









-
After millions of years of evolution, terrestrial plant organs exposed to air have been covered with a water-insoluble cuticular layer, which acts as a barrier between the plant organs and tissues and the external environment[1−3]. The plant cuticle is composed of the cuticular membrane of the epidermis and the wax layer. These tissues play an important role in water retention, insect resistance and maintenance of plant organ integrity[4]. Cuticular wax in plant epidermis is mainly divided into two layers: the inner wax embedded in a keratin polymer matrix and the outer wax located outside of the keratin polymer matrix[5]. Under natural conditions, cuticular waxes exhibit different crystal types, including plate, granular, stripped, and banded[5]. Very long chain fatty acids (VLCFAs) and their derivatives are the main components of plant cuticular wax. These derivatives are derived from C16 and C18 free fatty acids, including saturated alkanes, fatty alcohols, fatty aldehydes, ketones, and esters[6]. The cuticular structured terpenoids exist in the ceratine inner layer and include amorphous compounds that are difficult to observe by electron microscopy[7].
The biosynthesis of cuticular wax is affected by many internal and external factors. The chemical composition of epidermal wax of different horticultural crops was significantly different. Triterpenoids and beta diketones constitute the main wax components in mature blueberry fruit[8]. The content of long-chain fatty aldehydes in the cuticular wax is highest in Citrus unshiu and Citrus sinensis[9]. Alkanes, primary alcohols and triterpenoids are the main chemical constituents of epidermal wax in pear[10,11]. In apple, alkanes and fatty alcohols dominate the epidermal wax[12].
The wax composition of apple fruits also varies between different cultivars[13,14]. For example, the content of the secondary alcohol 10-nonacosanol (10-C29-ol) in the wax of the 'Jonagold' apple cuticular is 20.85%, while that of 'Elstar' apple cuticular is missing[15]. The composition of epidermal wax is also influenced by environmental factors such as wind, temperature and light intensity[16−18]. The effect of apple cuticle wax is widely studied on postharvest storage[19−21]. For example, the wax surfaces of some apple cultivars become greasy during storage[15,22,23], which has a negative impact on apple commodity sales. During storage, the contents of alkanes and alcohols in cuticular waxes decreased, while the contents of free fatty acids increased[12]. A study revealed that greasiness is caused by the apparent accumulation of wax components leading to the transformation of wax from a solid to a liquid phase[21], which eventually leads to the greasiness of the apple epidermis and affects fruit quality. Ethylene functions to change the content and composition of 'Starkrimson' fruit epidermal wax, which may change the post-harvest quality of the fruits and the susceptibility of the fruits to diseases during cold storage[19].
The physical structure of waxy crystal in fruit epidermis was also correlated with fruit quality. Pietrysiak & Ganjyal found that Listeria innocua usually adheres to the wax gap between the epicuticle calyx and stem calyx, while bacteria rarely attach to where the wax is smooth and dense at the equator[24]. Thus, the characteristics of the cuticular wax structure are closely related to the prevention and treatment of apple disease.
However, little is known about formation of apple fruit skin wax crystals or the biosynthesis of wax. Given the importance of apple cuticular wax in protecting fruit from biotic and abiotic stresses, we comprehensively analyzed the morphology of epidermal wax crystal, wax content and key gene expression levels of two apple cultivars, 'Golden Delicious' and 'Red Delicious' at different growth and development stages after fruit setting. The results revealed a critical period for cuticular wax biosynthesis in apple, laying a foundation for further study of the molecular and physiological regulatory mechanisms of wax biosynthesis in apple.
-
The apple fruits were sampled 30, 60, 90, 120, and 150 DAF. As shown in Fig. 1a, the apple fruits gradually grew larger with each developmental stage. The lightness of the skin decreased (Fig. 1b) and the redness increased in both cultivars (Fig. 1c). The yellowness of 'Golden Delicious' was gradually decreased, while that of 'Red Delicious' was not (Fig. 1d). Next, we analyzed the physiological characteristics and fruit quality of the two apple cultivars at different developmental stages, including fruit weight (Fig. 1e), fruit surface area (Fig. 1f), TSS (Fig. 1g), TA (Fig. 1h), pericarp tissue conductivity (Fig. 1i), and fruit firmness (Fig. 1j). The data showed that the two apple cultivars at different developmental stages exhibited similar growth patterns.
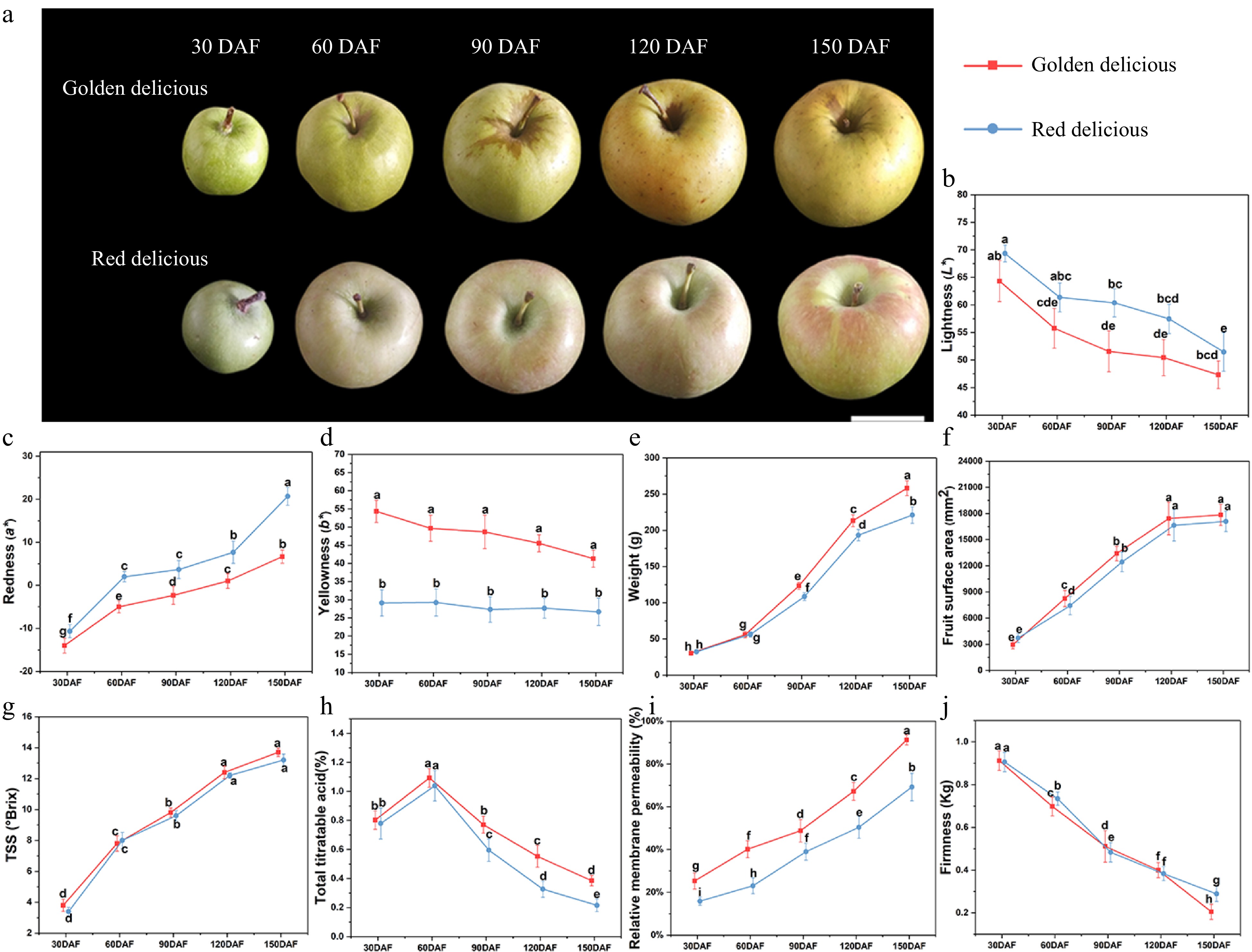
Figure 1. Changes of fruit appearance and quality during growth and development. (a) Fruit phenotypic, (b) lightness, (c) redness, (d) yellowness, (e) fruit weight, (f) fruit surface area, (g) TSS, (h) TA, (i) relative electrical conductivity of pericarp tissue, (j) fruit firmness (P/2 columnar probe diameter: 2 mm). Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan's new multiple range test. Each value is the mean ± standard error.
