








-
Apple (Malus × domestica Borkh.) is one of the most economically important fruits in the world. China is the biggest apple producer globally with a producing area of about 2.04 million hectares which produced 42.43 million tons of apple fruits in 2021 (
https://www.fao.org/statistics/en ). As a vegetatively propagated crop, infection by intracellular pathogens such as virus and viroid weaken tree vigor, cause graft incompatibility, result in a 15%−50% reduction in apple yield[1, 2].Apple stem grooving virus (ASGV) belongs to the Capollovirus genus of the family Betaflexiviridae. It widely occurs worldwide and is often found co-infected with other viruses, such as apple stem pitting virus (ASPV), apple chlorotic leaf spot virus (ACLSV), and apple mosaic virus (ApMV)[2−5]. ASGV is mainly transmitted by apple grafting and remains latent in most apple cultivars[3−5]. However, the infection of ASGV may cause graft-incompatibility, and reduce bud break[3]. ASGV can also infect other important crops, including citrus, pear, apricot, cherry, lily, ficus palmata, soybean[3, 6].
An accurate and rapid virus detection method is crucial for the management of viral diseases as well as for the implementation of virus eradication programs[7−9]. To date, reverse transcription polymerase chain reactions (RT-PCR) and enzyme-linked immunosorbent assays (ELISA) are the most widely used methods for detection of RNA viruses[10−12]. However, the preparation of antibodies is a necessity in ELISA test and lower sensitivity has often been reported in ELISA tests as compared with RT-PCR. The RT-PCR protocol is labor-intensive as it consists of the RNA extraction and purification, cDNA synthesis, PCR and detection of the PCR amplicons, which also require expensive equipment such as a PCR cycler[8]. These drawbacks greatly inhibit the broad use of virus detection, and particularly the on-site virus detection, for management of viral diseases. These drawbacks greatly inhibit the broad use of virus detection, particularly on-site virus detection, for managing viral diseases.
Recombinase polymerase amplification (RPA) technology depends on recombinases, strand-displacing DNA polymerases, and single-strand DNA-binding proteins for isothermal nucleic acid amplification and is known with the potential to replace PCR for the amplification of specific DNA segment[8, 13, 14]. RPA reactions can be rapidly performed at 37−42 °C with a high level of sensitivity[15, 16]. Moreover, RPA oligonucleotide probe can be recognized and cut by endonuclease IV (nfo) or exonuclease III (exo), so that the RPA amplification can be visualized by real-time fluorescence detection through nfo or exo probes or the use of LFD, therefore is free of nucleic acid electrophoresis[13, 17, 18]. Several studies have reported the use of RT-RPA-LFD for the detection of little cherry virus-2[19], penaeus stylirostris denso virus[20], bean common mosaic virus[18] and Potato Virus Y[21]. It is worth noting, that due to the high sensitivity of RPA reaction, crude RNA extract can be used for detection[22]. In this study, we developed a simple and sensitive method using RT-RPA-LFD technique for the rapid detection of ASGV. The crude extract of RNA was also tested in this study to facilitate the on-site detection of ASGV in the field.
-
To evaluate the effectivity of RT-RPA-LFD assay for the detection of ASGV in apple plants, ASGV-infected samples were collected from Apple Experiment Station in Shaanxi province, China. The total RNA as the template for RT-RPA-LFD reaction, the positive controls 'Gala' was obtained from Wang et al. [23], while ASGV-free 'Gala' was used as the negative control (Fig. 1c). RT-RPA-LFD results showed that a test line from positive samples, while no such bands can be observed from negative control (Fig. 1a). Similarly, the results of agarose gel electrophoresis of RT-RPA products show a band of 264 bp from the positive control, while no such band can be found in the negative control (Fig. 1b). These results indicate that the test line observed from the LFD was associated with the amplification of the ASGV genome by the RT-RPA reaction.
