








-
Meat is one of the most important foods globally as it provides proteins, fatty acids and many other nutrients. Due to the rising world population and individual incomes, the demand for meat is increasing year on year, especially in developing countries. The Food and Agriculture Organization of the United Nations (FAO) pointed out that meat production currently relies heavily on livestock, which poses severe implications for the environment, human health and animal welfare[1]. Alternative protein is a promising technology to solve the dilemma of meat consumption and animal husbandry. Among alternative protein technologies, cultured meat has the potential to produce 'real' meat with less implications than livestock farming[2]. As shown in Fig. 1, another advantage of cultured meat is that the production cycle is shorter than that of traditional animal husbandry.
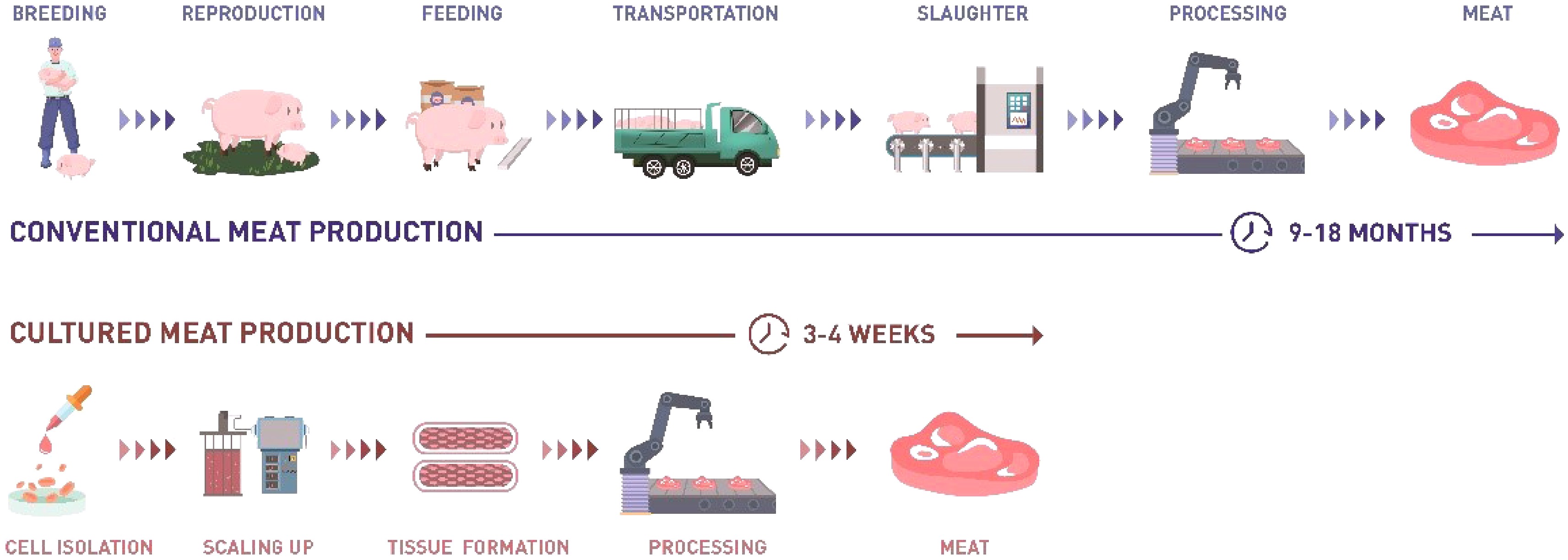
Figure 1. Comparison between traditional meat production and cultured meat production.
Cultured meat uses cultured animal cells to produce meat or meat proteins and fat tissue for human consumption according to the development and regeneration principles of mammalian tissues. The idea was first proposed by British Prime Minister Winston Churchill in 1931. In August 2013, Mark J. Post revealed the world's first cultured beef burger in London, which proved the feasibility of the concept of cultured meat for the first time[3]. Since 2013, societal attention and research on cultured meat have developed rapidly, to date more than 70 startup and scale-up companies are working in this space worldwide. In December 2020, the Singapore Food Agency approved Eat Just's cultured chicken to be sold in Singapore, which is the first regulatory approval for the production and sale of cultured meat globally.
While there has been a lot of research on technology, regulation and consumer acceptance of cultured meat, more progress is needed before cost-effective cultured meat can be produced commercially. Technically, cultured meat needs to optimize seed cells, cell media, scaffolds, and the biomanufacturing processes. At the regulatory level, although Singapore has approved cultured meat products, this approval only applies to a single product from a single species. Currently, it appears that every individual product has to be approved for each geography. How to form a relatively unified regulatory system needs further consideration.
