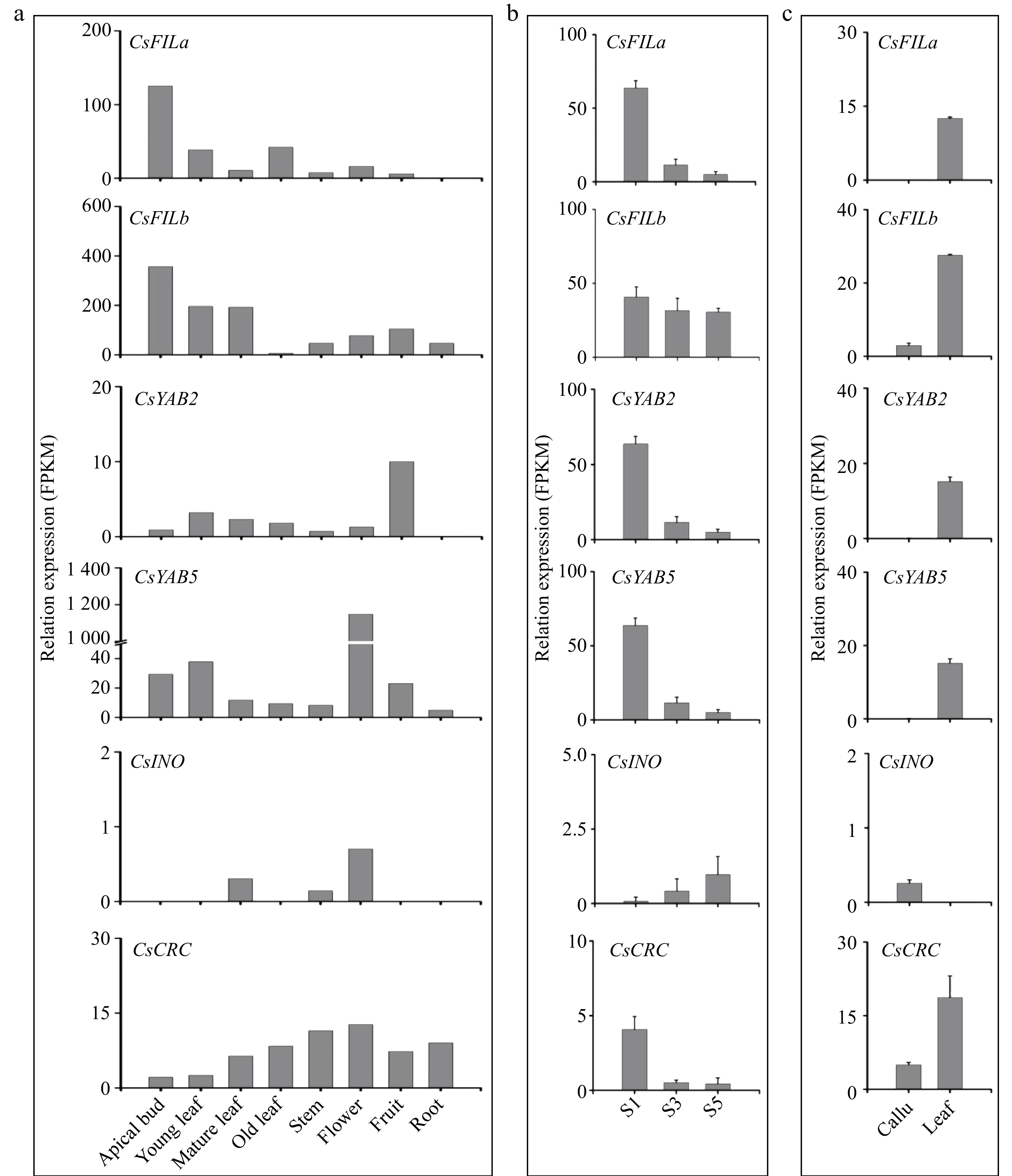
Figure 3.
Relative expression level of CsYABBY genes in different tissues, organs, or callus of tea plants. S1−S3, the different developmental stages of flowers. The data for (a) is derived from TPIA previously established by our laboratory[22].
