










-
Plant cell walls are complex and dynamic structures mostly composed of diverse polysaccharides and cell wall proteins (CWPs)[1]. Plant primary cell walls (PCWs) are dynamically modified, reorganized, and loosened to allow for wall relaxation and cell expansion[2]. Polysaccharides of plant PCWs mainly contain pectin, hemicelluloses and cellulose. In addition, some cells of certain plant tissues develop thick secondary cell walls (SCWs). These specialized cells form SCWs inside the PCWs upon completion of cell expansion, which provide mechanical strength and water-conducting capabilities[3,4]. CWPs play key roles in cell wall formation and modification and adaptation to the environment[5, 6]. Wood is a highly developed cell wall organization, of which the fibers and vessels develop thick SCWs. Some glycosylated proteins have been proposed to function in wood formation via genetic evidence[7].
Glycosylation is an important post-translational modification of proteins, affecting many protein functions and cellular activities. It is estimated that 50% of all proteins are glycosylated based on the fact that two-thirds of entries in the Swiss-Prot database were found to contain at least one N-glycosylation consensus (NXS/T)[8,9]. There are two main types of protein glycosylation: N-glycosylation and O-glycosylation in plants[10]. N-linked glycosylation is a common feature of plant proteins, particularly the CWPs that fulfill important roles in cell wall modification, sugar metabolism, signaling, and defense[11]. In addition, a structural role of hydroxyproline-rich O-glycoprotein extensins (EXTs) has been clearly assigned in maintaining the growing cell walls[12]. Therefore, it is necessary to identify the glycoproteins of tree stem xylem for the understanding of wood formation.
Lectin affinity chromatography (LAC) is a technique that uses different immobilized lectins to reversibly bind to specific sugar residues of the glycoproteins. Because of its specificity and rapidity, LAC has become a common tool for affinity purification of the glycoproteins[13]. Coupled with mass spectrometric (MS) technologies, a growing number of glycoproteome have been performed in different species. Over two decades, some glycoproteins have been identified as biomarkers or indicators of disease[14−16]. In plants, glycoproteome has been performed in tomato, Arabidopsis, rice, and cotton[17−22]. These studies indicate that the CWPs are a large number of glycoproteins. However, little is known about the identification of the glycosylated proteins in stem xylem in trees.
Poplar is a fast-growing tree with a large biomass accumulation in terrestrial ecosystems, extensively used in the pulp and paper industry, reforestation of land and bioenergy feedstocks. The objective of this study was to perform multiple lectin affinity-based glycoproteome of stem developing xylem in poplar. As a result, we identified 154 glycoproteins from the developing xylem of poplar. These glycoproteins were divided into nine functional groups, and 63% were located in the cell wall and the plasma membrane. In combination with the previously published research on functional reports, some glycoproteins are proposed to be involved in cell wall synthesis modification during wood formation. However, a large number of the genes encoding the identified glycoproteins are not still elucidated in function. Our study provides a significant foundation for further investigations into the potential roles of the identified glycoproteins in wood formation.
-
Total soluble proteins were extracted from developing xylem tissues of three-year-old poplar trees. The experiment was performed three times, and all extracts were mixed into total crude proteins. To enrich more numerous glycoproteins, we designed an affinity purification strategy, in which four kinds of plant lectins were integrated to enrich different types of glycoproteins (Fig. 1a). Concanavalin A (ConA) and wheat germ agglutinin (WGA) bind to the glycoproteins with mannosyl and glucosyl residues and N-acetyl-glucosamine and sialic acid, respectively[23−27]. Jacalin specifically binds galactosyl (β-1,3) N-acetylgalactosamine of O-linked glycoproteins, while peanut (PNA) can specifically recognize β-galactose[28, 29]. After total crude proteins flowed through the ConA-WGA-Jacalin-PNA affinity column, the captured proteins were eluted with the Elution Buffer containing 500 mM methyl D-glucopyranoside for Con A, 500 mM N-Acetyl-D-glucosamine for WGA, 500 mM N-Acetyl-D-galactosamine for PNA and 500 mM galactose for Jacalin, respectively. Afterwards, the eluted proteins were analyzed using Coomassie brilliant blue (CBB) or glycoprotein-specific (GS) staining on SDS-PAGE gels. The result showed that the bands of these purified proteins were similar on the CBB and GS staining gels (Fig. 1b), suggesting that these proteins purified using plant lectin affinity could be glycoproteins.
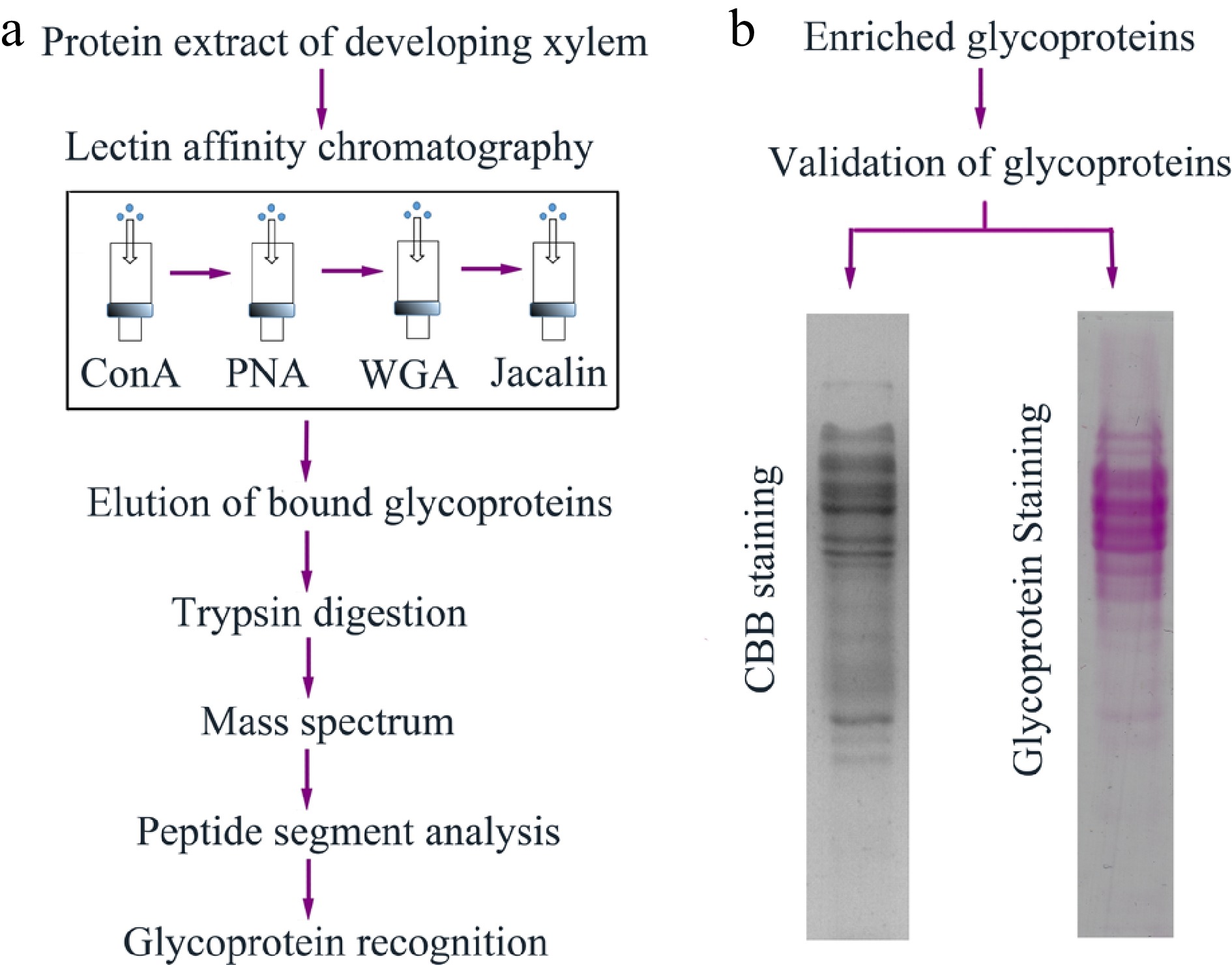
Figure 1.
