









-
Protein N-glycosylation is considered to be the most important and ubiquitous post-translational modification in living organisms. The biosynthesis of protein N-glycans in plants is a highly conserved process that takes place in the endoplasmic reticulum and the Golgi apparatus[1,2]. Although sharing the same core carbohydrate unit (Man3GlcNAc2) with other eukaryotes, plant protein N-glycans have their characteristic features, such as the occurrence of β1,2-xylosylation attached to core mannose residue and α1,3-fucosylation to innermost core GlcNAc residue (Fig. 1). Furthermore, plant glycoproteins lack terminal modifications by neuraminic acid residues or modifications with β1,4-linked galactose[3].
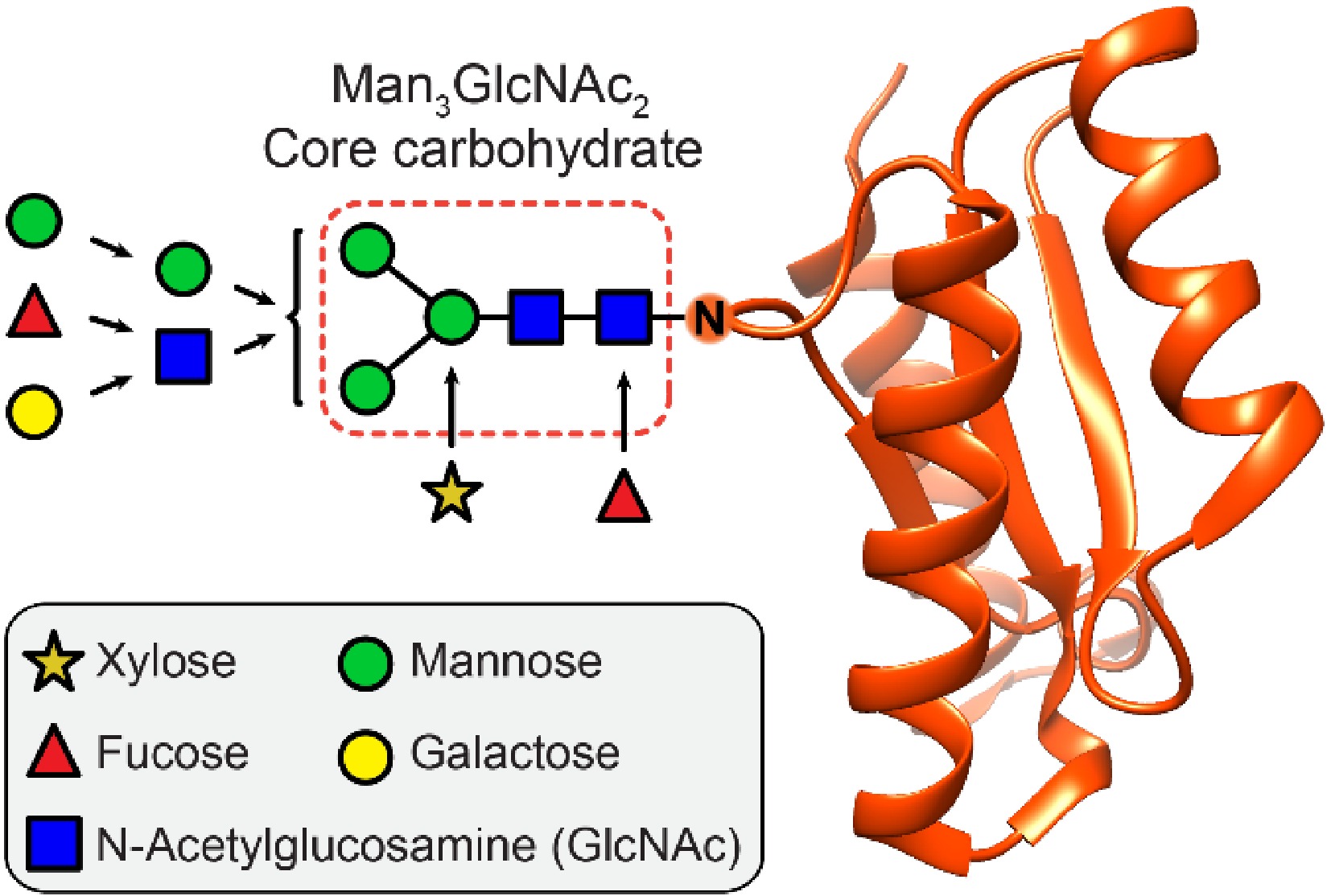
Figure 1. Structural composition of plant protein N-glycosylation. The Man3GlcNAc2 core carbohydrate is attached via an asparagine side chain (N) to the protein part (red). The core carbohydrate region can be decorated extensively by a variety of sugar moieties.
