










-
Tea, which is produced from Camellia sinensis buds or tender leaves, is the second most consumed beverage in the world. It originated in China, but it has become popular worldwide. According to the different manufacturing processes after harvesting, tea can be classified into six types[1]. Among them, the leaves used to make oolong tea are subjected to many stresses such as wounding stress, drought, heat, and UV/light radiation stress. Therefore, leaves processed to produce oolong tea are considered to be the best material for studying the biochemical responses of postharvest tea leaves to stress[2].
Wounding stress during oolong tea processing induces the production of many secondary metabolites like aroma compounds in tea leaves[2]. Aroma compounds, which are one of the major secondary metabolites, represent less than 0.03% of the tea product dry weight, but they affect tea flavor, making them key determinants of oolong tea quality[1]. The formation of tea aroma compounds mainly includes de novo synthesis and glycoside hydrolysis mediated by enzymatic reactions, as well as maillard reactions formed during thermophysical and chemical reactions[2]. More than 700 tea aroma compounds have been isolated and identified[1]. Among them, indole is an important aromatic compound in tea that endows oolong teas unique floral aroma[2]. Indole was significantly induced during the turn over stage of oolong tea processing due to the activation of the key synthetic gene Tryptophan synthase β-subunit 2 (CsTSB2)[3]. A recent study confirmed that DNA methylation is involved in the regulation of indole biosynthesis, implying that epigenetic regulation is involved in controlling aroma compound production in oolong tea processing[4]. In addition to the change in DNA methylation state of the genome, chromatin undergoes structural modifications in response to an exposure to external stresses.
The nucleosome is the basic unit of chromatin, consisting of a histone core around which 147 bp of DNA is encapsulated. Each histone core is an octameric disc composed of two copies of each of the four canonical histone proteins H2A, H2B, H3 and H4[5]. Histone modifications are key epigenetic modifications that regulate chromatin structure, involves the covalent alteration of histone amino residues, including acetylation, methylation, phosphorylation, and ubiquitination[6]. Unlike DNA methylation, histone acetylation loosens the nucleosome structure, making nucleoprotein-coated DNA more likely to bind to transcription factors, thereby activating gene transcription[7]. This process is controlled by histone acetyltransferase (HAT) and histone deacetylase (HDAC). Numerous studies have revealed the importance of histone acetylation for seed germination, morphogenesis, and stress response[8]. In recent years, some studies have also revealed its role in the regulation of secondary metabolism[9,10]. For example, in petunia, HAT is involved in regulating the formation of phenylpropanoid/benzenoid volatile compounds[9]. Therefore, we speculate that histone acetylation also plays an important role in the formation of tea aroma compounds, especially indole.
In this study, the oolong tea manufacturing process was used as a model for exploring whether histone acetylation contributes to the regulation of indole production in response to wounding stress. More specifically, we investigated the regulatory effects of histone acetylation on the biosynthesis of indole as well as the underlying mechanism. The results of this study indicate that histone acetylation affects the regulation of wounding-induced secondary metabolite biosynthesis.
-
During oolong tea processing, the unique turn over process induce the formation of a large amount of aroma compounds. Among them, indole is one of the characteristic aroma substances, giving oolong tea its unique floral aroma[2]. To verify whether histone acetylation modifications were involved, in this study tea branches were treated with histone deacetylase inhibitors. After 24 h of combinatorial histone deacetylase inhibitor feeding, the contents of histone 3 acetylation (H3ac) and histone 4 acetylation (H4ac) were detected, and it was found that both H3ac and H4ac were significantly increased (Fig. 1a), indicating the feasibility of inhibitor treatment. The samples were then subjected to mechanical wounding treatment and then tested for CsTSB2 expression level and indole content. After 24 h of treatment with the inhibitors, there was no significant difference between the treatment group and control groups (Fig. 1b & c). When the samples were subjected to mechanical wounding treatment, the CsTSB2 expression level decreased significantly, while the content of indole was also significantly decreased (Fig. 1b & c). Due to the fact that changes in metabolite content are expected to occur slightly later than the changes in gene transcription levels, the observed changes in the indole content was basically consistent with the changes in the expression of CsTSB2. These results indicate that histone acetylation modifications may be involved in regulating this process.
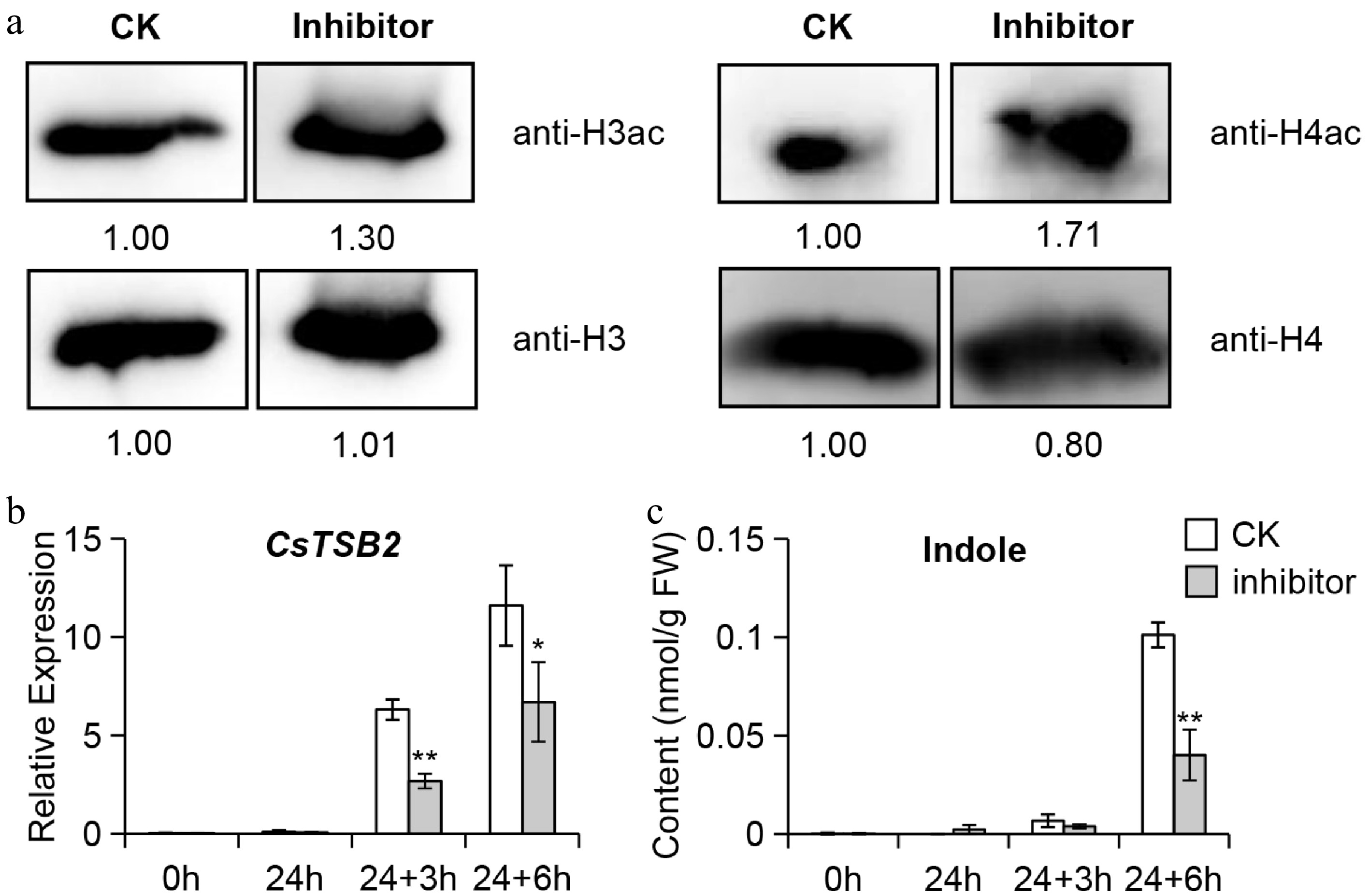
Figure 1.