The workflow of glycoprotein enrichment and identification in poplar developing xylem. (a) Enrichment and recognition of the glycoproteins. Crude proteins were extracted from poplar developing xylem, and the glycoproteins were bound to ConA, PNA, WGA and jacalin lectin affinity columns. The eluted glycoproteins were digested by trypsin and the peptide segments were further recognized through mass spectrum analysis. (b) Validation of the eluted glycoproteins. The glycoproteins separated on sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) gels were stained using Coomassie brilliant blue (CBB) staining and Pierce Glycoprotein Staining kit (detecting sugar moieties of the glycoproteins), respectively.
Identification of the proteins enriched by lectin affinity
-
These enriched proteins were digested with the trypsins and the peptide mixtures were analyzed by LC-MS/MS. Based on the annotation of the Populus protein database, the peptides were further identified and assembled into protein identifications (Supplemental Table S1). A total of 154 proteins were identified by MS and database retrieval (Table 1). Among them, 36 proteins were matched with a peptide sequence, and the remaining coincided with more than two peptides. In addition, the peptide number, the score, and coverage rate for each identified protein are shown in Table 1.
Table 1. Identification of the proteins enriched by lectin affinity from poplar developing xylem.
Protein name Gi number Phytozome
accession no.Peptides Score Cov % N-/O-linked sites Proteins acting on polysaccharides (47) GH3 Beta-xylosidase 222845455 Potri.001G206800 9 432 16 Y/Y GH3 Beta-xylosidase 222846715 Potri.001G089100 6 188 10 Y/Y GH3 Beta-xylosidase 222861083 Potri.014G122200 6 285 11 Y/N GH3 Beta-xylosidase 222844484 Potri.002G197200 1 76 1 Y/N GH3 Beta-glucosidase 222852772 Potri.007G114300 1 48 1 Y/Y GH5 Mannan endo-1,4-beta-mannosidase 222855167 Potri.006G109900 2 86 6 Y/Y GH16 Xyloglucan endo-transglycosylase 118481141 Potri.003G159700 6 483 26 Y/N GH16 Xyloglucan endo-transglycosylase 124109187 Potri.001G071000 6 414 28 Y/N GH16 Xyloglucan endo-transglycosylase 222857312 Potri.013G005700 2 203 9 Y/Y GH17 Glucan endo-1,3-beta-glucosidase 222850378 Potri.009G076500 3 86 8 Y/Y GH17 Glucan endo-1,3-beta-glucosidase 222858075 Potri.013G059700 12 1333 39 Y/N GH17 Glucan endo-1,3-beta-glucosidase 222862285 Potri.019G032900 10 1131 29 Y/Y GH17 Glucan endo-1,3-beta-glucosidase 222873604 Potri.018G150400 7 671 20 Y/Y GH 27 Alpha-galactosidase 222862356 Potri.019G056700 1 64 2 Y/Y GH28 polygalacturonase-like 222843096 Potri.002G162400 5 446 20 Y/N GH28 polygalacturonase 222863392 Potri.010G005500 3 167 9 Y/N GH28 polygalacturonase 222838571 Potri.008G211500 3 160 10 Y/N GH28 polygalacturonase 222861707 Potri.014G112100 2 134 7 Y/N GH28 polygalacturonase 222837934 Potri.003G131700 1 125 3 Y/Y GH28 polygalacturonase 222860156 Potri.011G159000 2 84 4 Y/N GH28 polygalacturonase 222843280 Potri.002G186900 2 60 6 Y/N GH28 polygalacturonase 222867323 Potri.016G051200 2 53 4 Y/Y GH31 Glucan 1,3-alpha-glucosidase 222853440 Potri.007G100000 10 287 13 Y/N GH31 Glucan 1,3-alpha-glucosidase 222856503 Potri.005G069000 4 97 6 Y/Y GH32 beta-fructofuranosidase 222868827 Potri.015G127100 2 46 5 Y/Y GH38 alpha-mannosidase 222843486 Potri.002G238200 14 859 19 Y/N GH38 alpha-mannosidase 222861848 Potri.014G143600 10 521 15 Y/N GH38 alpha-mannosidase 222859443 Potri.012G106500 5 374 7 Y/Y GH38 alpha-mannosidase 222859442 Potri.012G106400 1 50 7 N/Y GH51 Alpha-L-arabinofuranosidase 222853916 Potri.006G029900 2 269 5 Y/Y GH127 Beta-L-arabinofuranosidase 222845043 Potri.001G018200 5 154 7 Y/Y Alpha-fucosidase 222863630 Potri.010G047900 3 151 11 Y/N Xylose isomerase 222865922 Potri.T093900 13 998 37 N/N Pectinesterase 222861105 Potri.014G127000 2 285 6 Y/Y Pectinesterase 222844452 Potri.002G202600 1 263 3 Y/Y Pectin lyase 118488323 Potri.003G175900 2 71 6 Y/Y Acetylglucosaminyl transferase 222845138 Potri.001G068100 2 48 7 Y/N Glycopeptide N-glycosidase 222859921 Potri.011G109700 7 268 11 Y/Y Glucosidase II beta subunit 222872983 Potri.006G061600 1 74 4 Y/N Fasciclin-like arabinogalactan protein 222861509 Potri.014G071700 2 99 7 Y/Y Fasciclin-like arabinogalactan protein 47717933 Potri.015G129400 2 96 10 Y/Y Fasciclin-like arabinogalactan protein 118482997 Potri.012G127900 2 84 10 Y/Y Non-classical arabinogalactan protein 118482413 Potri.002G093100 1 82 10 N/Y Non-classical arabinogalactan protein 118481929 Potri.004G044700 1 79 6 Y/Y COBRA-like protein 118485798 Potri.010G001100 5 232 12 Y/Y COBRA-like protein 118488472 Potri.015G060100 4 145 11 Y/N COBRA-like protein 118482010 Potri.015G060000 1 50 2 Y/Y Oxido-reductases (43) Multicopper oxidase, SKU5-like protein 222871142 Potri.001G120300 13 1097 36 Y/Y Multicopper oxidase, SKU5-like protein 222840952 Potri.003G112700 12 944 34 Y/Y Multicopper oxidase, SKS1-like protein 222859558 Potri.012G126400 5 232 14 Y/Y Multicopper oxidase, SKS1-like protein 222868828 Potri.015G127200 3 167 8 Y/Y Multicopper oxidase, SKS4-like protein 118487967 Potri.004G010100 1 182 2 Y/N Multicopper oxidase 222857214 Potri.005G247700 10 1031 30 Y/N Multicopper oxidase 222842395 Potri.002G013700 14 997 27 Y/N Multicopper oxidase 222853065 Potri.007G038300 10 844 29 Y/Y Multicopper oxidase 118488761 Potri.001G219300 9 