








-
Marine ecosystems, comprising over 80% of lifeforms on earth, have a vast diversity of species. This includes more than 30,000 species of fish related to humans as a source of protein and industrial raw materials. However, less than 10% of marine bioresources are used for human food and other purposes[1]. Recently marine bio-resources have been widely explored as sources of bioactive secondary metabolites for functional food, cosmetic, pharmaceutical, and biomedical research[2]. Thousands of new active substances that exhibit physiological functions such as antioxidant, anti-inflammatory, anticancer, antiaging, antibacterial, and antiviral are being discovered from marine biological resources such as fish and shellfish, seaweed, and marine microorganisms. Still, only a few have been developed for consumer use.
Traditionally, dietary proteins are considered a source of energy and necessary amino acids, both of which are required to develop and maintain physiological homeostasis. Identification and characterization of plant and animal bioactive peptides have garnered considerable attention recently. Bioactive peptides are inert protein fragments inside the parent protein's sequence and exert various physiological functions once they are released by hydrolysis[3]. There is no strict definition for bioactive peptides; it often refers to peptides that have health benefits and are primarily used in functional foods. However, many peptides have good edibility and bioactivities and are employed in various applications, including functional meals, cosmetics, and pet food production[2,4]. These peptides have molecular weights of less than 2500 Da and a size of 2–20 amino acids. The bioactivity of these peptides depends on the composition and amino acid sequence. These peptides may have antihypertensive, antioxidative, hypocholesterolemic, antibacterial, immunomodulatory, opiate-like, mineral binding, and antithrombotic activities based on their structural features, amino acid composition, and sequencing[3]. Figure 1 depicts the spectrum of bioactivities demonstrated by numerous marine derived bioactive peptides. Several reviews have recently been published on biofunctional peptides derived from marine organisms[2,5−10].
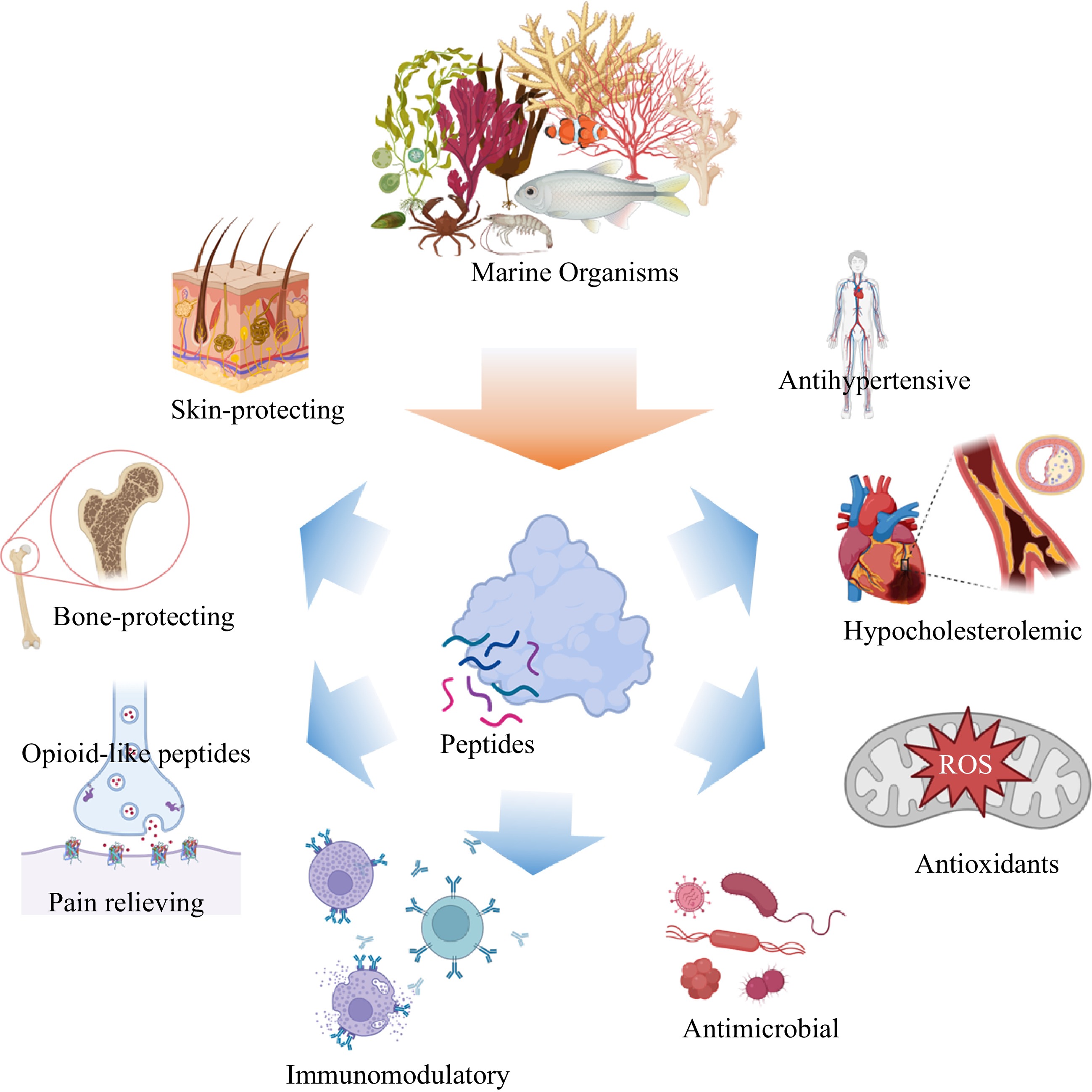
Figure 1.
Spectrum of bioactivities demonstrated by numerous marine derived bioactive peptides.
While it is commonly recognized that marine organisms (fish, shellfish, and algae) offer excellent nutrition, their functional properties have yet to be explored. Several nitrogenous compounds (proteins, peptides, and amino acids) with various biofunctional properties have recently been discovered in fish, shellfish, and seaweed. Furthermore, fish and shellfish, seaweed, and fisheries processing byproducts (waste) provide high-quality protein (10%–47%), making them excellent raw materials to produce functional peptides. Bioactive peptide identification from genomes and transcriptomes of non-model organisms has been made possible using high-throughput sequencing techniques[11]. While bioactive peptides are discovered in marine species, new functional peptides derived by artificially hydrolyzing proteins in marine organisms can be produced in vast quantities.
Fisheries processing by-products (intestines, fins, scales, bones, heads, shells, etc.) that consist of over 25% of the whole fish could be used to generate value-added products[12]. Most by-products are discarded during fisheries processing, causing environmental problems. It should be noted that these byproducts may contain a significant amount of proteins that could be used in generating functional peptides for animal feed production. Collagen, the primary component of fibrous proteins, is a material that has received immense importance due to its nutritional value. Collagen is partially hydrolyzed to produce gelatin, a soluble protein. The rheological properties of gelatin make it a desirable substance for numerous commercial applications. Gelatin is increasingly utilized by the food industry in massive quantities every year. Industrial collagen is mainly produced from cow or pig skin and used in cosmetics, functional foods, and medications. The use of marine-based collagen is acquiring rapid growth due to its unique qualities in comparison to collagen obtained from mammals, such as the minimal risk of disease transmission, lack of religious constraints, cost-effective production method, biocompatibility, low molecular weight, and ease of absorption by the human body[13]. Collagen is not easily hydrolyzed by proteolytic enzymes other than collagenase. Collagen can be a valuable raw material for producing bioactive peptides and can be relatively stable when the produced peptide is ingested[14]. Much of the fisheries' processing byproducts, including skins and bones, are primarily made of collagen . This highlights the importance of analyzing process scenarios and economic feasibility to outline methods to sustainably utilize biomass confining concepts of the circular economy. The present review on bioactive peptides of marine origin discloses the vast potential of value-added applications of protein biomass found in abundance in seaweed, fish and shellfish, microalgae, and by-products of fisheries processing.
-
Bioactive peptides can be produced by hydrolysis of food proteins with acid or alkaline, solvent extraction, enzymatic hydrolysis, microbial fermentation, ultrasound, pulsed electric field, pressurized liquid, and subcritical water extraction techniques[2]. Various problems occur when protein is hydrolyzedwith acid or base. When hydrolyzed with acids, essential amino acids such as tryptophan or cysteine become decomposed, and when hydrolyzed with alkali, toxic compounds such as lysinoalanine are produced. Hence the use of such methods to produce consumer products is prohibited by the Food Sanitation Act. On the other hand, techniques such as ultrasound, pulsed electric field, pressurized liquid, and subcritical water extraction are disadvantageous in terms of scaling-up methods for industrial integration. Hence enzyme assisted protein hydrolysis has been actively conducted targeting the production of bioactive peptides[15].