Analysis of (a) H3ac and H4ac protein content, (b) CsTSB2 expression level and (c) indole content after HDAC inhibitor treatment in tea plants. (a) Detection of H3ac and H4ac content after combinatorial histone deacetylases inhibitor feeding for 24 h. CK, methanol-water (1/19, v/v) treatment group. Inhibitor, combinatorial histone deacetylases inhibitor treatment group. (b) Detection of CsTSB2 expression level and (c) indole content after HDAC inhibitor treatment. 24 h: incubate for 24 h with HDACs inhibitor or methanol/water; 24 + 3 h: After incubating with HDACs inhibitor or methanol/water for 24 h, perform mechanical wounding treatment for 3 h; 24 + 6 h: After incubating with HDACs inhibitor or methanol/water for 24 h, perform mechanical wounding treatment for 6 h. Internal reference gene: CsEF1-α. Data are expressed as mean ± S. D. (n = 3). Significant differences between control and treatment at the same different treatment time are indicated (* p ≤ 0.05, and ** p ≤ 0.01).
Continuous mechanical wounding increased acetylation levels of CsTSB2 promoter
-
To further verify whether histone acetylation modification is involved, western blotting was used to detect the total H3ac and H4ac contents of samples treated with mechanical wounding treatment. It was found that the H3ac content increased and the H4ac content decreased after treatment (Supplemental Fig. S1a). As the total H3ac and H4ac content changes, there are many other physiological reactions such as dehydration during mechanical wounding treatment. Therefore, ChIP-qPCR was used to detect the specific changes in histone acetylation modifications on CsTSB2 promoter. Total H3 and H4 protein levels were measured as controls. It was found that the total amount of H3 and H4 proteins did not change significantly during mechanical wounding treatment (Supplemental Fig. S1b). Previous studies have demonstrated that jasmonic acid is significantly increased during mechanical wounding treatment, thereby regulating the expression of these synthetic genes through the key transcription factor CsMYC2a (Myelocytomatosis proteins 2a) of JA pathway[2, 11]. Because the binding site of CsMYC2a is G-box or G-box-like, primers were designed in the regions containing G-box or G-box-like on CsTSB2 promoters for ChIP-qPCR experiments (Fig. 2a). The H3 and H4 acetylation levels in the CsTSB2 promoter regions increased significantly (Fig. 2b & c). An increase in histone acetylation activates gene expression, which is consistent with the results of this study[12]. Thus, continuous wounding promotes the expression of CsTSB2 related to the synthesis of indole by enhancing histone acetylation, which demonstrates the importance of histone acetylation in this process.
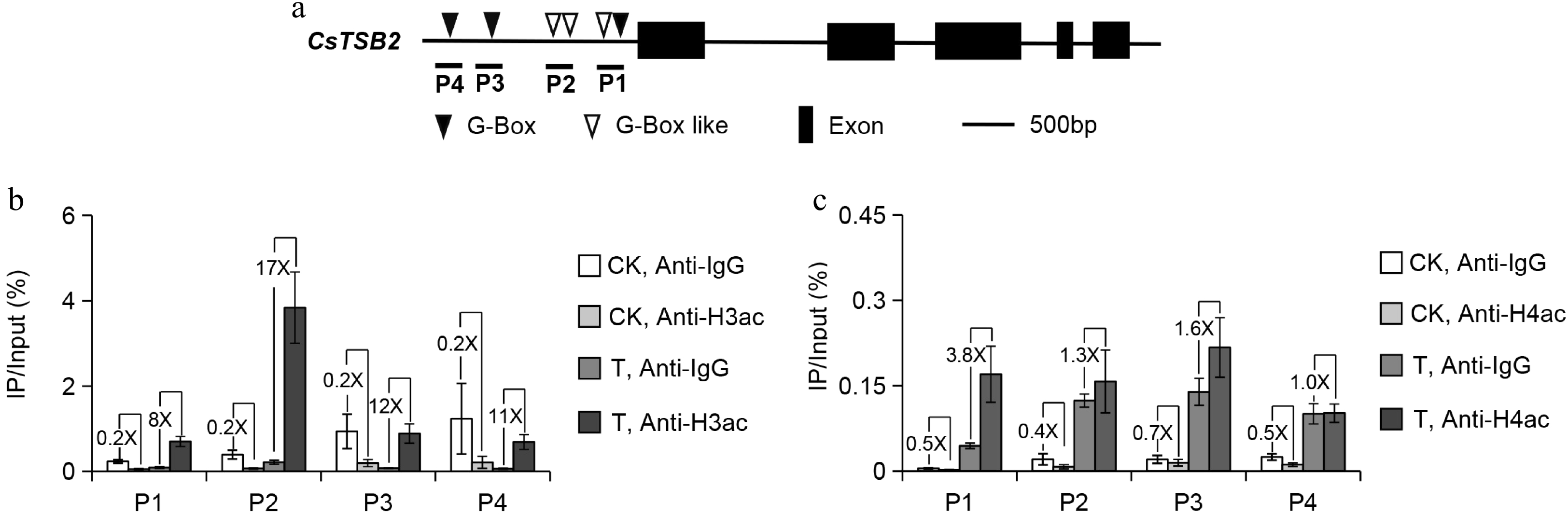
Figure 2.