526 26 Y/Y Multicopper oxidase 222849177 Potri.004G180500 12 470 26 Y/N Multicopper oxidase 222843342 Potri.002G227600 2 194 6 Y/Y Multicopper oxidase 222855045 Potri.006G087500 4 142 9 Y/Y Multicopper oxidase 222844867 Potri.001G000600 1 109 2 Y/N Multicopper oxidase 222849246 Potri.009G159700 4 56 9 Y/Y Laccase 222852007 Potri.010G183500 6 1201 16 Y/Y Laccase 222852006 Potri.008G073700 9 1113 23 Y/Y Laccase 222864170 Potri.010G183500 5 1006 13 Y/Y Laccase 222864171 Potri.010G183600 5 639 13 Y/N Laccase 222854184 Potri.006G096900 3 95 7 Y/Y Laccase 222849832 Potri.009G034500 3 79 7 Y/N Laccase 222850532 Potri.009G042500 1 65 2 Y/Y Laccase 222846554 Potri.001G054600 1 64 3 Y/N Laccase 3805960 Potri.010G193100 1 46 2 Y/Y peroxidase 115345276 Potri.003G214700 5 514 21 Y/N Peroxidase 118487605 Potri.005G195600 3 55 12 Y/Y FAD-Berberine enzyme 545aa 222860154 Potri.011G158700 7 323 15 Y/Y FAD-Berberine enzyme 222846288 Potri.001G462800 5 298 10 Y/N FAD-Berberine enzyme 222833370 Potri.006G128900 3 223 5 Y/N FAD-Berberine enzyme 222872123 Potri.011G159600 2 175 3 Y/N FAD-Berberine enzyme 222846286 Potri.001G462600 2 175 2 Y/Y FAD-Berberine enzyme 222858409 Potri.012G034700 2 175 3 Y/N FAD-Berberine enzyme 222834675 Potri.011G160300 2 173 5 Y/N FAD-Berberine enzyme 222872175 Potri.011G161500 2 89 4 Y/N FAD-Berberine enzyme 222846302 Potri.001G464700 1 83 2 Y/Y FAD-Berberine enzyme 222872118 Potri.011G161400 1 80 2 Y/N FAD-Berberine enzyme 222847838 Potri.001G470100 1 80 2 N/Y FAD-Berberine enzyme 222860155 Potri.011G158800 1 80 2 Y/Y Protein disulfide isomerase 222842706 Potri.002G082100 23 3116 50 Y/Y Protein disulfide isomerase 118485031 Potri.009G013600 14 648 29 Y/N Protein disulfide isomerase 222846968 Potri.001G183500 2 55 5 Y/Y Chitinase-like 118481023 Potri.010G141600 2 65 9 Y/Y Cytochrome P450 222868639 Potri.015G085800 1 56 N/Y Cu/Zn superoxide dismutase 4102861 Potri.005G044400 1 63 10 Y/Y Proteases (26) Aspartyl protease 118482048 Potri.001G028200 5 244 15 Y/Y Aspartyl protease 439aa 222847473 Potri.001G306200 1 90 3 Y/N Serine carboxypeptidase 222849960 Potri.009G003100 2 129 7 Y/N Serine carboxypeptidase 222850469 Potri.009G055900 4 88 8 Y/Y Serine carboxypeptidase S28 222854432 Potri.006G207900 3 499 10 Y/Y Serine carboxypeptidase S28 222836225 Potri.007G015400 6 428 14 Y/N Serine carboxypeptidase S28 118487876 Potri.007G015300 5 418 13 Y/N Serine carboxypeptidase S28 222853228 Potri.007G008100 4 224 12 Y/N Subtilase family protein 222860749 Potri.011G155400 4 269 7 Y/N Subtilase family protein 222875305 Potri.001G440300 5 243 9 Y/N Subtilase family protein 222848475 Potri.004G173900 1 81 1 Y/Y Subtilase family protein 222854095 Potri.006G076200 3 76 5 Y/Y Peptidase M20/M25/M40 222863686 Potri.010G076100 11 1315 33 Y/N Peptidase M20/M25/M40 118486005 Potri.009G169300 12 912 41 N/Y Peptidase M20/M25/M40 222837797 Potri.004G208100 4 526 12 N/Y Peptidase M20/M25/M40 222842722 Potri.002G085400 5 205 16 Y/Y Peptidase M20/M25/M40 222840651 Potri.003G045200 2 131 5 Y/N Peptidase M28 family 222855209 Potri.006G153300 4 178 10 Y/N Cysteine proteinase 222856445 Potri.005G256000 4 320 16 Y/N Cysteine proteinase 222843627 Potri.002G005700 4 223 21 Y/N Cysteine proteinase 222837653 Potri.004G207600 3 212 12 Y/N Cysteine proteinase 118482340 Potri.006G141700 1 59 3 Y/N Proteinase inhibitor 118485178 Potri.013G112800 3 56 39 N/Y Proteinase inhibitor 118482991 Potri.019G083300 2 51 20 N/Y Amidohydrolase family 222849678 Potri.009G067700 5 122 12 Y/Y Amidohydrolase family 222847228 Potri.001G273400 4 59 11 Y/Y Protein kinase (8) LRR protein kinase 222868332 Potri.016G144100 4 480 7 Y/Y LRR protein kinase 222853199 Potri.007G014700 3 231 4 Y/Y LRR protein kinase 222863806 Potri.010G103000 4 133 7 Y/Y LRR protein kinase 222854082 Potri.006G073900 4 110 6 Y/Y LRR protein kinase 222866571 Potri.018G107400 2 98 2 Y/Y LRR protein kinase 222852307 Potri.008G140500 1 45 1 Y/Y LRR protein kinase 222856570 Potri.005G083000 3 45 3 Y/Y LRR protein kinase 222852450 Potri.008G176900 1 47 2 Y/N Proteins with interacting/binding domains (9) Leucine-rich repeat protein 190897432 Potri.009G064300 9 663 42 Y/N Leucine-rich repeat protein 222853264 Potri.007G001000 5 348 12 Y/N Leucine-rich repeat protein 222854117 Potri.018G151000 2 132 4 Y/Y Leucine-rich repeat protein 222846498 Potri.001G017500 1 46 2 Y/N HSP70 family protein 222867185 Potri.016G019800 13 998 24 Y/N HSP70 family protein 222854802 Potri.006G022100 14 939 20 Y/Y HSP70 family protein 222841104 Potri.003G143600 1 62 2 N/Y Calreticulin family protein 222871704 Potri.005G015100 11 834 34 Y/Y Calreticulin family protein 118485765 Potri.013G009500 10 653 38 Y/Y Proteins related to lipid metabolism(6) Lipase/lipooxygenase 118483838 Potri.005G076900 5 287 40 Y/N Purple acid phosphatase 222865126 Potri.010G158400 3 119 7 Y/N Purple acid phosphatase 222851161 Potri.008G096000 3 109 6 Y/N HAD superfamily protein 222839124 Potri.004G232900 2 63 9 Y/N Type I phosphodiesterase 222855200 Potri.006G150900 2 160 7 Y/Y Type I phosphodiesterase 222872448 Potri.018G066600 1 79 3 Y/Y Amino acid metabolism (4) Amidase family protein 222869309 Potri.015G109400 4 180 12 Y/Y Methionine synthase 222850043 Potri.009G152800 2 56 4 Y/Y Methionine synthase 118483919 Potri.013G061800 2 56 4 Y/Y Cysteine desulfurase 222850426 Potri.009G066000 1 55 Y/Y Miscellaneous proteins (6) Germin-like protein 10 118482567 Potri.002G184900 4 623 19 Y/Y Cyclase family protein 222850275 Potri.009G097300 2 192 13 Y/N Cyclase family protein 118488222 Potri.001G301600 4 95 18 Y/Y Kelch repeat protein 222845394 Potri.001G178500 1 73 Y/N Nucleosome assembly protein 222854259 Potri.006G148600 1 56 3 Y/N Cupin domain protein 222858047 Potri.013G051600 1 55 7 Y/N Unknown function (5) Unknown protein (Duf642) 118486479 Potri.011G087500 4 124 16 Y/Y Unknown protein (Duf568) 118487890 Potri.002G249200 1 54 5 Y/Y Unknown protein (Duf2828) 222850304 Potri.009G091400 1 57 Y/Y Unknown protein 222846617 Potri.001G068800 1 80 6 Y/Y Unknown protein 118486279 Potri.019G076900 2 68 5 Y/Y Glycosylation site analysis of the identified glycoproteins