The CsYAB2 was relatively highly expressed in young and mature leaves compared with that of leaves at other developmental stages. Among different organs, CsYAB2 and CsYAB5 were highly expressed in fruits and flowers, respectively (Fig. 3a & b). In contrast, CsYAB5 expressed at a low level in other organs. This suggests that the CsYAB5 gene is involved in flower development. CsINO was expressed lower in all tissues or organs. CsCRC was relatively highly expressed in mature leaves, stem, flower and roots.
The expression of CsFILa, CsFILb, CsYAB2, CsYAB5, and CsCRC gradually decreased with development of flowers, contrary to the expression trend of CsINO (Fig. 3b). Almost all CsYABBY genes expressed lower or were not expressed in the callus (Fig. 3c). The expression pattern and level of CsYABBY genes in fresh leaves at different growth stages and organs or callus suggested that CsFILa, CsFILb and CsYAB2 may be more related to leaf development.
In order to examine the accuracy of the above gene expression profile data, the expression levels of CsFILa, CsFILb and CsYAB2 were checked again in leaves and flowers at different developmental stages by q-PCR. The results suggested that the expression trends of CsFILa, CsFILb and CsYAB2 by quantitative PCR analysis was basically consistent with that in the transcriptome. CsFILa was highly expressed in buds, the expression level decreased with the development of leaves. CsFILb was highly expressed in buds. The expression level of CsYAB2 increased with the development of leaves but was low in old leaves. CsFILa, CsFILb and CsYAB2 showed a relatively high expression in mature petals (Fig. 4).
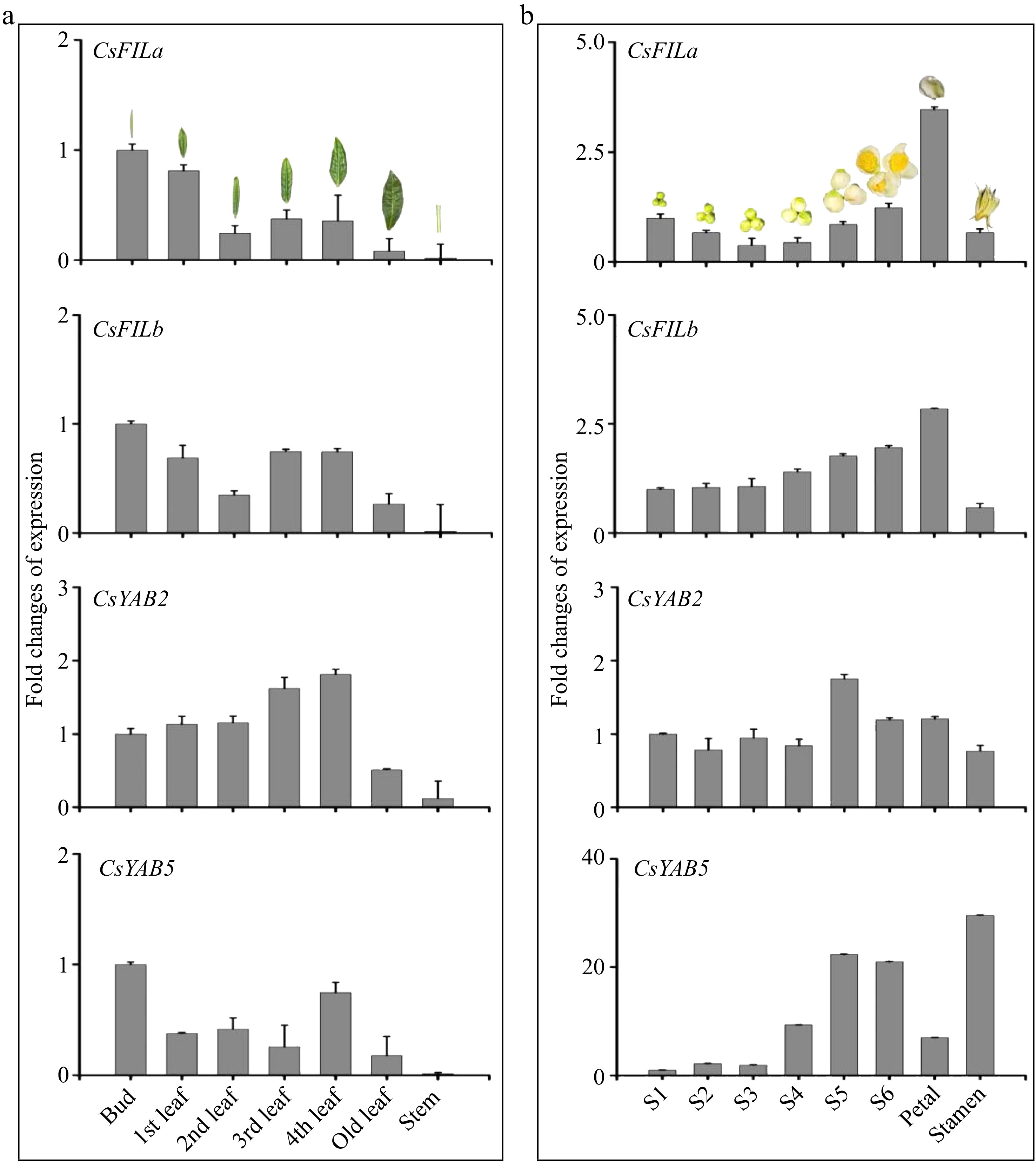
Figure 4.