Reported data indicate that the plant N-glycosylation pattern is associated with the regulation of plant growth and development[4]. For example, a mutation of the gcs1 gene (encoding the respective α-glucosidase 1, involved in the trimming of N-glycans) in Arabidopsis thaliana, induced abnormal embryonal development and reduced the accumulation of storage proteins[5]. Inactivation of α-glucosidase 1 in rice led to defects in root cell division and elongation[6]. Another type of exoglycosidases required for N-glycan processing are α-mannosidases (α-Man), which were also described to be essential for cell wall biosynthesis and the regulation of root development. The mutation of α-mannosidase genes or the inhibition of α-mannosidases by the glycosylation pathway inhibitor kifunensine resulted in shortened roots and altered cell walls[7]. The N-glycan processing glycosidases α-mannosidase and β-N-acetylhexosaminidase (β-Hex) were also reported to participate in the regulation of fruit softening and ripening[8−10]. The silencing of β-Hex or α-Man encoding genes, or the inhibition of their enzymatic activities by the iminosugar 1-deoxynojirimycin, were found to reduce the rate of softening in tomatoes[11,12]. However, the underlying physiological role of plant N-glycans during the ripening process still needs further validation and elucidation.
The melting peach is, in contrast to other non-melting peach cultivars, prone to an extremely rapid softening and therefore exhibits a short period from harvest to consumption of maximum 5−7 days (Fig. 2a)[13]. This short shelf-life significantly restricts its storage, transportation, and sale. A better understanding of how to delay the fruit softening would be of great benefit to an increased market value of melting peaches. In previous studies, Han et al. discovered that the enzymatic activity of β-Hex was associated with the softening and ripening of peach fruits, and treatment with the β-Hex inhibitor 2-Acetamido-1,2-dideoxynojirimycin (2-ADN) delayed the decrease of firmness and resulted in reduced ethylene production[14]. It suggested that the N-glycosylation in peaches is also involved in the regulation of fruit development and ripening[14]. However, these changes in the N-glycan pattern during fruit development and ripening of peach fruit were not reported in detail. Therefore, a thorough study of N-glycosylation at different stages is highly desirable. Here, we chose to analyze the fruit flesh of melting peaches (Prunus persica L. Batsch cvs Xia Hui 6), to study the changes of N-glycosylation during fruit development and ripening.
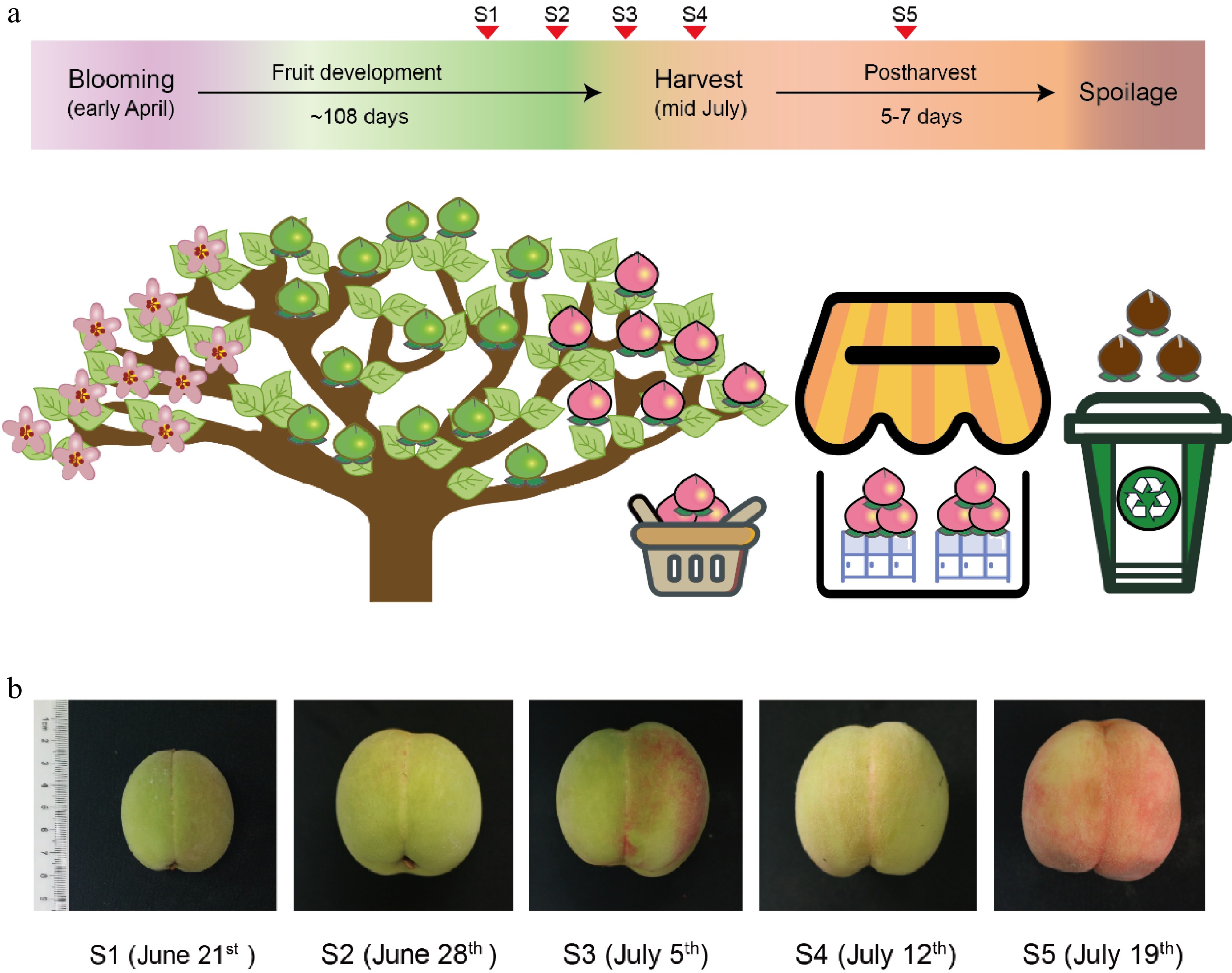
Figure 2. Peach fruit development and ripening. (a) Stages of fruit development before and after peach harvest. The red triangle markers show the time points of sample collection stages S1−S5. (b) Size and color of peach fruits at stages S1−S5 with the collection date shown in parenthesis.