Peptides are produced from fish and shellfish muscle proteins during post-storage by the action of native proteolytic enzymes such as calpains, which are calcium-dependent neutral cysteine proteases, and cathepsins, which are present in lysosomes[16]. Fermented fish products are popular foods in many countries in Asia. During fermentation, fish proteins are hydrolyzed by native proteolytic enzymes, digestive tract enzymes, and proteases produced by basophils. This fermentation process determines the consistency and quality change of the product. However, due to restrictions imposed by potential risk factors such as basophil enzymes, excessive salt content, and histamine generation, it can be difficult to market these products both at home and overseas.
Using proteolytic enzymes produced from plants, animals, or microbes, it is feasible to hydrolyze the proteins of seaweed, seafood, and by-products of seafood processing in order to create a wide variety of peptides with potential bioactivities. A number of excellent articles discuss current developments in the discovery and potential applications of peptides derived from edible seaweed, fish, and marine invertebrates[2,6,7]. Most of these studies are academic-oriented and carried out on a laboratory scale. Mince of samples in water (crude homogenates) as well as pulverized powder obtained from lyophilized or sun- or air-dried samples, samples with and without pretreatment or further protein enrichment by precipitation (ammonium sulfate, trichloroacetic acid, urea, and organic solvents), has been used for proteolysis and extraction of peptides[2]. These extracts are essentially a mixture comprising a wide array of components other than peptides. Membrane ultrafiltration of protein hydrolysates is generally employed as the first step in the bioassay-guided purification of peptides. After membrane ultrafiltration, the most active fraction is purified further using size-exclusion and/or ion exchange chromatography, which is either led by FPLC or conducted using open-column chromatography. Some studies bypass the ultrafiltration and use column chromatography to purify protein hydrolysates. RP-HPLC has been the last purification step in most studies performed for isolating the peptides, those that are being used for the determination of peptide sequence by either using liquid chromatography-tandem mass spectrometry or by Edman degradation[6].
Enzyme-assisted extraction (batch process) is relatively expensive as a scaleup process compared to other industrially feasible processes such as chemical hydrolysis and fermentation due to the cost of enzymes and maintaining conditions[17]. While batch processes are simple in terms of apparatus, operation conditions, and substrate concentration, they require a lot of enzymes, energy, and labor to operate. The inhibitory action of final products frequently affects the equilibrium and reduces the yield. Moreover, controlling the degree of hydrolysis, enzymatic autolysis, repeatability, and reproducibility for that matter are highly challenging, and the functional properties may vary depending on the batch. To overcome several of the above issues for industries, studies on continuous hydrolysis of proteins using immobilized enzymes have been conducted, but immobilization methods are challenging, and enzymatic activity may decrease (10%~90%) due to steric hindrance caused by immobilization[17]. Alternatively, immobilized enzymes may exhibit better functional properties than their soluble counterparts based on the enzyme used and the immobilization technique[18]. Yet studies in this regime are lacking for marine peptides and should be explored, for it has the potential to develop industrial processes for manufacturing functional peptides.
Numerous methods are being implemented to overcome problems associated with the batch process. Kim et al. report using a recirculating continuous three-stage membrane reactor[15]. In this method, a certain amount of enzyme is repeatedly used without immobilizing the enzyme, thereby continuously producing peptides with different functionalities depending on the molecular weight. An ultrafiltration membrane reactor system has been integrated to improve enzyme utilization efficiency in the 3-stage membrane enzyme reactor device. Despite the repeated re-use of the enzymes for 11 batches, no significant difference in the product quality was observed[19]. The enzyme used to hydrolyze the substrate in the reaction vessel is recycled, and product molecules smaller than the membrane's MWCO are separated from the substrate and the enzyme. The method shortens time, labor, and equipment installation[19].
Collagen peptides from fish are increasingly utilized in numerous biomedical and research applications. Collagen comprises multiple genetically distinct molecular species that collectively include the collagen family. Each molecular species is classified from type I to type X in their order of discovery[13,14]. Fibrous collagen is the most prominent molecular species in the collagen family, accounting for 85%–90% of total collagen, also known as type I collagen. In addition to being the main element of the dermis and tibia, type I collagen is also present in all of the body's organs. It has long been used as a raw material for leather or gelatin. Collagen from fish skin is primarily extracted by chemical hydrolysis using organic acids (generally 0.5 M acetic acid, at 4 °C for 24–72 h)[20−23]. Other than solubilizing non-crosslinked collagens, organic acids can break some of the interstrand crosslinks in collagen, increasing their solubility. Before acid extraction, non-collagenous proteins are removed by NaOH pretreatment (0.5–1.3 N NaOH for 12–36 h). Collagen that is insoluble in acids can be extracted by partial hydrolysis of the cross-linked bonds and peptide bonds by using enzymes such as pepsin, trypsin, alcalase, and papain. The extract is filtered, and collagen in the filtrate is precipitated by adding NaCl. The precipitate collected by centrifugation is re-dissolved in low concentrations of organic acids and gradually dialyzed against much lower concentrations of organic acids, followed by distilled water. Finally, collagen powder is obtained by lyophilization[20−23]. However, the above extraction process is unsuitable on an industrial scale, and the extraction methods must be optimized to maximize the yield.
-
Proteins derived from seaweeds, fish and shellfish, and fishery products are gaining popularity as a rich source of new bioactive peptides with distinct multifunctional properties. In recent years, many bioactive peptides have been discovered in the marine DNA pool, and their structural diversity has been extensively studied[12].
Seaweeds are used as a food material in Asian countries, including Korea, Japan, and China, and are renowned as sources of functional polysaccharides such as agar, carrageenan, fucoidan, and alginic acid. In the west, seaweeds are emerging as a potential source of bioactive peptides as well as a substitute material for meat protein. The protein content of seaweeds varies considerably between species, habitats, amount of sunlight, temperature, and seasons[24]. Red algae with the highest protein content generally consist of up to 47%, green algae 9%–26%, and brown algae 3%–15% from their dry weights. Among the red algae, Porphyra tenera and Palmaria palmata have reported high protein contents of 47% and 35% (w/w), respectively, whereas the green algae, Ulva pertusa, contains nearly 26% (w/w) proteins[5]. Seaweed protein contains all essential amino acids, high in glutamic acid and aspartic acid (7.9%–44.0% w/w), and low in threonine, lysine, tryptophan, histidine, cysteine, and methionine[5]. The content of these amino acids in seaweed is generally higher than in terrestrial plants. Some seaweeds contain relatively high amounts of essential amino acids. Examples include glycine in P. palmata, arginine in Porphyra sp., U. pertusa, and Chomdrus crispus, and proline in Ulva armoricana. Kelp, such as Laminaria digitata has a high content of alanine, and Undaria pinnatifida contains glycine, alanine, arginine, leucine, lysine, valine, and methionine[25].
Fish are the most abundant source of protein in marine life. The total protein content in bony fish muscle is composed of 60%–75% myofibril protein, 20%–35% sarcoplasmic protein, and 2-5% musculoskeletal protein. In cartilaginous fish muscles, sarcoplasmic protein is more abundant, representing about 10%, while in mammalian muscles, myofibril, sarcoplasmic and musculoskeletal protein are about 50%, 30%–35%, and 15%–20%, respectively[26,27]. Processing by-products discharged as waste from the seafood processing plants may contain a large amount of protein (25%–30%). It might be a low-cost, easy-to-obtain raw material for manufacturing protein hydrolysates and peptides[12]. Generally, when raw fish is processed in a fisheries processing facility, residuals such as the head, shell, intestines, and bones are acquired as by-products (residues) after harvesting the fish meat account for more than half of the fish body.
The amount of fish skin varies depending on the type of fish, but it is about 7%–12% depending on the weight of the fish. Hundreds of tons of fish skins are produced annually as by-products at fishery processing plants. The main proteins of fish skin are collagen and elastin, and the composition contains about 90% collagen, 1.5% elastin, and a small number of other glycoproteins[14].
-
It has been found that dietary proteins contain peptides that can influence physiological parameters in vivo. Some parameters include blood pressure, insulin, glucose homeostasis, blood cholesterol control, and immune function[3,4].
Previously fish protein hydrolysates were primarily utilized to make low-value animal feed for nutritional supplementation, fish feed, and seasoning to enhance food flavor. Southeast Asian, Scandinavian, and Eskimo societies have used fermented fish sauces or pastes as staple foods or flavoring[28]. Heart disease, diabetes, and cancer can be mitigated by consuming a variety of marine protein hydrolysates and peptides, such as those found in seaweed, fish, shellfish, and fish processing byproducts. They became available as functional food ingredients as scientific evidence revealed their role in the prevention and management of obesity-related chronic diseases[8]. Protein hydrolysates and peptides derived from marine organisms have various physiological functions. Marine peptides such as Tilapia piscidin 4, Kunitz-type peptides from Sea anemone (Heteractis crispa), and Epinecidin-1 from Orange-spotted grouper (Epinephelus coioides) are reported to contain multiple biological functions suggesting their potential therapeutic benefits[29−31].