Changes of skin gloss of apple fruit at different development stages
-
The skin gloss of apple is one of the important appearance qualities of fruit. We detected and analyzed the gloss of two apple cultivars at different development stages. As shown in Fig. 2, the apple cuticular gloss at angles of 20°, 60° and 85° were recorded. At each stage of growth and development, the 20° gloss value of the two cultivars did not differ significantly, and the gloss was between 0.8 GU and 1.5 GU (Fig. 2a). The values of 60° (Fig. 2b) and 85° (Fig. 2c) of the two cultivars varied significantly, both of which reached the lowest at 90 DAF and gradually increased during the subsequent development. At the end of development (150 DAF), 'Red Delicious' glossiness is significantly higher than 'Golden Delicious' (Fig. 2b & c).
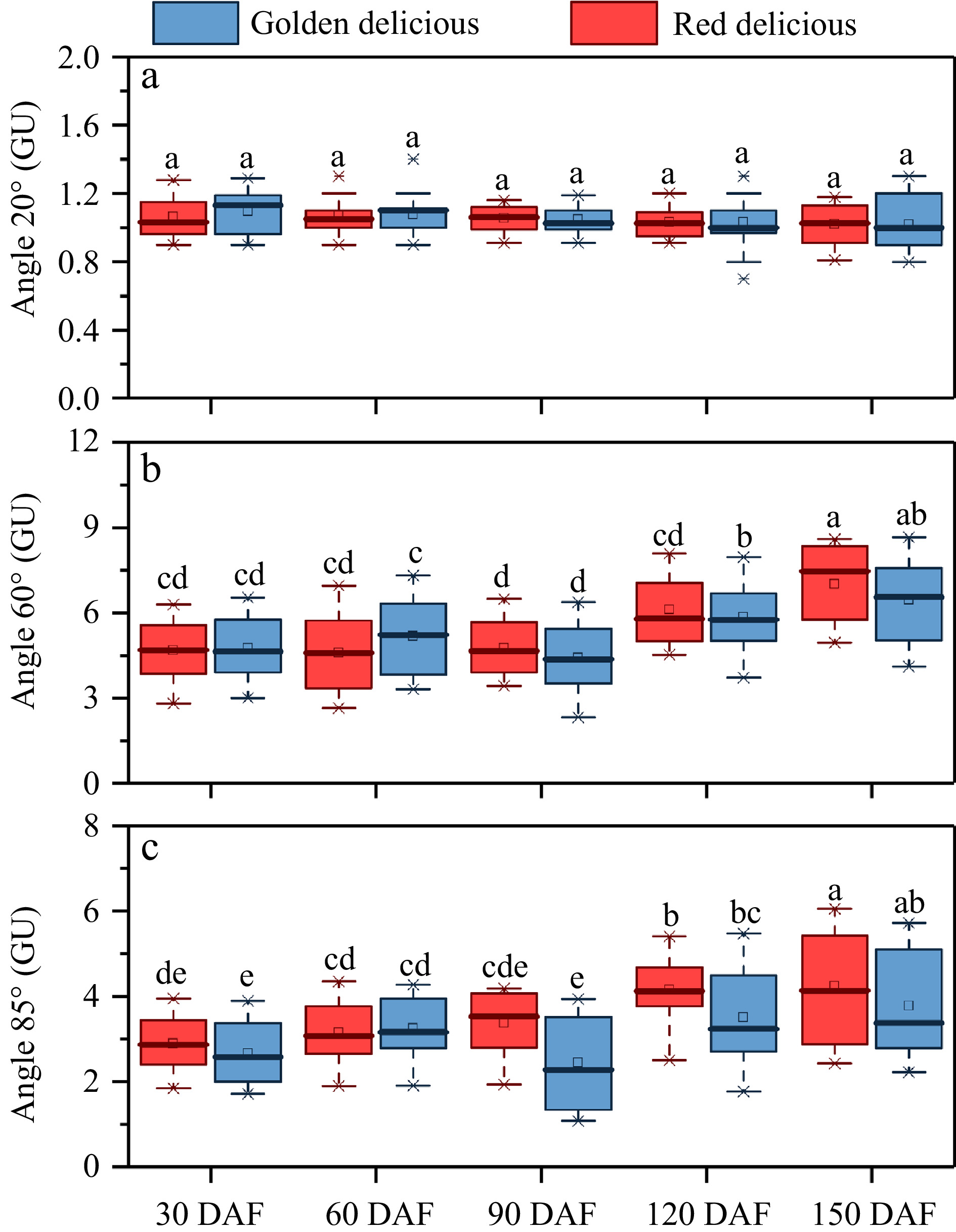
Figure 2. Changes in appearance gloss during fruit growth and development. (a) Angle 20°, (b) angle 60°, (c) angle 85°. Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan's new multiple range test. Box edges represent the 0.25 and 0.75 quantiles, and the bold lines indicate median values. Whiskers indicate 1.5 times the interquartile range. Each value is the mean ± standard error.
Wax morphology of the apple surface during fruit development
-
To understand the formation process of epidermal wax crystals at apple growth and development, the epidermal wax morphology was observed by environmental field SEM. A microscopic examination indicated an obvious increase in the epidermal wax coverage of both apple cultivars at 90 DAF under a low power microscope (×150) (Fig. 3a, G1−G5 and R1−R5). Wax accumulated gradually from 90 to 120 DAF. During subsequent development, we observed a continuous accumulation of wax in the epidermis of the two cultivars. At 150 DAF, the wax in the apple epidermis almost completely covered the cuticle. The wax deposition processes in the two apple cultivars were similar.
