









-
Abiotic stresses widely affect plant growth and development[1]. Salt stress is one of major abiotic stresses which affects plant cellular and physiological changes, resulting in ion imbalances and toxicity[2], causing osmotic stress, reducing nutrient balance, stomatal activity, hydraulic conductivity, and ability of photosynthesis[3]. Thus, to survive, plants must respond in a timely manner to improve their adaptation and tolerance to salt stress. However, the process of adaptation is complex, with changes in gene expression, proteins, and metabolites, etc[4, 5]. In addition, the hormones maintain a balance between normal growth and resistance of plants under salt stress by interacting with each other[6], and changes of plant hormone-related signaling and regulatory pathways affect plant adaptation to salt stress[7], which involves significant changes in specific transcription factors and expression of related genes[8]. Therefore, hormones play an important role in response to salt stress.
Melatonin (MT) is an important class of indole-like compound that are widely existed in animals[9]. In higher plants, the content of it is in minute amounts but varies in different tissue organs[10]. Recently, the function of exogenous application of MT has been widely studied in tree plants, including regulation of flowering, fruit quality, stress resistance, etc. Among them, the most widely studied is that MT plays a critical role in response to both biotic and abiotic stresses[11−13]. For example, exogenous treatment of MT could reduce drought-induced growth inhibition and the production of superoxide (O2–) and hydrogen peroxide (H2O2) in apple rootstocks with alleviating damage to photosynthesis by protecting the integrity of membranes[14]. In addition, application of exogenous MT could significantly increase the accumulation of antioxidant substances which enlarge the antioxidant capacity of mitigating the oxidative damage caused by salt stress[15]. In pear, MT could increase the maximum photosynthetic rate in the later stage of fruit development by increasing the content of soluble sugar[16]. In jujube, during the process of cold storage, application of exogenous MT could delay the degradation of jujube fruit nutrients by inhibiting the related enzyme activity and gene expression, and improve the antioxidant capacity to maintain the quality of postharvest jujube fruits[17, 18].
Jujube (Ziziphus jujuba Mill.) is native to China, belonging to the Ziziphus (Ziziphus Mill.) genus in the family of Rhamnaceae, and has important economic and ecological values[19, 20]. Sour jujube (Ziziphus jujuba var. spinose), as a rootstock of jujube, is also one of the dominant fruit trees in China[21]. The fruit of sour jujube is specifically rich in vitamin C, flavonoids, triterpenoids, and has special nutritional composition and medicinal values. In addition, the seedlings of sour jujube are highly resistant to drought and salinity stress. However, in response to salt stress, the growth of seedlings and seed germination could be negatively affected[22]. Thus, improving the ability of sour jujube to salt stress is still necessary and whether the exogenous application of MT could regulate the salt resistance ability of jujube tree has not been reported.
In the current study, sour jujube seedlings were used as plant materials with different concentration treatments of sodium chloride (NaCl) and MT, water treatment was used as control, phenotypic observation, and transcriptome analysis were performed to determine the potential molecular mechanisms behind them. The results could provide the theoretical reference for application of MT to regulate salt stress in sour jujube.
-
Sour jujube seeds were sown in nutritional bowls and grown in a laboratory cultivating room with a 12 h light/12 h dark photoperiod and constant temperature (23 °C) and humidity (70%). When the seedling grew seven to nine leaves, the healthy and consistent growth seedlings were selected and divided into five treatment groups: water treatment (W), 200 mmol/L, and 400 mmol/L NaCl treatment (Na), 10 µmol/L MT treatment for 6 d (root application once every two days, three times in total) prior to 200 mmol/L NaCl treatment, 10 µmol/L MT treatment for 6 d (root application once every 2 d, three times in total) prior to 400 mmol/L NaCl treatment (MNa), three biological replicates for each treatment were performed. The phenotypic observation was conducted on day 1, 3, and 8. Meanwhile, at the corresponding time, the leave samples were collected and stored at −80°C for transcriptome sequencing analysis.
RNA extraction, library preparation and sequencing
-
Total RNA was extracted from the leaves of sour jujube which were selected from water treatment (W), 400 mmol/L NaCl treatment (Na), and 10 µmol/L MT plus 400 mmol/L NaCl treatment (MNa) groups at 0 and 192 h by using RIzol® Reagent kit. After checking of RNA concentration and quality, 1 μg of total RNA was used to construct the RNA-seq transcriptome library with TruSeqTM RNA sample preparation Kit and RNA-seq transcriptome library was sequenced by Illumina HiSeq xten/NovaSeq 6,000 sequencer (2 × 150 bp read length) at the Shanghai Majorbio Bio-pharm Biotechnology Co. (Shanghai, China)[23].
Transcriptome data and gene annotation analysis
-
The raw data obtained by sequencing were filtered to obtain high-quality sequences. Firstly, the raw paired end reads were trimmed and quality controlled by SeqPrep (
https://github.com/jstjohn/SeqPrep ) and Sickle (https://github.com/najoshi/sickle ) with default parameters. After filter out adapters and low-quality sequences, the clean reads were separately mapped using TopHat (http://tophat.cbcb.umd.edu/ , version 2.0.0)[24]. Then using RSEM (http://deweylab.biostat.wisc.edu/rsem/ )[25] to calculate gene expression levels as transcripts per million reads (TPM). Majorbio cloud platform (www.majorbio.com ) was used to summarize and describe gene function annotation.Differential expression genes and functional enrichment analysis
-
Using DESeq2[26] software to analyze the differential expression genes (DEGs) between groups of transcriptome data of different treatments in sour jujube seedlings, and the screening threshold was set as |log2 FC| ≥ 1 and p-adjust values < 0.05. The software Goatools (
https://github.com/tanghaibao/GOatools )[27] and KOBAS (http://kobas.cbi.pku.edu.cn/home.do ) was further used for Gene Ontology (GO) and Kyoto Encyclopedia of Genes and Genomes (KEGG) enrichment analysis, respectively. In addition, the STRING database (http://string-db.org/ ) was used for protein interactions analysis and their orthologs in Arabidopsis were used as references. The figures were drawn using cytoscape software[28].Identification of transcription factor analysis
-
The transcription factor database PlantTFDB (Plant,
http://planttfdb_v4.cbi.pku.edu.cn/ )[29, 30] was used to predict transcription factors. According to the prediction results, the transcription factor families were counted and stress-related transcription factors were screened. -
To gain insight to whether MT treatment could rescue the damage caused by salt stress, different concentration of NaCl and combination of NaCl and MT were applied to sour jujube. Compared to the control, 200 mmol/L NaCl treatment could not induce the damage within 8 d. However, under 400 mmol/L NaCl treatment, the leaves of sour jujube seedlings appeared yellowing, curling, and gradually wilting within 8 d compared to the control (Fig. 1a & b). While the symptoms of sour jujube under the MT + NaCl group were also yellowing and curling, but not as severe as with only NaCl treatment (Fig. 1b & c, Supplemental Fig. S1). The phenotypical results indicated that 400 mmol/L NaCl treatment could induce leaf damage of sour jujube, while 10 µmol/L MT treatment had some alleviating effect on NaCl induced leaf damage. Therefore, for the next transcriptome analysis, the 400 mmol/L NaCl and MT + NaCl treatments were selected subsequently.

Figure 1.
Time course of phenotypic characteristics of sour jujube seedlings under (a) control group, (b) 400 mmol/L NaCl treatment group, and (c) 10 µmol/L MT + 400 mmol/L NaCl treatment group respectively, within 0 h, 24 h, and 192 h from left to right. Bar: 1 cm.
Transcriptome sequencing data and quality assessment
-
In order to initially reveal the molecular mechanism of how exogenous MT treatment alleviates the salt stress induced damage in sour jujube seedlings, transcriptome analysis was performed and the sequencing data were obtained (Table 1). Among them, the raw reads obtained from each treatment were higher than 44.5 million reads, while the clean reads obtained after filtering and quantization, were higher than 44.1 million reads. In addition, error rates ranged from 0.0243% to 0.0251%, GC content was from 44.43% to 44.97%, and base scoring Q30 was above 93.96 %. The comparison efficiency of clean reads matching with the jujube reference genome for each sample ranged from 83.08% to 89.08%, among which 11.66% to 12.32% were matched to multiple positions and 71.00% to 77.35% were matched to the unique position, indicating that the data obtained by transcriptome sequencing were of high quality and could be used for subsequent analysis.