Antihypertensive activity
-
Hypertension is a hallmark of lifestyle-related disorders such as cardiovascular, chronic kidney diseases, and dementia and is one of the major causes of death today. Systolic blood pressure (> 115 mmHg) was projected to be responsible for 8.5 million deaths in 2015. Hypertension affects an estimated 1.28 billion people worldwide between the ages of 30 and 79, with two-thirds of those individuals residing in low- and middle-income nations. It is anticipated that by 2025, 1.06 billion individuals over the age of 20 will have hypertension[32]. Essential or primary hypertension is the most common chronic disease and is considered a risk factor for cardiovascular disease, stroke, and renal failure. Angiotensin-converting enzyme (ACE), which is present in the lungs, blood vessel walls, brain, and kidneys, is a dipeptidyl-carboxypeptidase, an enzyme that cleaves peptides at the C-terminus into a dipeptide unit. ACE converts angiotensin I into angiotensin II, an active vasoconstrictor that increases blood pressure. Further, ACE decomposes and inactivates bradykinin, which has arterial relaxation and blood pressure-lowering effects, thereby increasing blood pressure[33]. Hence, ACE inhibitors suppress these responses and lower blood pressure by vasodilation, making them useful as hypertension medications.
The first ACE inhibitory peptide has been identified from a dietary protein hydrolysate produced by a microorganism-derived collagenase. Since then, ACE inhibitory peptides have been extracted from various food protein digests due to the comparatively simple method for assessing the ACE inhibitory activity. Marine organisms such as katsuobushi, tuna, sardines, salmon, laver, seaweed, and krill have been found to contain many ACE inhibitory peptides (Table 1)[34]. These ACE inhibitory peptides have a relatively low molecular weight and short amino acid sequences. However, only a few of these peptides reduce blood pressure when fed to humans. Saito & Hagino (2005) extracted ACE inhibitory peptides from enzymatic digests of various dietary proteins and investigated their ability to lower blood pressure[34]. It was found that ACE inhibitory activity did not necessarily result in a lowering of blood pressure. While there are many ACE-inhibitory peptides, some show only enzyme-inhibitory activity but not blood pressure-lowering properties since the broad substrate-specificity of ACE[34].
Table 1. ACE inhibitor peptides obtained from marine organisms.
Peptide sequence Bioactivity * (ACE inhibition IC50; μM) Reference Cuttlefish (Sepia officinalis) crude protease (from Bacillus mojavensis A21) hydrolysate Ala-His-Ser-Tyr 11.6 [35] Gly-Asp-Ala-Pro 22.5 Ala-Gly-Ser-Pro 37.2 Asp-Phe-Gly 44.7 Cuttlefish (S. officinalis) muscles hepatopancreas enzyme hydrolysate Ala-Phe-Val-Gly-Tyr-Val-Leu-Pro 25.66 [36] Glu-Lys-Ser-Tyr-Glu-Leu-Pro 14.41 Val-Glu-Leu-Tyr-Pro 5.22 (non-competitive) and lowering of systolic blood pressure in spontaneously hypertensive rats (SHR) Lizard fish (Saurida elongata) muscle protease hydrolysate Gly-Met-Lys-Cys-Ala-Phe 45.7 ± 1.1 [37] Arg-Val-Cys-Leu-Pro 175 [38] Grass carp (Ctenopharyngodon idella) muscle alcalase hydrolysate Val-Ala-Pro 18.6 (competitive) [39] Atlantic salmon (Salmo salar) skin collagen hydrolyzed by Alcalase followed by papain Ala-Pro 0.060 ± 0.001 mg/mL [40] Val-Arg 0.332 ± 0.005 mg/mL Skipjack (Katsuwonus pelamis) roe alcalase hydrolysate Asp-Leu-Asp-Leu-Arg-Lys-Asp-Leu-Tyr-Ala-Asn 67.4 [33] Met-Cys-Tyr-Pro-Ala-Ser-Thr 58.7 Met-Leu-Val-Phe-Ala-Val 3.07 Ser-Pro 0.06 ± 0.01 mg/mL [41] Val-Asp-Arg-Tyr-Phe 0.28 ± 0.03 mg/mL Val-His-Gly-Val-Val 0.90 ± 0.16 mg/mL Tyr-Glu 0.80 ± 0.03 mg/mL Phe-Glu-Met 2.18 ± 0.20 mg/mL Phe-Trp-Arg-Val 0.76 ± 0.10 mg/mL Pacific cod (Gadus macrocephalus) skin gelatin pepsin hydrolysate Gly-Ala-Ser-Ser-Gly-Met-Pro-Gly 6.9 [42] Leu-Ala-Tyr-Ala 14.5 Flounder fish (Paralichthys olivaceus)
muscle pepsinMet-Glu-Val-Phe-Val-Pro 79 (competitive) [43] Val-Ser-Gln-Leu-Arg-Thr 105 (non-competitive) Haruan (Channa striatus) Thermolysin hydrolysate Val-Pro-Ala-Ala-Pro-Pro-Lys 0.45 [44] Asn-Gly-Thr-Trp-Phe-Glu-Pro-Pro 0.63 U. pinnatifida (wakame) peptide from hot water extract Ile-Tyr, Phe-Tyr, Tyr-His, Lys-Tyr Respective IC50 values are 2.7, 3.7, 5.1, 7.7 [45] Jellyfish (Rhopilema esculentum) alcalase hydrolysate Val-Lys-Pro and Val-Lys-Cys-Phe-Arg Respective IC50 values are 1.3 and 34.5 [46] Blue mussel (Mytilus edulis) protein hydrolyzates Ile-Lys, Tyr-Glu-Gly-Asp-Pro, Trp-Phe,
and Ser-Trp-Ile-Ser-SerRespective IC50 values are 0.77, 0.19, 0.40, and 0.32 mg/mL [47] in vitro gastrointestinal digest of Isochrysis zhanjiangensis Glu-Thr-Thr 15.08 (non-competitive) [48] Sonicated Spirulina sp. protease K digest TVLYEH and LQAGGLF 2.88 and 66.83 (competitive) [49] * The IC50 value of ACE inhibitory activity is given in the unit μM if not specified otherwise. Information acquired from in vitro inhibition, and molecular docking investigations indicates peptides' potential for treating hypertension. Before clinical trials, the in vivo bioactivity assessment using the SHR model is a key step in demonstrating the antihypertensive effectiveness of ACE-inhibiting protein hydrolysates or peptides. Protein hydrolysates or peptides of many seaweeds, fish and shellfish, and seafood processing by-products have shown antihypertensive effects in SHR. A peptide derived from the fermented blue mussel M. edulis, EVMAGNLYPG, at 10 mg/kg, was found to lower systolic blood pressure by 25 mmHg units compared to the control group[50]. It is slightly less potent than the ACE inhibitor Captopril in reducing systolic blood pressure. After hydrolyzing katsuobushi (dried bonito flakes) with various proteolytic enzymes and analyzing the ACE inhibitory activity of the hydrolyzed product, the hydrolyzed product of thermolysin revealed the best ACE inhibitory action (IC50 = 29 µg/mL)[51]. This substance (250 mg/kg) had a considerable anti-hypotensive effect when given orally to SHR[52].
There have been only a few clinical trials involving marine ACE inhibitory peptides, although several have shown antihypertensive effects in the SHR model. When the hydrolysate of seaweed (U. pinnatifida) protein extract was administered to two groups of hypertensive subjects for more than 8 weeks (300 mg/day), the systolic blood pressure was significantly reduced by 11 mmHg units compared to the control group. An equivalent reduction was observed for the 500 mg/day group after 5 d of treatment[53]. Per another study, when hypertensive participants were given 1.6 g of oligopeptide derived from Parphyra yezoensis for 12 weeks, it showed a substantial antihypertensive impact after 8–12 weeks[54]. In a clinical trial, the antihypertensive effects of katsuobushi oligopeptides (1.5 g/day) were compared in a randomized, double-blind, cross-over study with 65 borderline and moderately hypertensive subjects who indicated systolic and diastolic blood pressure higher than 130 and 85 mmHg. Katsuobushi oligopeptide fractions obtained by ultra-filtration showed stronger antihypertensive activity with no side effects. The consumption of 1.5 g of the ultra-filtered fraction significantly decreased blood pressure by inhibiting ACE[52]. Based on the findings of this study, katsuobushi oligopeptides containing functional food were given governmental approval as an antihypertensive food[55].
Although the relationship between ACE inhibitory peptide structure and activity has not been fully understood, various structural characteristics have been discovered to influence its ACE inhibitory function. According to the literature following trends are apparent in ACE inhibitory peptides[56−59].