ChIP-qPCR analysis of (b) histone 3 acetylation (H3ac) and (c) histone 4 acetylation (H4ac) enrichment of CsTSB2 promoter after wounding treatments. (a) The precise location of primers used for ChIP-qPCR are indicated. CK: No-treatment at 3 h; T: Continuous wounding treatment at 3 h; TSB, Tryptophan synthase β-subunit; P, Promoter. Anti-IgG antibody was used as a negative control. The 'number X' represents the difference multiplier between the two groups. Data are expressed as mean ± S. D. (n = 3).
CsHDA8 may be a key suppressor regulating CsTSB2 expression
-
Continuous wounding can regulate the biosynthesis of indole by modulating the histone acetylation levels of CsTSB2. Changes in histone acetylation levels are regulated by HATs and HDACs, therefore the changes in the transcription of HAT and HDAC genes in tea leaves were analyzed. Homologous alignment were made with reported HDACs in Arabidopsis thaliana, rice, soybean, tomato and sorghum. A phylogenetic tree was constructed using MrBayes v3.2.7a, and 19 CsHDACs family members were identified in tea, including nine members of the RPD3 family, five members of the HD2 family and five members of the SIR2 family (Supplemental Fig. S2). The same method was used to identify tea CsHATs (Supplemental Fig. S3). Eight tea CsHATs were indentified, including four GCN5-related N-terminal acetyltransferase family members (CsHAG1-3 and CsHAG5), three p300/CREB-binding protein activator family members (CsHAC1, CsHAC1-Like, and CsHAC12), and one TATA-binding protein-associated factor family member (CsHAF1).
Possible regulators were identified based on the expression levels of related genes. The expression levels of most of the CsHAT genes changed during the continuous wounding treatment (Supplemental Fig. S4). Previous studies have confirmed that aroma synthesis-related genes are expressed in large amounts during this treatment and that histone acetyltransferase acts as a positive regulator, and the candidate gene must be upregulated in the process[2,7]. Meanwhile, as histone acetylation modification is a post-transcription modification, its changes should precede the expression of aroma synthesis genes. Indole was already significantly increased at 3 h of mechanical wounding treatment, so the genes related to histone acetylation modification that already showed significant changes at 3 h would be the candidate genes. However, only CsHAG3 showed the expected trend. In addition to CsHATs, histone deacetylase, which also regulates the level of histone acetylation, may have played a role in this process. Because HDACs are negative regulators, those that were down-regulated in this process were screened. Most of the CsHDAC expression levels tended to be down-regulated by continuous wounding, with the most significant and consistent decreases observed for CsHDA2, CsHDA5, CsHDA8, CsHDA15, CsHDA19-1, CsHDT1-3, CsSRT1-1, CsSRT1-3 and CsSRT1-4 (Fig. 3 & Supplemental Fig. S5). Hence, those transcript level changes during oolong tea processing were analyzed. The results (Supplemental Figs S6 & S7) indicated that the expression of only CsHDA8 was significantly down-regulated in the key aroma formation stage (i.e., turn over stage). On the basis of these findings, we speculated that CsHDA8 may be the key HDAC regulating the synthesis of indole during oolong tea processing.
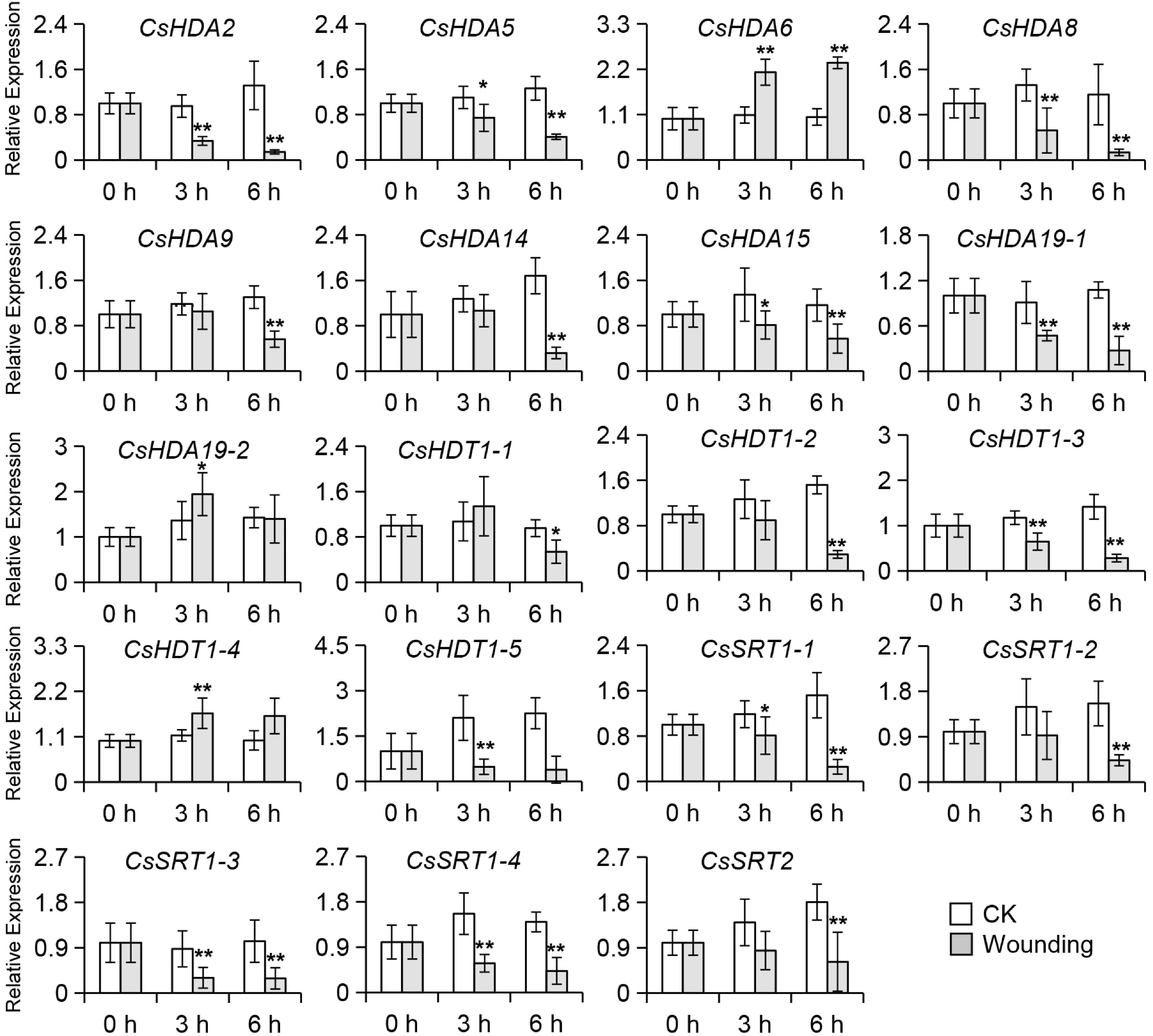
Figure 3.