-
N- and/or O-glycosylation sites of the identified proteins were analyzed by NetNGlyc, NetOGlyc and GlycoEP tools, and the results are shown in Supplemental Table S2. Of all 154 proteins, 153 proteins contained N- and/or O-glycosylation sites and only one did not contain the glycosylation sites. Among them, 143 proteins had the N-glycosylation sites; 56 proteins contained 1−3 N-glycosylation sites, and 41 and 46 proteins with 4−6 and 7~ N-glycosylation sites, respectively (Supplemental Fig. S1a). However, O-glycosylation of protein and its types are more complex in plants and the current tools are difficult to accurately predict O-glycosylation sites in plant proteins. The analysis suggested that the 56 proteins might contain 1−3 O-glycosylation sites, and 17 and 19 proteins with 4−6 and 7~ O-glycosylation sites, respectively (Supplemental Fig. S1b).
To confirm whether the N/O-glycosylation prediction of the identified proteins is reliable, we selected two proteins with less glycosylation sites predicted by the tools for verification. A selected superoxide dismutase (PtSOD) contained two N- and one O-glycosylation sites, while the HAD (PtHAD) only contained two N- glycosylation sites (Supplemental Fig. S1). We first generated PtSOD- and PtHAD-transgenic Arabidopsis plants, respectively. RT-PCR analysis showed that the three transgenic lines overexpressed PtSOD and PtHAD genes, respectively (Fig. 2a, b). Because of the expressed PtSOD or PtHAD with the fusion of FLAG tag, we further detected PtSOD or PtHAD protein level in transgenic lines by Western blot using anti-FLAG antibody. The data showed high PtSOD or PtHAD protein levels in transgenic lines (Fig. 2c, d). In addition, Western blot analysis revealed that the molecular weights (MWs) of the expressed PtSOD and PtHAD proteins were 38 and 37 kDa, respectively (Fig. 2d). The MWs are much bigger than those calculated based on the amino acids of PtSOD or PtHAD proteins, suggesting that protein modification might occur. Next, protein extracts from the PtSOD- or PtHAD-transgenic plants were digested by O-glycosidase and PNGase F, respectively. Western blot analysis showed that the digestion of O-glycosidase or PNGase F accelerated the migration rate of PtSOD on SDS-PAGE gels (Fig. 2e), indicating that the PtSOD protein is both N- and O-glycosylated. The migration of PtHAD digested by PNGase F was accelerated on the SDS-PAGE gel, while that of PtHAD digested by O-glycosidase did not change (Fig. 2f). This suggests that PtHAD protein has N-glycosylation sites and no O-glycosylation sites. Taken together, this data indicates that analysis of N- and/or O-glycosylation sites of the identified proteins by the bioinformatic tools is reliable.

Figure 2.
Verification of N- and/or O-glycosylation in PtSOD and HAD proteins. (a), (b) Analysis of PtSOD or PtHAD gene expression by RT-PCR in their transgenic Arabidopsis plants. AtActin2 serves as a control gene. (c), (d) Western blot analysis of PtSOD1-Flag and HAD-Flag protein levels in corresponding transgenic plants usig anti-FLAG antibody. Coomassie brilliant-stained Rubsico large subunit proteins indicate the loading amount of each sample on SDS-PAGE gels as control. (e), (f) Migration analysis of PtSOD and PtHAD proteins on SDS-PAGE gels. Protein extracts with/without the digestion of PNGase F (N) or O-glycosidase (O) were separated on 10% SDS-PAGE gels and followed by immunoblotting with anti-FLAG antibody.
Functional classification and localization of the identified glycoproteins
-
The identified glycoproteins were classified into nine functional groups based on gene annotations and/or known domains (Fig. 3a), which include protein acting on carbohydrates (30.5%), oxido-reductase (27.9%), proteases (16.9%), protein kinases (5.8%), proteins with interaction domain (5.8%), lipid metabolism (3.9%) and amino acid metabolism (2.6%). The group of proteins acting on carbohydrates mainly includes beta-xylosidase (GH3), xyloglucan endo-transglycosylase (GH16), glucan endo-1,3-beta-glucosidase (GH17), polygalacturonase (GH28), alpha-L-arabinofuranosidase (GH51) and fasciclin-like arabinogalactan protein (FLA). Oxido-reductase cluster contained multicopper oxidases, laccases, FAD-berberine enzymes and peroxidases. Protease is the third largest cluster of the identified glycoproteins, including aspartyl proteases, serine carboxypeptidases, subtilases, peptidases, cysteine proteinases. LRR protein kinases and leucine-rich repeat proteins were also important functional groups of the identified glycoproteins.