Characteristics of a peptide that preferentially influences the ACE inhibitory activity:
● Dipeptides with Arg are observed to be more potent ACE inhibitors (Pro-Gly has indicated an IC50 of < 1,000.00 µM while Pro-Arg has indicated an IC50 of 4.10 µM. Tyr-Pro has shown an IC50 of 720.00 µM for ACE inhibition, while Tyr-Pro-Arg has demonstrated an IC50 of 16.50 µM).
● Positively charged amino acids are in the middle of the peptide. In long-chain peptides, the peptide conformation, or the geometric structure at the specific binding site of ACE will greatly influence its ACE inhibitory function.
● Dipeptides having amino acid residues with hydrophobic or bulky side chains are preferred ACE inhibitors[57].
● Tripeptides could be more potent ACE inhibitors compared to dipeptides.
Characteristics at the C-terminus of a peptide that preferentially influence the ACE inhibitory activity:
● Peptides containing three hydrophobic (aromatic) amino acid residues at the C-terminus[35,57].
● Predominant content of Ile and Val (branched-chain aliphatic amino acids) and C-terminal Leu.
● A positive charge on the amino or guanidine terminus of C-terminal Lys or Arg side chains[58,59]. (The removal of Arg from ALPMHIR (IC50 of 42.60 µM) has produced less-active analogs, ALPMH (IC50 of 521.00 µM), and the removal of R from AVPYPQR (IC50 of 15.00 µM) have produced AVPYP (IC50 of 80.00 µM)).
Characteristics at the N-terminus of a peptide that influence the ACE inhibitory activity:
● Tyr, Val, Phe, Trp, Lys, and Ile (hydrophobic amino acids) are generally found in the N-terminus of ACE inhibitory di- and tripeptides, and they indicate IC50 values within the range of 1.5–45.0 M.
Mediators of the renin-angiotensin system (RAS) pathway and ACE inhibitors are used to treat hypertension and heart disease. Synthetic ACE inhibitors like Captopril, Enalapril, Alacepril, and Lisinopril are commonly used to treat and prevent hypertension. However, they can produce side effects, including coughing, allergic reactions, loss of taste, and skin rashes. Thus, natural compounds have become increasingly important as medicinal agents for treating and controlling hypertension.
Antioxidant activity
-
Most living organisms maintain life with the energy obtained by oxidizing oxygen through respiration. Toxic and reactive compounds, also known as reactive oxygen species (ROS) that inevitably harm cells, are produced as byproducts of metabolic processes that require oxygen. ROS, including free radicals, damages cells and organelles and causes oxidation of amino acids and protein degradation. Moreover, ROS causes oxidative degradation of nucleotides and base modification, resulting in nuclear damage. This damage leads to cellular apoptosis or mutations. ROS also affects aging. Free radical theory dictates that aging results from cumulative oxidative damage to the body's cells and tissues, primarily caused by aerobic metabolism. The production of free radicals is promoted with increasing age. On the other hand, interactions between transition metal ions and lipid hydroperoxides result in excessive free radical production, which promotes lipid oxidation[60]. Many chronic diseases are associated with oxidative stress, including heart disease, stroke, arteriosclerosis, diabetes, and hypertension. Other than implications for humans, deterioration of food quality caused by the oxidation of unsaturated fatty acids is a key issue in the food industry. Antioxidants such as butylated hydroxytoluene, tert-butylhydroquinone, and propyl gallate or metal ion chelators have been added to food to suppress fatty acid oxidation; however, their use has been restricted due to their harmful side effects[61]. Therefore, it is crucial to develop natural antioxidants that are safe for health to cope with oxidative stress and improve food quality.
The algae form part of a diverse group of marine organisms that have evolved complex biochemical pathways to survive in highly competitive ocean conditions. These severe environmental variables, such as depth, irradiation, dehydration, freezing, low temperature, heavy metals, and salinity, stimulate the generation of excess reactive oxygen species (ROS) in seaweed cells[62]. To counteract this, seaweeds have naturally evolved with antioxidants and antioxidant enzymes such as catalase, superoxide dismutase, glutathione reductase, and glutathione peroxidase.
While the relationship between amino acid type, position, and hydrophobicity in peptides and antioxidant activity is not yet apparent, it is believed to have a profound effect on the peptide's antioxidant activity. Amino acids such as His, Tyr, Leu, Met, and Cys are generally recognized as antioxidants, and these amino acids are known to provide protons to electron-deficient radicals indicating radical scavenging activity[3,12]. Chelation of metal ions is another aspect that would inhibit lipid oxidation. Generally, peptides containing histidine have been shown to behave as metal ion chelators[60]. Furthermore, antioxidative peptides protect cells from ROS damage by inducing genes that encode antioxidative enzymes. For example, the dipeptide Met-Tyr derived from sardine muscle stimulates heme oxygenase-1 (HO-1) and ferritin in endothelial cells[63]. The peptide purified from the skin gelatin tryptic hydrolysate of Hoki (Johnius belangerii) increases superoxide dismutase, catalase, and glutathione peroxidase in Hep3B cells[64]. Hydrophobic amino acids such as Gly, Val, Ala, Pro, and HyPro are abundantly present in gelatin. These amino acids have a high affinity for the lipid system and thus have a strong inhibitory effect on lipid peroxidation[8]. Therefore, it is presumed that fat-soluble radicals (e.g., hydrophobic peroxyl radicals) generated by the oxidative attack of unsaturated fatty acids are neutralized by antioxidant peptides containing hydrophobic amino acids. Molecular weight is a deterministic factor of a peptide that determines its antioxidant potential.
Table 2 summarizes several types of algae, shellfish, and fish-derived peptides that exhibit radical scavenging activity. As reported in previous studies, peptides have shown better, or similar antioxidant activity compared to α-Tocopherol and synthetic antioxidants such as butylated hydroxytoluene[65,66].
Table 2. Antioxidant peptides obtained from marine organisms.
Peptide sequence Bioactivity Reference Jumbo squid (Dosidicus gigas) skin gelatin tryptic hydrolysate P1: Phe-Asp-Ser-Gly-Pro-Ala-Gly-Val-Leu
P2: Asn-Gly-Pro-Leu-Gln-Ala-Gly-Gln-Pro-Gly-Glu-ArgLipid peroxidation inhibition is higher than α-tocopherol. Hydroxyl radical scavenging IC50 = 90.90 (P1) and 100.72 μM (P2). Improve the viability of t-Butyl hydroperoxide-induced lung fibroblasts. [60] Alaska pollack skin gelatin serial hydrolyzed
by Alcalase, Pronase E, and collagenase in
a three-step recycling membrane reactor.Gly-Pro-Hyp-Gly-Pro-Hyp-Gly-Pro-Hyp-Gly-Pro-Hyp-Gly High antioxidant activity based on TBA assay. Increase viability in tertbutylhydroperoxide (t-BHP)-induced rat liver cells. [67] Giant squid (D. gigas) muscle protein ultrafiltration Asn-Ala-Asp-Phe-Gly-Leu-Asn-Gly-Leu-Glu-Gly-Leu-Ala Improve the viability of cytotoxic embryonic lung fibroblasts by a significant margin at a concentration of 50 g/mL. Suppress radical-induced oxidation of membrane lipids. Radical scavenging activity carbon centered (IC50 396.04 μM), hydroxyl (IC50 497.32 μM), and superoxide radicals (IC50 669.34 μM). [65] Asn-Gly-Leu-Glu-Gly-Leu-Lys Significantly enhance the viability of cytotoxic embryonic lung fibroblasts at 50 μg/mL. Suppress radical-induced oxidation of membrane lipids. Radical scavenging activity carbon centered (IC50 396.04 μM), hydroxyl (IC50 497.32 μM), and superoxide radicals (IC50 669.34 μM). Fermented marine blue mussel
(Mytilus edulis)His-Phe-Gly-Asp-Pro-Phe-His IC50 values for radical scavenging activity, superoxide (21 μM), hydroxyl (34 μM), carbon-centered (52 μM), and DPPH radicals (96 μM). Fe2+ chelating effect. Increase t-BHP-induced WI-38 cell viability. [66] Hard-shelled mussel (Mytilus coruscus)
papain hydrolysateSer-Leu-Pro-Ile-Gly-Leu-Met-Ile-Ala-Met In AAPH administered mice, inhibited malondialdehyde levels per thiobarbituric acid reactive substances assay, and increased superoxide dismutase and glutathione-s-transferase activities. [68] Krill (Euphausia superba) protein pepsin hydrolysate Ala-Met-Val-Asp-Ala-I le-Ala-Arg IC50 value of 0.87 for DPPH radical scavenging assay and ORAC value of 1.56 mM Trolox equivalents/mM peptide. Reduce H2O2-induced oxidative stress in Chang liver cells by increasing superoxide dismutase, glutathione peroxidase, and catalase activities. [69] Antimicrobial activity
-
Antimicrobial peptides are one of the main factors of innate immunity of living organisms, and they play a crucial role in the front line of defense. Pioneering research has been undertaken to show the antimicrobial action of peptides present in humans, animals, and amphibians. In its evolution, the amino acid sequences of antimicrobial proteins and peptides evolved by combating pathogenic microorganisms and have three-dimensional structures of interest[70]. Furthermore, it is recognized that the degradation of microbe cell membranes is the primary method by which peptides exert their antimicrobial activity. However, the precise mechanism of action of specific peptides is unclear. In addition to membrane degradation, modes of action targeting intracellular molecules of microbes have recently been discovered[70]. Antiviral activity of these peptides proceeds by either blocking viral or cell membrane receptors, inhibiting or modulating the host cell’s gene expression profile. Most antibacterial peptides are cationic and act by disintegrating the lipid bilayer of the bacterial cell membrane. The amphipathic structure of these peptides facilitates their ability to penetrate the cytoplasmic membrane of microorganisms by attachment and destabilization[71]. Some peptides are found to modulate DNA replication, transcription, and protein synthesis in host cells, while some could kill antibiotic-resistant bacteria.