Expression levels of CsYABBY genes in different tissues or organs of tea plants by quantitative PCR analysis. S1−S6, the different developmental stages of flowers. 1st−4th leaf, the different developmental leaf of tea plants.
The above results suggested that CsFILa, CsFILb and CsYAB2 were positively correlated with leaf development. Therefore, these three genes were selected for the further study of gene function identification.
Functional characterization of CsCsFILa, CsFILb, and CsYAB2
-
To identify the function of CsFILa, CsFILb, and CsYAB2, the ORF of CsCsFILa, CsFILb, and CsYAB2 was cloned into PCB vector which was triggered by a 35S promoter, the recombinant vector was then transferred into Arabidopsis by the floral dip method. PCR amplification showed that the CsFILa, CsFILb, CsYAB2 genes has been successfully transformed in Arabidopsis (Supplemental Fig. S2). The whole growth cycle of CsYAB2 overexpressed plants was normal and the phenotype was similar to that of wild-type plants. (Fig. 5a). There is no specific phenotype in CsCsFILa and CsFILb overexpressing plants before the four- euphylla seedling stage (7-day-old) or in the newly grown young leaves. But, the leaves at the middle growth stage of the transgenic lines (T4, 30-day-old) curled to the abaxial side and became long-narrow (Fig. 5a). The curling degree of the leaves increases with the growth of the leaves (Fig. 5b). Leaves of CsFILa overexpressed plants curled more from mature than young leaves in contrast with the CsFILb transgenic plant.
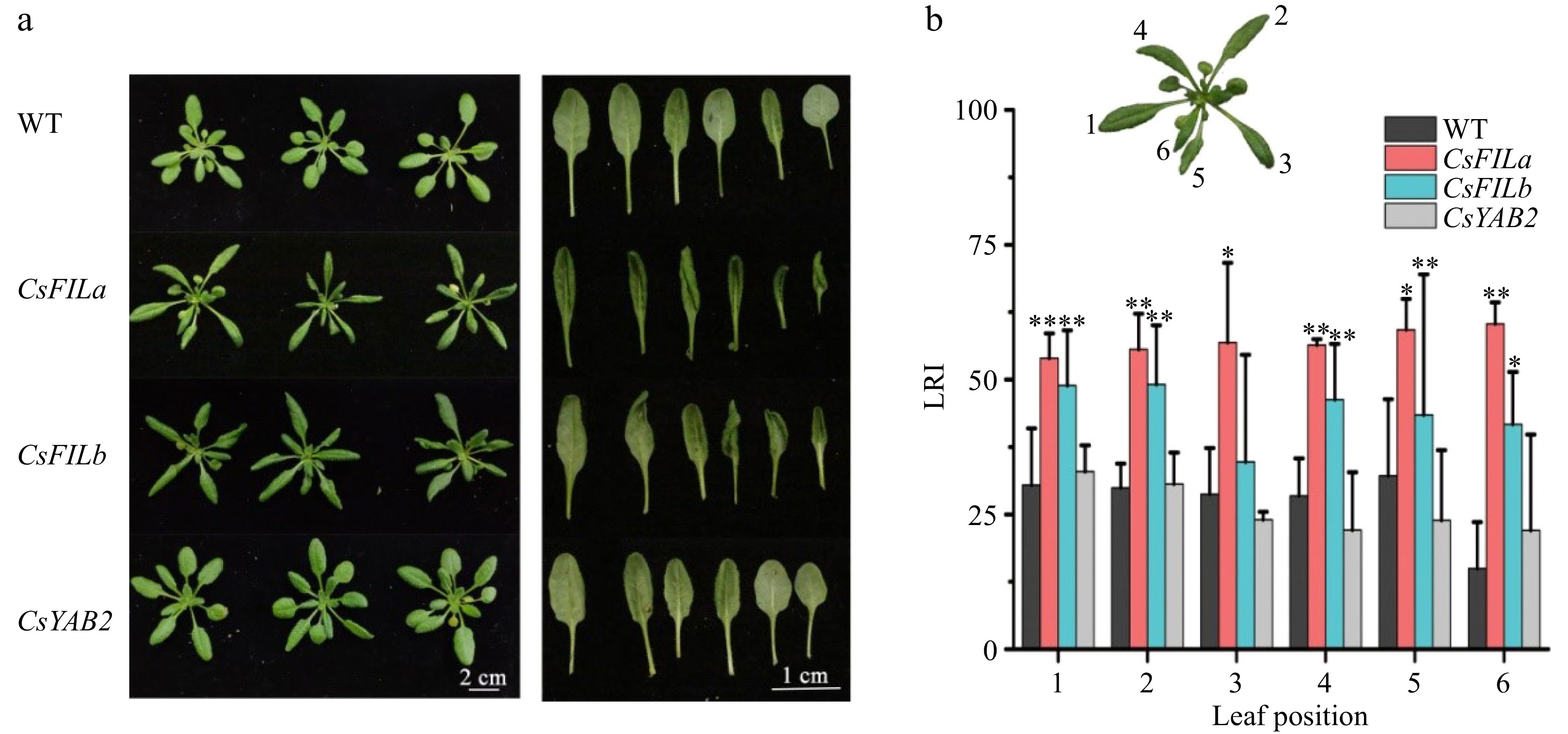
Figure 5.
The curl phenotypes of leaves caused by overexpression of CsFILa, CsFILb and CsYAB2 in Arabidopsis. (a) Phenotypes of leaves of Arabidopsis overexpressed of CsFILa, CsFILb and CsYAB2 (T4 generation). (b) The rolling index of leaves at different developmental stage from the transgenic Arabidopsis. The leaf rolling index (LRI) was calculated according to the formula: LRI = (Lw − Ln) / Lw × 100%[21]. Significant differences are represented by asterisks. P-value less than 0.05 and 0.01 indicate statistical significance and extreme significance, respectively (n = 3, * P < 0.05, ** P < 0.01).