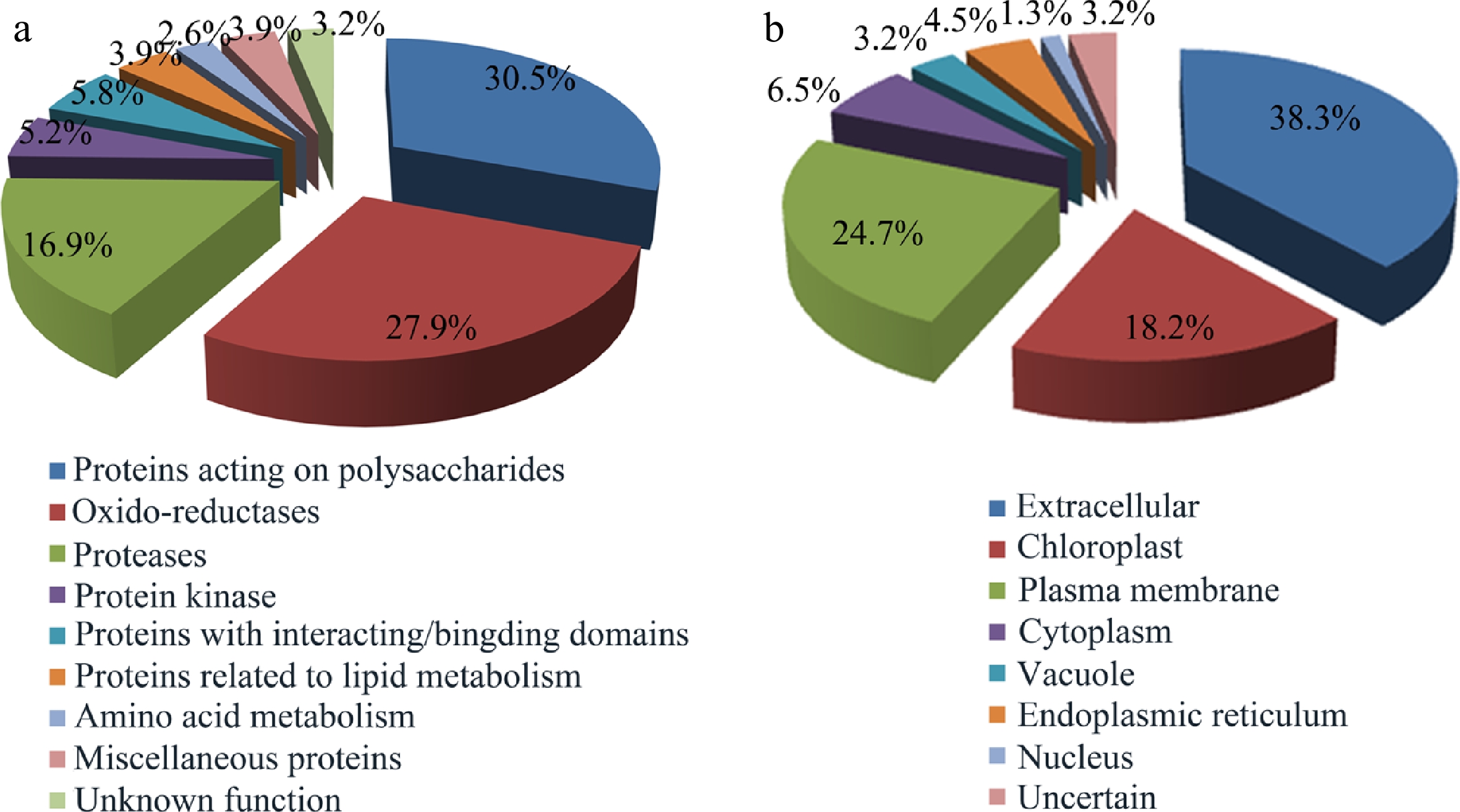
Figure 3.
Functional classification and localization of the identified glycoproteins. (a) Functional classification of glycoproteins identified from Populus developing xylem. Please refer to Supplemental Table S1 for detailed analysis. (b) Subcellular localization of the identified glycoproteins predicted by Plant-mPLoc, ngLOC, ProtComp9.0, WoLF PSORT and YLOC.
Subcellular localization of the identified glycoproteins was analyzed using five bioinformatic tools including Plant-mPLoc, ngLOC, ProtComp 9.0, WoLF PSORT and YLOC. As shown in Fig. 3b, the 38.3% of glycoproteins identified were extracellular proteins (cell wall protein), and the 24.7 and 18.2% of the identified glycoproteins were located in the plasma membrane and chloroplast, respectively. In addition, a small proportion of glycoproteins were located in cytoplasm (6.5%), endoplasmic reticulum (4.5%), vacuole (3.2%), or nucleus (1.3%). Most glycoproteins were secreted proteins, which contain the signal sequences (Table 1).
Expression profiles of the genes encoding the identified glycoproteins
-
To analyze the potential roles of these glycoproteins in developing xylem, the digital expression profiles of the genes encoding the glycoproteins were collected from poplar electronic fluorescent pictograph (eFP) browsers (Supplemental Fig. S2). Of all 154 encoding genes, the 34 genes have no corresponding data in the eFP database. The eFP data showed that profiles of 52 gene expression were high in developing xylem, and transcription levels of 21 genes were high in roots. In addition, 21 and 14 genes were highly expressed in female and male catkins, and only 11 genes in young leaves.
To test the eFP data above, we further examined expression profiles of the 52 genes using RT-qPCR. The results showed that only 25 genes were highly expressed and their expression profiles were more specific in xylem (Fig. 4). These xylem-expressed genes encode laccase, FLA, peroxidase, methionine synthase, and cysteine protease. Laccase, peroxidase and FLA play a key role in secondary cell wall (SCW) formation, and cysteine protease is involved in the process of programmed cell death (PCD) and SCW thickening[30−33]. These expression profiles suggest that many glycoproteins identified should be of importance for wood cell wall synthesis and modification in poplar.