Changes in expression levels of CsHDACs in tea leaves exposed to wounding treatment. HDA: histone deacetylase-A; SRT: sirtuin; HDT: histone deacetylase. Internal reference gene: CsEF1-α. Significant difference between control and wounding treatment are indicated (* p ≤ 0.05, and ** p ≤ 0.01). Data were expressed as mean ± S. D. (n = 3).
CsHDA8 is localized in the nucleus and cytoplasm
-
Histone deacetylases are mainly localized in the nucleus or cytoplasm or they may be shuttled between the two[13]. The HDACs located in the nucleus are primarily involved in regulating histone structures and functions, whereas the HDACs localized in the cytoplasm, chloroplasts, and mitochondria may also participate in the post-translational modification of certain proteins and the regulation of secondary metabolism. The RPD3/HDA1 subfamily members are distributed in the nucleus, cytoplasm, and some organelles. In contrast, the SIR2-like subfamily members are mainly located in the nucleolus, whereas the HD2 subfamily members are specifically localized in the nucleolus, suggesting that they may be involved in regulating ribosomal structures and functions[14]. In the current study, CsHDA8-YFP was detected in the nucleus and cytoplasm (Fig. 4a), which is in accordance with the localization of the known RPD3/HDA1 family members. Thus, CsHDA8 may function as an HDAC. However, the function of CsHDA8 in the cytoplasm will need to be determined in future investigations.
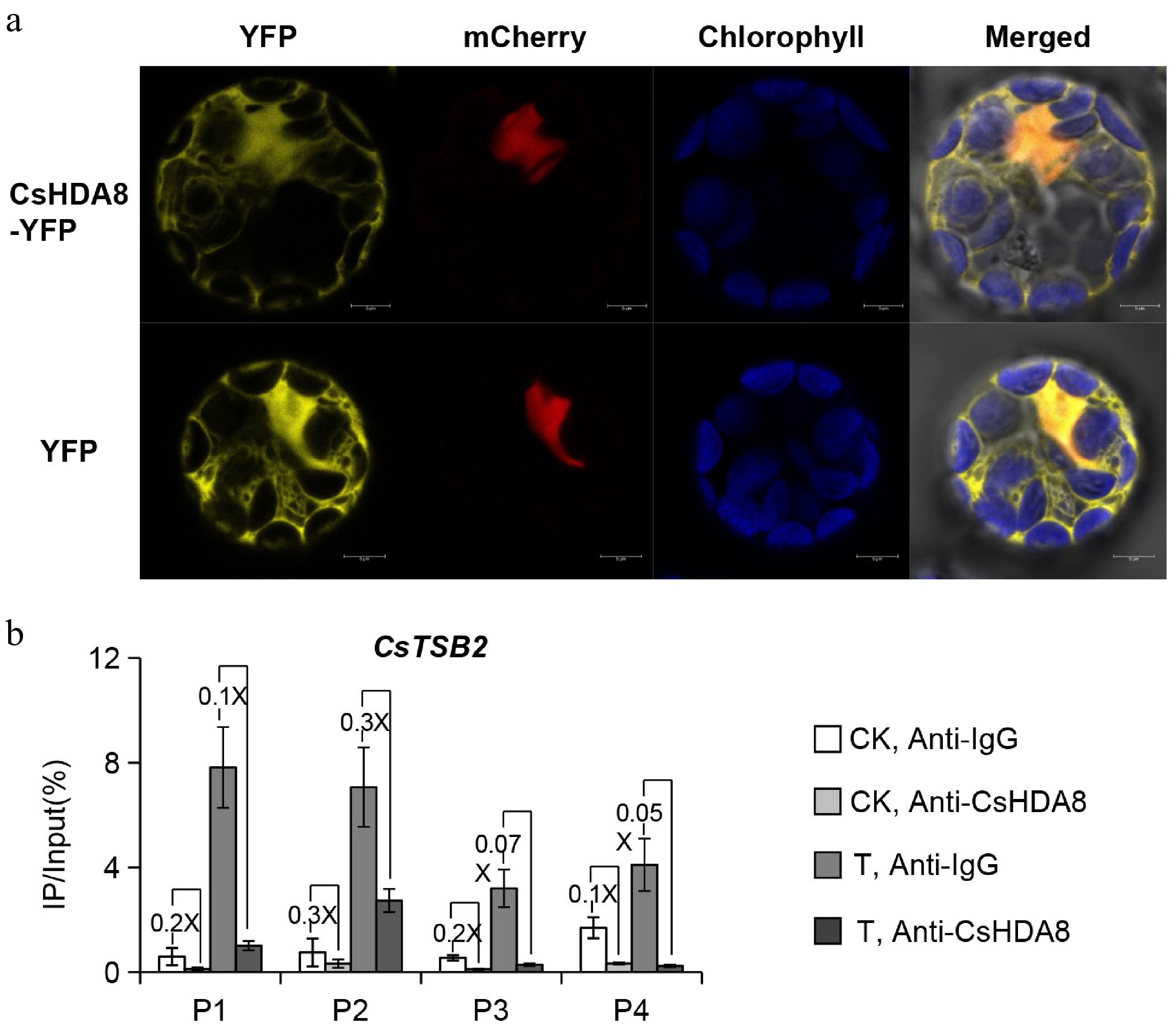
Figure 4.