-
Melting peaches (Prunus persica L. Batsch cvs Xia Hui 6) obtained from the Jiangsu Academy of Agricultural Sciences (Nanjing, China) were used in the study. Fruits without visible defects were randomly collected from one designated peach tree (approx. 8 years old) at the following developmental stages: 80 d after full bloom (S1), 87 d after full bloom (S2), 94 d after full bloom (S3), and 101 d after bloom (= harvest, S4); one additional sample cohort was taken 7 d after harvest (S5). Five replicate samples (from individual peaches) for each stage were collected, immediately frozen in liquid nitrogen, and stored at −80 °C for further analysis.
Extraction and determination of exoglycosidase activity
-
One gram of frozen flesh of peach fruits was ground to a fine powder in liquid nitrogen, transferred to a 2 mL microtube, and mixed with 1 mL of extraction buffer (50 mM sodium acetate buffer, pH 5.5 containing 0.5% polyvinyl pyrrolidone). The water-soluble protein was extracted overnight at 4 °C. The extract was centrifuged at 12000 g for 5 min and the supernatant was collected. Colorimetric exoglycosidase assays were performed based on the method described by Wang et al.[15]. Briefly, 25 μL reaction mixtures consisting of 2 μL of the respective p-nitrophenyl-sugar substrate (pNP-β-D-N-acetylglucosaminopyranoside, pNP-α-D-mannopyranoside, and pNP-β-D-galactopyranoside, 20 mM each), 2.5 μL of sodium acetate (1 M, pH 3.5), 5.5 μL of distilled water, and 15 μL of enzyme extract were used to test the enzymatic activity. The reaction was incubated at 37 °C for 30 min and stopped by the addition of 25 μL sodium carbonate solution (1 M). The color intensity of released p-nitrophenol was measured at 405 nm using a microplate reader.
Preparation N-glycans from peach fruit
-
The preparation of N-glycans was based on the method described by Du et al.[16]. One gram of peach fruit was homogenized in ice-cold water. The homogenate was mixed thoroughly with an equal volume of 40% (w/v) trichloroacetic acid (TCA), then centrifuged at 12000 g for 30 min at 4 °C. The debris was collected and carefully washed with deionized water five times to remove free sugars. The precipitate was then suspended in water and heated at 95 °C for 15 min. 0.5 milliunits of recombinant peptide N-glycosidase Dj (PNGase Dj)[17] was used to release N-glycans in 50 mM acetic buffer (pH 4.0) by incubating the mixtures for 16 h at 37 °C. The supernatant was collected by centrifugation at 12000 g for 20 min and transferred to a Supelclean ENVI Carb solid-phase extraction tube (Supelco, 500 μL bed volume), which initially was flushed with 3 mL of 80% acetonitrile containing 0.1% trifluoroacetic acid (TFA, v/v), and then equilibrated with 3 mL of distilled water. The liberated N-glycans were eluted with 1.5 mL of 40% acetonitrile containing 0.1% (v/v) TFA. The collected fraction was dried with centrifugal concentration.
N-glycan profiling by ultra-performance liquid chromatography
-
Dried N-glycan samples were labeled with 10 μL of 2-aminobenzamide (2-AB) labeling solution (35 mM 2-AB and 0.1 M sodium cyanoborohydride in dimethyl sulfoxide/acetic acid 7:3 v/v) at 65 °C for 4 h as previously described by Zhang et al[18]. The reaction product was mixed with 35 μL acetonitrile and 40 μL of the mixture was removed for UPLC analysis. A 35 μL sample was injected into a Shimadzu Nexera UPLC system equipped with an RF-20Axs fluorescence detector and separated by an Acquity ethylene-bridged hybrid (BEH) glycan column (Waters, 1.7 μm pore size, 2.1 × 150 mm). Solvent A was aqueous 50 mM ammonium formate buffer, pH 4.5, and solvent B was acetonitrile. The flow rate was set at 0.5 mL/min. The gradient of solvent A was increased linearly from 5%−12% for the first 6 min and increased to 44% over the next 39 min. Fractions collected from the UPLC were then dried by vacuum concentration for further mass spectrometric analysis. 2-AB labeled dextran was used to calculate the relative retention times of N-glycans based on their glucose unit (GU) values by comparison with previously assigned standard N-glycans[19].