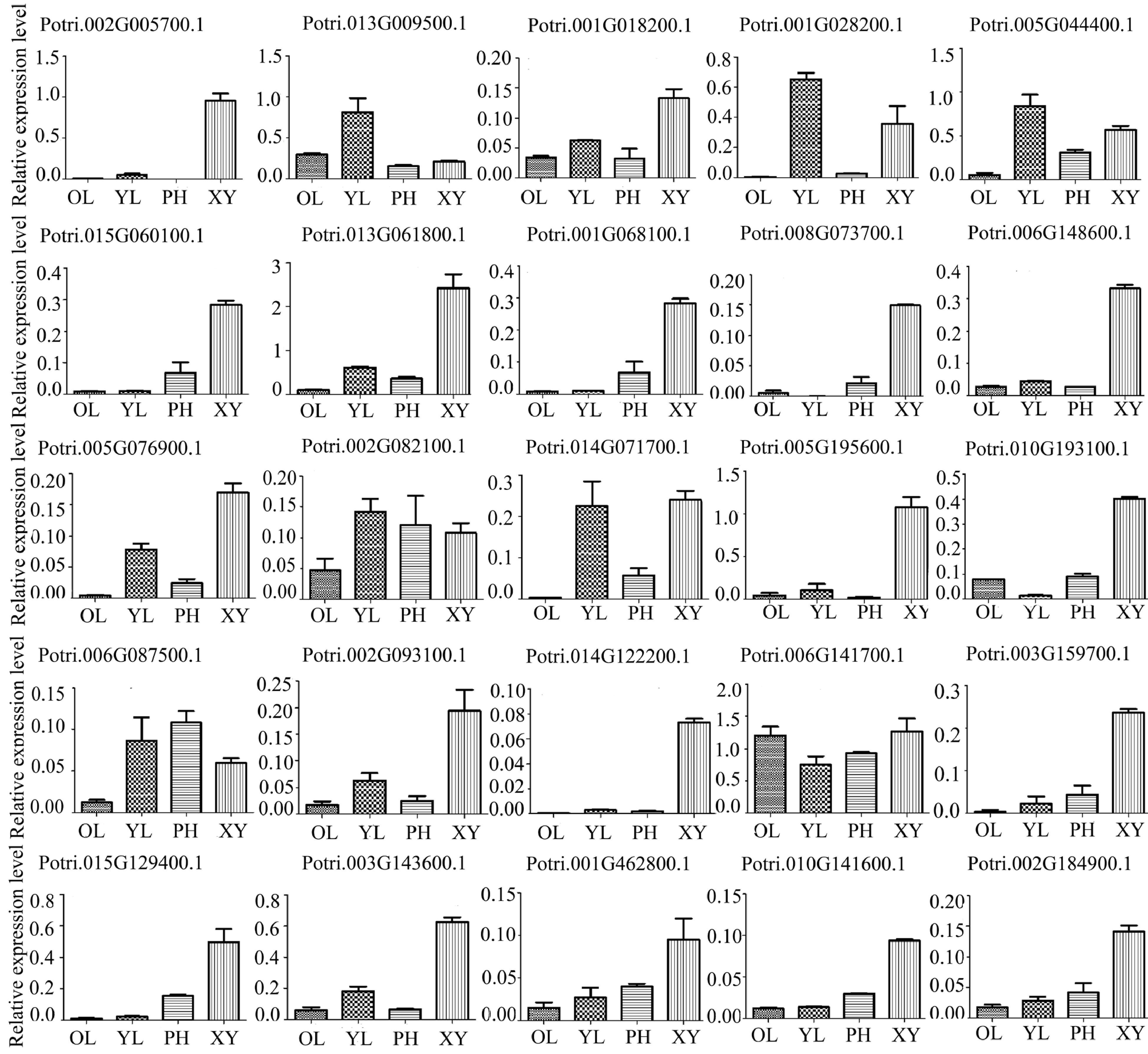
Figure 4.
Expression profiles of 25 genes encoding the glycoproteins in different poplar tissues using RT-qPCR analysis. Different tissues included phloem (PH), xylem (XY), young leaf (YL) and mature leaf (OL). The expression of PtActin2 was used as an internal control. Data are means ± standard error of three technical replicate results.
-
Lectin affinity enrichment is based on the specific binding interaction between lectins and unique glycan structures attached to glycoproteins. A variety of lectins can selectively bind to oligosaccharides, and enrich different types of the glycans of glycoproteins[34−36]. To date, most of the work using LAC for targeted glycoprotein enrichment in plants has focused on N-glycosylation, and binding specificity of the lectin for O-glycosylation is less satisfactory. To capture O-glycoproteins as far as possible, we made serial columns of concanavalin A and jacelin in tandem to isolate O-glycoproteins from the developing xylem in poplar. As a result, many O-glycoproteins were identified in the present study. In addition, some studies rely on two-dimensional electrophoresis (2-DE), which has limitations when used to separate and identify certain types of proteins, such as those that are membrane-associated, less abundant, or have extreme pIs or MWs[37]. Overall, our strategy of this study is a relatively unbiased technology that can more comprehensively identify glycoproteins.
Protein glycosylation occurs in the proteins in the secretory pathway, so a convenient indicator for evaluating the identified glycoproteins is to use software packages searching signal peptides. Up to 89% of the glycoproteins identified from poplar developing xylem were suggested to have signal peptides (Supplemental Table S3). This is much higher than other plant extracellular proteomics, based on the way that LAC is not used[38, 39]. Proteins with signal peptides entering the secretory pathway do not necessarily target the cell wall, but may remain on the endomembrane system, such as endoplasmic reticulum, Golgi apparatus, and other organelles, including vacuoles and chloroplasts. We analyzed the localization of identified glycoproteins using the software subcellular localization website. According to predictive analysis (Fig. 3), the proportion of proteins (38.3%) was located in cell wall, while most of the remaining proteins might be in the plasma membrane (24.7%). It provides a hint that xylem synthesis and modification might be mediated by a number of the glycoproteins in the cell wall and/or plasma membrane.
In this study, most of the identified glycoproteins clustered a functional group of protein acting on carbohydrates (Table 1), suggesting their involvement in wood formation in poplar. Wood formation undergoes a genetically controlled xylogenesis process, which includes cambia cell division, cell differentiation and expansion, SCW synthesis and PCD. As shown in Fig. 5, a number of the glycoproteins identified are involved in wood formation. The PCWs of the growing xylem cells are mainly composed of pectins (such as rhamnogalacturons and homogalacturonans), cellulose, and hemicellulose (xylogucan and mannan). Here we have identified 33 GHs in poplar secondary xylem (Table 1), which belong to 11 types of glycoside hydrolases (GHs). GHs are important cell wall polysaccharide-modified enzymes that participate in the division and expansion of plant cells and their substrates are pectin and hemicellulose[40−46]. GH16, GH31 and GH51 may act on modification of xylans in cell wall, and GH28 can hydrolyze pectin[47, 48]. GH38 may be involved in the modification of the mannose and GH32 as the invertase functions in carbohydrate allocation[49−51]. GH3 and GH5 have broader substrates. It is reported that they are involved in modification and hydrolysis of hemicellulose, as well as lignification and secondary growth[41,45]. In addition to GHs, pectin modifying enzymes, pectin esterase and pectin lyase, affect the plasticity and fluidity of cell walls and play a decisive role in the final shape and size of cells[52, 53]. In addition, peptidase and serine carboxypeptidase affect cell expansion, but the mechanism is unknown[54].