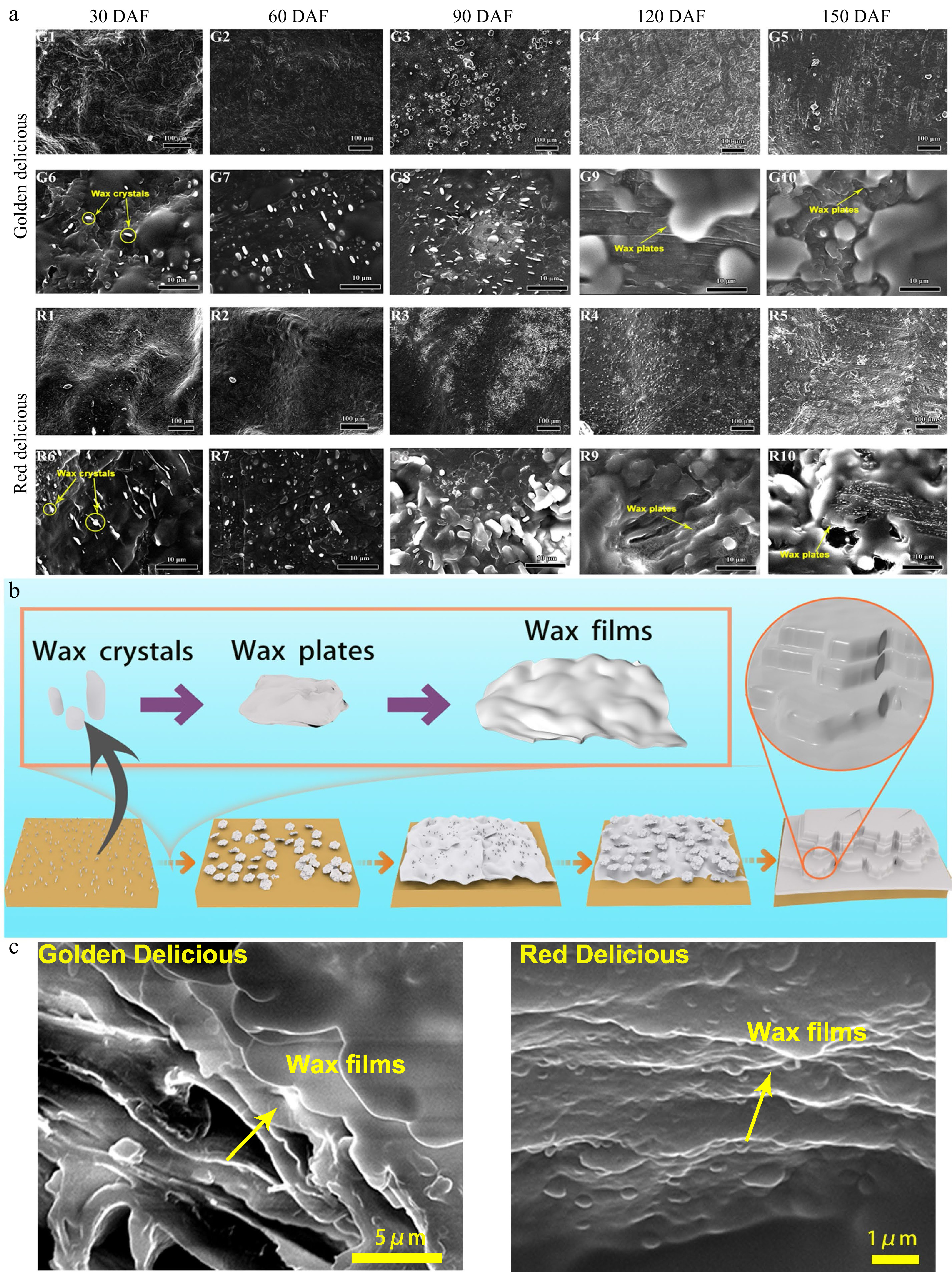
Figure 3. Changes of wax crystal in the epidermis of apple fruit during growth and development. (a) Environmental field scanning electron micrographs of the apple fruit surface. G1–G5 and R1–R5: Low magnification scanning electron microscopic observations (×150) of 'Golden Delicious' and 'Red Delicious' growing at 30, 60, 90, 120, and 150 DAF; G6–G10 and R6–R10: High magnification scanning electron microscopic observations (×2,500) of 'Golden Delicious' and 'Red Delicious' growing at 30, 60, 90, 120, and 150 DAF. (b) Diagram of wax model of apple epidermis at different growth and development stages. (c) Wax lamellar structures of 'Golden Delicious' and 'Red Delicious' apple fruit at 150 DAF.
At the same time, we observed the crystal structure of the epidermal wax in the two apple cultivars by high power microscopy (×2,500). As shown in Fig. 3a (G6-G10 and R6-R10), wax plates accumulated and their structures changed during fruit development of both cultivars. A single wax crystal was gradually deposited into a wax plate, and the wax morphology changed. Both apple cultivars exhibited irregular epidermal wax crystals beginning 30 DAF (Fig. 3a, G6 and R6). A large number of small plates appeared on the surface of 'Red Delicious' at 90 DAF. They were in an amorphous state and loosely arranged (Fig. 3a, R8). From 120 to 150 DAF, the wax plates became overlapped and fused into wax films in both 'Golden Delicious' and 'Red Delicious' (Fig. 3a, G9-G10 and R9-R10). However, at 150 DAF, 'Red Delicious' was observed to have many small wax grains on large wax plates (Fig. 3a, R10) while 'Golden Delicious' was not (Fig. 3a, G10).
In order to further intuitively describe the formation process of apple epidermal wax in different development stages, we illustrated the formation sketches of different stages of apple epidermal wax (Fig. 3b). During the formation of wax in the apple epidermis, the first observed wax crystals are granular wax crystals. With apple fruit growth and development, wax particles gradually become larger, and form wax plates. Then, adjacent plates contact each other to form a larger plate, here named wax film, which covers the fruit cuticle. The thickness and layer numbers of wax film vary with cultivars. We photographed the epidermis of two apple cultivars at the end development stage (150 DAF), and observed the lamellar structure of their edge positions (Fig. 3c). Although the epidermal wax morphology of the two apple cultivars is different, the lamellar structure observed is similar.
Changes in the wax chemical components of the cuticle during development
-
To determine the composition and synthesis of epidermal wax during fruit development, we extracted epidermal wax from apple fruits at each development stage for analysis. Total wax content increased gradually from 30 to 150 DAF, and wax synthesis of both apple cultivars was quicker from 60 to 90 DAF (Fig. 4a & b). Total wax content of 'Red Delicious' increased 2,753.4 mg m−2 (Fig. 4b), and that of 'Golden Delicious' increased 2,250.6 mg m−2 from 60 to 90 DAF (Fig. 4a). A significant increase of wax content (2,353.1 mg m−2) in 'Golden Delicious' was observed from 120 to 150 DAF (Fig. 4a).