Expression patterns of CsYABBYs in tea leaves on picked shoots
-
Picking young leaves or shoots is the first step in tea processing. For white tea processing, the leaf will be wilted for 24−48 h after picking, and then further processing begins. As shown in Fig. 6a−c, leaf spreading and growth rate on picked shoots were significantly lower than those in field plants. The length and width of tea leaf in the field increased by 32% and 77% within 48 h, respectively. Correspondingly, the length and width of the picked leaf increased by 18% and 51%, respectively. This showed that fresh leaves are still developing and unfolding slowly after picking (Fig. 6b & c). When the tea leaves in the field spread from the bud, the expression levels of CsFILa, CsFILb and CsYAB2 increased rapidly within 24 h. Correspondingly, the expression levels of CsFILa, CsFILb and CsYAB2 decreased obviously in leaves on the picked shoots within 48 h (Fig. 6d & e). The above results suggested that the slow growth of the leaves of the picked shoots is positively correlated with the expression of CsYABBY genes.

Figure 6.
Leaf development and CsYABBYs expression pattern in leaves of the picked and unpicked shoots. (a) The development of leaf on the picked or unpicked shoots within 48 h. (b) Variation of width and length of leaf on the unpicked shoots within 48 h. (c) Variation of width and length of leaf on the picked shoots within 48 h after picking. (d) The expression pattern of CsFILa, CsFILb and CsYAB2 in tea leaves on the unpicked shoots. (e) The expression pattern of CsFILa, CsFILb and CsYAB2 in tea leaves on the picked shoots.