MALDI-TOF MS analysis of N-glycans
-
Collected and vacuum-dried fractions from the UPLC runs were dissolved in 2 μL of distilled water and analyzed by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry using a Bruker Autoflex Speed instrument (equipped with a 1000 Hz Smarbeam-II laser). 6-Aza-2-thiothymine was used as a matrix. The obtained mass spectra were analyzed by Bruker Flexanalysis software version 3.3.80, and manually annotated with the glycan structural software GlycoWorkbench (version 1.1)[20].
Statistical analysis
-
The data are presented as mean values accompanied by standard deviation (SD) values derived from sample triplicates. The differences among the five sample collection time points were analyzed by one-way analysis of variance (ANOVA) followed by Tukey's posthoc test using the PASW Advanced Statistics 18 software package. P-values less than 0.05 were defined as significant. The Pearson's correlation coefficient was used to quantify the relationship between the N-glycan species and exoglycosidase activities.
-
To understand the regulatory function of N-glycosylation on peach fruit development and ripening in more detail, it is necessary to analyze its dynamic compositional changes. Here, we collected samples at different periods from the early stage of fruit development to the late stage of fruit ripening, including three stages of pre-harvest (S1, S2, and S3), one harvest stage (S4), and one postharvest stage (S5) (Fig. 2b). N-glycans were enzymatically released from each sample by PNGase Dj digestion, fluorescence-labeled with 2-AB, and subsequently detected using HILIC-UPLC.
The UPLC chromatogram showed that the 2AB-labeled N-glycans of the peach fruit sample S1 were eluted between 15 to 40 min and consisted of 15 individual sample peaks (Fig. 3a). The glucose unit (GU) values of each sample peak are listed in Table 1. Each peak was then collected and analyzed with MALDI-TOF-MS for assigning each corresponding N-glycan structure (Fig 3b & Table 1). Nine of these structures (M3, MMX, GnMX, MMXF, GnMXF, GnGnMXF, GGnGnXF, GGXF, and (GF)(GF)XF) were complex-type N-glycans, five N-glycans (M5, M6, M7, M8, and M9) belong to the high-mannose type, and one N-glycan fraction (peak 6) belongs to the class of hybrid-type N-glycans (M4X).
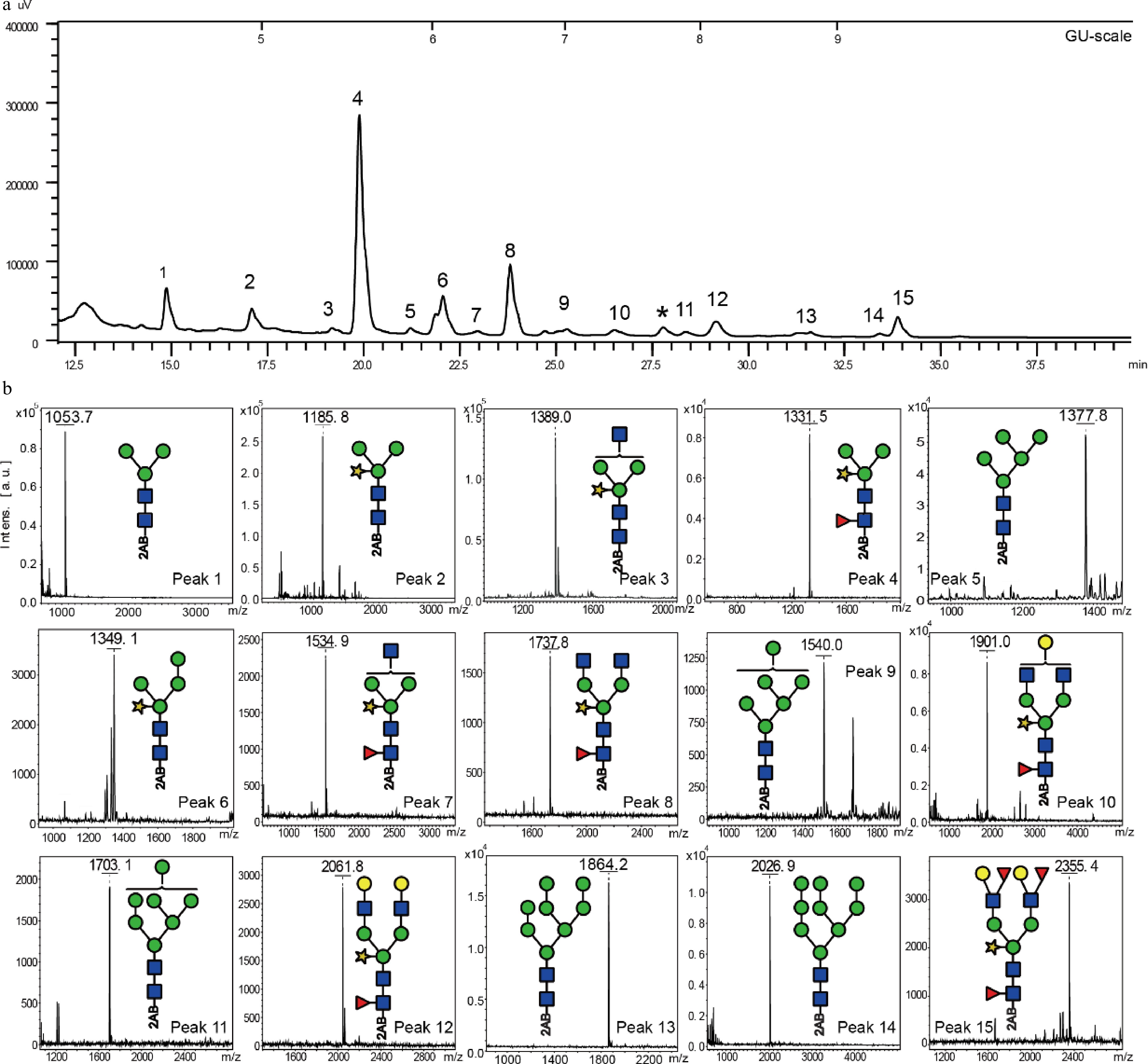
Figure 3. Analysis of peach N-glycans at stage S1. (a) Representative UPLC chromatogram of peach fruit N-glycans from stage S1. (b) MALDI-ToF mass spectra of the individual fractions 1−15. The peak fraction, * was determined as a non-N-glycan oligosaccharide corresponding by mass to Hex9 (m/z = 1618.06 [M+Na]+).