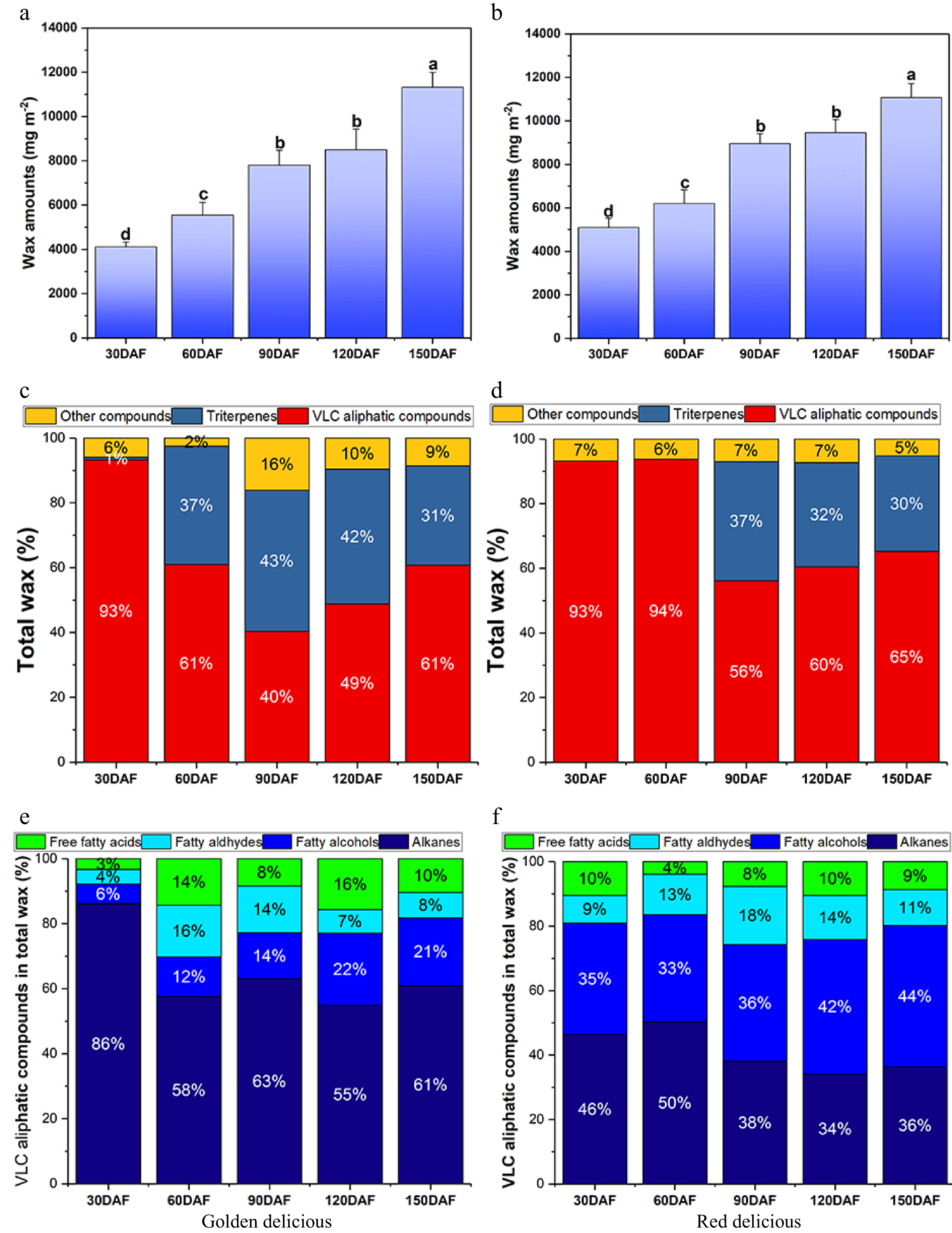
Figure 4. Wax content and chemical composition of the two apple cultivars at different developmental stages. (a), (b) Represent the total wax content per unit area of 'Golden Delicious' and 'Red Delicious' fruits, respectively. Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan's new multiple range test. Each value is the mean ± standard error. (c), (d) Represent the percentage of different components in total wax of 'Golden Delicious' and 'Red Delicious' fruits. (e), (f) Represent the percentage of VLCFA derivatives of 'Golden Delicious' and 'Red Delicious' fruits.
Subsequently, we analyzed the wax chemical components extracted from the two cultivars by GC-MS. A total of 44 compounds were identified, including 37 VLCFA derivatives and six triterpenoids. Then PCA analysis was performed on the wax compounds in the two cultivars at different developmental stages (Supplemental Fig. S1). On the score plot, PC1 (39.8%) and PC2 (18.4%) explained 58.2% of the variation (Supplemental Fig. S1a). At the 30 DAF stages, the difference of wax components between the two varieties was small, but gradually separated after 90 DAF, indicating that the difference of wax components between the two varieties was relatively large (Supplemental Fig. S1a). The main components of total wax included very long chain fatty acids (VLCFAs), triterpenes, and other compounds. The percentage of different wax components varied at different developmental stages (Fig. 4c & d). In the 30 DAF stages, VLCFA contents were higher in both apple cultivars. During subsequent development (60 to 90 DAF), the proportion of VLCFA compounds decreased gradually. While after 90 DAF, the proportion of VLCFA compounds gradually increased in both cultivars (Fig. 4c & d). They were almost undetectable in 30 DAF 'Golden Delicious', but increased significantly after 30 DAF (Fig. 4c). The triterpenes were undetectable in 'Red Delicious' before 60 DAF (Fig. 4d).
VLCFAs and their derivatives are the main components of cuticular wax. Next, we analyzed the VLCFA derivatives in the wax layer of the epidermis during development of 'Red Delicious' and 'Golden Delicious' fruits, and found that they are mainly composed of alkanes, fatty alcohols, fatty aldehydes and free fatty acids (Supplemental Table S2, Fig. 4e & f). Saturated alkanes (C16–C31) were the main components of cuticular wax in 'Golden Delicious' apples. In particular, the content of alkanes in the VLCFAs was 3,259.34 mg m-2 during the early developmental stage (30 DAF) (Supplemental Table S2), accounting for 86% of all VLCFA contents (Fig. 4f). During the subsequent growth and development, the alkane content initially decreased and then increased, and finally reached 4,257.56 mg m−2 (Fig. 4f, Supplemental Table S2). However, compared with 'Golden Delicious' fruit, the percentage content of alkanes, fatty alcohols, aliphatic aldehydes and free fatty acids in 'Red Delicious' fruit remained at a relatively stable level during development (Fig. 4e). Notably, alkanes and fatty alcohols, two main VLCFA derivatives in the epidermal wax of 'Red Delicious' fruit, showed a tendency to decrease at first and then increase (Supplemental Table S2).