Similarly, antifungal peptides are effective at damaging the fungal cell wall. Antiparasitic peptides act by killing cells via interacting with the cell membrane. Physicochemical properties such as charge, length, helicity, hydrophobicity, solubility, and amphipathicity affect the activities of these antimicrobial peptides. DRAMP lists 22,407 entries of antimicrobial peptides to date (
http://dramp.cpu-bioinfor.org ). Most eukaryotic antimicrobial peptides can be categorized under one of the following groups: (a) cysteine deficient linear, α-helical peptides, (b) extended linear peptides, with an unusual proportion of specific amino acids (often proline, histidine, tryptophan, or glycine), or (c) looped peptides with disulfide bonds[71].Antibiotic use in feed additives has been linked to the development of antimicrobial resistance, which can increase disease morbidity[70]. Despite the limited number of in vivo studies, antimicrobial peptides appear to be a viable alternative to antibiotics in animal nutrition as they would increase feed efficiency while reducing certain animal diseases. Antimicrobial peptides are extensively researched for industrial applications, and contribute to the field of drug development as antibiotic alternatives. Furthermore, antimicrobial peptides are explored for use as food additives to prevent food spoilage in place of organic acids such as sorbic acid and sodium acetate. Table 3 discloses facts on marine-derived antimicrobial peptides obtained from a wide variety of marine invertebrates, such as crabs, oysters, lobsters, shrimps, and sea urchins.
Table 3. Antimicrobial peptides derived from marine organisms.
Peptide sequence Bioactivity Reference American lobster (Homarus americanus) hemocytes Gln-Tyr-Gly-Asn-Leu-Leu-Ser-Leu-Leu-Asn-Gly-Tyr-Arg Effective against some gram-negative bacteria. Indicate protozoastatic activity against ciliate parasites. [72] Rainbow trout (Oncorhynchuss mykiss) omBD-1 ASFPWACPSLNGVCRKVCLPTELFFGPLGCGKGFLCCVSHFL β-defensin-like peptide sequence. Provide resistance against viral hemorrhagic septicemia rhabdovirus (VHSV) infection in fish cell line Epithelioma papulosum cyprinid. [73] Channel catfish(Ictalurus punctatus) HbβP-1 AANFGPSVFTPEVHETWQKFLNVVVAALGKQYH Homologous to β-chain of hemoglobin. Antiparasitic activity against Ichthyophthirius multifiliis and Aeromonas hydrophila infection in channel catfish. [74] Hemocyte extracts of Spider
crab (Hyas Araneus)MERRTLLVVLLVCSCVVAAAAEASPSRWPSPGRPRPFPGRPKPI Arasin 1, effectively inhibits the growth of Corynebacterium glutamicum, Listonella anguillarum, and E. coli. [71] The skin mucus of many organisms contains antimicrobial peptides as a primary defense against invading pathogens. The skin of catfish (Pelteobagrus fulvidraco) reports containing antimicrobial peptides, which function as the first line of defense against pathogens. Pelteobagrin (amino acid sequence: GKLNLFLSRLEILKLFVGAL) is an antimicrobial peptide isolated from P. fulvidraco mucus that exhibits antimicrobial activity against fungi, Gram-positive and Gram-negative bacteria[75]. Pelteobagrus has been confirmed to kill bacteria by acting on the cell wall or membrane of bacteria and is expected to have the potential to be applied in the aquaculture and functional foods industry. Similarly, the skin mucus of Japanese eel (Anguilla japonica) is reported to contain antibacterial peptides (GCPQTPRCTNYAEKGXCPPN) that could effectively inhibit the growth of bacteria, A. hydrophila, Edwardsiella tarda, Aeromonas sp. and Micrococcus luteus[76]. Epinecidin-1 (FIFHIIKGLFHAGKMIHGLVTRRRH), derived from Orange-spotted grouper (E. coioides), is a potent multi-functional antimicrobial peptide that has antibacterial, antifungal, antiprotozoal, antiviral, immunomodulatory, anticancer, and wound healing properties[29]. This peptide has much potential for future research and medication development. Epinecidin-1 is assessed in Phase II clinical studies to treat venous leg ulcers. Myticalins are another group of antimicrobial peptides found in order Mytiloida first discovered from Mytilus galloprovincialis[11]. They are rich in proline and, like many other proline-rich antimicrobial peptides, exhibit a non-lytic mode of action by penetrating the bacterial cytosol and interfering with ribosomal protein synthesis. As such, it is essential to develop improved antibacterial peptides that can outperform the resistance caused by pathogenic microorganisms compared to traditional antibiotics. Marine-derived antimicrobial peptides are promising candidates with a broad spectrum of antimicrobial activity. However, these peptides are rarely used as antibiotics.
Skin protective activity
-
The use of cosmetics has become a daily routine, with the core ideology being personal care and morale-boosting. Environmental pollutants are becoming crucially influential in premature aging, wrinkling, uneven tone, and dark spots. Natural extracts with antioxidant properties have gained increased attention due to their safety despite active synthetic chemicals in contemporary cosmetics. Peptides that show antioxidant activities could be potent anti-aging cosmetics. The cosmeceutical potential of peptides derived from marine organisms has been the subject of several recent reviews[9,77]. Table 4 discloses specific bioactivities attributed to marine peptides that render them potential ingredients for cosmetics manufacturing.
Table 4. The cosmetic potential of marine peptides and protein hydrolysates.
Protein source and hydrolysis method Bioactivity Reference Chlorella-derived peptides hot water extract Reduce UVB-induced production of MMP-1 MCP-1, expression of CYR61, c-fos, c-jun, and increase procollagen and TbRII expression, in skin fibroblasts via attenuating AP-1 expression and production of CYR61 and MCP-1. [78] Tuna roe trypsin and alkaline 1:2 hydrolysate. Peptides Ile-Cys-Arg-Asp and Leu-Cys-Gly-Glu-Cys Inhibit UVB-induced apoptosis and alter Keap1/Nrf2-ARE pathway in HaCaT keratinocytes and mice. Inhibit the release of the proinflammatory cytokines. [79] Simulated gastrointestinal digest of grass carp fish scale gelatin hydrolysate (Phe-Thr-Gly-Met-Leu) Tyrosinase inhibition with an IC50 value of 1.89 mmol/L and inhibition of melanin production in the zebrafish model. [80] Skin collagen of Atlantic Codfish (Gadus morhua). Water retaining ability. No toxicity or inflammatory response in HaCaT cells. [77] Tilapia collagen nanofibers Significantly increase HaCaT cell proliferation and accelerates epidermal differentiation through upregulating involucrin, filaggrin, and type I transglutaminase expression. Accelerate wound healing in Sprague–Dawley rats by promoting re-epithelialization. [20] Type I collagen from Magalaspis cordyla and Otolithes ruber bones Accelerate wound healing in Wistar rats [21] Tilapia and grey mullet collagen Enhances cell adhesion, antibacterial activity (Streptococcus mutans, Staphylococcus aureus, E. coli and Bacillus subtills), faster wound healing in Sprague Dawley male rats. [22] Mrigal carp (Cirrhinus cirrhosus) scale collagen Efficient cell growth and proliferation on the collagen sponge. Fibroblast and keratinocyte co-culture indicated the development of a stratified epidermal layer. Faster wound healing, dermal reconstitution, and re-epithelialization in Wistar rats. [81] Low-molecular-weight collagen peptides from Starfish (Asterias pectinifera) Promotes wound healing, bone regeneration, and skin protection. Reduce MMP-1 expression caused by ultraviolet radiation-induced photoaging in CCD-986sk human dermal fibroblasts. [23] Collagen is an imperative component for maintaining skin health. It is a central extracellular matrix element, representing 75% of its weight[82]. Collagen forms a fibrous network supporting keratinocytes, fibroblasts, melanocytes, immune system cells, other fibers, hyaluronic acid, reticulin, and elastin. During skin aging, the network of dermal collagen fibers becomes more disordered. Additionally, metalloproteinases, which break down collagen fibers, are produced at higher rates, which reduces the creation of new extracellular matrix components. Clinical evidence suggests that collagen consumption (especially with high Hyp, Gly, and Pro contents) improves skin elasticity, hydration, and dermal density, reducing wrinkles without adverse side effects[82].