The subcellular localization and binding level of CsHDA8. (a) The subcellular localization and binding level of CsHDA8 in Arabidopsis thaliana protoplasts. CsHDA8 and nuclear-localized mcherry are co-transferred into A. thaliana protoplasts. (b) ChIP-qPCR detection the binding level of CsHDA8 to CsTSB2 promoter after mechanical wounding treatments in tea plant. CK: No-treatment at 3 h; T: Continuous wounding treatment at 3 h. Anti-IgG antibody was used as a negative control. The specific positions represented by P1, P2, P3, and P4 in the figure are shown in Fig. 2a. The 'number X' represents the difference multiplier between the two groups. Data are expressed as mean ± S. D. (n = 3). HDA8, histone deacetylase 8. TSB, Tryptophan synthase β-subunit.
Continuous mechanical wounding treatment reduced CsHDA8 enrichment at the CsTSB2 promoter
-
A ChIP-qPCR experiment was performed to examine the binding of CsHDA8 to CsTSB2 promoter regions associated with the synthesis of indole in response to a continuous wounding treatment. The binding of CsHDA8 to CsTSB2 promoter regions decreased significantly after the treatment, suggestive of a regulatory role for CsHDA8 (Fig. 4b). The decreased binding of CsHDA8 to the promoter regions of CsTSB2 during continuous wounding led to an increase in the histone acetylation level in the examined promoters. The resulting up-regulated gene expression increased the biosynthesis of indole. Histone acetylations, which regulate the transcription of target genes, mainly occur on positively charged lysine residues. The acetylation neutralizes the positive charge, which adversely affects the binding of the lysine residue to DNA with phosphate groups. This leads to the loosening of the chromatin structure and increased gene expression[15].
-
Since tea plants have not yet established a stable genetic transformation system, there has been limited research on the epigenetic regulation in them. Therefore, studying epigenetic regulatory mechanisms in tea remains challenging. Verifying whether epigenetic modifications are involved in the regulation of secondary metabolites has become a major objective. In this study, experiments involving HDAC inhibitors and ChIP-qPCR were conducted to obtain in vivo evidence of the involvement of epigenetic modifications. The known HDAC inhibitors have been classified into the following four groups according to their structures: hydroxamic acid salts, fatty acids, cyclic peptides and benzamides. These HDAC inhibitors are widely used for cancer research and treatment[16]. There has recently been a gradual increase in their use in plant biology research. Among them, TSA was the first to be identified and is most often used in plant research. It has inhibitory effects on all zinc ion-dependent HDACs[17]. In an earlier investigation, a TSA treatment altered the response of the peanut AhHDA1 gene to drought stress, reflecting its repressive effect on HDACs in plants[18]. Therefore, TSA has been used in several studies, including those on plant cell dedifferentiation, pollen germination and pollen tube growth[19,20]. Additionally, TSA treatments reportedly affect the secondary metabolite synthesis pathway of Populus tomentosa, indicating that TSA is useful for studies on plant secondary metabolite regulation[21]. Sodium butyrate is a fatty acid that specifically inhibits the activity of the HDACs RPD3 and HD2, whereas nicotinamide and benzamide specifically inhibit SIRT1 and SIRT2. Recent studies demonstrated that the combined use of sodium butyrate and nicotinamide can increase gene editing efficiency in maize and wheat, suggesting they may also affect other biological functions when mixed[22]. Although the HDAC inhibitors in tea plants have not been extensively researched or used, other inhibitors are critical for studying the biological functions of tea secondary metabolites. For example, the use of polyphenol oxidase (PPO) inhibitors confirmed CsPPO is a key regulator of tea flavonol glycoside formation[23]. The use of ion channel inhibitors has also clarified the mechanisms mediating the transport of aluminum and fluoride ions in tea[24]. These studies revealed the importance of using inhibitors to conduct tea plant biology research, especially for in vivo analyses.
We previously determined that combining the HDAC inhibitors TSA, SBHA, sodium butyrate, and nicotinamide (1:1:1:1) for a treatment of tea leaves can effectively inhibit HDAC activities[25]. Therefore, the same treatment was used in this study. As expected, both H3ac and H4ac were significantly increased after inhibitor treatment, while the indole content was significantly changed. At the same time, using ChIP-qPCR, a method often used to study histone modification, further verified the results[4]. This method was developed to examine protein–DNA interactions. Formaldehyde immobilizes protein–DNA complexes in living cells, target proteins are enriched using specific antibodies, and purification and detection steps reveal the dynamic interaction between the trans-acting factor and the target DNA[26]. In this study, H3ac- and H4ac-specific antibodies were used to detect acetylated histones in CsTSB2 promoter before and after the continuous wounding treatment. There was a significant post-treatment increase in the H3ac and H4ac levels in the examined promoters, which likely resulted in the loosening of the chromatin structure to activate the expression of the corresponding genes, leading to the biosynthesis of aroma compounds. This evidence suggests that the combination of HDAC inhibitor treatment and ChIP analysis can provide in vivo evidence for tea plant research and may help elucidate the molecular mechanisms involved.
CsHDA8 may be a key suppressor that regulates the formation of indole under stress conditions
-
Histone deacetylases form a large gene family that is widely found in eukaryotes. Plant HDACs can be classified into the following three subfamilies: RPD3/HDA1, SIR2, and HD2 (specific to plants), of which the RPD3/HDA1 subfamily is the largest and its members are the most studied (e.g., HDA6 and HDA19)[27]. In A. thaliana, AtHDA6 is vital for plant development, flowering, hormone responses, and abiotic stress responses[28]. Additionally, AtHDA19 has crucial effects on seed development, flowering, floral organ development, hormone responses, and abiotic stress responses[28]. In some life cycle processes (e.g., flowering or embryogenesis), AtHDA6 and AtHDA19 have redundant functions[29]. Other HDACs have also been characterized. The considerable size of the HDAC family may reflect the functional diversity among its members. Several studies verified the importance of other HDACs for plant growth and development, reproductive growth, the cell cycle, and stress responses[28,30].