To identify the main VLCFA derivatives of the epidermal wax, further analysis was performed on the composition of alkanes, fatty alcohols, fatty aldehydes and free fatty acids. As shown in Fig. 5a & e, the main components of saturated alkanes were C29 compounds, followed by C27 in both apple cultivars. More fatty alcohols were detected in the cuticular wax of 'Red Delicious' compared with 'Golden Delicious' apples at the developmental period (Supplemental Table S2). In 'Red Delicious', the highest content of long chain fatty alcohols was 10-nonacosanol (10-C29-ol) (Fig. 5f). The change in its content at the developmental period was similar to that of C29 saturated alkanes. In 'Golden Delicious', the content of 10-C29-ol was lower than that of 'Red Delicious' varieties (Fig. 5b). The changes of fatty aldehydes in epidermal waxes of the two cultivars were similar, and the substance with the highest content was triacontanal (C30) (Fig. 5c & g). The free fatty acids in the wax of 'Red Delicious' mainly consisted of palmitic acid (C16) and stearic acid (C18) at the 30 DAF stage (Fig. 5h), and then decreased gradually. The long-chain fatty acids (≥ C24), such as C30 and C28, increased significantly from 90 to 120 DAF (Fig. 5h). The content of C16 and C18 fatty acids in 'Golden Delicious' was the highest at 60 DAF (Fig. 5d), and then decreased, while the change of long-chain fatty acids (≥ C24) content was similar to that of 'Red Delicious' apple.
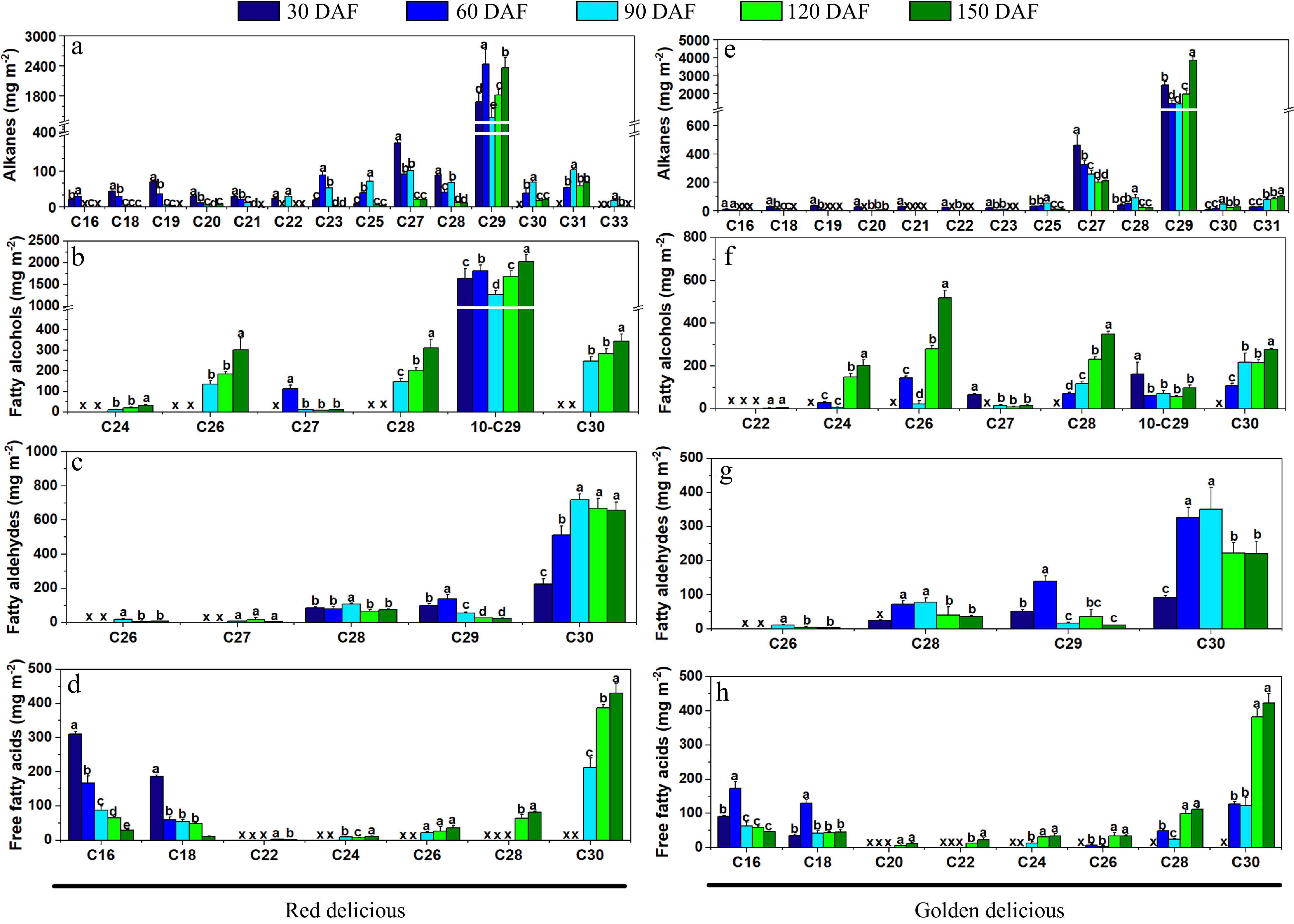
Figure 5. Concentrations of the cuticular wax composition of 'Golden Delicious' (a−d) and 'Red Delicious' (e−h) apples at growth and development. (a), (e) Alkanes. (b), (f) Fatty alcohols. (c), (g) Fatty aldehydes. (d), (h) Free fatty acids. Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan's new multiple range test, x indicates that the composition is not detected. Each value is the mean ± standard error.
Quantitative expression of wax synthesis-related genes at different developmental stages
-
To understand the transcription patterns of wax-related genes during apple growth and development, qRT-PCR was performed to examine the expression levels of 12 genes associated with wax synthesis in the peel and flesh of the two apple cultivars at different stages (Fig. 6). MdCER1, MdCER2 and MdCER6 encode the biosynthesis of long-chain alkanes in epidermal wax of apple fruits, while MdCER4 encode the synthesis of primary alcohols in epidermal wax. The expression of these genes in peel tissue was much higher than that in flesh tissue in both cultivars (Fig. 6a−d). The expression of MdCER4 gene in 'Red Delicious' apple peel was much higher than that of 'Golden Delicious' during all development stages (Fig. 6c). MdLACS2 and MdLACS4, which encode VLCFA derivatives, act as positive regulators in wax accumulation. They showed different expression patterns in the growth and development stages of the two varieties (Fig. 6e & f). MdKCS1 and MdKCS7/2 encode rate-limiting enzymes for de-novo synthesis of fatty acids. The expression of MdKCS1 in peel was significantly higher than that in flesh (Fig. 6g). The expression of MdKCS7/2 was similar in the two cultivars, which is higher in peel than that in flesh (Fig. 6h). The MdLTPG1 (lipid transfer protein) gene showed a higher transcription level in fruit peel of both cultivars, compared to that in flesh (Fig. 6i).