Moreover, several studies have reported that low molecular weight peptides from collagen hydrolysates of marine organisms increase fibroblast proliferation, stimulate the formation of connective tissue components, and provide protective effects against UV and other harmful environmental factors[9]. Recent studies show that the Nile tilapia skin could be successfully used as a xenograft for treating burns in human skin[83]. A wide array of studies was recently undertaken to explore the uses of tilapia skin collagen in biomedical engineering[84,85]. Aminated collagen synthesized from collagen purified from the fish, Larimichthys crocea, has been used to manufacture a hybrid hydrogel dressing containing oxidized sodium alginate and antimicrobial peptides (bacitracin and polymyxin B sulfate)[86]. The dressing had a similar strain to human skin, adjustable gelation time, and stable rheological properties. Besides the antimicrobial activity, the dressing increased migration, the proliferation of cells, and accelerated angiogenesis in vitro, indicating that it could enhance collagen deposition and tissue formation, thereby accelerating epithelial regeneration. Although much information is available on marine peptides and their cosmeceutical potential, many species remain underutilized.
Appetite regulating activity
-
Peptides derived from marine organisms have indicated various other physiological functions. Among them is the appetite-regulating function, where peptides from marine organisms aid in alleviating chronic diseases such as obesity, type-two diabetes, cardiovascular disease, and cancer. Cholecystokinin (CCK) is an important hormone that modulates appetite and hunger. CCK content can be used as a biomarker for evaluating satiety in vivo. Numerous studies have shown that marine peptides could modulate CCK production. The release of CCK from gut endocrine cells was shown to be significantly stimulated by a low molecular weight peptide (1.0–1.5 kDa) extracted from a hydrolysate of shrimp head protein[87]. In addition, it was confirmed that plasma glucose decreased when the protein hydrolysate of P. yezoensis was added to rat feed[88]. As such, it is expected that products containing protein-based bio-functional ingredients may assist in preventing and managing obesity.
Bone-protecting activity
-
A healthy bone structure depends on a balance between bone formation and resorption. Aging is a major contributor to bone mass and strength decline, leading to osteoporosis. Globally more than 200 million people have osteoporosis[89]. A growing body of evidence suggests that peptides derived from edible marine organisms are related to bone health. The tetrapeptide, Met-Pro-Asp-Trp derived from Nannochloropsis oculata by alcalase hydrolysis, induced the differentiation of osteoblasts through Smad and mitogen-activated protein kinases signaling pathway in human osteoblastic MG-63 cells and mouse mesenchymal stem cells[90]. Peptides derived from blue mussel (Mytilus edulis) (PIISVYWK and FSVVPSPK), Belanger's croaker (Johnius belangerii) bone pepsin hydrolysate (KSA), and Ark shell protein (AWLNH and PHDL) report to promote osteogenic differentiation of pre-osteoblasts through activation of the canonical Wnt/β-catenin signaling[91−93].
According to Kim et al., peptides derived from tuna pyloric caeca crude enzyme fish skin gelatin increase calcium absorption in calcium-deficient rats and increase the calcium deposition and strength of the femur[94]. A novel calcium-binding peptide (Val-Leu-Ser-Gly-Gly-Thr-Thr-Met-Ala-Met-Tyr-Thr-Leu-Val) isolated from the pepsin hydrolysates of pollock bone was recovered by hydroxyapatite affinity chromatography. It showed high affinity and calcium solubilizing effects similar to casein phosphopeptide[95]. Peptide (Val-Leu-Ser-Gly-Gly-Thr-Thr-Met-Tyr-Ala-Ser-Leu-Tyr-Ala-Glu) purified via hydroxyapatite affinity chromatography of hoki (J. belangerii) frame by-products (flesh and skeleton) pectinolytic hydrolysates indicated increased calcium-binding activity and bioavailability[96].
Cardioprotective activity
-
It has been reported that protein hydrolysates and peptides derived from seaweed, fish, shellfish, and fishery by-products exhibit anticoagulant activity[97]. In various animal trials, samples containing protein hydrolysates have dramatically lowered total cholesterol, free cholesterol, triglyceride, and phospholipid levels[45]. Anticoagulant protein with a molecular weight of about 26.0 kDa was isolated and purified from the blood ark shell (Scapharca broughtonii), inhibited coagulation factor IX in the intrinsic blood coagulation pathway, and inhibited blood coagulation by significantly delaying the blood clotting time[98].
A single-chain monomer protein with a molecular weight of 12.01 kDa extracted from yellowfin sole (Limanda aspera) using α-chymotrypsin inhibited blood clotting activator XII. It indicated an aggressive action on integrin, a platelet membrane glycoprotein involved in platelet aggregation, thereby inhibiting blood coagulation[97]. Finding a study that reports the anticoagulant activity of peptides derived from marine organisms or protein hydrolysates in vivo was challenging.
Anticancer activity
-
Enzymatic hydrolysates and peptides of marine proteins have been discovered to possess anticancer properties. Oyster (C. gigas) protein Bacillus sp. SM98011 protease hydrolysates have dose-dependently inhibited the growth of transplantable sarcoma-S180 in BALB/c mice while increasing natural killer cells activity, lymphocytes proliferation in the spleen, and phagocytic rate of macrophages[99]. Herein the antitumor activity is thought to result from the immunostimulatory effects. Giant squid (D. gigas) gelatin Esperase hydrolysate has shown anti-proliferative effects in human breast cancer (MCF-7) and glioma (U87) cell lines with IC50 values 0.13 and 0.10 mg/mL, respectively[100]. Steam-cooked tuna fish (Thunnus tonggol) dark muscles papain and protease hydrolysates indicated antiproliferative activities against the MCF-7 cell line[101]. Purified peptides Leu-Pro-His-Val-Leu-Thr-Pro-Glu-Ala-Gly-Ala-Thr and Pro-Thr-Ala-Glu-Gly-Gly-Val-Tyr-Met-Val-Thr indicated antiproliferative effects with respective IC50 values of 8.1 and 8.8 μM. The hexapeptide (Phe-Ile-Met-Gly-Pro-Tyr, FIMGPY) isolated from the hydrolysate of cartilage protein from Skate (Raja porosa) indicated anti-proliferation effects in HeLa cells (IC50 of 4.81 mg/mL) and induced apoptosis[102]. The peptide tilapia piscidin 4 (TP4), originally found as an antimicrobial peptide, is reported to possess strong anti-cancer activity[30]. In glioblastoma cells (U87MG and U251), TP4 promoted mitochondrial hyperpolarization and dysfunction, which elevated intracellular ROS, DNA damage, and necrotic cell death[103]. TP4 promoted necrosis in synovial sarcoma cells by inducing calcium overload, oxidative stress, and mitochondrial hyperpolarization[30]. Moreover, TP4 triggered non-small cell lung cancer cell death via necrosis[104] and inhibited the proliferation of triple-negative breast cancer cells via inducing an AP-1 protein FOSB, whose expression disrupts calcium homeostasis . These outcomes suggest that certain features enable peptides to demonstrate anti-cancer activity, which needs further investigation.
Pain relieving effects
-
There are also reports of studies examining the effects of protein hydrolysates and peptides derived from marine species on the nervous system[10,105]. Opioid-like peptides have been shown to positively affect motivation, emotion, stress, behavior, appetite, and pain management. These peptides bind with specific opioid receptors in the immune, endocrine, nervous, and digestive systems and show similar actions. However, their effects are weaker than those of the native opioid ligands. Therefore, these much safer natural products appear to be able to replace opioid drugs used for treating moderate to severe pain, coughing, and diarrhea[105]. Gabolysat PC60, fish protein hydrolysate sourced from cod and mackerel, is being used as a nutritional supplement with anxiolytic properties similar to diazepam on sympathoadrenal activity, activating the hypothalamic-pituitary-adrenocortical axis and increasing GABA content in hippocampus and hypothalamus during resting and stress conditions[105]. Moreover, marine proteins and peptides inhibit the progression of neurodegenerative diseases such as Parkinson's, Alzheimer's, and multiple sclerosis[10]. Neuroprotective peptide (HTP-1, Gly-Thr-Glu-Asp-Glu-Leu-Asp-Lys) isolated and purified from the Hippocampus trimaculatus prevented amyloid-β42 induced rat pheochromocytoma (PC12) cell death and up-regulated Bcl-2 expression indicating its desirable effects against Alzheimer's disease[106]. A tripeptide (Trp-Leu-Pro) isolated from Halocynthia roretzi alleviated 6-Hydroxydopamine (6-OHDA)-induced PC12 neuronal cell apoptosis, increased glutathione peroxidase activity and promoted cell viability indicating it can modulate neurodegenerative diseases associated with oxidative stress[107]. Kunitz-type peptides derived from venomous animals have demonstrated analgesic, anti-inflammatory, antihistamine, and many other biological activities. Sea anemone (H. crispa) derived Kunitz-type peptides, the lead peptide being InhVJ (57 amino acid residues), are reported to alleviate 6-hydroxydopamine-induced neurotoxicity in neuroblastoma Neuro-2a cells by reducing ROS production and increasing viability[31]. One of the main factors limiting the use of pharmaceuticals for treating central nervous system diseases is the permeability of the blood-brain barrier (BBB). The effectiveness of marine peptides in treating diseases of the central nervous system requires further testing on laboratory animals.