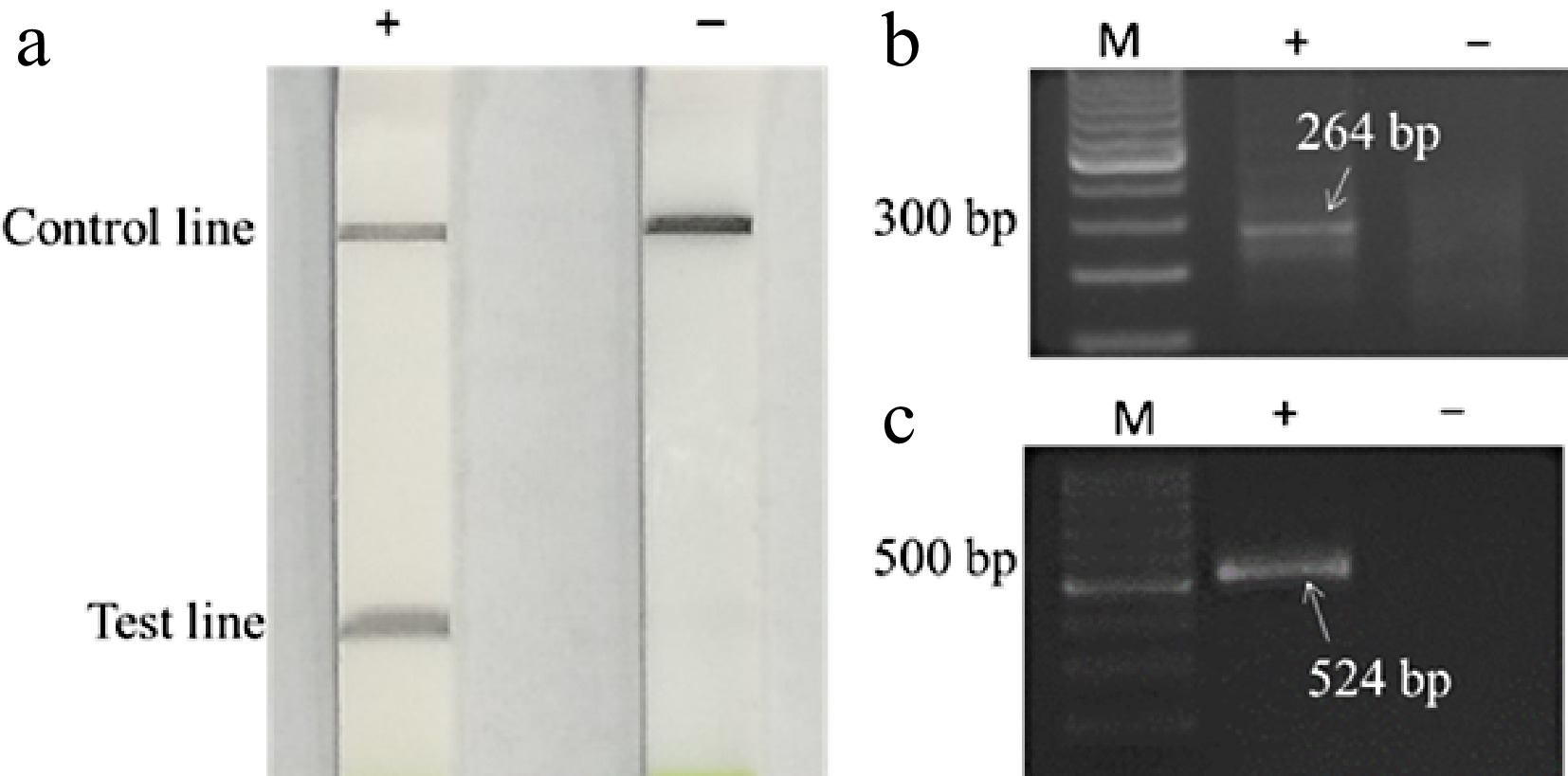
Figure 1.
One-step RT-RPA and RT-PCR detection of ASGV. (a) One-step RT-RPA-LFD detection of ASGV. (b) One-step RT-RPA-gel detection of ASGV. (c) RT-PCR-gel detection of ASGV. In two gel images, lane M is a 100 bp DNA marker; lean '+' is positive; lean '−' is negative.
To optimize RT-RPA reaction conditions for ASGV detection. The RT-RPA reaction mixture was incubated at 40 °C for 0, 10, 20, 30 and 40 min. The LFD results showed that the strongest test line was found when the reaction time was 30 min and 40 min (Fig. 2a). The agarose gel electrophoresis further showed that the brightest band was obtained after 30 min of incubation (Fig. 2a). The optimization of the RT-RPA reaction temperature found the RPA reaction performed at 39 °C and 40 °C exhibited strongest bands on LFD, while weaker signals were obtained from the lower (38 °C) and the higher (41 °C) temperatures. Accordingly, the agarose gel electrophoresis produced clear and strong bands when RPA reaction was maintained at 39 °C and 40 °C for 30 min (Fig. 2b).

Figure 2.
Optimization of one-step RT-RPA reaction conditions for ASGV detection. (a) Optimization of ASGV detection for different reaction time (0, 10, 20, 30 and 40 min) at 40 °C. (b) Optimization of ASGV detection at various temperatures 37 °C, 38 °C, 39 °C, 40 °C and 41 °C for 30 min.
Specificity and sensitivity of RT-RPA-LFD
-
The in vitro apple plants 'Yanfu 8' and 'Gala' single-infected with ACLSV, ASPV or ASGV were used as test materials, the in vitro 'Yanfu-8' co-infected with three viruses (ASGV, ASPV and ACLSV) was used as a positive control, and the virus-free 'Gala' was served as a negative control for both RT-PCR and RT-RPA-LFD detection. In RT-PCR detection, only one band of 794 bp, 370 bp, and 524 bp was shown from the sample single-infected by ACLSV, ASPV and ASGV, respectively (Fig. 3a−c). In the RT-RPA-LFD detection for ASGV, only the sample infected by ASGV showed positive results, while no such bands were detected from samples single infected by either ACLSV or ASPV (Fig. 3d).