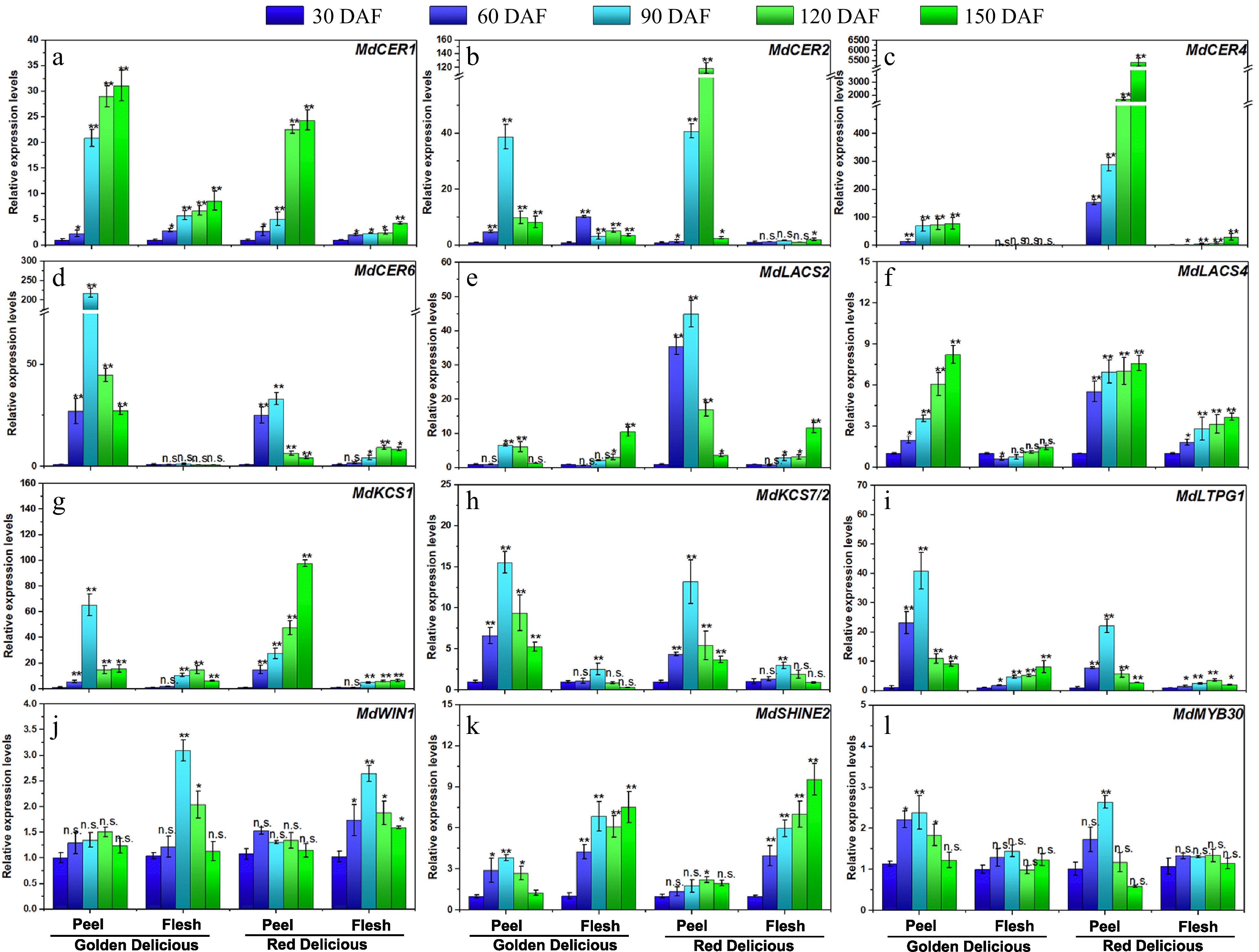
Figure 6. Expression analysis of wax-related genes at growth and development of 'Golden Delicious' and 'Red Delicious'. The asterisks indicate a statistically significant difference (two-tailed Student's t-test, * p ≤ 0.05, ** p ≤ 0.01). n.s. represent no significant difference (p > 0.05). The columns and bars represent the mean and standard error, respectively (n = 3).
In addition, we examined the transcription levels of several wax-related transcription factors. The expression of MdWIN1 was not significantly changed in apple peel, but differed in flesh during fruit development, which reached the highest level at 90 DAF (Fig. 6j). The expression pattern of MdSHINE2 in flesh is higher than that in peel during fruit development in both cultivars (Fig. 6k). The expression of the MdMYB30 gene reached the highest level in peel of the two cultivars at 90 DAF, but was not significantly changed in the flesh tissues (Fig. 6l).
-
Apple cuticular gloss is one of the most important appearance qualities, however, few studies report on fruit gloss. Cuticular wax directly determines fruit gloss[5,21]. In this study, we used a gloss meter to evaluate the skin gloss of apple fruits during growth and development. We found that the gloss value of both cultivars was lowest at 90 DAF, and increased gradually during subsequent development (Fig. 2b & c). At the same time, correlation analysis showed that Angle 20° (Supplemental Fig. S3a & S3d), Angle 60° (Supplemental Fig. S3b & S3e) and Angle 85° (Supplemental Fig. S3c & S3f) had significant correlation (R2 > 0.95), moderate correlation (0.5 < R2 < 0.8), and low correlation (0.3 < R2 < 0.5) with cuticular wax amounts, respectively. These results suggest that 90 DAF is the critical point of apple epidermal wax synthesis.
The wax in the outer layer of the apple peel has a typical crystal structure, which is called wax crystal. Previous studies have suggested that wax in apple peel generally has a plate structure, and that there are notable differences among cultivars[14]. Curry used laser confocal SEM to observe the thickness of apple epidermis wax, and reported that it ranged from 2 to 4 μm[22]. Yang et al. discovered that some apples are prone to become greasy during storage, wax crystals are reduced, and crystal fusion occurs[21]. However, changes in the structure of the wax during apple growth and development have been rarely studied. Here, scanning electron microscopy showed that epidermal wax accumulated gradually during growth and development in both cultivars (Fig. 3a). This is also similar to the results of gloss changes during development (Fig. 2a).
Three-dimensional wax crystals are produced on plant surfaces by self-assembly of wax compounds and affect light reflection at the cuticular interface[5]. Changes in wax composition may lead to changes in wax morphology and appearance of the apple. However, there are few studies on the morphological characteristics and formation process of apple epidermal wax. In our study, although the epidermal wax of both cultivars exhibited different microstructures, their synthetic patterns were similar at the same growth and development period (Fig. 3b). These results are also similar to the pattern of wax deposition in Citrus sinensis fruits during growth and development[25].
Changes of wax composition during fruit growth and development
-
The content of fatty alcohols in the cuticular wax of 'Golden Delicious' apples was very low, while the proportion of long-chain alkanes was almost the same compared with 'Red Delicious' apples[14]. The difference in these components resulted in the change of the morphology of epidermal wax (Fig. 3a). Nonacosane (C29) and 10-Nonacosanol (10-C29-ol) are the main components of epidermal wax along with VLCFA compounds, which play an important role maintaining the 3D structure of the apple epidermal wax[21]. These are similar results to those of the present study. Aldehydes contribute to water retention during post-harvest storage[9]. Similarly, the types of fatty aldehydes vary with species. Fatty aldehydes in blueberries are dominated by C30 and C28 compounds, which is similar to apples[8,26]. C24 and C28 are the main aldehydes in Newhall navel orange wax[25,27]. In our results, the long-chain fatty aldehydes in apple epidermis wax were mainly C30, and their content was highest at 90 DAF (Fig. 5c & g). During this period, the content of saturated alkanes and fatty alcohols per unit area decreased (Fig. 5a & e). We suspect that this may be due to the rapid expansion of apple surface area during this period (Fig. 1f), resulting in a decrease in the total amount of epidermal wax per unit area. The increase in fatty aldehydes and free fatty acids made up for the decrease in wax coverage.