Table 1. Overview of N-glycan structures determined in peach fruit sample S1.
Peak number Retention time GU value Structure The theoretical electron charge-mass ratio (m/z) Detection value of charge-mass ratio (m/z) Name 1 14.90 4.33 1053.4 1053.7 M3 2 17.15 4.90 1185.4 1185.8 MMX 3 19.25 5.43 1388.5 1389.0 GnMX 4 19.97 5.62 1331.5 1331.5 MMXF 5 21.35 5.94 1377.5 1377.8 M5 6 22.18 6.18 1347.5 1349.1 M4X 7 23.11 6.39 1534.6 1534.9 GnMXF 8 23.98 6.63 1737.6 1737.8 GnGnMXF 9 25.48 7.00 1539.5 1540.0 M6 10 26.78 7.38 1899.7 1901.0 GGnGnXF 11 28.66 7.87 1701.9 1703.1 M7 12 29.45 8.15 2061.8 2061.8 GGXF 13 31.69 8.76 1863.7 1864.2 M8 14 33.79 9.47 2025.7 2026.9 M9 15 33.92 9.71 2353.9 2355.4 (GF)(GF)XF Note: Changes of the peach N-glycome during development and ripening
-
The N-glycan profiles of the peach fruit samples from stages S2 to S5 were also analyzed by UPLC. The N-glycan structures previously determined for the S1 sample were also detected in the samples from stages S2 to S5, although the relative contents of the individual N-glycan structures varied significantly (Supplemental Fig. S1). The quantitative changes of N-glycans 1−15 were calculated based on comparing the corresponding peak areas to the sum of peak areas 1−15 (Table 2). Not unusual for N-glycans from plant materials, the MMXF glycan (4) was the main component (varied from 43.6 ± 1.0% to 55.6 ± 5.1%) throughout stages S1-S5. GnGnMXF (8) was the second most abundant N-glycan with the relative contents varying between 9.1 ± 0.6% and 14.7 ± 0.4%. The relative contents of the rest of the N-glycan species did not exceed 10%. A significant increase in MMXF was observed during fruit ripening, especially at stage S5. A diametric change in abundance was observed for the GnGnMXF glycans, whose contents significantly declined during peach maturation. A decreasing trend in abundance was also observed for another terminal GlcNAc modified N-glycan, GnMX (3) (Supplemental Fig. S2). No significant changes in relative contents were found for MMX (2), M5 (5), GnMXF (7), and GGnGnXF (10). For other N-glycans the relative content changes were less than 5%. In a previous report, Priem and Gross found that vacuum infiltration of tomato fruits with 5 μg/mL Man3XylFucGlcNAc solution (a truncated MMXF (3) substructure) triggered fruit ripening, which indicated that the trimming of terminal GlcNAc residues in N-glycans may have a regulatory function[21]. The herein shown changes of the MMXF, GnGnMXF, and GnMX structures may be therefore also key to N-glycans for the regulation of fruit ripening.
Table 2. The relative contents of different N-glycans from peach fruit at stages S1−S5.