Expression patterns of CsYABBYs in tea leaves under biotic and abiotic stresses
-
To investigate the role of CsYABBYs in the development of leaves under biotic and abiotic stresses, the expression levels of CsFILa, CsFILb, CsYAB2 in leaves treated by drought (PEG), salinity (NaCl), cold and methl jasmonate were acquired from the Tea Plant Information Archive (TPIA;
http://tpia.teaplant.org , Supplemental Fig. S3)[22]. The expressions of CsFILa, CsFILb, CsYAB2 did not fluctuate regularly in tea plants exposed to salt and methl jasmonate. Under low temperature stress, the expression of CsFILb decreased by half compared with the control. The expressions of CsFILa, CsFILb, CsYAB2 were sensitive to PEG, and decreased dramatically in 24 h, 48 h and 72 h treatments (Supplemental Fig. S3).The expression levels of the three genes in leaves treated by drought (simulation by PEG) were verified via qRT-PCR. As shown in Fig. 7a−c, spreading and growth of leaves on picked shoots treated by 10% and 15% PEG almost stopped. Moreover, the decrease levels of CsFILa expression surpassed 60% and 80% in picked leaves under 10% and 15% PEG treatment for 24 h and 48 h, respectively (Fig. 7d & e). The above results suggested that CsFILa positively regulated the growth of tea leaves on picked shoots under drought treatment.
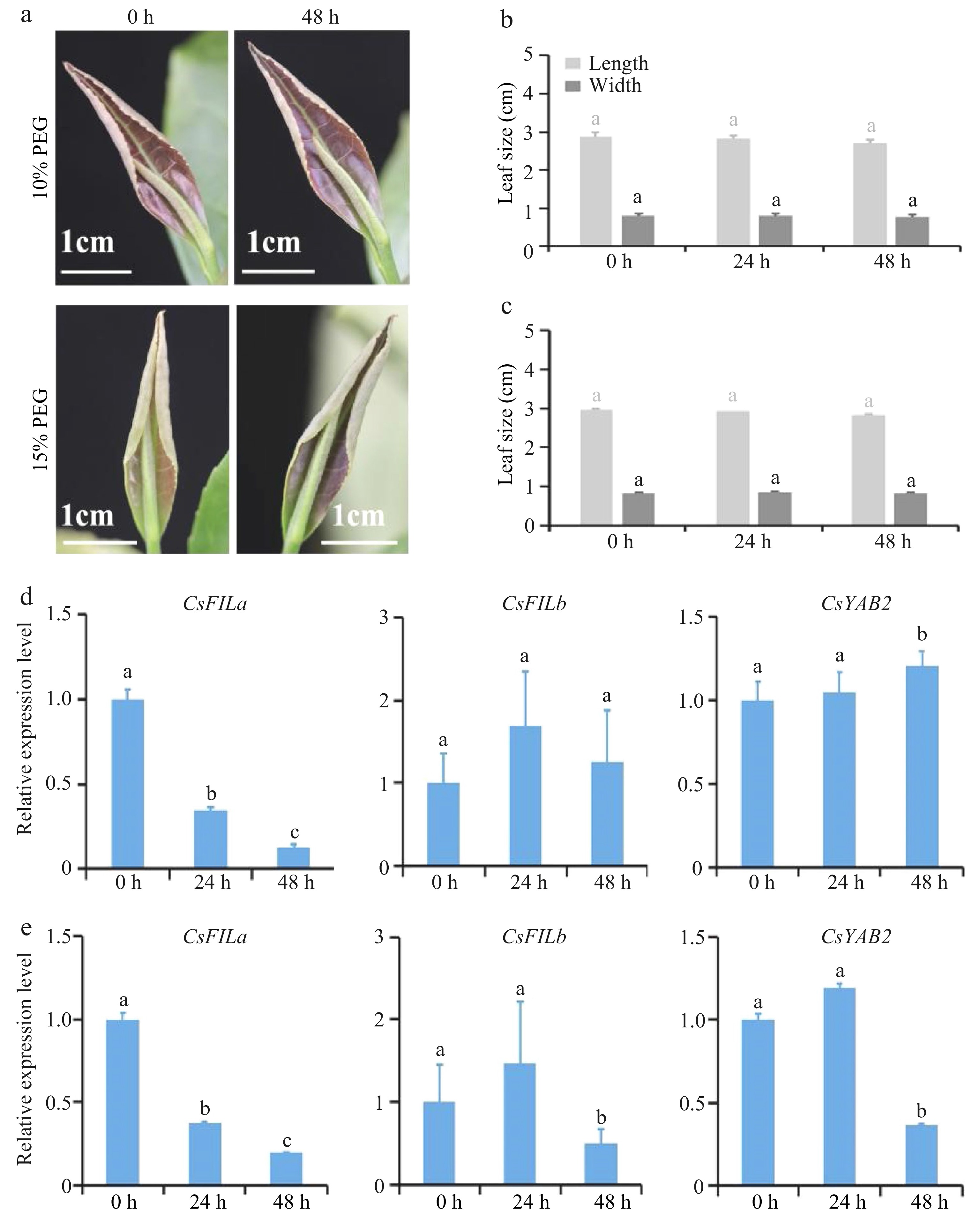
Figure 7.