Triterpenoids belong to the inner layer of wax in epidermal wax. In the latest research, 42 triterpenoids were isolated by high resolution liquid chromatography, and almost all of them were wax components[28]. Triterpenoids are also abundant in the wax of apple epidermis, accounting for 30%−40 % of the total wax in apple peel[14]. This is similar to our results. In 'Red Delicious', triterpenoids were synthesized in large quantities before and after 90 DAF, while they were synthesized by 60 DAF in 'Golden Delicious' (Fig. 4c & d, Supplemental Table S2). Six triterpenoids were identified in the two cultivars, including β-Sitosterol, γ-Sitosterol, α-Amyrin, Ursolic acid (most abundant), Ursolic aldehyde and Lupeol (second most) (Supplemental Fig. S2). Triterpenoids have numerous biological activities, including participating in fruit disease resistance. For example, the triterpenoids in the epidermal wax of Asian pear fruit (cv. Pingguoli) are responsible for inhibiting the germination and growth of Alternaria alternate[29]. Triterpenoids are located in the inner layer of the epidermis and do not form wax crystals; therefore, we did not analyze the triterpenoids in detail in the current research.
Changes in expression of wax synthesis-related genes at different developmental stages
-
Wax-related genes have been extensively studied in model plants, especially those involved in regulating alkane synthesis. The CER (Eceriferum) gene family has been previously identified to control the biosynthesis of plant wax. AtCER1[30] and AtCER2[31] in Arabidopsis act as an aldehyde decarbonylase to complete wax synthesis by catalyzing the conversion of long-chain aliphatic aldehydes to alkanes. In apple, MdCER1 and MdCER2 protein is highly homologous to Arabidopsis AtCER1 and AtCER2, and over-expression of MdCER1[32] and MdCER2[33] can increase wax accumulation. AtCER4 has been identified as an aliphatic acyl-CoA reductase that forms fatty acyl-CoA reductase, which is specific to long-chain fatty acids and is responsible for primary alcohol synthesis of waxes[34]. In Arabidopsis, AtCER6 is the key wax gene that has been studied, and it is necessary for extending C24 ultra-long chain fatty acids. The gene encodes ketolipoacyl-CoA synthase, which acts on long chain fatty acids[35]. In this study, the expression of the MdCER4 gene was very active during the growth and development stage of 'Red Delicious' peel tissues (Fig. 6c), and a large number of fatty alcohols were detected at different growth and development stages (Fig 5b). Long chain acyl-CoA synthase (LACS) is a member of the acyl-activated enzyme superfamily and plays an important role in wax synthesis[36]. In apple, MdLACS2 and MdLACS4 genes has been identified, they are highly expressed in the pericarp tissue, and can also significantly promote the accumulation of epidermal wax[37,38]. The β-ketoester-CoA (KCS) is a rate-limiting enzyme in wax anabolism. In apple, KCS family genes are highly expressed in the peel tissues[39], which is similar to our results (Fig. 6g & h). In Arabidopsis, LTPG1 is a glycosylphosphatidylinositol-anchored lipid transfer protein, and its transcription is regulated by AtLTPG1[40]. The function of MdLTPG1 may be related to lipid exports[41]. MdLTPG1 transcription is similar to other wax-related genes in the pericarp tissue, and the gene expression level was highest at 90 DAF, followed by a significant downward trend (Fig. 6i).
WIN1 (WAXY INDUCTION/SHINE1), is a transcription factor containing the AP2 domain, regulates AtLACS2 and AtLCR to participate in Arabidopsis epidermal wax biosynthesis[42]. MdSHINE2 is highly homologous to AtSHINE2 in Arabidopsis thaliana and its function has been identified in apple. MdSHINE2 positively regulates epidermal wax biosynthesis[43]. In our results, the expression of MdWIN1 and MdSHINE2 in flesh tissue was higher than that in skin tissue, indicating its specific function in flesh tissue (Fig. 6j & k). MdMYB30 is a transcription factor that regulates cuticular wax biosynthesis at the transcriptional level and plays an important role in wax biosynthesis and disease resistance in apples[44]. It was found that its expression reached the highest level at 90 DAF stage in both cultivars, while its expression was not significant in flesh tissue (Fig. 6l).
Based on our results, most wax-related structural genes exhibited higher transcription levels in apple peel, compared to flesh, during fruit development. However, the expression of transcription factors MdWIN1 and MdSHINE2 is higher in flesh. Meanwhile, the expression of structural genes in different fruit development stages changed much more than that of transcription factors, indicating the direct function of structural genes in mediating wax biosynthesis. Finally, it was found that the transcript levels of eight of the 12 genes studied reached a peak at 90 DAF, suggesting that 90 DAF might be a crucial period for wax accumulation.
-
In the current study, wax biosynthesis was investigated during apple growth and development. According to the SEM, skin gloss value, and GC-MS results, 90 DAF was the critical period for wax biosynthesis in apple fruit. The VLCFA compounds that determine apple cuticular wax were mainly long-chain fatty alkanes. The wax composition of 'Red Delicious' apples also contained a large amount of secondary alcohols, which decreased initially and tended to increase later. The qRT-PCR results showed that the expression levels of wax synthesis-related genes increased significantly at 90 DAF, also demonstrating that 90 DAF was a key period for epidermal wax synthesis. In general, wax was deposited gradually on the surface of the apple peel during fruit development, which might play an important role in water retention, disease resistance, and external quality of apple fruits.
-
The 'Red Delicious' and 'Golden Delicious' apple (Malus domestica Borkh.) cultivars were picked from an apple orchard in Tai'an, Shandong, China. Both cultivars were picked 30, 60, 90, 120, and 150 days after flowering (DAF). The standards for picking the samples were: 1) apple skin has no mechanical damage, 2) no physiological diseases, and 3) fruit size and appearance are uniform. The apples were picked into polyvinyl chloride bags after harvest.
Determination of fruit quality
-
The calculation formula of fruit surface area was based on the method of Yin et al.[29] with slight modification. We used vernier calipers to measure and record the transverse diameter (l), longitudinal diameter (d) and height (h) of the apple fruit. Fruit surface area was calculated using the formula: A (mm2) = 4 × π × (l + d + h)2/36.
Pericarp tissue of the same thickness was peeled and an 8 mm round punch was used to cut the pericarp tissue. Ten pieces of tissue were taken as a group, and the thickness of the peel was consistent and damage was minimized. This process was repeated 10 times for each period (i.e., DAF) of fruit sampled. The pericarp tissue was vacuumed in deionized water. S1 conductivity was measured after the pericarp sank to the bottom. Afterwards, plant tissue samples were boiled for 20 minutes. S2 conductivity was measured after cooling the tissue to room temperature (± 25 °C). Relative conductivity (S) was calculated as S = S1/S2 × 100%.