Peak number N-glycans Relative content (%) S1 S2 S3 S4 S5 1 M3 7.1 ± 1.0b 10.3 ± 1.8a 6.1 ± 0.8b 5.9 ± 0.7b 4.8 ± 2.2b 2 MMX 6.8 ± 2.1a 7.1 ± 1.1a 6.0 ± 1.0a 7.7 ± 0.6a 6.4 ± 1.8a 3 GnMX 1.3 ± 0.1ab 1.5 ± 0.3a 1.0 ± 0.2bc 1.0 ± 0.1bc 0.7 ± 0.2c 4 MMXF 46.3 ± 4.4b 45.2 ± 1.4b 43.6 ± 1.0b 48.4 ± 1.6b 55.6 ± 5.1a 5 M5 1.1 ± 0.1a 1.3 ± 0.4a 1.5 ± 0.9a 1.5 ± 0.3a 0.9 ± 0.3a 6 M4X 10.1 ± 2.0a 8.5 ± 1.0ab 7.8 ± 0.1b 9.6 ± 0.3ab 9.9 ± 0.6a 7 GnMXF 0.9 ± 0.2a 1.5 ± 0.9a 1.2 ± 0.2a 0.6 ± 0.1a 0.8 ± 0.2a 8 GnGnMXF 14.2 ± 1.5a 14.7 ± 0.4a 14.6 ± 0.6a 10.9 ± 1.7b 9.1 ± 0.6b 9 M6 1.7 ± 0.2b 3.0 ± 0.4a 2.2 ± 0.4ab 3.2 ± 1.1a 2.8 ± 0.3a 10 GGnGnXF 2.1 ± 0.3a 1.7 ± 0.5a 1.7 ± 0.1a 1.6 ± 0.1a 1.6 ± 0.2a 11 GGXF 3.6 ± 1.4a 0.7 ± 0.2b 1.3 ± 0.2b 1.5 ± 0.8b 1.6 ± 0.3b 12 M7 1.0 ± 0.5c 2.5 ± 0.5b 4.9 ± 0.5a 3.3 ± 0.6b 2.6 ± 0.2b 13 M8 0.6 ± 0.3b 0.3 ± 0.1b 1.1 ± 0.1a 1.0 ± 0.2a 0.5 ± 0.1b 14 M9 0.7 ± 0.4ab 0.4 ± 0.1b 1.1 ± 0.3a 0.8 ± 0.5ab 1.0 ± 0.2ab 15 GFGFXF 2.6 ± 1.5b 1.5 ± 0.3b 6.0 ± 1.4a 2.9 ± 0.5b 2.3 ± 0.2b The relative content of each N-glycan is expressed as mean ± SD (n = 3). Different letters indicate significant differences (p < 0.05) in the relative content of N-glycans among different stages. Changes of exoglycosidase activities
-
Based on the metabolic pathway known for N-glycans, a series of exoglycosidases were described to participate in the truncation and modification of these carbohydrates[22]. Among them, β-Hex was found to be associated with fruit development and ripening of peaches, but the involvement of other N-glycan processing exoglycosidases was not yet reported[14]. We investigated the changes in the activities of three main types of exoglycosidases (α-Man, β-Hex, and β-galactosidase (β-Gal)) suspected to be involved in the processing of N-glycans at different maturity stages (Fig. 4a−c). As shown in Fig. 4a−c, the activities of α-Man, β-Hex, and β-Gal all showed increasing trends, with the lowest enzymatic activities measured in immature fruits (S1) and highest activities measured in ripe fruits (S5). The activity of β-Hex was the highest among the three enzymes, which increased slightly during stages S1 to S3 and climbed rapidly in stages S4 and S5. β-Hexosaminidases are responsible for the removal of terminal GlcNAc residues from N-glycans and can catalyze the conversion of GnGnMXF to MMXF[8,14]. Therefore, the increase of β-Hex activity during the fruit development and ripening may also be the cause for the increase of MMXF structures and the decrease of GnGnMXF structures. A similar increase of activity over time was observed for α-Man activity, which releases terminal mannose from N-glycans. However, no decrease of the relative content of N-glycans M8 (13) and M9 (14) was observed in our study, indicating no degradation of the larger size high-mannose type N-glycans by α-Man. We also found an increase of β-Gal activities between immature stages and mature stages of peach fruits. β-Gal activities were described to be important for the hydrolysis of xyloglucan, rhamnogalacturonan, and hemicelluloses in plant cell walls, and were previously associated with fruit ripening[23]. Liu et al. reported that the silencing of the BGAL genes in peach fruits responsible for the β-Gal activity resulted in the inhibition of cell wall degradation and delayed ripening[24]. N-glycans with terminal galactose moieties could also be possible substrates of β-Gal enzymes. However, in this study no removal of terminal galactose from N-glycans was observed, suggesting that the measured β-Gal activities in the peach fruits target only specific cell wall polysaccharides instead of glycoproteins.
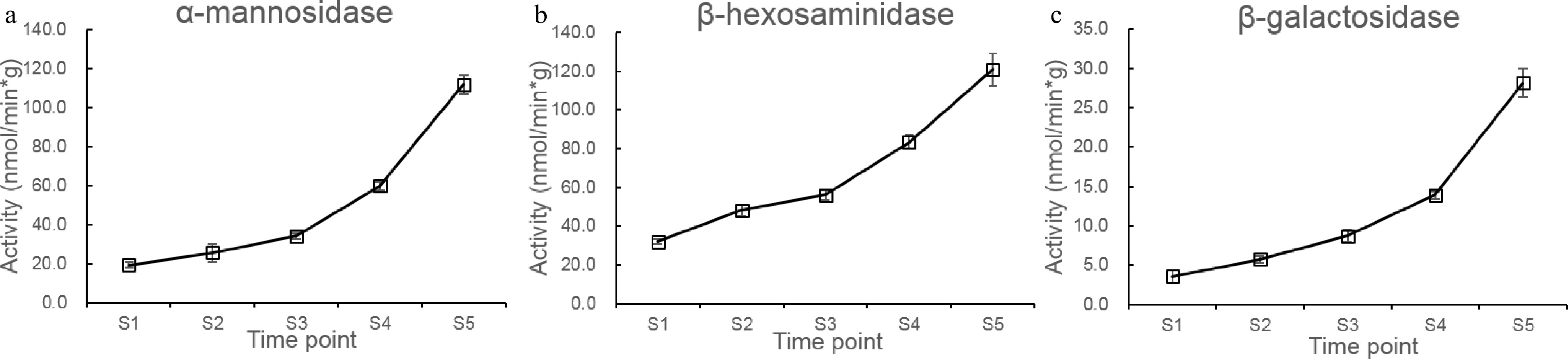
Figure 4. Changes in the exoglycosidase activities at stages S1 to S5. (a−c) changes of β-Hex, α-Man, and β-Gal activities at stages S1 to S5.