Leaf development and CsYABBYs expression pattern in leaves of the picked shoots treated with 10% and 15% PEG. (a) The developmental phenotype of the leaf on the picked shoots treated with 10% and 15% PEG within 48 h. (b) Variation of width and length of the leaf on the picked shoots treated with 10% PEG within 48 h. (c) Variation of width and length of the leaf on the picked shoots treated with 15% PEG within 48 h. (d) The expression level of CsFILa, CsFILb and CsYAB2 in the leaf on the picked shoots treated with 10% PEG. (e) The expression level of CsFILa, CsFILb and CsYAB2 in the leaf on the picked shoots 15% PEG.
-
In dicotyledons, the establishment of leaf polarity is the core of leaf development. In the development of leaves, there are three axial polar differentiations: proximal and distal axis, basal and apical axis, abaxial-adaxial axis[23]. These three polar processes are interrelated, among which the establishment of the abaxial-adaxial axis is the most important. Homeodomain-leucine zipper family genes such as AtPHB, AtPHV and AtREV regulate adaxial cell fate[1]. The phb phv rev mutants can only develop into a needle cotyledon and hypocotyl with the characteristics of abaxial transformation, without any bilateral symmetrical structure[24]. This indicates these three genes are involved in the regulation of differentiation in the adaxial polarity of leaf. In Arabidopsis, YABBY family YAB2, YAB3, FIL and KANKDI(KAN) family KAN1, KAN2, KAN3 control the abaxial polarity of leaf. KAN regulates YABBY positively and PHB negatively. PHB negatively regulates the expression of KAN and YABBY during the process of regulating the adaxial polarity of leaf.
The function of ortholog genes of YABBY is conserved among different species. In most dicotyledonous angiosperms, members of the YABBY family usually contain five groups. The Arabidopsis genome contains six YABBY genes, AtFIL, AtCRC, AtINO, AtYAB2, AtYAB3, and AtYAB5. Six CsYABBY genes were found from the tea transcriptome and genome based on YABBY domain. CsFILa and CsFILb showed similar expression trends in various tissues, which may indicate their functional similarity (Fig. 3). The expression pattern and level of CsYABBY genes in fresh leaves at different developmental stages and organs or callus suggested that CsFILa and CsFILb are probably involved in leaf development of the tea plant. The leaves of Arabidopsis overexpressing CsFILa and CsFILb curled to the abaxial side and became long-narrow (Fig. 4a). The curling degree of the leaves increases with the growth of leaves. Similar results have been reported that GmFILa (a soybean YABBY gene) was overexpressed in Arabidopsis thaliana, which also caused leaf curling but does not change the leaves' internal structure[25].
In addition to regulating the adaxial polarity of leaf, YABBY transcription factors also repressed the expression of shoot apical meristem (SAM) regulators, KNOX and WUSCHEL, in the growing leaf[26]. AtFIL/AtYAB3 up-regulates (is up-regulated by) AtKAN1 and AtARF4, which forms a positive feedback loop[27,28]. Loss of YABBY function altered distribution of auxin signaling and changed normal auxin metabolism in YABBY multiple mutant Arabidopsis[5]. That means the YABBY gene also performs a feedback regulation toward auxin biosynthesis. Abiotic stresses inhibit plant growth and development. Drought is one biotic stress which is a common environmental factor causing a reduction in crop productivity worldwide[29,30]. The polar auxin transport is blocked when plants are exposed to abiotic stress[31].