Finally, how to get consumers to accept and be satisfied with cultured meat is a necessary part of commercial success. In this paper, we review the methodologies of cultured meat and discuss the technologies to achieve low-cost cultured meat industrial production. We also discuss the regulation and consumer acceptance and how both accelerate commercialization.
-
Cultured meat is muscle tissue and fat tissue generated in vitro by culturing and biomanufacturing animal cells and tissues for human consumption[2]. Depending on cell types and complexity, cultured meat products fall into three main categories, including: 1) cells as an ingredient, 2) muscle and fat organoids, and 3) full thickness complex tissue (Fig. 2). The ultimate goal of cultured meat is to create full thickness complex tissue containing muscle fibers, fat tissue, blood vessels, nerves and extracellular matrix, which is currently difficult. The medium-term goal is to produce cultured meat products that contain large amounts of muscle fibers, that are the basic building blocks of muscle and contain the most important source of protein in meat, accounting for about 80%−90% of the total protein. In addition, it is highly conducive to meat-like organoleptic features to interlace the muscle fibers with fat tissue. The short-term goal of cultured meat is to produce meat-like products, containing cells and an edible scaffold. Either way, cultured meat requires functional seed cell, low-cost serum-free medium, tissue engineering and scaling up processes. The continuous research in these fields can reduce the production cost of cultured meat for large-scale production purposes.
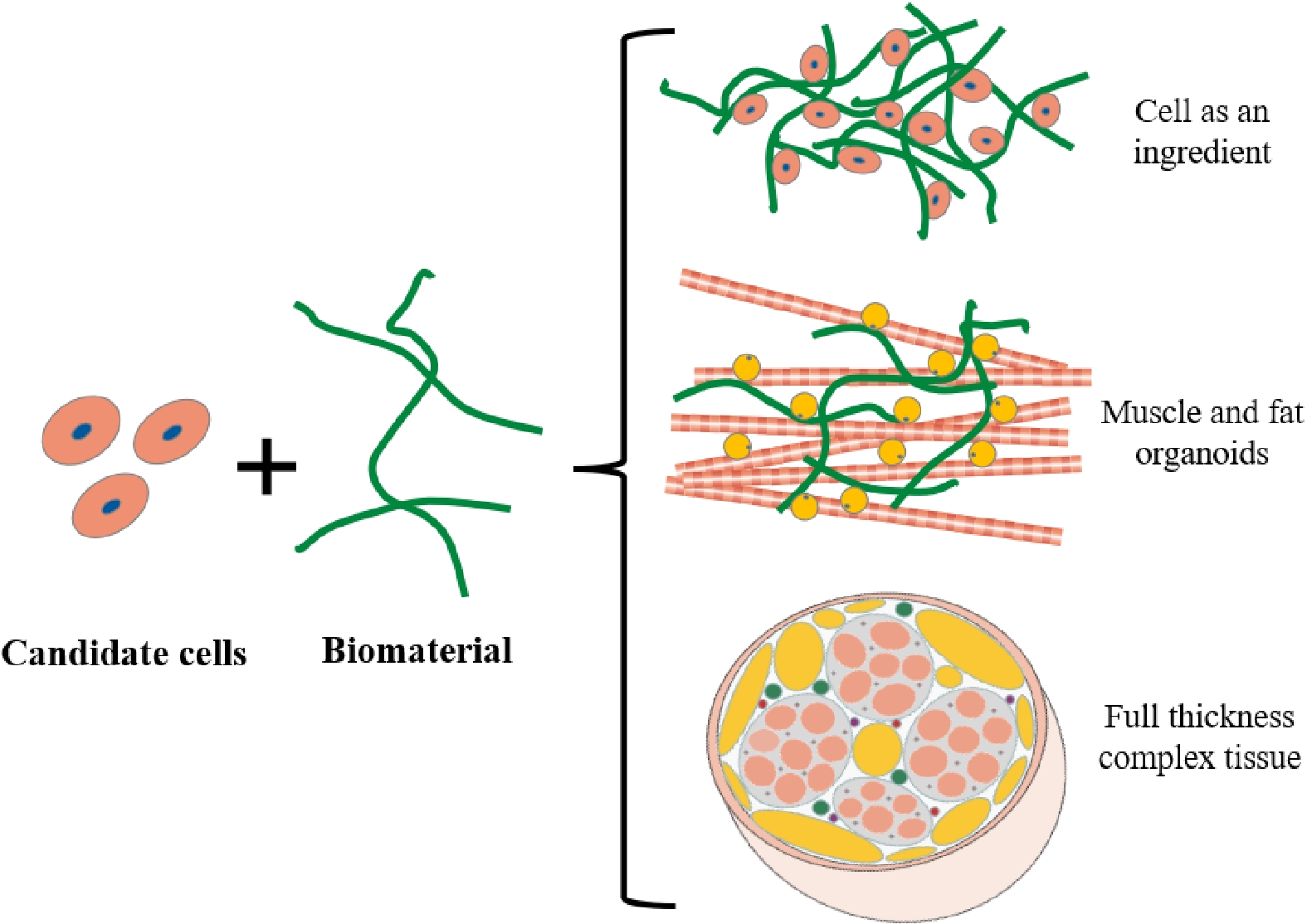
Figure 2. Three categories of cultured meat. 1. Direct combination of cells and biomaterials; 2. muscle and fat organoids generated by different cell types, that are combined by biomaterials; 3. complex muscle tissue, including muscle, fat, blood vessels, fibrous tissue, etc.