In this study, CsHDA8 was revealed to be a key HDAC regulating indole formation during continuous wounding treatment. Moreover, it belongs to the RPD3/HDA1 subfamily and is exclusive to plants. To date, it has only been identified in A. thaliana, tomato, soybean, and tea[13,25,31−33]. In tea, CsHDA8 expression is significantly down-regulated by drought stress, salt stress, and methyl jasmonate treatment, indicating that CsHDA8 may be an important abiotic stress response factor[31]. This is consistent with the findings of an earlier study on soybean[32]. Another study revealed a strong correlation between CsHDA8 expression and theanine and catechin biosynthesis, suggesting that CsHDA8 is a vital regulator of secondary metabolite production in tea plants, which is also supported with our previous findings[31]. Drought stress, insect feeding (Supplemental Fig. S8) and mechanical wounding treatment can induce a large number of changes in secondary metabolites[25]. Meanwhile, the expression of CsHDA8 was significantly reduced under all these stress treatments, indicating a strong correlation between CsHDA8 and the formation of secondary metabolites. There are relatively few reports on HDA8, and its function has not been thoroughly characterized, likely because most studies on epigenetic regulation in plants have focused on growth, development, or stress responses, with little attention paid to the regulation of secondary metabolism. In fungi, HDACs were the first histone-modifying enzymes reported to regulate the formation of secondary metabolites[34]. Therefore, HDACs may also play an important role during the regulation of plant secondary metabolism. Because tea plants are rich in secondary metabolites, clarifying the functions of CsHDACs may help to elucidate the mechanisms regulating secondary metabolism in tea plants, while also expanding our understanding of general HDAC functions.
Reduced binding of CsHDA8 on the CsTSB2 promoter after mechanical wounding induces the expression of CsTSB2
-
In this study, the continuous wounding treatment inhibited the binding of CsHDA8 to CsTSB2, which increased the acetylation of the promoter sequences. The subsequent chromatin loosening resulted in transcriptional activation. However, the current research evidence can only prove that CsHDA8 is involved in regulating the expression of CsTSB2, but its specific binding site, whether it can directly bind to the promoter of aroma synthesis genes, etc. is still unknown. In the future, further verification is required through experimental systems such as electrophoretic mobility shift assay (EMSA) and Yeast one-hybrid assay. Histone deacetylases are typically recruited to target genes by transcription factors. Moreover, HDACs interact with transcription factors and help regulate transcription by decreasing the degree of histone acetylation in the target gene. This down-regulation of gene expression can be reversed in two ways. First, the weakening of the interaction between HDACs and transcription factors releases them from the target gene, leading to increased acetylation[28]. Second, other proteins outcompete HDACs for the binding to transcription factors, thereby restricting their ability to deacetylate histones in the target gene. For example, cold stress induces the binding between High expression of osmotically responsive gene1 (HOS1) and the histone deacetylation recruitment factor Flowering locus VE (FVE), which then releases HDA6 and activates the expression of Flowering locus C (FLC), thereby inhibiting flowering[35]. Therefore how CsHDA8 binds to CsTSB2 promoter to function and whether there are transcription factors that mediate this process remains to be further investigated. Moreover, histone acetylation and other epigenetic modifications, such as DNA methylation or histone methylation, often synergistically regulate gene transcription. In A. thaliana, AtHDA6 can interact with methyltransferase (AtMET1) to maintain transposon silencing by regulating histone acetylation and DNA methylation[36]. We previously demonstrated that the expression of the DNA methyltransferase gene Domains rearranged methyltransferase 3 (CsDRM3) decreases significantly after a mechanical wounding treatment; this leads to decreases in DNA methylation and the H3K9me2 modification in the CsTSB2 promoter, which induces the binding of CsMYC2a and activates CsTSB2 expression[4]. It is possible that DNA methylation, histone methylation, and histone acetylation jointly regulate indole biosynthesis. However, the specific regulatory mechanism is unknown. Additionally, whether CsHDA8 can interact directly with CsDRM3 is unclear and the epigenetic modifiers regulating H3K9me2 to control this process are unknown. Future research will need to further elucidate the mechanism underlying the biosynthesis of aroma compounds in tea.
-
This study included HDAC inhibitors and ChIP-qPCR analysis to generate in vivo evidence in tea plants and may be relevant for future basic research on tea. Moreover, an in-depth analysis of CsHDA8, a key HDAC that regulates the stress-induced production of indole, revealed that continuous wounding decreases CsHDA8 expression and the binding of CsHDA8 to CsTSB2 promoter in tea leaves. The resulting increase in histone acetylation and loosening of the chromatin structure activates the expression of CsTSB2. These changes ultimately lead to a rapid increase in indole content. In summary, HDACs may mediate the biosynthesis of quality-related secondary metabolites in response to the continuous wounding during oolong tea processing (Fig. 5). The results of this study broaden our understanding of how histone deacetylation influences the regulation of secondary metabolism in plants, and also provides evidence of the involvement of epigenetic regulation in the production of quality-related secondary metabolites in non-model plants exposed to abiotic stress.