Anti-diabetic activity
-
Despite their regular metabolism, protein hydrolysates and peptides from marine organisms can be used to treat hyperglycemia. Glucose uptake-stimulating peptides can improve pancreatic β-cell homeostasis and glucose-stimulated insulin secretion, inhibit α-amylase and α-glucosidase activities and mediate mechanisms that increase insulin sensitivity in target cells[108]. Evidence is limited on using bioactive peptides from marine organisms to manage type 2 diabetes.
Isolation of anti-diabetes peptides from Spirulina platensis following ultrasound + subcritical water extraction (USW) was reported by Hu et al.[109]. The peptide with the sequence LRSELAAWSR has displayed strong α-glucosidase and DPP-IV inhibitory properties with IC50 values of 134.2 and 167.3 μg/mL and moderate α-amylase inhibition (IC50 = 313.6 μg/mL). Moreover, USW significantly increased the glycogen content, pyruvate kinase, and hexokinase activities in insulin-resistant HepG2 cells supporting potential anti-diabetic activity. Peptides purified from Protease XXIII and orientate hydrolysates of tuna cooking juice (PGVGGPLGPIGPCYE, CAYQWQRPVDRIR, and PACGGFWISGRPG) have indicated potential Dipeptidyl peptidase IV (DPP-IV) inhibitory activity with IC50 values of 116.1, 78.0 and 96.4 μM respectively[110]. DPP-IV inhibitors are potential targets in the treatment of diabetes as the inhibition helps to increase the postprandial insulin level. DPP-IV inhibitory peptides have also been isolated from skin gelatin hydrolysates of Tilapia and Halibut, among which IPGDPGPPGPPGP from Tilapia skin gelatin has indicated an IC50 of 65.4 μM[111]. Daily administration of tilapia skin gelatin hydrolysate to streptozotocin-induced diabetic rats has improved their glucose tolerance attributed to the plasma DPP-IV inhibition and increased glucagon-like peptide-1 and insulin secretion.
Zhu et al. have evaluated the modulatory effects of oligopeptides derived from marine salmon skin on type 2 diabetes mellitus-associated hyperglycemia and β-cell apoptosis in streptozotocin and high-fat diet-induced rats[112]. The oligopeptide treatment has significantly reduced oxidative stress and inflammatory responses in rats and attenuated hyperglycemic conditions. Clinical evaluation of collagen peptides of fish collagen hydrolysate in Chinese patients diagnosed with type 2 diabetes has revealed that daily supplementation of 13 g of the collagen peptides for three months has significantly lowered fasting blood glucose and insulin contents, total cholesterol, total triglycerides, low-density lipoprotein, free-fatty acids, and human glycated hemoglobin A1c, but have increased insulin sensitivity and HDL content[113]. Further, the treatment has substantially decreased nitric oxide and serum high-sensitivity C-reactive protein while increasing bradykinin, prostacyclin, and adiponectin, suggesting its desirable effects in improving glucose and lipid metabolism in diabetes patients.
Improvement of sexual and reproductive health
-
Reproductive and sexual health problems, including sexual dysfunction and infertility, are common concerns affecting millions worldwide. Literature and scattered evidence report that bioactive compounds from marine animals such as seahorses, oysters, sea cucumbers, and spoon worms could significantly improve hormonal imbalances, infertility, sexual dysfunctions, and impotence. According to traditional Chinese medicine, seahorse (Hippocampus kuda) consumption improves male fertility and reproductive dysfunctions[114]. A peptide extract from an autoclaved and ultrasonicated extract of Urechis unicinctus was reported to ameliorate erectile dysfunction in streptozotocin-induced diabetes in rats (oral administration) through elevating NO/cGMP signaling and upregulation of cavernosal nNOS and eNOS transcription and translation in penile tissues[115]. A commercial oyster peptide hydrolysate (source unknown) has been reported to significantly improve the functions of the hypothalamus-pituitary gonad axis in male Sprague Dawley rats during heavy load training[116]. The supplementation has lowered oxidative stress in the testis by upregulating the expression of the steroidogenic enzyme that promotes spermatogenesis and steroidogenesis. Another commercial oyster peptides sample (source unknown) was reported to effectively raise blood androgen levels in cyclophosphamide-induced male rats, improve the impaired pathological state and oxidative stress in kidneys, and enhance the state of the kidneys and testicles, all of which have a positive impact on procreative capacity[117]. Male reproductive infertility and dysfunctions are linked to diabetes. Seahorse (Hippocampus kuda) protein alcalase hydrolysate has improved male reproductive dysfunction in streptozotocin-induced diabetic rats fed with a high-fat diet[118]. The daily supplementation of 20 mg/kg of the hydrolysate has increased antioxidant enzyme activity, inhibited lipid peroxidation, restored morphology of seminiferous tubules in the testis, and increased the level of luteinizing hormone, follicle-stimulating hormone, testosterone, sperm count, and motility.
There is still very much a gray area due to the lack of in-depth understanding of the action mechanisms of these bioactives in humans, which needs to be proven by clinical studies.
-
The soy and dairy industries dominate the highly competitive market for bioactive peptide ingredients. Numerous drugs, cosmetics, functional foods, and nutritional supplements are already available on the market and contain bioactive peptides from terrestrial organisms, both in pure form and as unpurified blends[119]. To the authors’ knowledge, functional foods, dietary supplements, drugs, and cosmetics containing peptides from marine organisms with approved health claims are scarce or are not reported in the literature . There are numerous possibilities for manufacturing novel products that contain bioactive peptides from edible marine organisms.
There are two marine peptide-derived pharmaceutical products on the market: ziconotide, a native marine peptide, and brentuximab vedotin, a peptide derivative[120]. Ziconotide is a peptide with 25 amino acids first isolated from the marine mollusk Conus magus. It produces analgesic effects by binding to presynaptic N-type channel α1B subunit inhibiting the release of neuromodulators and neurotransmitters. Brentuximab vedotin is derived from dolastatin 10 (N,N-dimethyl Val-OH, L-valine, (3R,4S,5S)-dolaisoleucine, (2R,3R,4S)-dolaproine and (S)-dolaphenine) isolated from Dolabella Auricularia, a marine mollusk produced by the symbiotic cyanobacterium Caldora penicillate.
Clinical trials are currently underway in the United States and Europe for a number of other marine peptide-derived compounds. Plitidepsin is a depsipeptide under phase III clinical trials isolated from the tunicate Aplidium albicans that inhibits SARS-CoV-2 replication by targeting eukaryotic translation-elongation-factor-1A[120]. Glembatumumab Vedotin is another antibody-drug conjugate derived from dolastatin 10 that targets transmembrane glycoprotein NMB (GPNMB) in GPNMB-positive metastatic triple-negative breast cancer[121]. Dermochlorella is an oligopeptide-containing extract from Chlorella sp. that possesses skin protective effects and is actively researched as a cosmetic ingredient for skin firmer and toner[122]. As mentioned previously, Katsuobushi oligopeptides are undergoing clinical trials for their blood pressure-lowering activity[34,52]. Currently, venoms peptides from cone snails (conotoxins) are undergoing broad investigations for therapeutic applications while 6 of them (ω-CVID (AM336), Contulakin-G (CGX-1160), χ-MrIA (derivative) (Xen-2174), α-Vc1.1 (ACV-1), Conantokin-G (CGX-1007), κ-PVIIA (CGX-1051), and μO-MrVIB (CGX-1002)) have reached clinical trials[52]. Although numerous peptides have shown a wide variety of bioactivities, their effectiveness in treating human diseases requires further evaluations by preclinical trials.
-
According to the present state of knowledge, various obstacles need to be tackled in marine peptide-based functional food research. One challenging aspect of peptide research is purifying target peptides via conventional techniques such as membrane separation, dialysis, gel-, ion exchange-, and reverse-phase high-performance liquid chromatography (RP-HPLC) for laboratory-scale evaluations due to the trace amounts of active peptide/s recoverable from hydrolysates and fermented extracts[123]. On the other hand, affinity chromatography is a more effective technique, which relies on the formation of reversible intersections between an immobilized molecule and target ligands to be purified. Methods such as magnetic affinity separation using surface-modified agarose microspheres with immobilized affinity ligands yield a higher proportion of active peptides[37].