Seed cells
-
Meat proteins or fat should be the minimum necessary ingredients for cultured meat according to the three goals. Embryonic stem cells (ESC), induced pluripotent stem cells (iPSC) and mesenchymal stem cells (MSC) are pluripotent or multipotent stem cells with the ability to differentiate into muscle cells, stromal cells and fat cells, but they require precise regulation of cell fate. At present, ESC and iPSC are rarely used for cultured meat production and research whereas some startups use MSC or adipose tissue derived stem cells for fat-related products. Muscle stem cells (MuSC) are unipotent adult stem cells that contribute to muscle regeneration and form muscle fibers in vitro and therefore appear to be the most suitable cell source for skeletal muscle fiber based cultured meat. In addition to the cells mentioned above, other muscle tissue can also be consumed as meat, including smooth muscle and heart muscle. Smooth muscle tissue can be produced from smooth muscle cells, and cardiac muscle tissue can be produced from cardiomyocytes derived from embryonic stem cells. Another important protein in muscle tissue is extracellular matrix protein, accounting for about 10%−20%. The main matrix protein in meat is collagen, which can be produced by fibroblasts and smooth muscle cells, or adipose tissue derived stem cells. More research is needed on how to isolate the cell candidates from animal tissues and culture them at low cost and apply them to create different cultured meat products.
However, primary adult stem cells have the disadvantage that they generally have limited replicative capacity compared to ESCs or iPSCs. In addition to primary adult stem cells, several research institutions and startups have therefore also used spontaneous or gene-edited immortal cell lines, or small molecule modified cells as seed cells for cultured meat production[2]. Cell lines obtained by this method are generally more replicative, which means that the efficiency of future scaling-up is higher, and the cost lower than that for primary cells. However, there are still regulatory and consumer acceptance problems of such schemes that modify cell biology, more so with genetic modification than with small molecule modification.
Low-cost serum-free medium
-
Cell culture medium contains basal culture medium and serum to provide nutrients for cell growth. However, serum of animal origin contains undefined substances and is not sustainable, which doesn’t fit with the purpose of cultured meat. Cell culture medium is one of the main factors contributing to the high cost of cultured meat. Therefore, the development of low-cost serum-free medium will become an important prerequisite for the industrialization of cultured meat. Serum-free media have been developed for ESC and MSC. It is important to develop serum-free formulations suitable for cultured meat seed cells[4].
Serum-free medium development strategies usually involve optimizing basal media and compounds that affect specific cellular pathways, such as growth factors, hormones and small molecules. DMEM/F12 is a nutrient-rich basal medium, which has been greatly optimized in the field of biopharmaceuticals to reduce cost and be used for the growth of different cells due to its sufficient, possibly excessive, nutrient content. For cell specific pathway molecules, progress in research has led to a deep understanding of cell proliferation and differentiation, e.g. FGF signaling pathway in muscle stem cell proliferation. The rapid development of transcriptomics, proteomics and metabolomics can also help better understand the nutritional requirements and metabolic characteristics of cells, which can improve the efficiency of serum-free medium development. In addition, high-throughput screening tests and high-throughput microscopy allow rapid screening of compounds that regulate self-renewal and differentiation of cells, facilitating serum-free medium development[5]. Several biopharmaceutical media companies have a large array of serum-free formulations, and in-depth analysis can identify a range of common nutrients to help accelerate the development of serum-free media for new cell lines. Finally, the use of high-throughput workstations and rapid cell metabolism detection combined with artificial intelligence for medium optimization and prediction, can further help improve the efficiency of medium optimization.