Table 1. Statistical analysis of transcriptome sequencing data.
Samples Raw reads Clean reads Error rate (%) GC content (%) Q30 (%) Total mapped Multiple mapped Uniquely mapped W1 45076920 44509554 0.0250 44.54 94.13 39226588 (88.13%) 5230113 (11.75%) 33996475 (76.38%) W2 45847360 45407490 0.0251 44.65 93.96 40451096 (89.08%) 5338687 (11.76%) 35112409 (77.33%) W3 44540110 44091906 0.0250 44.43 94.09 39056270 (88.58%) 5141396 (11.66%) 33914874 (76.92%) Na1 55405174 54910544 0.0247 44.83 94.34 48658620 (88.61%) 6669193 (12.15%) 41989427 (76.47%) Na2 51733218 51190904 0.0247 44.65 94.32 45581570 (89.04%) 6270880 (12.25%) 39310690 (76.79%) Na3 48546886 48083922 0.0243 44.92 94.73 41217079 (85.72%) 5925798 (12.32%) 35291281 (73.40%) MNa1 53456026 52992586 0.0247 44.97 94.40 44028697 (83.08%) 6403570 (12.08%) 37625127 (71.00%) MNa2 47546416 47139186 0.0247 44.47 94.40 41628008 (88.31%) 5603741 (11.89%) 36024267 (76.42%) MNa3 55938524 55430944 0.0249 44.50 94.12 49350021 (89.03%) 6474285 (11.68%) 42875736 (77.35%) Correlation analysis between the samples
-
To confirm the relationship of all samples, the correlation and principal component analysis (PCA) analysis were performed. As shown in Fig. 2, the R2 values among the three replicates of the same treatment were all larger than 0.856 and most of them were above 0.913, while the correlation among the three treatment groups were significantly different. For the PCA analysis, three replicates of the same treatment clustered together, indicating high reliability among the samples. In addition, the three treatment groups scattered and distributed separately in different areas implied weak correlation and more DEGs existed among them.
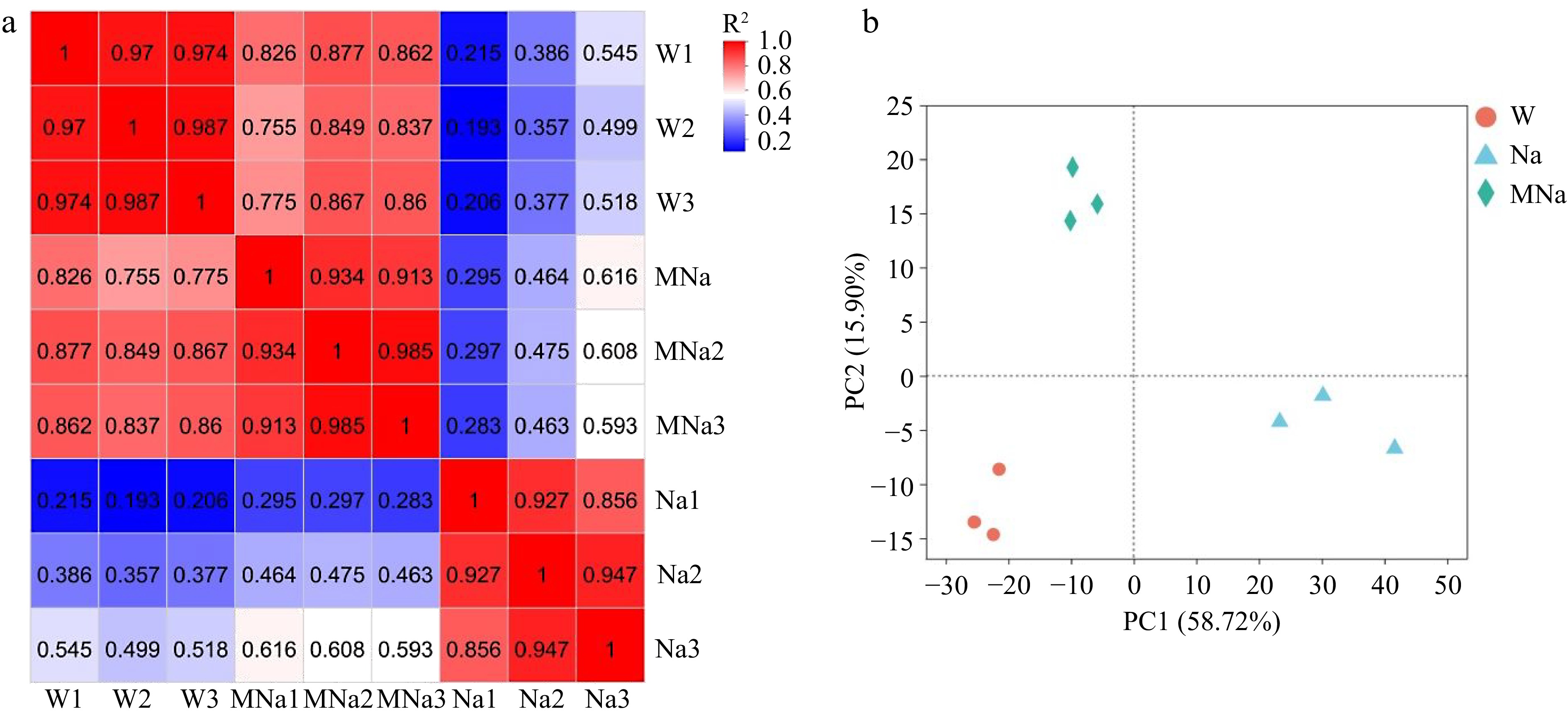
Figure 2.
Overview of the transcriptome samples. (a) Heat map of pearson correlation analysis. (b) Principal component analysis plot. W, Na, and MNa represented water, NaCl, and MT + NaCl treatment, respectively.
Identification of differentially expressed genes (DEGs) among different pair groups
-
To identify the number of DEGs that were significantly regulated among different groups, firstly we constructed the volcano map of DEGs, as shown in Fig. 3a & b, in the three comparative groups (W vs Na, W vs MNa, and Na vs MNa), the number of DEGs which were upregulated decreased and more were downregulated in Na vs MNa group. Compared with the W group, 6,292 (3,564 upregulated and 2,728 downregulated) and 2,795 (1,455 upregulated and 1,340 downregulated) DEGs were identified in the Na and MNa treatment groups, respectively. While 4,198 (1,363 upregulated and 2,835 downregulated) DEGs were identified in the MNa treatment group compared with the Na treatment group. Further comparison of the proportion of DEGs in different treatments by Venn diagram (Fig. 3c) showed that there were 523 DEGs co-expressed in the three treatment groups and more DEGs were identified in MNa vs Na than that in MNa vs W group, suggesting that melatonin could alleviate the salt stress induced damage might by inducing more stress-related DEGs.