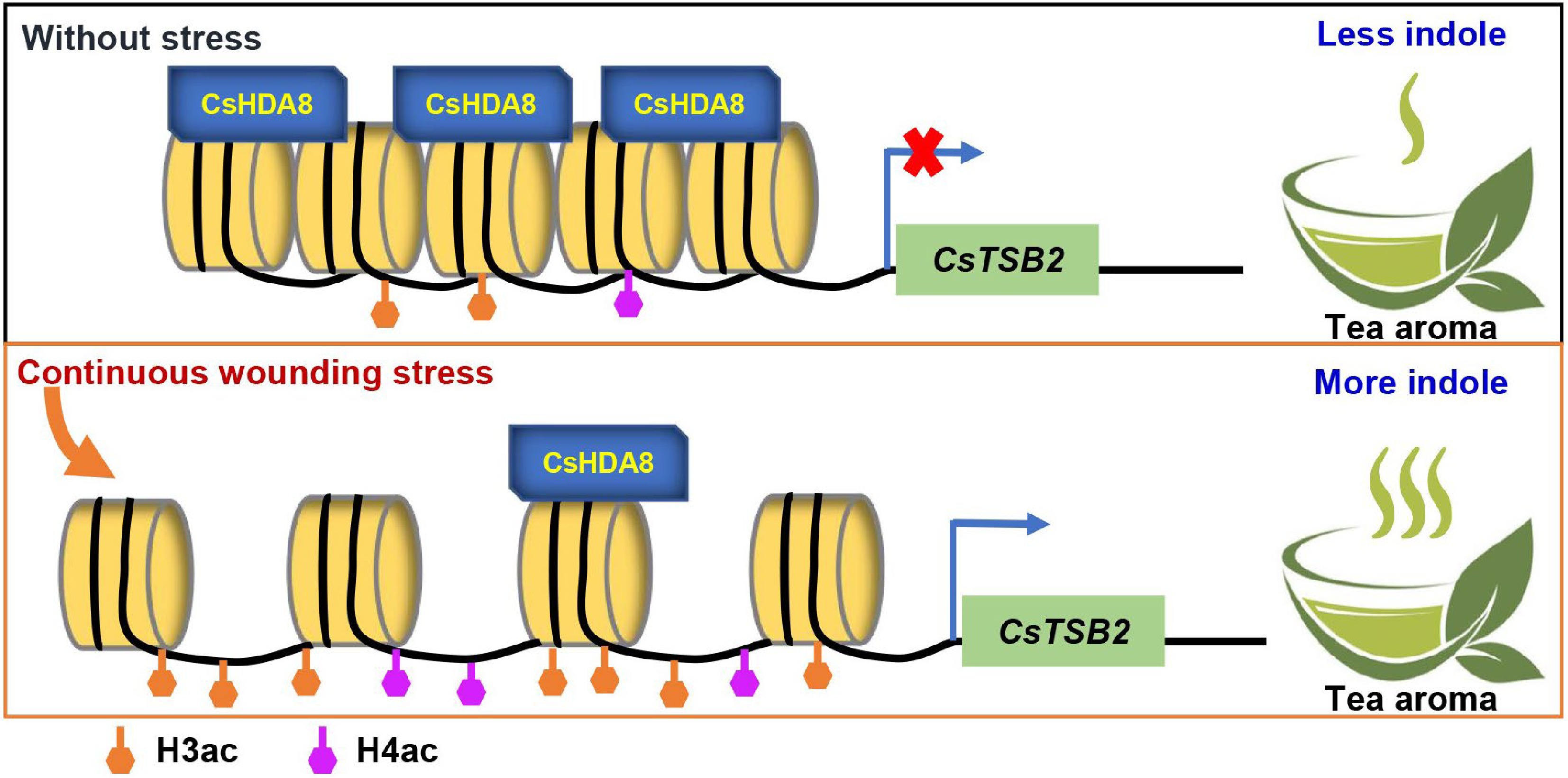
Figure 5.
Mechanism of CsHDA8 involvement in regulating the synthesis of indole induced by mechanical wounding during the processing of oolong tea. H3ac, Histone H3 acetylation; H4ac, Histone H4 acetylation. ×, Repress expression.
-
Tea samples (C. sinensis 'Jinxuan') were obtained from Yingde City (Guangdong Province, China). C. sinensis 'Jinxuan' is widely planted in South China and is often selected for processing into oolong tea. The samples used in the experiments were tea branches containing one bud and two or three leaves. Arabidopsis thaliana (Columbia-0) for subcellular localization experiment was planted in incubator (22 °C temperature, 16 h light/8 h dark photoperiod, and 60% humidity).
Manufacturing processes of oolong tea
-
the method for oolong tea processing refers to the methods used in a previous study[4]. One bud and two and/or three leaves of C. sinensis 'Jinxuan' were used. Samples at eight time points (including freshly plucked (P), solar withering (SW), indoor-withered (IW) and turned over for five times (T1−T5)) were collected. Three independent biological samples were prepared.
Wounding treatment
-
one bud and two and/or three leaves of C. sinensis 'Jinxuan' were used. Plucked samples were placed in a shaker at 25 °C for continuous shaking for 0, 3, and 6 h, respectively. The samples placed in the same environment and kept stationary were used as the control groups. Three independent biological samples were prepared.
Histone acetylation inhibitor treatment
-
method for histone acetylation inhibitor treatment was carried out as per the published reference, with modification[25]. The branches with one bud and two and/or three leaves of C. sinensis 'Jinxuan' were collected and exposed to the following treatments: (1) cultured in methanol-water (1/19, v/v); (2) cultured in combinatorial histone deacetylase inhibitors containing 0.25 mM Trichostatin A (TSA), 0.25 mM suberoylanilide hydroxamic acid (SBHA), 0.25 mM sodium butyrate, and 0.25 mM nicotinamide in methanol-water (1/19, v/v). The above treatments were carried out for 24 h, and the samples were then subjected to wounding treatment, i.e. the samples were placed in a shaker at 25 °C for continuous shaking for 0, 3, and 6 h, respectively. The samples only cultured in methanol-water were set as the control group. Three independent biological samples were prepared.
Extraction and analysis of indole content
-
Method for indole content analysis was carried out as per the published reference, with modification[11]. Tea samples (500 mg) were extracted with 2.7 mL dichloromethane, and 5 nmol ethyl n-decanoate was used as an internal standard. The extracted samples were separated and analyzed on GC-MS QP2010 SE (Shimadzu Corporation, Kyoto, Japan). Authentic standards were used to perform the qualitative and quantitative analyses.
Nuclear protein extraction and immunoblot assay
-
The method for nuclear protein extraction was as referred to in the published reference, with modification[37]. The samples were ground to powder and fully homogenized with cold lysis buffer (20 mM Tris-HCl [pH 7.4], 25% glycerol, 20 mM KCl, 20 mM EDTA, 2.5 mM MgCl2, 250 mM sucrose). Filtration and centrifugation were carried out to obtain nuclear proteins. Finally, the nuclear proteins were resuspended in nuclei storage buffer (20 mM Tris-HCl [pH 7.4], 25% glycerol, 2.5 mM MgCl2, 15.1 g/100 mL sucrose, PI) and 0.2 mg/ml bromophenol blue was added for protein detection. The protein samples were boiled and separated by 10% SDS-PAGE and detected with anti-H3 (catalog no. ab1791), anti-H4 (catalog no. Ab10158), anti-H3ac (catalog no. 06-599), anti-H4ac (catalog no. 06-866) and anti-actin antibodies, respectively. The gray-scale intensity of protein bands were analyzed by Image Lab software (Bio-Rad, USA).