Microalgae have a high production and biomass conversion rate and produce proteins, essential amino acids, and other healthy nutrient contents with long-term sustainability compared to terrestrial crops[123]. However, extracting proteins from microalgae is challenging due to glycoside side chains on proteins and linkages between proteins and cell wall polysaccharides . To overcome this issue, alkaline solutions combined with reducing agents such as β-mercaptoethanol have been used to dissociate proteins from polysaccharides, resulting in an improved yield of alkaline soluble proteins. However, these processes have limitations in food applications[2]. The use of food-grade Reducing agents such as N-acetyl-L-cysteine (NAC) is advisable to ensure that food safety protocols are being followed.
Understanding the structure-activity relationship of peptides is paramount for developing treatment options and predicting their favorable and unfavorable side effects. Peptides exhibit bioactivities by directly associating/binding with the target or regulating cellular signaling pathways and/or the immune system[56,57]. Deep learning-based algorithms should be developed to analyze the structure-activity relationship of peptides. Comparison between studies is often challenging due to the numerous types of assays and assay conditions used in different laboratories to assess the bioactivities of peptides from various sources. Comparisons would be more meaningful if assay conditions were standardized and preferably identical positive controls were established between studies to check reproducibility or variations.
Molecular docking is a novel and advancing technique that generates knowledge of the structure-activity relationships between signaling molecules and peptides. Therefore, in silico approaches can have significant potential in the identification of action mechanisms, the discovery of novel bioactive peptides, as well as in the mining of complex protein hydrolysates for the identification of bioactive peptides[2].
The antimicrobial action mechanism of most reported peptides remains elusive due to the lack of further investigations. In an intriguing approach, Pan et al. used electron microscopic analysis to study the invasive mechanism of Epinecidin-1 on bacteria[124]. Epinecidin-1 was reported to attach to the bacteria's surface and decrease the negative surface charge. This peptide induces bacterial membranes to develop a saddle splay membrane curvature, followed by pore development, blebbing, budding, vascularization, and lysis. This approach would be an attractive way to visualize and study the antimicrobial action mechanism of peptides.
The stability of peptides remains another obstacle in developing functional foods. Peptides should withstand processing conditions with different pH and temperature treatments[3,6,14]. The stability could be investigated by comparing liquid chromatograms of peptides after subjecting them to incubation under a pH range of 2–10 and a temperature range of 4–100 °C for 2 h. Some bioactive peptides have failed to demonstrate bioactivity following oral administration in vivo, presumably due to gastrointestinal protease hydrolysis[36]. In vitro incubation with gastrointestinal enzymes has provided a straightforward process for predicting the fate of these peptides after oral administration[125].
Both from a commercial and efficacy standpoint it would be desirable if these marine organisms contained a higher concentration of these bioactives. Commercial species, such as sea cucumbers and seahorses, are being depleted due to overexploitation in an unsustainable manner and due to habitat loss. As part of aquaculture, it is important to study the factors that influence the synthesis of specific bioactive metabolites, breed, and optimize cultivation conditions to protect wild populations while simultaneously increasing the yield and production of these bioactive compounds. Effective and clean hydrolysis, extraction, and purification methods for marine bioactive compounds are crucial for developing and applying consumer products, particularly for quality control and efficacy[18].
Recent developments in understanding structure-activity relationships based on machine learning techniques, such as artificial neural networks-based quantitative structure-activity relationship models with chemical synthesis, could be integrated with marine peptide research. Based on the large pool of data on ACE inhibitory peptides, it will be possible to produce computer models that are capable of distinguishing the structure-activity relation of ACE inhibitory peptides[56]. Moreover, it is necessary to develop standards and conformity assessment criteria to compare the bioactivities.
Using high-throughput sequencing methods and bioinformatics tools on non-model organisms has opened up new avenues for bioactive peptide identification from genomes and transcriptomes[11]. With suitable modifications, this strategy appears particularly attractive for discovering bioactive gene-encoded products in marine organisms. Moreover, it helps to overcome the inherent difficulties of traditional wet-lab approaches.
Marine peptides have the potential to be applied in food, pharmaceutical, and cosmetic industries considering their advantages over the sources such as land animals. Additionally, marine peptides at therapeutic concentrations have not been associated with adverse effects, and they are abundantly available, free from religious restrictions, and simple to obtain[77]. As a sustainable approach for the biotechnological valorization of fish byproducts, marine collagen can replace conventional sources of collagen used in cosmetics[9]. Studies should be focused on evaluating their transcutaneous delivery, bioactive potency, stability and compatibility with cosmetic formulations, side effects, toxicity, and production costs[77].
Loss of bioavailability due to gastric digestion is a major issue for bioactive peptides when they are implemented in producing pharmaceuticals and dilatory supplements. Recent pharmaceutical research has expanded and diversified on aspects including targeted drug delivery and controlled drug release using micro-/nano-capsules[126]. They offer protection for the encapsulated bioactive agent against degradation, the ability to control the release rate, and ease of administration. Liposomes are among the most widely studied nanoparticles and can enclose lipophilic, amphiphilic, and hydrophilic drug molecules inside the phospholipid layers. Encapsulating peptides in liposomes is reported as an excellent approach to increasing their bioavailability[23].
-
In the medical field, interest is increasing from treatment-oriented to preventive medicine, but very few drugs can be called preventive drugs. As a means of reducing morbidity in adults from diseases such as cancer, arteriosclerosis, and diabetes, it is essential to actively introduce the concept of prevention by food rather than prevention by medicine. Recent developments in medical science have altered how people see their health, changing how natural foods and other natural items are sought. In spite of their low efficacy, natural bioactive substances are increasingly being used for the prevention or treatment of diseases. These include proteins and peptides that play an essential role in the body, modulating cell signaling pathways and performing various biological activities aside from providing nutritional functions.
Marine organisms are enriched with bifunctional secondary metabolites that evolved due to extreme environmental conditions. They have unique structures and properties compared to terrestrial organisms. For these reasons, research to develop new bioactive lead materials from marine organisms and use them in preserving human health has recently been in the spotlight. In the literature, a number of ACE inhibitory peptides have been discovered from marine protein hydrolysates, some of which may contribute to the reduction of hypertension. Further, peptides derived from marine organisms have been reported to possess antioxidant, antimicrobial, skin-protecting, apatite regulating, bone-protecting, cardioprotective, anticancer, pain relieving, anti-diabetic, and sexual and reproductive health benefits. Consequently, these peptides can be used to manufacture nutraceuticals, functional foods, dilatory supplements, pharmaceuticals, and cosmetics. More study is required to confirm the therapeutic effectiveness of these protein hydrolysates or peptides and to comprehend the structure-activity link of these substances in the body. Computer-based prediction models should be developed to understand the structure-activity relationship of peptides and thereby formulate peptide-based cosmetics and drugs. Appropriate measures should be taken to assess the quality and efficacy of functional foods and dietary supplements containing peptides derived from marine organisms and to standardize products through accredited and certified bodies.
This research was funded by a grant from the Natural Science and Engineering Research Council (NSERC) of Canada, grant number RGPIN201804680.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fernando IPS, Jayawardena TU, Wu J. 2023. Marine proteins and peptides: Production, biological activities, and potential applications. Food Innovation and Advances 2(2):69−84 doi: 10.48130/FIA-2023-0009 

Marine proteins and peptides: Production, biological activities, and potential applications
- Received: 10 September 2022
- Accepted: 21 February 2023
- Published online: 17 April 2023
Abstract: Marine protein hydrolysates and peptides have grown in popularity due to their biological activities and robust properties. They are increasingly studied in the functional food, pharmaceutical, and cosmeceutical sectors. This article discusses the current knowledge about preparing protein hydrolysates and peptides from seaweed, seafood, and seafood processing byproducts. Gaps in knowledge and technical expertise required for their industrial integration have been identified. The desire for natural substances to use as functional food has gained prevalence as consumers have become more aware of the adverse side effects of synthetic drugs. Aging-related chronic diseases, including cancer, arteriosclerosis, and diabetes, can be prevented by actively introducing food-based functional ingredients. Marine-derived proteins and peptides still face several hurdles to commercialization, such as scaling up production and maintaining a sustainable supply of raw materials. Further understanding of the physiological functionalities, action mechanisms, and clinical efficacy of these peptides and proteins would facilitate their use in biomedical applications and as functional ingredients in food and cosmetics.
-
Key words:
- Marine /
- Proteins /
- Bioactive peptides /
- Protein hydrolysates /
- Fish proteins /
- ACE-inhibitory peptides