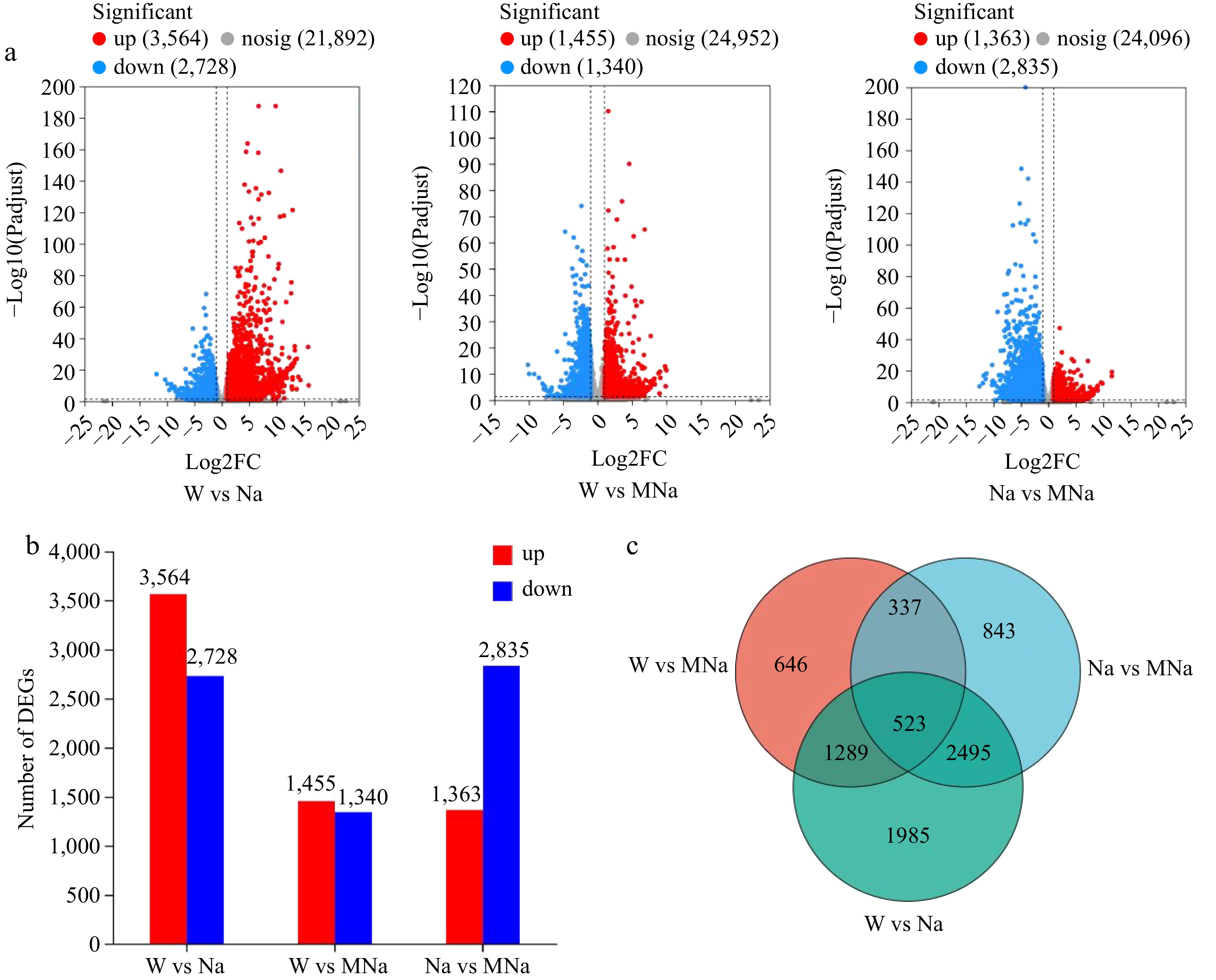
Figure 3.
Identification of the number of differential expression genes (DEGs) in sour jujube seedlings under different treatments. (a) Volcano plots analysis of the number of DEGs. (b) Number of up and down regulated DEGs. (c) Venn diagrams of DEGs.
GO annotation analysis of DEGs among different treatments
-
To gain insight into the biological functions of DEGs among the three treatment groups, GO annotation analysis was performed. As shown in Fig. 4, among the top 20 metabolic pathways mapped to DEGs, the molecular function was mainly enriched in catalytic activity (GO:0003824) and binding (GO:0005488) pathways. The biological process was mainly enriched in metabolic process (GO:0008152), cellular process (GO:0009987), biological regulation (GO:0065007), localization (GO:0051179), response to stimulus (GO:0050896), etc. While in the cellular component, the cell part (GO:0044464) accounts for the most, followed by the membrane part (GO:0044425), membrane (GO:0016020), and organelle (GO:0043226).
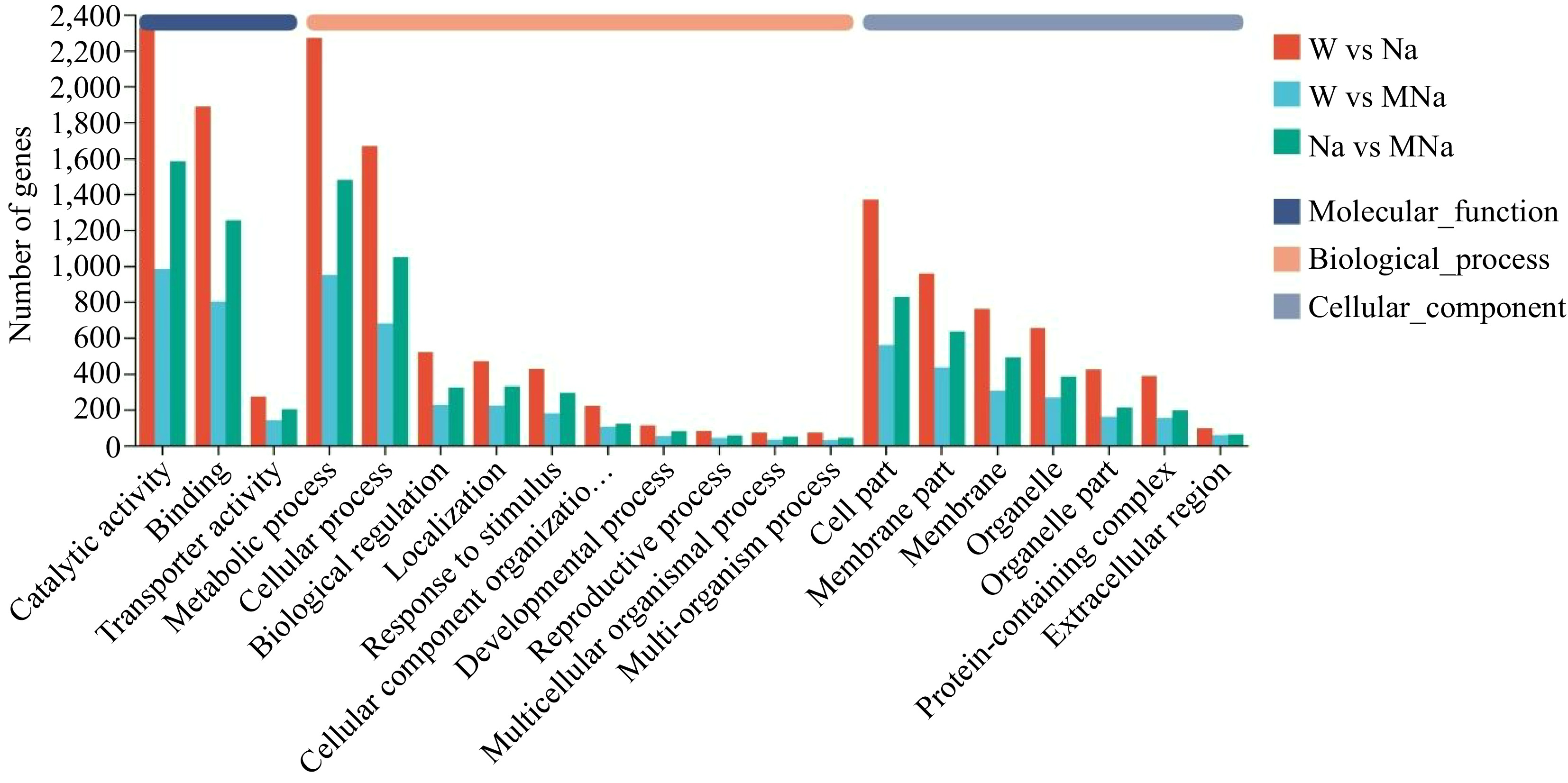
Figure 4.
Histogram of annotation analysis of gene ontology among different pairwise groups.