The fruit firmness was measured by fruit texture analyzer (Stable Micro Systems, Godalming, UK). A P/2 columnar probe (diameter: 2 mm) was used to measure fruit firmness. The pretest velocity was 2 mm s−1, measurement velocity was 1 mm s−1, and post-test velocity was 5 mm s−1. The data were automatically analyzed and calculated by Texture Exponent 32 software. Five fruits were selected as one repeat at each stage, and three replicates were determined.
Apples were randomly selected from each group, and total soluble solids (TSS) and titratable acids (TA) were determined from each apple. TSS value of fruit was measured using a digital hand-held refractometer (Atago PAL-1, Japan). TA value was measured by acid-based titration.
Fruit glossiness and surface color measurement
-
Apple fruit epidermis gloss is a physical quantity. In order to evaluate the ability of fruit epidermis to reflect light at different growth and development stages, a Micro-TRI-gloss tester (BYK-4563, Germany) was used to measure fruit surface gloss. The measuring mouth of the instrument was aligned with the equatorial plane of the fruit for testing, and 10 points were randomly selected for each apple for testing. The color of the apple surface was determined by a chroma meter CR-400 (Konica Minolta Sensing Inc., Japan). Lightness (L*), Redness (a*) and Yellowness (b*) values were recorded on the equatorial surface of the fruit.
Electron microscopic observations
-
The microstructures of cuticular wax in apple were observed by scanning electron microscopy (SEM) using the method of Yang et al.[45] with sight modification. Five apples at each growth and development stage were randomly selected and apple pericarp tissue (about 5 mm in diameter) were cut from the equatorial zone of the fruit with an anatomical blade. The disks were then frozen using liquid nitrogen, and then transferred to FDU-1110 vacuum freeze dryer (EYELA, Tokyo, Japan) for 24 h for tissue dehydration. The freeze-dried materials were coated with platinum target JFC-1600 ion sputtering apparatus (JEOL, Tokyo, Japan). We used JSM-6610 SEM (JEOL, Tokyo, Japan) to observe the epidermal wax structures.
Cuticular wax extraction and analysis
-
The apple cuticular wax was extracted using the chloroform extraction method as per Yang et al.[45]. Apple fruits were soaked and eluted with 200 ml chloroform for 45 s. The chloroform was transferred to a rotary evaporator for concentration. Subsequently, the samples were blow-dried under N2 flow. The wax samples were stored at −80 °C. The experiment was repeated three times.
The sample was re-dissolved with 18 ml chloroform and 2 ml methanol, and 1 ml (1 mg ml−1) n-tetracosane (Sigma Aldrich, China) was added as internal standard. One ml wax solution was added into a centrifuge tube and dried under N2 flow, 300 μl bis-N,O-(trimethylsilyl)trifluoroacetamide (BSTFA, Sigma Aldrich, China) was added for derivatization (70 °C, 40 min). After derivatization, the samples were injected into the GC-MS (QP-2010 Plus, Shimadzu, Tokyo, Japan). Chemical composition of cuticular wax was determined by gas chromatography-mass spectrometry (GC-MS) as described by Yang et al.[21]. NIST 17 MS Library was used for comparative analysis of the detected compounds. Internal standard method was used to calculate the content of wax chemicals detected. Triterpenoids were quantified by external standard method based on calibration curve[9,46].
RNA isolation and quantitative real-time polymerase chain reaction (qRT-PCR)
-
RNA was extracted from pericarp and flesh tissues using an RNA extraction kit (Tiangen, Beijing, China). Subsequently, cDNA samples were prepared using an RNA reverse transcription kit (TaKaRa, Dalian, China). The sample preparation procedure for qRT-PCR was according to the method of An et al.[47]. The results were normalized to those of Md18S. Three biological replicates were performed for each sample. qRT-PCR reaction conditions were as follows: denaturation at 95 °C for 10 min; 25−28 cycles, 95 °C 30 s, 58 °C 30 s, 72 °C 30 s; 72 °C, 5 min. Specific primers are shown in Supplemental Table S1.
Statistical analysis
-
All statistical analyses were carried out using the software Origin Pro 2017. Data were analyzed using the two-tailed student t-test and Duncan's new multiple range test. 3D MAX 2016 software was used to construct the model of epidermal wax at different developmental stages.
- This work was supported by grants from the National Key Research and Development Program (2018YFD1000200) and the National Natural Science Foundation of China (32072539 and 31772275).
- The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 PCA of wax compounds in ‘Golden Delicious’ and ‘Red Delicious’ apple fruit at different growth and development stages. (a) Scatter plot of PCA scores and (b) Loadings plot of the PCA.
- Supplemental Fig. S2 Concentrations of the cuticular triterpenoids composition of ‘Golden Delicious (a) and ‘Red Delicious (b) apples at growth and development. Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan’s new multiple range test, x indicates that the composition is not detected. Each value is the mean ± standard error.
- Supplemental Fig. S3 Correlation analysis between wax amounts and fruit cuticular gloss during growth and development of two apple cultivars. Data were analyzed using Pearson's correlation coefficients. (a-c) represent ‘Golden Delicious’ Angle 20°, Angle 60° and Angle 85°, respectively. (d-f) represent ‘Red Delicious’ Angle 20°, Angle 60° and Angle 85°, respectively.
- Supplemental Table S1 Primer pairs used for the qRT-PCR analysis.
- Supplemental Table S2 Concentrations of wax fractions of ‘Golden Delicious’ and ‘Red Delicious’ apples during Developmental period. Different letters in the same fraction indicate significant differences at p ≤ 0.05 according to Duncan’s new multiple range test. n.d. represent not detected. Each value is the mean ± SE. The concentrations were determined by semi-quantification.
- Copyright: © 2022 by the author(s). Exclusive Licensee Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Y, Wang X, Cao Y, Zhong M, Zhang J, et al. 2022. Chemical composition and morphology of apple cuticular wax during fruit growth and development. Fruit Research 2: 5 doi: 10.48130/FruRes-2022-0005

Chemical composition and morphology of apple cuticular wax during fruit growth and development
- Received: 03 November 2021
- Accepted: 16 March 2022
- Published online: 13 April 2022
Abstract: Cuticular wax covers the surface of the apple epidermis and plays an important role in water loss and pathogen resistance. However, little is known about wax biosynthesis during apple fruit development. In this study, the morphological structure and chemical composition of wax and the transcription patterns of wax-related genes were investigated in the 'Red Delicious' and 'Golden Delicious' cultivars. The results showed that accumulation of the waxy layer was synchronized with enlargement and maturation of fruit. The crystal structure, major chemical components (very long chain fatty acids and triterpenes) of wax and the expression levels of wax-related genes varied significantly 90 days after flowering (DAF), indicating that the critical period for biosynthesis of cuticular wax was about 90 DAF. This finding is valuable for further study of the regulatory mechanism of wax biosynthesis at both the molecular and physiological levels.
-
Key words:
- Apple /
- Cuticular wax /
- Wax crystal structure /
- Wax component /
- Gene expression