After the formulation is developed, the next step will be to generate low-cost food-grade alternatives of these media. Several media companies already have well-established supply chains of basic ingredients. Synthetic biology can be used to produce other functional factors at a low cost and improve the stability of culture media[6]. In addition, the use of plant and microbial hydrolysates to replace the relevant components in the cell culture medium can also contribute to low-cost food-compatible medium. Finally, if some key compounds cannot be removed or replaced by food-grade ingredients, it is necessary to trace their metabolokinetics in cell culture and seek ways to replace them by compounds with similar function.
Scaling up
-
Cultured meat requires a large number of cells to form cultured meat products. How to culture seed cells on a large scale is an important problem restricting the industrialization of cultured meat. The traditional two-dimensional culture method has disadvantages such as the low surface-area-to-volume-ratio, the inability to monitor culture conditions in real time and the cumbersome passaging process, which is not suitable for scaling up.
It is necessary to consider suspension culture, either microcarrier-based, cell aggregate-based or as single cells, in suitable bioreactors. Suspension culture technologies have been developed for embryonic stem cells, mesenchymal stem cells and fibroblasts. Muscle stem cells are a special kind of stem cell due to their adhesion and cell fusion ability, so microcarrier-based suspension culture seems to be a better choice. However, lessons from biopharmaceutical applications indicate that it is difficult to achieve low-cost culture based on microcarrier suspension culture. It appears inevitable to develop suspension culture for muscle and fat precursor cells that employ inexpensive, biodegradable microcarriers[7] or dispense with microcarriers altogether. Using food-grade microcarriers could potentially realize integration of cell proliferation and differentiation in one bioreactor step. However, it is necessary to consider how to multiply cells more than 106 times on the microcarriers to obtain sufficient scaling up in a bioreactor run.
The bioreactors used for mammalian scale-up production are mainly stirred tanks and rocking bioreactors. In future, it is necessary to design larger bioreactors to promote the large-scale production of seed cells for cultured meat.
Cultured meat tissue formation
-
In order to produce proper cultured meat for consumption, these cells need to undergo differentiation towards muscle protein forming fibers and proper fat cells or tissues. Tissue engineering techniques can be used to produce meat-related tissue. Scaffold biomaterials are the key components of cultured meat and are used for cell proliferation and differentiation to form tissues. Non-animal sources of polysaccharides, proteins and other materials can be used as scaffolds for cultured meat. The characteristics of scaffolds will also have an impact on the final product of cultured meat. Therefore, it is necessary to combine the seed cells, scaffolds and appropriate tissue engineering techniques to make the most suitable cultured meat products.
There are two approaches for tissue engineering including bottom-up and top-down, both of which can be used to build cultured meat products. Two recent articles take both approaches respectively. Some studies and startups used a top-down approach to develop cultured meat tissue by adding cells into edible porous scaffolds[8]. In fact, using the top-down approach, it has become a trend in cultured meat field to culture cells into a structured plant scaffold to produce a hybrid culture meat/plant-based product with acceptable texture and taste. This approach can quickly implement the construction of larger muscle tissues, but it has drawbacks including poor tissue function and non-uniform cell distribution. The bottom-up approach can generate tissue by pre-mixing cells and biocompatible materials to form tissue by 3D printing[9] or structured molding. In this way, smaller tissues are generally made, which need a final assembling process in a patty or other defined meat products. Hybrid products may achieve lower-cost cultured meat products and may be easier to industrialize.
Differentiation of muscle and fat tissue at scale requires the development of novel, dedicated bioreactors where tissues are generated in an automated fashion. Differentiation conditions for muscle and fat tissue are very different and it is easier to keep these processes separated. When complex multi-cell type tissues are cultured, one of the challenges is to define conditions where the cell types can simultaneously differentiate. The aspect of differentiation has not yet received much attention, but is of significant importance for the realization of bona fide cultured meat.
-
Cultured meat is produced in a different way to traditional meat production, so regulation needs to be considered. In December 2020, the Singapore Food Agency (SFA) became the first national regulatory authority globally to approve a cultured meat product. During the regulatory process (which took over two years), the cultured chicken meat produced by Eat Just was approved and used as an ingredient in the company's cultured chicken products. This approval is not an overall approval for cultured meat products, but only for the product and manufacturing process of Eat Just. Safety and nutrition indicators such as microbial content, amino acid composition and other indicators have been rigorously reviewed. Singapore's unique cultured meat regulatory system has also attracted more cultured meat companies to focus on Singapore as first entry into the market.