KEGG enrichment analysis of DEGs among different treatments
-
To comprehensively understand the biological pathways in DEGs among different treatments and determine the main pathways of MT regulated in salt stress, KEGG pathway enrichment analysis was performed. As shown in Fig. 5, 20 top pathways were enriched in these pairwise groups. Among them, DEGs in the W vs Na group were significantly enriched in glyoxylate and dicarboxylate metabolism (map00630), photosynthesis-antenna proteins (map00196), glycolysis/gluconeogenesis (map00010), and monoterpene biology (map00902) pathways. In W vs MNa group, DEGs were significantly enriched in tryptophan metabolism (map00380), anthocyanin biosynthesis (map00942), flavonoid biosynthesis (map00941), phenylpropanoid biosynthesis (map00940), and indole alkaloid biosynthesis (map00901) pathways. In Na vs MNa group, DEGs were significantly enriched in galactose metabolism (map00052), monoterpenoid biosynthesis (map00902), phosphatidylinositol signaling system (map04070), MAPK signaling pathway-plant (map04016), and arginine and proline metabolism (map00330) pathways. These results indicated the key DEGs in these pathways might play important roles in NaCl and MT + NaCl treatments.
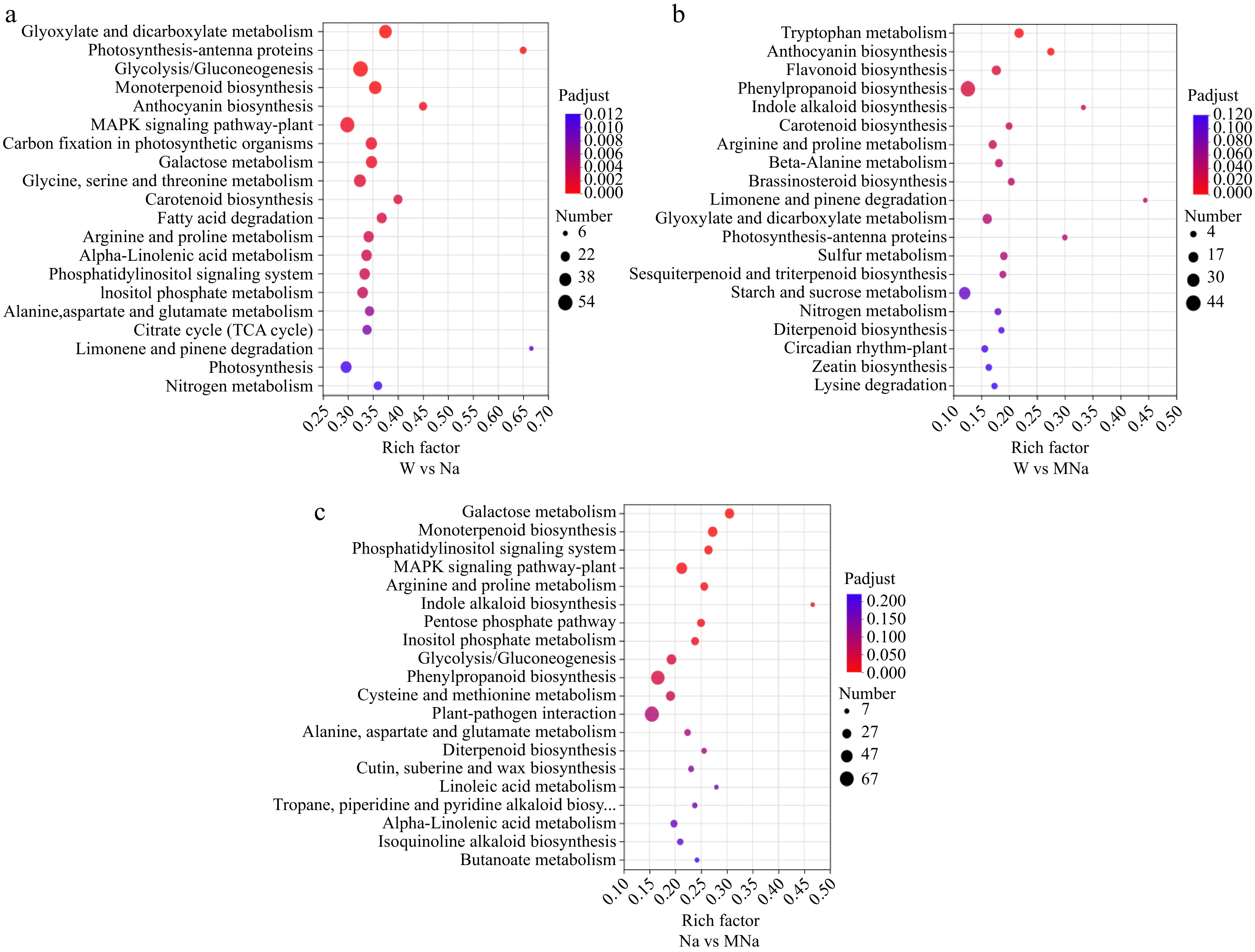
Figure 5.
The top 20 KEGG pathways were analyzed in sour jujube under different pairwise groups, such as (a) W vs Na, (b) W vs MNa, and (c) Na vs MNa. The vertical axis represents the pathway name, the horizontal axis represents the rich factor, the size of dots in the pathway represents the number of DEGs, and the p-adjust value is reflected by the color of the dots.
However, a large number of DEGs were enriched in plant hormone signal transduction (map04075) pathway as well as MAPK signaling pathway and phosphatidylinositol signaling pathway between different treatments, especially in the Na vs MNa group, which might further indicate these pathways function more importanly in MT allevated salt stress inducd damage in sour jujube and should be focused on and deeply analyzed.
Key DEGs involved in MAPK signaling pathway
-
MAPK pathway has been demonstrated to function importantly in various abiotic stresses. The above analysis indicated it might function importanly in MT allevated salt stress inducd damage in sour jujube. As shown in Fig. 6, in the Na vs MNa group, a total of 37 DEGs were enriched in eight sub-pathway of MAPK signaling pathway and we mainly focused on two of them which are related to salt stress (Fig. 6a). Among them, comparing to NaCl treatment, gene13139 encoding mitogen-activated protein kinase kinase kinase 1 (MEKK1) was downregulated in MT + NaCl treatment, while gene6673 encoding abscisic acid receptor PYR/PYL family (PYL) was upregulated. In addition, eight genes, including gene7093, gene7094, gene32214, gene3897, gene21562, gene28461, gene7270, and gene31283 encoding protein phosphatase 2C (PP2C) were downregulated and gene6889 encoding mitogen-activated protein kinase kinase kinase 17/18 (MAPKKK17_18) was downregulated, while gene8078 encoding mitogen-activated protein kinase kinase 3 (MKK3) was upregulated. These results indicated that the gene6673 encoding PYL and gene8078 encoding MKK3 might function positively in MT allevated salt stress inducd damage in sour jujube. To further discuss and excavate the interaction of these key genes, the corresponding protein interaction prediction analysis was performed. As shown in Fig. 6b, gene8078 (mitogen-activated protein kinase kinase 3-like), gene6889 (mitogen-activated protein kinase kinase kinase 2), gene7270 (probable protein phosphatase 2C 51), gene31283 (protein phosphatase 2C 37-like), gene21562 (probable protein phosphatase 2C 51), and gene6673 (abscisic acid receptor PYL4-like) in both pathways were also key nodes which could further regulate the other genes in response to salt stress within MT treatment.