Identification of C. sinensis Histone deacetylase (CsHDACs) and Histone acetyltransferase (CsHATs) genes
-
The reported gene sequences of histone deacetylases (HDACs) and histone acetyltransferases (HATs) in A. thaliana, rice, soybean, tomato and sorghum were obtained from the NCBI database (
www.ncbi.nlm.nih.gov/ ). The downloaded sequences were aligned in the TPIA database (http://tpia.teaplant.org/Blast.html ) to obtain tea CsHDACs and CsHATs sequences, and the repeated sequences were deleted.Phylogenetic construction
-
MrBayes v3.2.7a was used to perform Bayesian inference (BI) of phylogeny using the best-fit model and the Markov Chain Monte Carlo (MCMC) method with 200,000 samplings every 10 generations[38,39]. The phylogenetic trees were viewed and edited using FigTree v1.4.4 (
http://tree.bio.ed.ac.uk/software/figtree ).Analysis of related gene expression levels
-
The method for gene expression analysis was as per the reference[4]. Quick RNA isolation Kit (Huayueyang Biotechnology) was used to obtain the total RNA. PrimeScript® RT reagent Kit (Takara) was used to reverse transcription. Finally, quantitative real-time PCR (qRT-PCR) was used to determine the gene transcription expression level. Supplemental Table S1 shows the qRT-PCR primers. Changes in mRNA levels of related genes were normalized to that of CsEF-1α and CsACTIN1.
Chromatin immunoprecipitation assay
-
The method for chromatin immunoprecipitation (ChIP) assays were as per the reference with modifications[26]. The extracted chromatin was sheared to an average length of 500 bp through sonication, and then immunoprecipitated with a specific antibody anti-CsHDA8 (customized by Huaan Co., Ltd), anti-H3ac (catalog no. 06-599) or anti-H4ac (catalog no. 06-866). The reaction with pre-immune serum anti-IgG (catalog no. 12-370) was used as the negative control. The input sample was equal amounts of the sonicated chromatin solution. The cross-linking was then reversed and the amount of each immunoprecipitated DNA fragment was determined by quantitative PCR using gene specific primers (Supplemental Table S2). The 2−ΔCt (= 2-[Ct(ChIP)-Ct(Input)]) was determined as the input percentage.
Subcellular localization assay
-
The CsHDA8 sequence was fused to the marker gene EYFP and inserted into the pSAT6-EYFP-N1 transient expression vector (forward, 5'-CGGTACCGCGGGCCCGGGATGTGGGGGAGTCCCAACCC-3' and reverse, 5'-CACCATCAGGATCCCGGGGGATCCTTTCAAGAACGGTA-3'). Subcellular localization assay was carried out as per the previously reported method[4]. A. thaliana leaves of approximately 4 weeks old were used for the experiments. Protoplasts were isolated by removing the lower epidermis with tape and then using the prepared enzyme solution (1.5% cellulose R10, 0.3% macerozyme, 0.4 M mannitol, 20 mM KCl, 20 mM MES, 100 mM CaCl2 and 1% BSA). After 2 h of enzymatic digestion, the isolated protoplasts were collected. The samples were then washed with W5 solution and MMG solution was added to complete the protoplasmic preparation. The constructed CsHDA8-YFP plasmid and the nuclear localization vector mCherry were co-transferred into A. thaliana protoplasts using the PEG permeation method. After co-transformation for 12 h in darkness, the signal localization of YFP and mCherry was observed under a confocal microscope (Zeiss LSM 510, Carl Zeiss, Jena, Germany).
Statistical analysis
-
SPSS package (Version 23.0) was used in this study. Differences between the two groups were verified with Two-tailed student's t test, and differences among more than two groups were determined using one-way analysis of variance (ANOVA) follow by Duncan's multiple comparison tests. p ≤ 0.05 was considered as significant.
This study was supported by the financial support from the National Natural Science Foundation of China (31870684), the Foundation of Science and Technology Program of Guangzhou (202102020806), the Key Project of Universities in Guangdong Province (2021ZDZX4066), the Young Elite Scientists Sponsorship Program by China Association for Science and Technology (2020QNRC001), and the Guangdong Provincial Special Fund For Modern Agriculture Industry Technology Innovation Teams (2022KJ120). We thank Dr. Guangyi Dai at the South China Botanical Garden, Chinese Academy of Sciences for the analysis of subcellular location.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Xiaochen Zhou, Jie Yang
- Supplemental Fig. S1 Protein detection results after mechanical wounding treatment in tea plant.
- Supplemental Fig. S2 Phylogenetic analysis of histone deacetylase homolog protein in plants.
- Supplemental Fig. S3 Phylogenetic analysis of histone acetyltransferase homolog protein in plants.
- Supplemental Fig. S4 Changes in expression levels of CsHATs in tea leaves exposed to wounding treatment.
- Supplemental Fig. S5 Changes in expression levels of CsHDACs in tea leaves exposed to wounding treatment.
- Supplemental Figs S6 Changes in expression levels of CsHAG3 and CsHDACs in tea leaves during the manufacturing process of oolong tea.
- Supplemental Fig. S7 Changes in expression levels of CsHAG3 and CsHDACs in tea leaves during the manufacturing process of oolong tea.
- Supplemental Fig. S8 The expression level of CsHDA8 after tea geometrid treatment.
- Supplemental Table S1 The primers used for qRT-PCR in this study.
- Supplemental Table S2 The primers used for ChIP-qPCR in this study.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou X, Yang J, Jian G, Zeng L, Gu D, et al. 2022. Role of histone deacetylase CsHDA8 in regulating the accumulation of indole during the oolong tea manufacturing process. Beverage Plant Research 2:17 doi: 10.48130/BPR-2022-0017 

Role of histone deacetylase CsHDA8 in regulating the accumulation of indole during the oolong tea manufacturing process
- Received: 29 June 2022
- Accepted: 29 August 2022
- Published online: 01 October 2022
Abstract: Aroma is an important quality-related trait of oolong tea. The compounds contributing to oolong tea aromas are mainly produced in response to stress during the enzyme-active stage of the tea manufacturing process. Epigenetic regulation affects stress-induced secondary metabolite production; however, the mechanisms regulating oolong tea aroma compound formation are unclear. In this study, the effect of histone acetylation on indole formation, an important aroma of oolong tea, was revealed and CsHDA8 was identified as a candidate regulator. Additionally, CsHDA8, which was localized in the nucleus and cytoplasm, may function as a histone deacetylase. Moreover, the binding of CsHDA8 to CsTSB2 promoter related to indole decreased during a wounding treatment, which resulted in increased histone acetylation and gene expression. The study results may clarify the regulatory effects of histone acetylation on the formation of tea secondary metabolites, with potential implications for studies on the epigenetic regulation in other non-model horticulture plants.
-
Key words:
- aroma compound /
- Camellia sinensis /
- CsHDA8 /
- histone modification /
- oolong tea /
- stress response