Relationship between changes in N-glycan pattern and exoglycosidase activities
-
To determine the association between N-glycan and the various tested exoglycosidases, the relative contents of N-glycans and enzyme activities at each stage were subjected to correlation analysis. The results showed that the relative contents of the MMXF glycan (4) were negatively correlated with β-Hex activities (p < 0.05), while the relative contents of the GnGnMXF glycan (8) were positively correlated with β-Hex activities (p < 0.05) (Table 3). These results indicate that β-Hex activity is associated with the truncation of N-glycans containing terminal GlcNAc residues. This relationship between N-glycans and β-Hex activity was previously shown by Meli et al. using RNA interference to inhibit β-Hex activity in tomato plants, which resulted in increased amounts of N-glycans with terminal GlcNAc moieties[12]. In the same work, they also found that the amount of terminal mannosyl N-glycan increased when RNA interference was used to inhibit α-Man activity. This finding is in contrast to our study, as we did not observe any decrease in the relative contents of high-mannose type N-glycans, despite the measured increase of α-Man activity. The correlations between high-mannose type N-glycans and α-Man were not statistically significant. Also, no statistically significant correlation was found for the other combinations of N-glycan structures and exoglycosidases. The results indicated that only the changes in abundance of the MMXF and GnGnXF N-glycan structures (4 and 8) are associated with the development and ripening of peach fruits. Two possible hypotheses were suggested for the role of N-glycans in fruit ripening. The first hypothesis suggests that specific N-glycans serve as intrinsic hormones to trigger fruit ripening via signaling cascades[25,26]. The second hypothesis postulates that changes of N-glycans which are still conjugated to the host protein can directly affect the diffusion and solubility of glycoprotein in the cell wall to reduce fruit firmness (Fig. 5)[3]. Fig. 5 also shows the dynamic relationship and dependence among the various N-glycan species, which in many cases can be directly linked to increased glycosidase activities. The data on the changes of the conjugated N-glycan structures obtained in this study, during the development and ripening process of peach fruits, provide further evidence for the latter hypothesis.
Table 3. The Pearson's correlation analysis of N-glycan relative contents and exoglycosidase activities.
Peak number N-glycans R value β-Hex α-Man β-Gal 1 M3 NC −0.68ns NC 2 MMX −0.03ns −0.09ns NC 3 GnMX −0.86ns NC NC 4 MMXF 0.89* NC NC 5 M5 NC −0.42ns NC 6 M4X NC 0.41ns NC 7 GnMXF −0.57ns NC NC 8 GnGnMXF −0.94* NC NC 9 M6 NC 0.48ns NC 10 GGnGnXF NC NC −0.64ns 11 GGXF NC NC −0.26ns 12 M7 NC 0.11ns NC 13 M8 NC −0.06ns NC 14 M9 NC 0.52ns NC 15 GFGFXF NC NC −0.13ns NC: not calculated; ns: no significant difference. 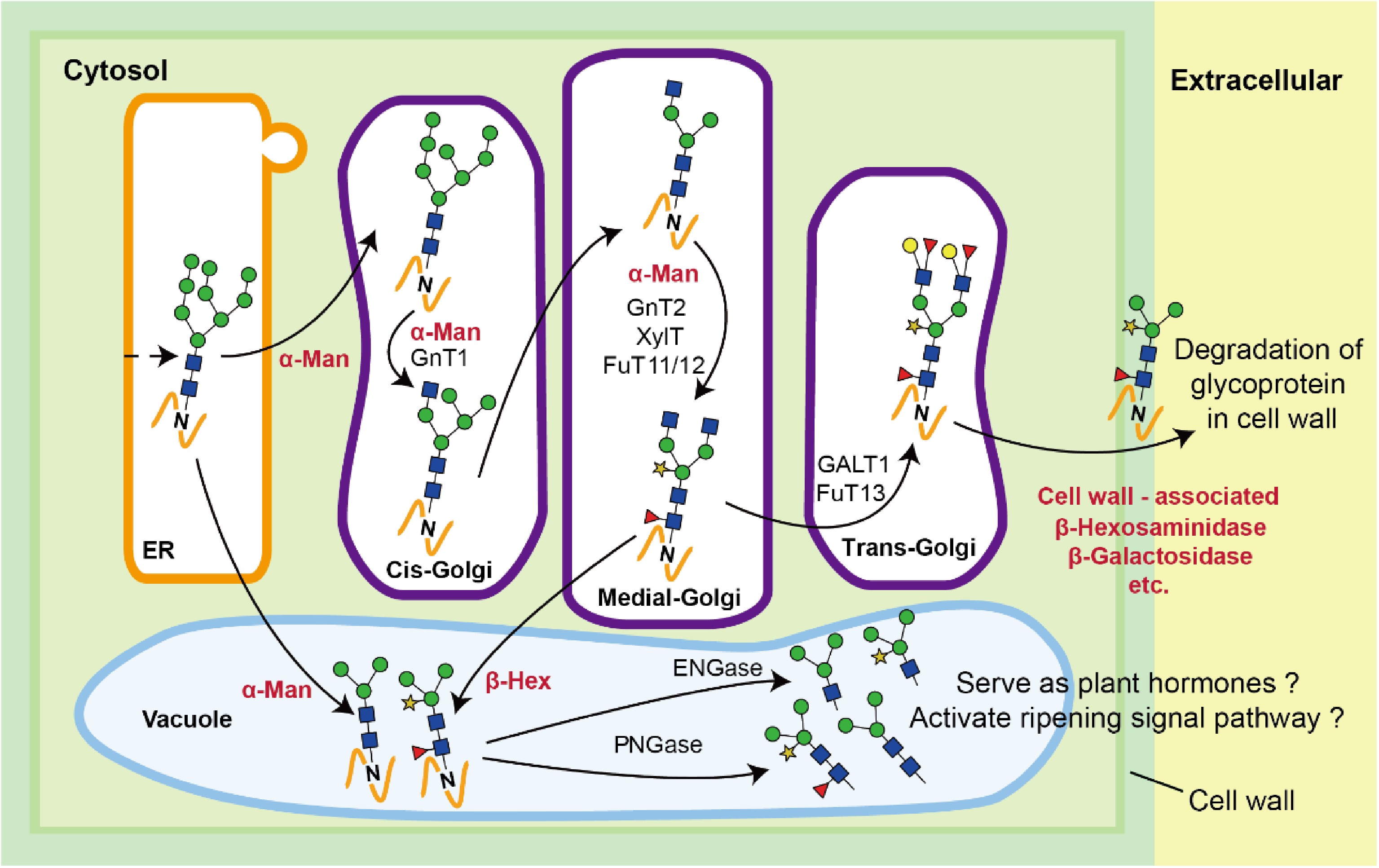