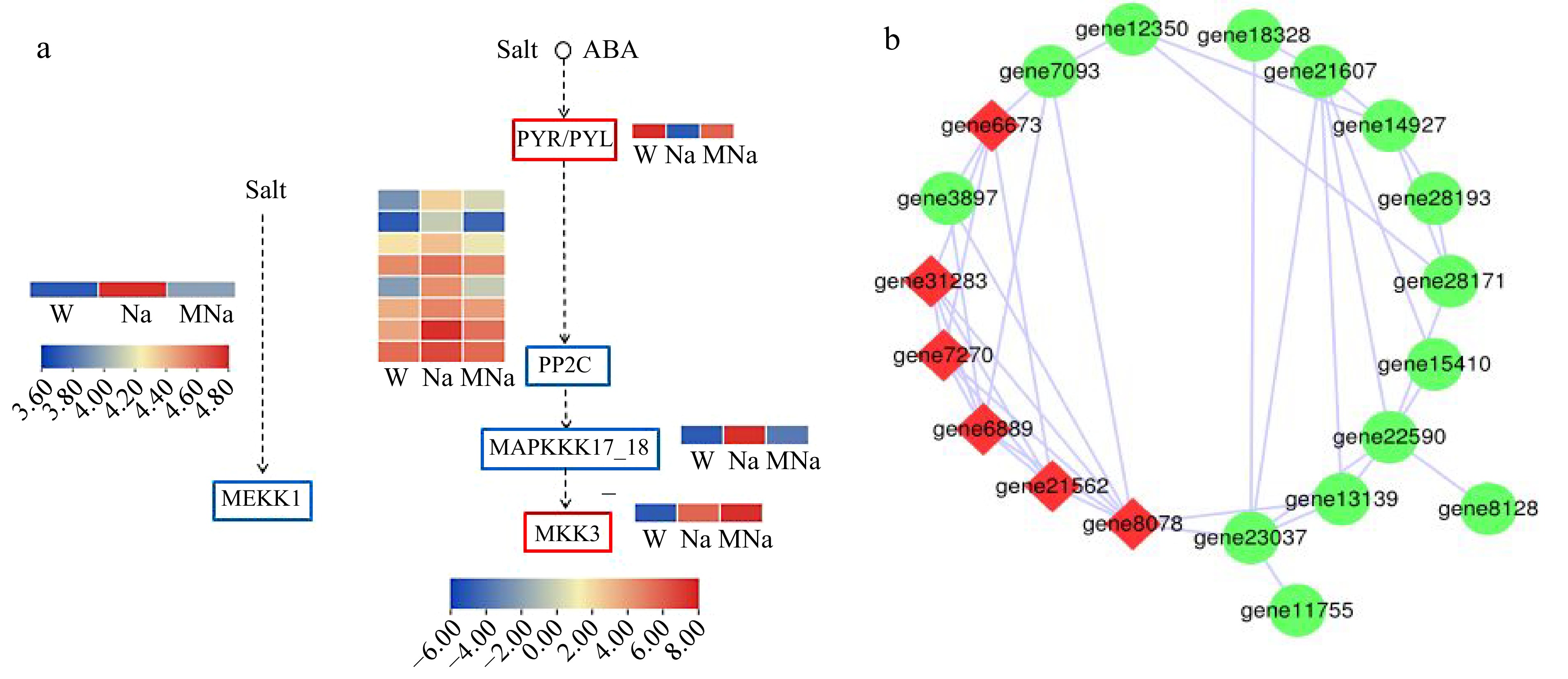
Figure 6.
Key DEGs analysis on MAPK signaling pathway. (a) DEGs analysis in two key MAPK pathways in response to salt stress within MT treatment. Red border represents genes which are upregulated, blue border represents genes which are downregulated. Arrows only represent legend indications. (b) Protein interaction network analysis. Node size represents the strength of protein interactions. Red color indicates the key node proteins.
DEGs involved in phosphatidylinositol signaling system
-
The calcium signaling pathway plays a crucial role in plant perception and response to stress signals[31]. In phosphatidylinositol signaling system pathway, a total of 23 DEGs were enriched under Na vs MNa group, among which genes, including gene11755, gene15410, gene11271, gene8128, were regulated under exogenous melatonin treatment. Compared to NaCl treatment, gene15410 expression was upregulated in MT + NaCl treatment, while gene11755, gene11271, and gene8128 expression was downregulated (Fig. 7a). In addition, protein interaction prediction analysis revealed that gene15254 (inositol polyphosphate multikinase beta-like, transcript variant X1) and gene15254 (probable CDP-diacylglycerol-inositol 3-phosphatidyltransferase 2) are key nodes in exogenous MT treatment under salt stress (Fig. 7b).
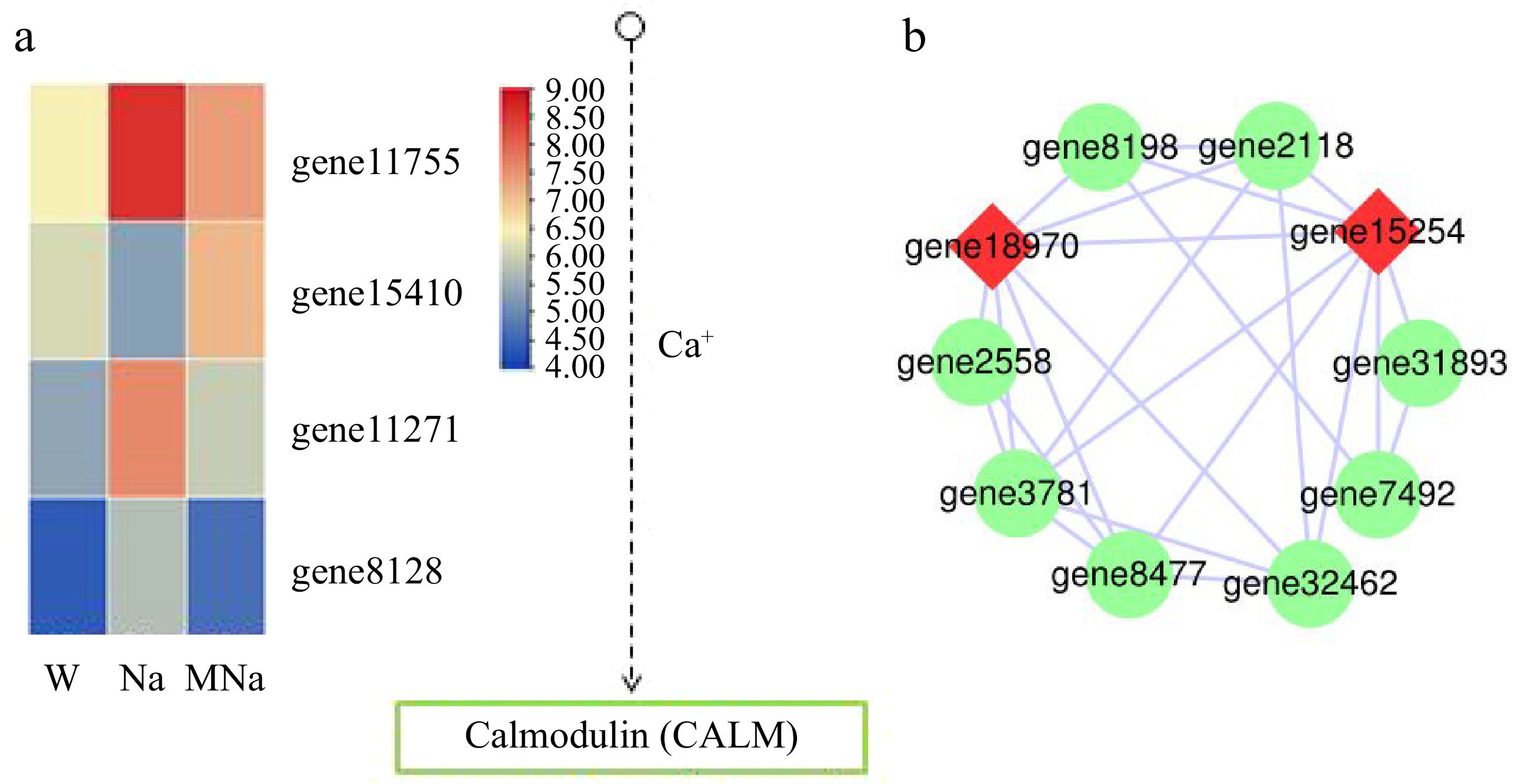
Figure 7.
Key DEGs analysis on phosphatidylinositol signaling system. (a) DEGs analysis in Ca2+ signaling pathway. Arrows only represent legend indications. (b) Protein interaction network analysis. The node size represents the strength of protein interactions. Red color indicates the key node proteins.