In the United States, the Food and Drug Administration (FDA) has conducted many discussions with United States Department of Agriculture (USDA) on the regulation of cultured meat and formulated a regulatory framework (excluding seafood). The FDA will oversee cell collection/storage and all culture inputs and processes until biomass is harvested from bioreactors, and the USDA will oversee subsequent processing and labeling of meat. In the European Union, cultured meat produced without genetic modification, is regulated by the EU's Novel Foods Regulation of the European Union. The company must obtain authorization from the European Commission before its cultured meat product can enter the market. The application should consist of safety information, the identity of the product, its production process, compositional data and specifications, proposed uses, use level and anticipated intake of the product[2]. In China, cultured meat products should be regulated as 'New Food Raw Materials'. Cultured meat companies should submit applications to the National Health Commission, including raw material development reports, safety assessment reports, an analysis of the production process and other materials.
For the regulation of cultured meat, it will be very important to have uniform safety standards that should include the identity of the product, its production process, compositional data and specifications, proposed uses, use level and anticipated intake of the product. Important safety information contains absorption, distribution, metabolism and excretion (ADME); nutritional and toxicological information and allergenicity of cultured meat product. International consensus on such safety information would greatly simplify the regulatory landscape.
-
As a novel food, the consumers' acceptance of cultured meat will determine commercial success. A considerable number of surveys have been conducted on consumers' acceptance of cultured meat, which provides ample support for the entry of cultured meat into the market. In western countries[2], when consumers receive more information about cultured meat, they are more inclined to try cultured meat products. The social benefits or the similarities to conventional meat are more appealing than the technological innovations around cultured meat. Price is also an important factor, as consumers are more willing to buy cultured meat when the price is lower. The acceptance of cultured meat is higher in men, younger people, and omnivores than in women, older people and vegetarians. Cultured meat is more attractive among American and Asian consumers than in Europe. Our lab in China also produced China’s first cultured pork and held a tasting event in June, 2020 (Fig. 3). According to a recent survey in China, knowledge of cultured meat has increased people's acceptance. More than 70% of respondents were willing to taste or purchase cultured meat[10].

Figure 3. On June 6, 2020, cultured pork was tasted in Nanjing Agricultural University.
Almost all of the current surveys are based on hypothetical cultured meat products. Because there are no commercially available cultured meat products, researchers can't observe consumer preferences in practice. However, these surveys can help cultured meat companies to develop targeted marketing strategies. The focus of future consumer acceptance studies will be in Singapore, as soon as cultured meat products become more widely available. With availability of products, the next level of consumer acceptance studies will provide more insight into the attributes of cultured meat products that are important for consumers.
-
The progress of science and technology, coupled with the attention to livestock meat production, has promoted the industrialization of cultured meat. Although cultured meat products have been approved in Singapore, low-cost cultured meat industrial production remains a challenge. Producing meat-like products, containing cells that express meat proteins within an edible, plant-based scaffold may be easier to commercialize than pure cultured meat products. The seed cells, low-cost serum-free medium, scaling up and cultured meat tissue formation still need more research and development. For the regulation of cultured meat, a uniform safety standard is essential and current consumers' acceptance survey will help cultured meat companies to develop targeted marketing strategies. We believe that the cultured meat industry will develop rapidly with the synergy of technology, regulation and consumer acceptance and bring tastier, healthier and more sustainable meat products for consumers in the future.
- The authors declare that they have no conflict of interest.
- Copyright: © 2021 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ding S, Post MJ, Zhou G. 2021. Perspectives on cultured meat. Food Materials Research 1: 3 doi: 10.48130/FMR-2021-0003

Perspectives on cultured meat
- Received: 30 October 2021
- Accepted: 30 November 2021
- Published online: 14 December 2021
Abstract: Cultured meat is a promising alternative protein technology that aims to produce healthier, safer, and more sustainable meat. It is muscle and fat tissue generated in vitro by the culturing and biomanufacturing of animal cells for human consumption. However, producing cultured meat at large scale and low-cost is still a difficult task. Technology, regulation and consumer acceptance are all challenges for cultured meat production. In this perspective, we divided the industrial production of cultured meat into three goals according to the cell types and complexity. We discuss the technologies used to achieve low-cost cultured meat industrial production. We also discuss the regulation and consumer acceptance and how both can accelerate commercialization.
-
Key words:
- Perspectives /
- Cultured /
- Meat