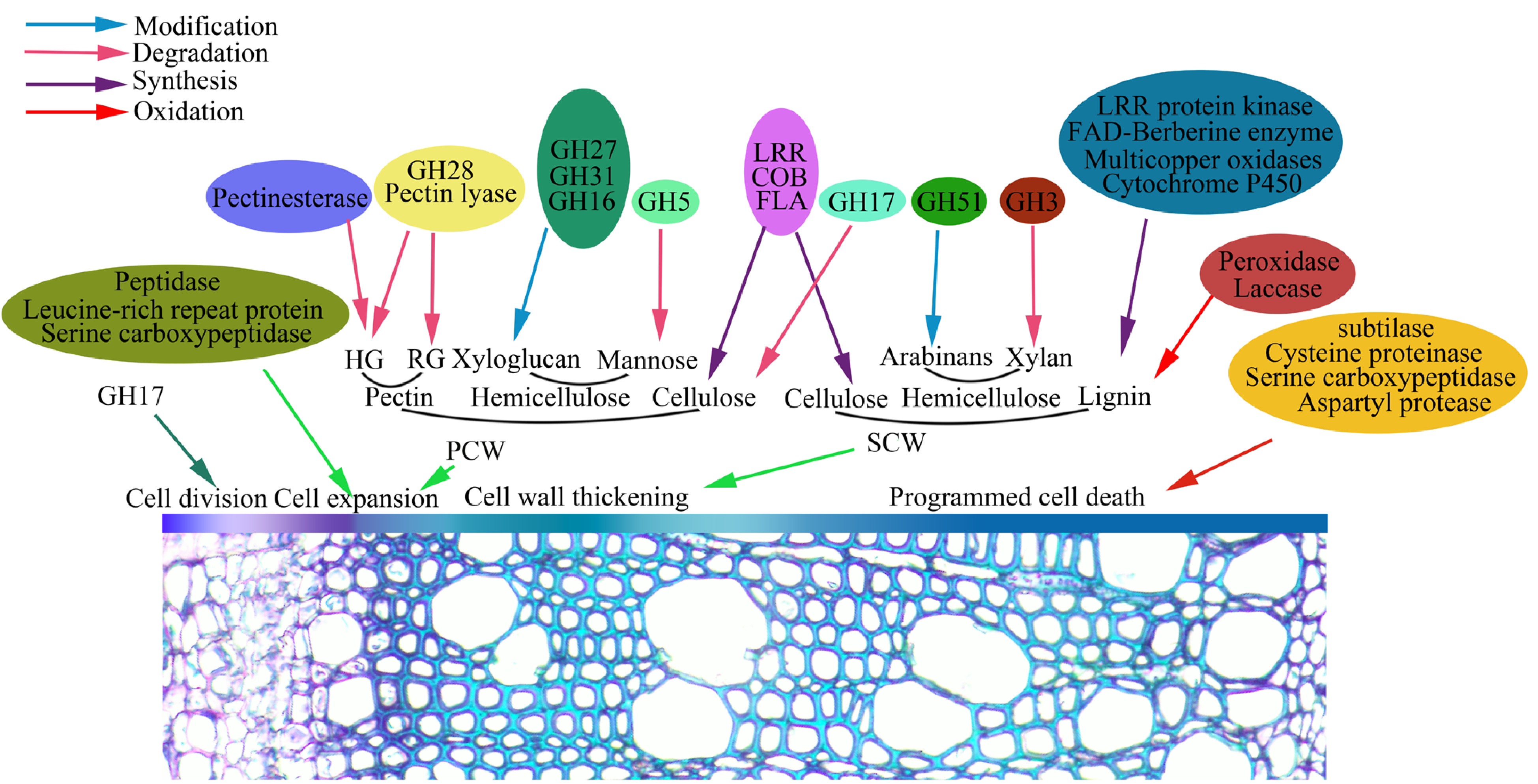
Figure 5.
Identified glycoproteins are proposed to be involved in wood formation that includes cambia cell division, cell differentiation and expansion, SCW synthesis and PCD.
When xylem cells reach the final size and shape, a thicker SCW is produced continuously. In this study, dozens of glycoproteins including laccase, peroxidase and methionine synthase, are specifically expressed in secondary xylem at the transcriptional level, suggested by the RT-PCR analysis (Fig. 4). Lignin is one of main components of secondary xylem (wood) in trees. In this study, 38 glycoproteins identified might be involved in lignin biosynthesis, which include BBE, laccase and peroxidase (Table 1). BBE-like proteins, as monolignol oxidoreductases, may participate in the oxidation of lignin required for polymerization processes, while laccase and peroxidase are responsible for the polymerization of the lignins[55−58]. Other glycoproteins, such as FLAs, COBRA-like protein and LRR protein kinase participate in cell wall thickening, for example, cellulose deposition in the SCW is implemented by the COBRA-like protein[59−62]. Additionally, we also identified several types of proteases such aspartyl protease, serine carboxypeptidase and cysteine proteinase, which are involed in cell death of xylem fibers and vessels[63−65]. Overall, these glycoproteins could be served or proposed as the players in wood formation in poplar. Recently, poplar mannanase PtrMAN6 with the N-glycosylation plays a role in coordinating cell wall remodeling with suppression of secondary wall thickening[66]. Another study reveals that glycan synthesis levels of the AGP proteins change in wood formation[67]. Thus, it is inferred that protein glycosylation as a regulatory way should be involved in wood formation. We are now attempting to detect the roles of glycosylation sites of the glycoprotein in this process through genetic studies.
-
In this study, we used Populus simonii × P. nigra as plant material. Three-year-old trees grown in a forest farm at Northeast Forestry University (Harbin, China, longitude 127°18′0″; latitude 45°2′20″) were selected for correcting developing xylem tissues. Arabidopsis thaliana (Columbia ecotype) plants were grown in the greenhouse (16 h light/8 h dark) at a light intensity of 120 μmol photons m− 2 s− 1 at 22 °C. The CDS of PtHAD or PtSOD was amplified using the xylem cDNAs as a template, and the DNA fragements were constructed into pGWB11 vector with the fusion of FLAG tag for overexpression of PtHAD or PtSOD. After DNA sequencing, the resultant constructs were introduced into A. tumefaciens strain GV3101 for Arabidopsis transformation using the floral-dip method.
Protein extraction
-
Developing xylem tissue was corrected from one young tree on June 15, and we repeated it three times. After the bark was peeled, the xylem tissue was quickly frozen with liquid nitrogen and developing xylem cells were scraped from the outside to the inside. Three corrected sample materials were mixed and ground into a powder. Approximately 50 g powder was saturated into 120 ml protein extraction buffer (50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, 1 mM MnCl2). After the mixture was shaken on ice for 30 min, the homogenate was centrifuged at 40,000 g for 30 min at 4 °C. The supernatant was used for enrichment of glycoproteins using lectin affinity chromatography.