DEGs involved in plant hormone signal transduction pathway
-
It has been suggested that plant hormone signaling pathways are associated with biotic and abiotic stresses in plants[32, 33]. In the Na vs MNa group, a total of 42 DEGs were enriched in the plant hormone signal transduction pathway (Fig. 8b), participated in the regulation of eight hormone sub-pathways (Fig. 8a). In the case of auxin signaling, compared to NaCl treatment, six upregulated genes encoded auxin-responsive protein IAA (AUX/IAA), auxin response factor (ARF), and auxin responsive GH3 gene family (GH3), and eight downregulated genes encoded AUX/IAA, ARF, GH3, and SAUR family protein (SAUR) were identified in MT + NaCl treatment. In the case of cytokinin (CTK) signaling, one upregulated gene encoded histidine-containing phosphotransferase protein (AHP) and one downregulated gene encoded two-component response regulator ARR-A family (ARR-A) were identified. Then in the case of other six hormones signaling, several upregulated and downregulated genes were identified. Among them, genes which encoded DELLA protein (DELLA), abscisic acid receptor PYR/PYL family (PYR/PYL), ABA responsive element binding factor (ABF), and jasmonic acid-amino synthetase (JAR1_4_6) were upregulated, demonstrating these genes positively correlated with MT regulated salt stress tolerance in sour jujube. Further protein interaction prediction analysis of 42 DEGs revealed that gene15814 (protein TIFY 10A, transcript variant X1), gene6673 (abscisic acid receptor PYL4-like), gene12590 (auxin response factor 5), gene23244 (auxin-responsive protein IAA27), gene31283 (protein phosphatase 2C 37-like), gene27460 (two-component response regulator ORR10-like), gene14927 (ethylene receptor 2, transcript variant X1), gene28171 (EIN3-binding F-box protein 1-like), and gene20606 (jasmonic acid-amido synthetase JAR1-like) interacted more strongly with other proteins and played an important role in the process of melatonin regulated salt stress tolerance (Fig. 8c).
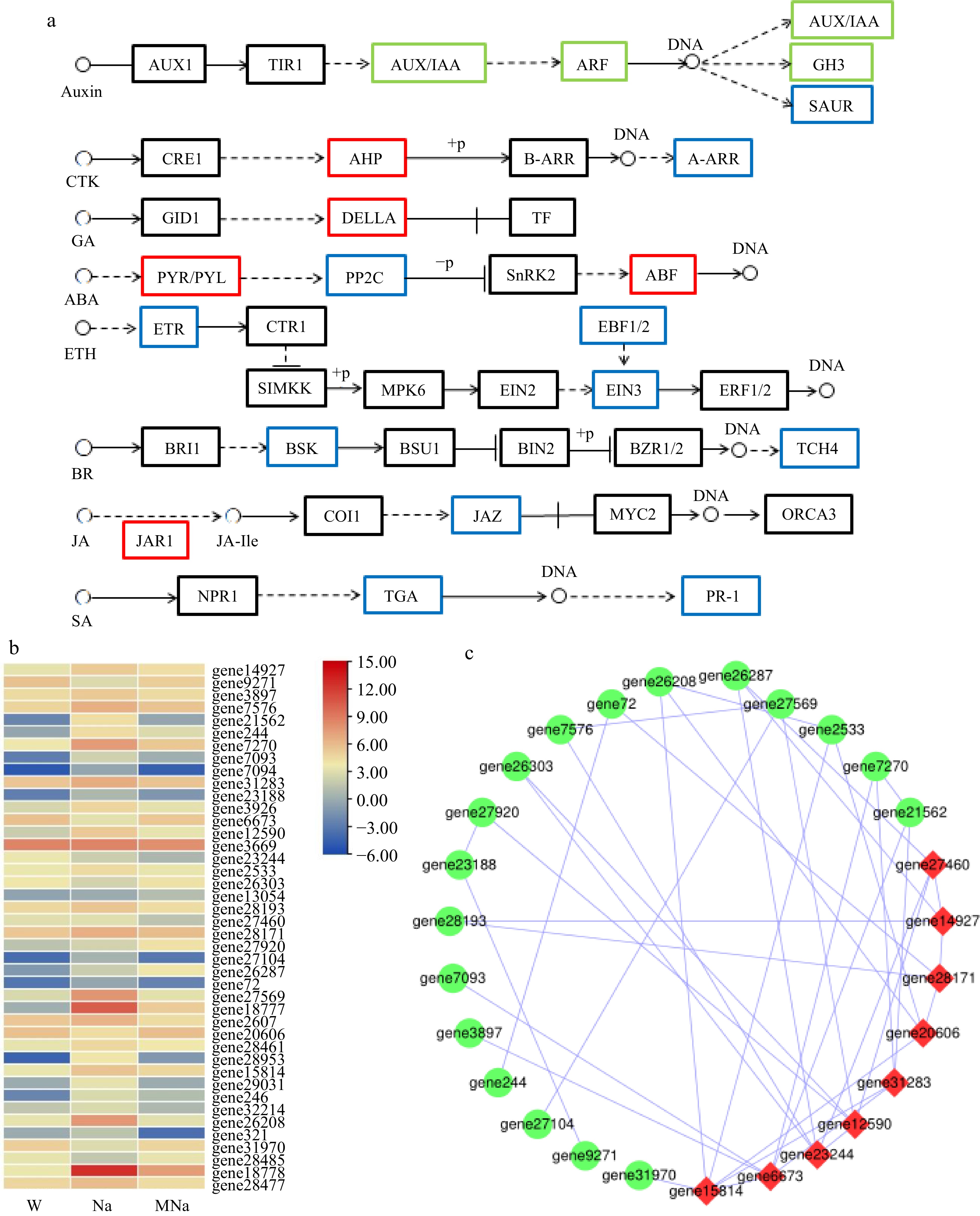
Figure 8.
Key DEGs analysis on plant hormone signal transduction pathway. (a) DEGs analysis in hormone pathways in response to salt stress with MT treatment. Red border represents upregulated genes, blue border represents downregulated genes, and green border represents both up and down regulated genes. Arrows only represent legend indications. (b) Heatmap analysis of related DEGs. (c) Protein interaction network analysis. Node size represents the strength of protein interactions. Red color indicates the key node proteins.
Expressed transcription factors analysis among these treatments
-
Numerous transcription factors (TFs) have been shown to play an important role in the stress response of plants[34]. It is reported that the MYB transcription factor family was involved in the regulation of plant cellular processes and plays a crucial role in various biotic and abiotic stresses[35]. WRKY plays a key role in regulating the stress response of plants and can improve salt tolerance[36]. NAC is involved in the regulation of different developmental processes in plants and has a positive regulatory role in stress-induced senescence in Arabidopsis[37]. Here, these three transcription factor families were induced in the top six TFs in different comparative groups. Among them, in the W vs Na group, 31 DEGs belonging to MYB transcription factor family, 30 DEGs of WRKY transcription factor family and 26 DEGs of NAC transcription factor family were identified (Fig. 9a). While in the Na vs MNa group, 32, 25, and 16 DEGs of MYB, WRKY, and NAC family have been found, respectively (Fig. 9b). The DEGs belonging to MYB, WRKY, and NAC family between the W vs Na and Na vs MNa groups might function importantly in MT regulated salt tolerance in sour jujube.