In the present study, the growth rate of the tea leaf on the picked shoots was slower than that of the unpicked in the fields within 48 h. The expression levels of CsFILa, CsFILb and CsYAB2 decreased obviously in picked leaves within 48 h. When picked shoots of tea plants were treated with both 10% and 15% PEG for 24 h and 48 h, CsFILa expression levels surpassed 60% and 80%, compared to the control, respectively. Correspondingly, the development speed of plant leaves slowed down significantly, or even stopped. This may be due to drought restricting auxin synthesis and transport[32,33], which in turn reduced CsFILa gene expression. Transcription factors related to leaf development, FIL/YAB3, KAN1 and ARF4, established a positive feedback loop and their expressions were regulated by auxin signal[27]. It was reported the OsPIN3t acts in auxin polar transport in rice, the expression of which increased by NAA treatment and decreased by 1-N-naphthylphthalamic acid treatment (drought treatment)[33]. Our work suggested that YABBY transcription factors played an important role in leaf development under drought stress.
Wilting after tea picking of tea leaves or tender shoots is a common step in tea processing. For white tea processing, the leaf will be wilted for 24−48 h after picking, and then processing will continue. During this wilting process, fresh leaves still develop and expand slowly. The expansion degree of leaves is greatly affected by the degree of environmental humidity. This physiological activity is believed to be closely related to the expression of YABBY transcription factors.
-
Six YABBY transcription factors were identified from tea plants and regulated establishment of leaf polarity and development. The expression of CsFILa and CsFILb decreased with the development of leaves. The mature leaves of CsFILa and CsFILb transgenic Arabidopsis curled from the adaxial side to the abaxial side and became long-narrow. CsFILa, CsFILb and CsYAB2 genes positively regulated the growth of leaf on the picked shoots.
This work was supported by the Natural Science Foundation of China (31870677, 31870676, 31902069, U21A20232); the National Key R&D Program of China (2018YFD1000601); Collegiate Collaborative Innovation Foundation of Anhui Province (GXXT-2020-081).
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Typical and famous green teas in china.The processing raw materials of these teas are strictly limited by the development and shape of the tea leaf.
- Supplemental Fig. S2 The verification of transgenic plants of CsFILa, CsFILb and CsYAB2 in T1 generation by qRT-PCR with transcript template using end-to-end primers. Primers were listed in Table S1.
- Supplemental Fig. S3 The expressing level of CsFILa, CsFILb, CsYAB2 in leaves treated by salinity (NaCl, A), drought (PEG, B), cold (C), methl jasmonate (D). The data was acquired from Tea Plant Information Archive (TPIA;http://tpia.teaplant.org).
- Supplemental Table S1 Primers used in the study.
- Supplemental Table S2 Information of the YABBY family in the phylogenetic tree.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shen Y, Li X, Ma G, Zhao Y, Jiang X, et al. 2022. Roles of YABBY tanscription factors in the regulation of leaf development and abiotic stress responses in Camellia sinensis. Beverage Plant Research 2: 4 doi: 10.48130/BPR-2022-0004 

Roles of YABBY transcription factors in the regulation of leaf development and abiotic stress responses in Camellia sinensis
- Received: 01 December 2021
- Accepted: 30 December 2021
- Published online: 15 February 2022
Abstract: The formation of leaf polarity directly determines the development of leaf spreading and type, and affects the physiological functions of photosynthesis, transpiration and stress resistance of plants. Many famous Chinese teas are highly dependent on the morphological characteristics and size of the tea leaves. To date, there have been few reports on the establishment of tea leaf polarity and leaf type development. The plant-specific transcription factor YABBY family gene has been reported to be involved in the formation of leaf polarity and growth. Herein we describe the isolation and functional characterization of transcription factor YABBY genes of the tea plant (Camellia sinensis). Six YABBY transcription factors were isolated from tea plants and classified into five clusters in the protein phylogenetic tree. Among them, CsFILa and CsFILb were highly expressed in buds and mature petals, and their expression decreased with the stage of leaf development. The Arabidopsis transgenic lines of CsFILa and CsFILb showed abaxial curled and long-narrow leaves. The above results indicated that CsFILa and CsFILb regulated the leaf growth of tea plants. The growth and expansion rate of leaf on the picked shoots decreased significantly after being plucked from tea plants, especially those suffering from drought stress. In this process, CsFILa expression levels decreased 60%−80% compared to the control. These data indicated YABBY genes play important roles in the leaf development of Camellia sinensis.
-
Key words:
- Camellia sinensis /
- YABBY /
- Leaf polarity /
- Leaf development /
- Drought /
- Withering /
- White tea