Lectin affinity chromatography
-
The crude protein was used for lectin affinity chromatography. Four plant lectins, concanavalin A (Con A), Triticum vulgaris (WGA), peanut (PNA) and jacalin, are used to specifically enrich different sugar residues of various glycoproteins. We added 0.5 ml ConA-Sepharose 4B (27700, Supelco), WGA-Agarose (L1882, Sigma-Aldrich), PNA-Agarose (AL-1073, Vector Laboratories), and Jacalin-Sepharose (6561, Biovision) to the columns, respectively. After 5 ml binding buffer (20 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, 1 mM MnCl2) was added to clean the column, the crude protein in the supernatant was successively passed through the four columns. Then, each column was washed with 50 ml of binding buffer to remove the unbound proteins. After discarding the binding buffer, the bound glycoproteins were respectively eluted from the four columns with 0.5 ml of elution buffer (20 mM Tris-HCL, pH 7.4, 300 mM NaCl, 1 mM MgCl2, 1 mM CaCl2, 1 mM MnCl2) containing 500 mM methyl α-D-glucopyranoside (M9376, Sigma-Aldrich) for ConA, 500 mM N-Acetyl-D-glucosamine (A8625, Sigma-Aldrich) for WGA, 500 mM N-Acetyl-D-galactosamine (A2795, Sigma-Aldrich) for PNA, or 500 mM galactose (G0750, Sigma-Aldrich) for jacalin. The eluted fractions of the sample were pooled and filtered through Microcon YM-10 centrifugal filter devices to a volume of ~0.2 ml. The sample was used for protein identification.
Protein identification and database searching
-
The sample was digested with porcine trypsin (Promega) at 37 °C overnight, as described previously[68]. After protein digestion of trypsin, the short peptides obtained were performed for LC-MS/MS analysis, as described previously[69]. The MS/MS spectra were searched against the NCBInr protein databases and phytozome databases using Mascot software (Matrix Sciences, UK). The search criteria included a mass accuracy of 0.3 Da, with one missed cleavage allowed, carbamidomethylation of cysteine as a fixed modification, and oxidation of methionine as a variable modification. A highly confident protein identification met the following criteria: (a) top hits in the database searching report; (b) a probability-based MOWSE score of greater than 55 (p > 0.01); (c) more than two peptides matched with a nearly complete y-ion series and complementary b-ion series present. Based on the MASCOT probability analysis, the significant hits were accepted as the identification of each protein.
Microarray data, RNA isolation and RT-qPCR analyses
-
Tissue-specific expression data were downloaded from poplar eFP browser. The heat map was generated by Heat map illustrator (HemI) with the default settings. Total RNA was extracted from plant tissues using plant RNA Extraction Kit (Bio-Flux, China). For each sample, 1 μg of total RNA was reverse-transcribed into total cDNAs using the PrimeScript RT reagent Kit (TaKaRa, China). The qRT-PCR experiments were performed with SYBR Green (TaKaRa, China) in the ABI Prism 7500 system (Applied Biosystems, USA) according to the manufacturer's instructions. The reaction mixture (20 μl) consisted of 10 μl 2× TB Green Premix Ex Taq II (Tli RNaseH Plus), 0.8 μl of each gene-specific primer, 0.4 μl ROX Reference Dye II, 1 μl cDNA template and 7 μl distilled deionized H2O. The PCR parameters as follows: 95 °C for 30 s; 40 cycles of 95 °C for 5 s, 60 °C for 15 s, 72 °C for 30 s. PtActin2 was used as an internal control and the comparative Ct (2−ΔCᴛ) method was used to calculate gene expression levels. Three technical replicates were carried out for each sample.
Deglycosylation experiment and Western blot
-
Transgenic plant materials were ground in liquid nitrogen and homogenized in protein extraction buffer (50 mM Tris-HCl, 200 mM NaCl, 2% SDS, 5 mM DTT, pH 8.0). The suspensions were centrifuged at 18,000 g for 5 min, and the supernatant (protein extract) was used for protein deglycosylation with/without PNGase F and/or O-glycosidase (New England Biolabs, UK). The treated protein extract was resolved in 10% SDS-PAGE gel and transferred into a PVDF membrane. Western blotting was performed using anti-FLAG antibody (Abmart, China) and Pierce ECL chemiluminescent Substrate (Thermo, USA).
Bioinformatics analysis
-
TProtein sublocalization was predicted based on five bioinformatics tools including Plant-mPLoc, ngLOC, ProtComp 9.0, WoLF PSORT and YLOC. Signal peptides were analyzed using the SignalP 4.1 Server (
www.cbs.dtu.dk/services/SignalP ). Glycosylation sites were analyzed using three tools including NetNGlyc 1.0, NetOGlyc 4.0 and GlycoEP. The research was supported by the Fundamental Research Funds for the Central Universities (2572021DT01), the National Natural Science Foundation of China (31770637) and Zhejiang Provincial Natural Science Foundation of China (No. LY20C160010).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Hao Cheng, Jinwen Liu
- Supplemental Table S1 The single spot in 2D gels was identified to contain more than a protein.
- Supplemental Table S2 The subcellular localization prediction of the proteins identified in Populus developing xylem.
- Supplemental Table S3 The glycosylation sites prediction of the expressed proteins identified in developing xylem in Populus
- Supplemental Fig S1 Prediction and analysis of the number of glycosylation sites in glycoproteins. Glycosylation site prediction of identified proteins with three bioinformatic softwares. (1. NetNGlyc 1.0; 2. NetOGlyc 4.0; 3. GlycoEP). Refer to the supplemental Table S3 for detailed analysis.
- Supplemental Fig S2 Hierarchical clustering of the identified glycoprotein expression profiles in different tissues. The microarray data were downloaded from the Poplar eFP browser. Color scale at the right of the dendrogram represents log2 expression values.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Cheng H, Liu J, Zhou M, Cheng Y. 2022. Lectin affinity-based glycoproteome analysis of the developing xylem in poplar. Forestry Research 2:13 doi: 10.48130/FR-2022-0013 

Lectin affinity-based glycoproteome analysis of the developing xylem in poplar
- Received: 11 July 2022
- Accepted: 04 October 2022
- Published online: 27 October 2022
Abstract: Glycosylation is a significant post-translational modification of proteins, and some glycoproteins serve as players in plant cell wall synthesis and modification. Wood is a highly developed cell wall organization, and protein glycosylation as a regulatory mechanism may be involved in wood formation. Here, a lectin affinity-based glycoproteome was performed in stem developing xylem of poplar. After enrichment, trypsin digestion, LC-MS/MS analysis and peptide identification, we identified 154 glycoproteins from poplar developing xylem, which were classified into nine functional groups mainly including protein acting on carbohydrates, oxido-reductase, proteases, and protein kinases. Further, N- and/or O-glycosylation sites of the identified proteins were analyzed using bioinformatic tools, and deglycosylation experiments in the selected PtSOD and PtHAD proteins verified the reliability of the identified glycoproteins. Analysis of protein subcellular localization showed that a total of 63% of the identified glycoproteins were extracellular proteins or located in the plasma membrane. Poplar eFP and RT-qPCR data showed that a number of the genes encoding these glycoproteins such as laccase, peroxidase and cysteine protease, have highly preferential expression profiles in the developing xylem. Together with previously published research, most identified glycoproteins could be involved in wood cell wall synthesis and modification in poplar. Thus, our study provides some potential wood formation-related glycoproteins to be determined during tree stem development.
-
Key words:
- Glycoproteome /
- Xylem /
- Poplar /
- Lectin /
- Wood formation