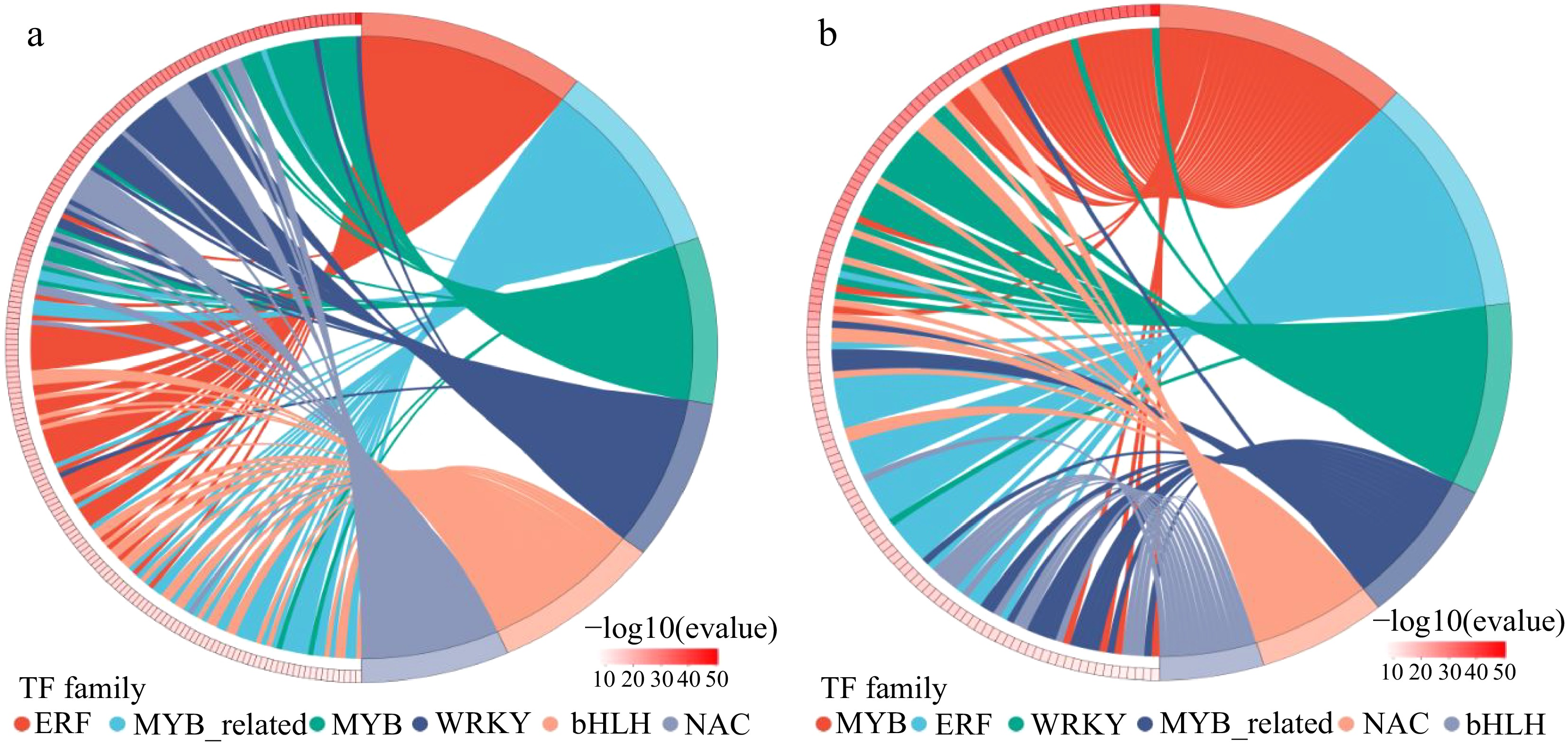
Figure 9.
Distribution map of expressed transcription factor families in (a) W vs Na group and (b) Na vs MNa group. The horizontal coordinates are different transcription factor families, the vertical coordinates are number of genes falling into that transcription factor family.
-
When plants suffer salt stress, harmful cellular changes and phenotypical symptoms, including compatible osmolytes, ion homeostasis, reactive oxygen species (ROS), hormones, are always observed[38]. For example, under salt stress, cucumber seedlings have restricted chlorophyll synthesis, reduced germination rate, inhibited growth, and reduced biomass and yield[39, 40]. In citrus seedlings, salt stress significantly reduced plant height, leaf area, leaf water content, chlorophyll content, and restricted ion uptake[41, 42]. In short, the growth status of different plant species was negatively affected by salt stress to some extent. As an effective growth regulator, melatonin can alleviate the adverse effects of salt stress on fruit trees[43]. After exogenous application of melatonin, the fruit tree had increased nutrient uptake, improved leaf cuticle, reduced osmotic and water loss, and decreased leaf wilting under salt stress[44]. Moreover, MT treatment could also reduce production of stress induced substances, such as ROS, lipid peroxides, H2O2, and MDA, and limited losses of photosynthetic pigments and improved defense systems[45]. For example, in banana seedlings, the application of exogenous MT can enhance salt tolerance by increasing the content of proline content and soluble protein, slowing down the degradation of chlorophyll, reducing membrane permeability, increasing the accumulation of MDA, and enhancing antioxidant defense activity. In our current study, NaCl treatment could induce leaf curling and yellowing, while MT treatment could alleviate the salt stress induced damage in sour jujube seedlings. However, which physiological parameters involved in this process should be further elucidated.
To initially reveal the molecular mechanism of MT treatment on sour jujube seedlings in response to salt stress, the transcriptomic analysis was performed. It was found more DEGs were identified in MNa vs Na group than that in MNa vs W group. KEGG enrichment analysis showed these DEG mainly involved in plant hormone signal transduction pathway as well as MAPK signaling pathway and phosphatidylinositol signaling pathway. In the study of salt stress on banana, cotton, and rapeseed seedlings, plant hormone synthesis and its signal transduction, lignin and fatty acid metabolism, and phenylalanine metabolism play important roles in MT regulated salt stress[38, 46]. Together with our results, these major metabolism pathways, especially the plant hormone signal transduction pathway may be involved in the potential mechanism of MT effect on salt stress.
Plant hormones play an important role in resistance to abiotic stresses[47−49]. ABA is a plant defense hormone that improves plant resistance. Under salt stress, endogenous ABA could be significantly induced which could activate the synthesis of related protein kinases to improve salt stress[50]. GA is involved in multiple processes of plant growth and can promote the growth of tree stems, which is a stimulating factor in plants[51]. In response to salt stress, GA levels could be reduced which further inhibited plant growth[52]. Other hormones could also regulate the plant growth and development and function importantly in response to stresses. For example, the increase of SA level is related to the improvement of active oxygen scavenging and antioxidant capacity, and plays an important role in improving plant abiotic stress. JA can also improve salt tolerance by enhancing its antioxidant capacity. According to reports, MusaNAC29 transcription factor is involved in regulating drought and salt tolerance in bananas by altering the content of plant hormones such as SA and JA[53]. MT is an important regulator of the expression of plant hormone-related genes and plays an important role in response to salt stress. In cotton, salt stress could decrease the endogenous contents of MT and enhance the accumulation of IAA, GA, BR, and ABA[46]. In banana, auxin, cytokinin and gibberellin signaling were identified function importantly under MT + NaCl treatment. In our study, 42 DEGs involving in different hormone signal transduction pathways were identified. Among them, genes encoded DELLA, PYR/PYL, ABF, and JAR1 were upregulated, demonstrating these genes positively correlated with MT regulated salt stress tolerance in sour jujube. However, the biological function of their involvement in MT regulated salt tolerance in sour jujube should be further demonstrated. If conditions permit, further exploration will be conducted on the changes in plant hormone content of jujube seedlings under salt stress accompanied with MT treatment to further confirm how MT regulated the changes of hormones to alleviate the damages induced by salt stress.
As well as plant hormone signal transduction pathways, Ca2+ and MAPK signaling pathways act together within exogenous MT regulated salt stress in sour jujube seedlings. Among them, the MAPK signaling pathway has two key sub-pathways which were related to salt stress, and 12 DEGs involved in this process. Under different abiotic stress with MT treatment, the function of the MAPK pathway has also been demonstrated. For example, the number of DEGs involving in MAPK signaling pathway was significantly affected by exogenous MT treatment under stress in maize[31]. Under salt stress, the expression of MAPK genes was significantly upregulated in plum with exogenous MT treatment[54]. In oat seedlings, MAPKs involving in the ABA signaling pathway participated in drought stress[55]. Here, the gene6673 encoding PYL and gene8078 encoding MKK3 might function positively in MT allevated salt stress induced damage in sour jujube and deserve further study.
Ca2+ plays a regulatory role in stress response during plant growth and development. When plants are subjected to various external stimuli, such as temperature, light, salt, and drought, Ca2+ will be released into the cytoplasm through Ca2+ channels to generate specific signals for signal transduction[56]. In our study, exogenous application of melatonin treatment induced the expression of gene11755, gene15410, gene11271, and gene8128 under salt stress, thereby regulating calmodulin protein (CALM), which may promote Ca2+ uptake to mediate signal transduction and mitigate the damage caused by salt stress to sour jujube seedlings. However, all the above pathways, especially the corresponding DEGs should be functionally demonstrated in future studies.