Figure 5. Schematic diagram of the regulatory mechanism of N-glycosylation in fruit ripening. PNGase, peptide N-glycosidase; ENGase, endo-β-N-acetylglucosaminidase; FuT11/12/13, α1,3/4-fucosyltransferase isoforms 11/12/13; XylT, xylosyltransferase; GalT1, galactosyltransferase isoform 1; GnT1/2, β1,2-N-acetylglucosaminyltransferase isoforms 1/2.
-
In conclusion, this study identified the N-glycosylation fingerprints of melting peach fruits at different developmental and ripening stages and summarized its changes. It was found that the truncation of terminal GlcNAc residues, which is caused by the increase of β-Hex activity, correlates well with the ripening process of peach fruit. This study lays the foundation for further elucidating the physiological function of N-glycosylation in fruit development and ripening. The inhibition of β-Hex by genetic or biochemical means could be a valuable strategy to delay the softening and ripening in peach fruits.
- This work was supported by the Priority Academic Program Development of Jiangsu Higher Education Institutions (grant KYZ201824 to Ting Wang), and the National Natural Science Foundation of China (NSFC grants 31471703, 31671854, 31871793 and 31871754 to Josef Voglmeir and Li Liu).
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 HPLC analysis of N-glycans from peach fruits at different stages.
- Supplemental Fig. S2 Changes in the relative contents of N-glycans at stages S1 to S5.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang T, Jia X, Liu L, Voglmeir J. 2021. Changes in protein N-glycosylation during the fruit development and ripening in melting-type peach. Food Materials Research 1: 2 doi: 10.48130/FMR-2021-0002

Changes in protein N-glycosylation during the fruit development and ripening in melting-type peach
- Received: 08 October 2021
- Accepted: 27 October 2021
- Published online: 09 November 2021
Abstract: The posttranslational modification of proteins with complex carbohydrate moieties (glycosylation) regulates the process of fruit ripening. Exoglycosidases are enzymes that can trim this protein glycosylation and are therefore considered to be important targets in the control of fruit ripening and softening. Melting-type peaches are popular seasonal fruits in many Asian regions, but the extremely short shelf-life of the peach fruits significantly hampers their economic value. To investigate the effect of the protein glycosylation and exoglycosidase activities on the development and ripening of the peach fruit, the fruit flesh of the melting peach cultivar 'Xia hui 6' at five different maturity stages were analyzed. The N-glycan profile of each sample was characterized and quantified by HILIC-UPLC and MALDI-TOF mass spectrometry, revealing two characteristic N-glycan structures (MMXF and GnGnMXF) which were strongly affected by the state of maturity. Furthermore, it was shown that one of the endogenous exoglycosidase activities analyzed (β-N-acetylhexosaminidase, β-Hex) correlated with the MMXF and GnGnMXF N-glycan structures (p < 0.05) in an obverse manner. These findings lay the foundation for further elucidation of the physiological functions of protein glycosylation in peach fruit development and ripening.
-
Key words:
- N-glycosylation /
- Melting peach /
- Fruit development /
- Fruit ripening /
- Prunus persica