As regulators of genes, transcription factors have been confirmed to perform vital functions in regulating abiotic stress responses in plants[57]. It is shown that different types of transcription factors, mainly including AP2/ERF, NAC, bZIP, MYB, WRKY can play a crucial role in response to various stress[58]. In the current study, the transcription factors MYB, WRKY, and NAC were identified, which are associated with adversary stresses, and possibly regulate a large number of DEGs to promote MT induced salt tolerance in sour jujube seedlings.
To take all the data together, we come to a proposed model where MT treatment could regulate the MAPK pathway and the genes related to hormones to confirm MT induced salt tolerance in sour jujube. Meanwhile, calcium signaling was also involved (Fig. 10). The current results and proposed model could provide some theoretical basis for understanding the molecular mechanism of how applying exogenous MT alleviated the damage caused by salt stress to sour jujube seedlings, but the specific regulatory mode and pathway need to be further explored.
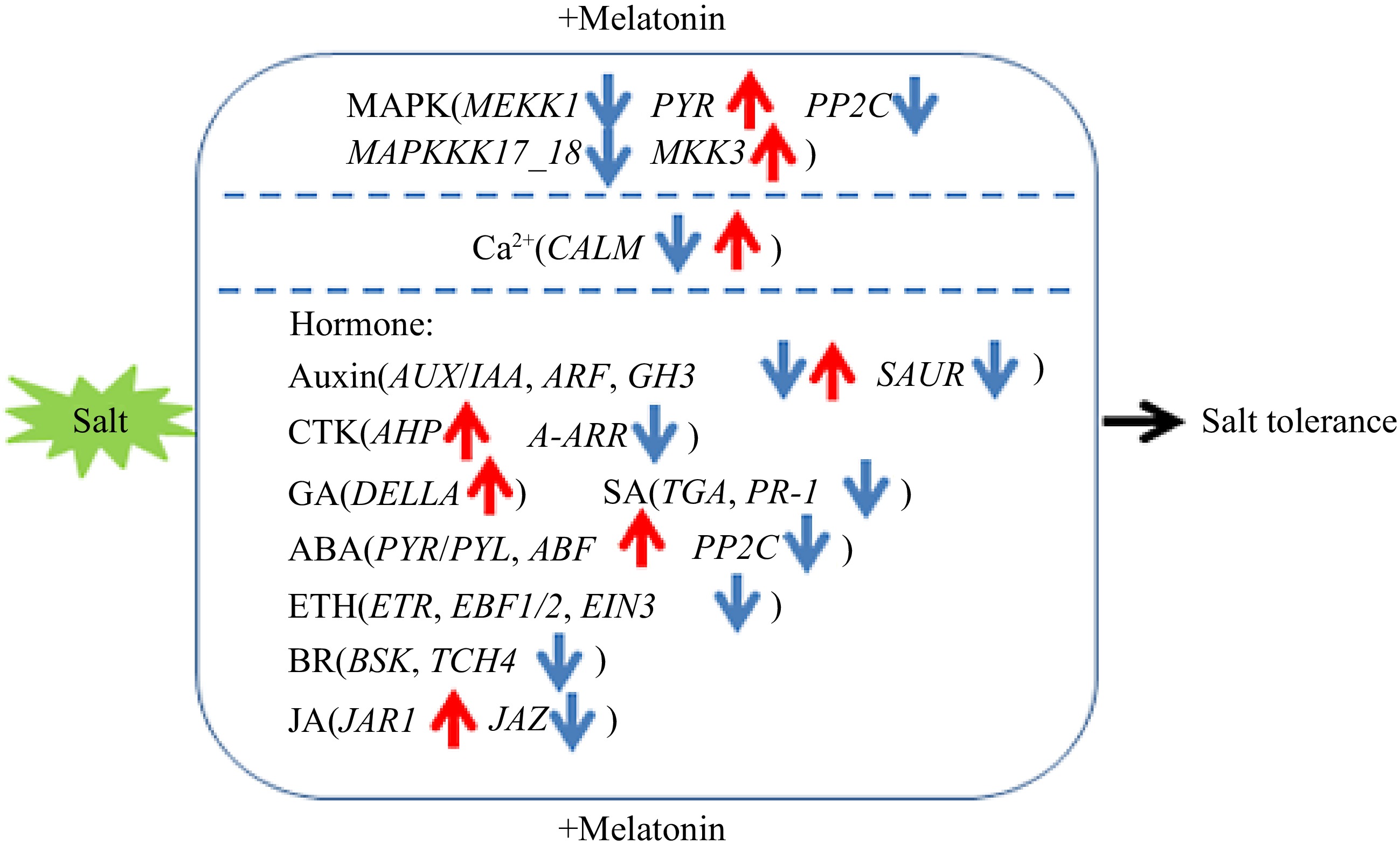
Figure 10.
Proposed model of exogenous melatonin application on effects of sour jujube seedlings in response to salt stress. Blue arrows represent regulatory downregulated genes and red arrows represent regulatory upregulated genes.
This research was funded by Provincial Supporting Program of Hebei for the Returned Oversea Scholars (C20210114); Science and Technology Project of Hebei Education Department (QN2022017); Fundamental Scientific Research Fund of Universities in Hebei Province (KY2021059); China Agriculture Research System (CARS-30-2-07); Natural Science Foundation of Hebei province (C2020204082); Funds for Hebei Jujube Industry Technology Research Institute after Operation Performance (205676155H); Young Talent Project of Hebei Agricultural University Foundation (YJ201853).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Wenhui Zhu, Haonan Cao
- Supplemental Fig. S1 Overall observation of phonotypical changes of sour jujube seedlings after192 h NaCl (a) and MT plus NaCl (b) treatments, respectively.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhu W, Cao H, Wang Q, Niu Y, Sadeghnezhad E, et al. 2023. Transcriptome analysis revealed MAPK and hormone pathway involving in exogenous melatonin-regulated salt tolerance in sour jujube. Fruit Research 3:19 doi: 10.48130/FruRes-2023-0019 

Transcriptome analysis revealed MAPK and hormone pathway involving in exogenous melatonin-regulated salt tolerance in sour jujube
- Received: 04 May 2023
- Accepted: 29 June 2023
- Published online: 03 August 2023
Abstract: Melatonin (MT) plays an important role in plant growth and development, and response to biotic and abiotic stresses. Salt stress is one of the major abiotic stresses affecting plant growth and distribution. Sour jujube mainly grows in mountainous sandy land and faces the threat of salt stress. Thus, to explore whether exogenous MT application could improve the salt stress tolerance of sour jujube and reveal the molecular mechanism behind it, the phenotypic and transcriptomic analyses of different concentrations of MT treatments prior to sodium chloride (NaCl) treatment to sour jujube was investigated. Firstly, exogenous MT treatment has a certain alleviating effect on salt stress induced damage of sour jujube. Transcriptome analysis showed that 6,292, 2,795, and 4,198 differentially expressed genes (DEGs) were identified in water treatment (W) vs 400 mmol/L NaCl treatment (Na), W vs 10 µmol/L MT plus 400 mmol/L NaCl treatment (MNa), and Na vs MNa comparative groups, respectively. KEGG enrichment analysis of DEGs showed that multiple metabolic pathways were involved in MT regulated effect on salt stress, including mitogen-activated protein kinase (MAPK) signaling pathway, phosphatidylinositol signaling system, and plant hormone signal transduction pathways etc. In addition, some transcription factor families, such as MYB, WRKY, and NAC, were also activated by exogenous MT treatment under salt stress. The results suggest that MT can improve the tolerance of sour jujube to salt stress mainly through the regulation of MAPK, hormone signaling pathways, and related transcription factors, which could lay the foundation for further study on the molecular mechanisms of how exogenous MT regulates salt stress tolerance in sour jujube.
-
Key words:
- Sour jujube /
- Melatonin /
- Salt stress /
- Transcriptome /
- MAPK /
- Hormones