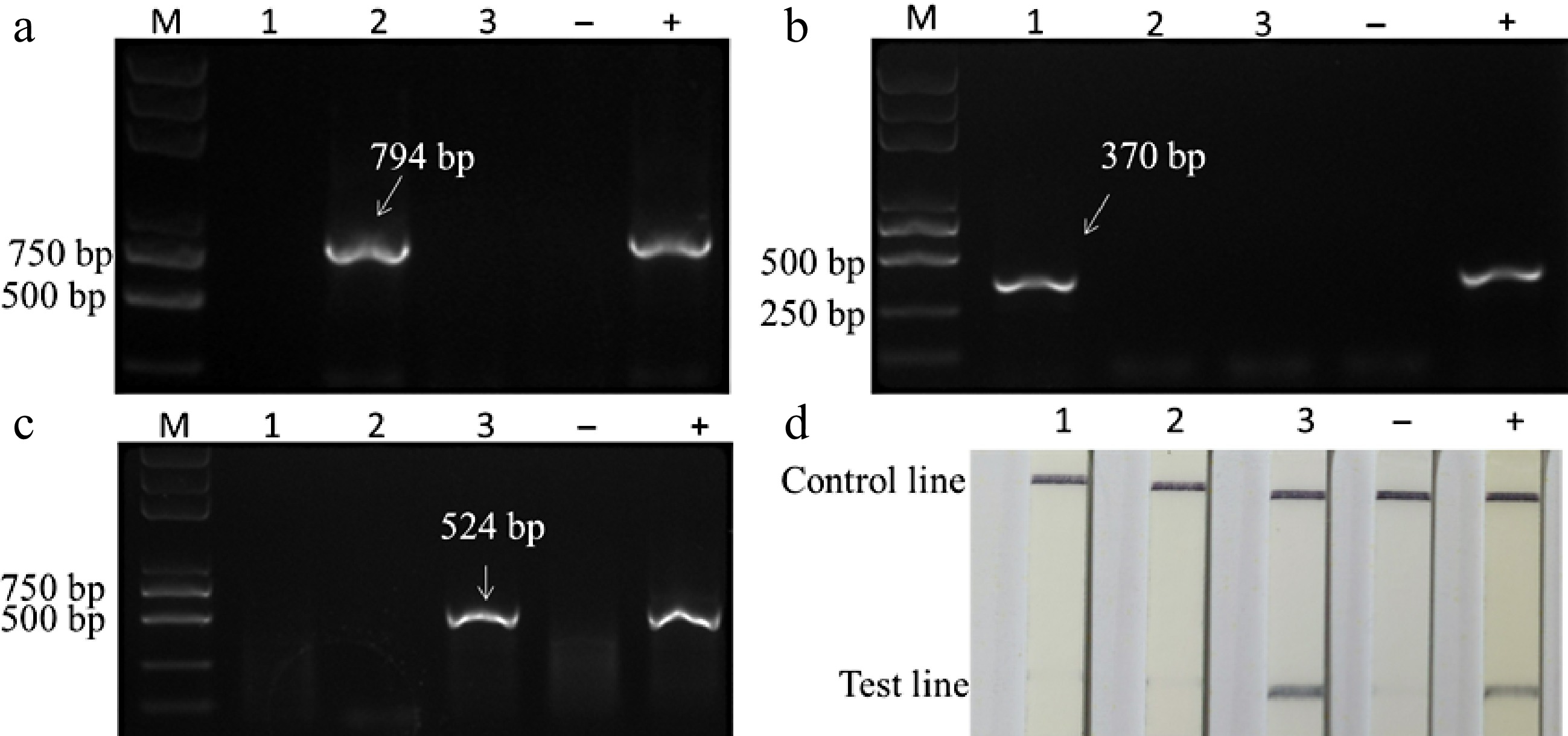
Figure 3.
Specificity of one- step RT-RPA-LFD assays. (a) RT-PCR detection of ACLSV. (b) RT-PCR detection of ASPV. (c) RT-PCR detection of ASGV. (d) One- step RT-RPA-LFD detection of ASGV. In all images, lane 'M' is 2000 bp DNA marker, lanes 1−3 are samples Yanfu 8-3, Yanfu 8-2 and GL-1 which was single-infected by ACLSV, ASPV and ASGV, respectively. '−' negative control, '+' positive control.
In sensitivity tests, the positive sample of total RNA (500 ng/μL) was serially diluted to 10−8 in a 10-fold gradient as a sensitivity comparison templates. The results show that RT-RPA-LFD can detect to 10−6 (Fig. 4a), while RT-PCR can detect to 10−4 (Fig. 4b). Therefore, RT-RPA-LFD is 100 times more sensitive than RT-PCR (Fig. 4).
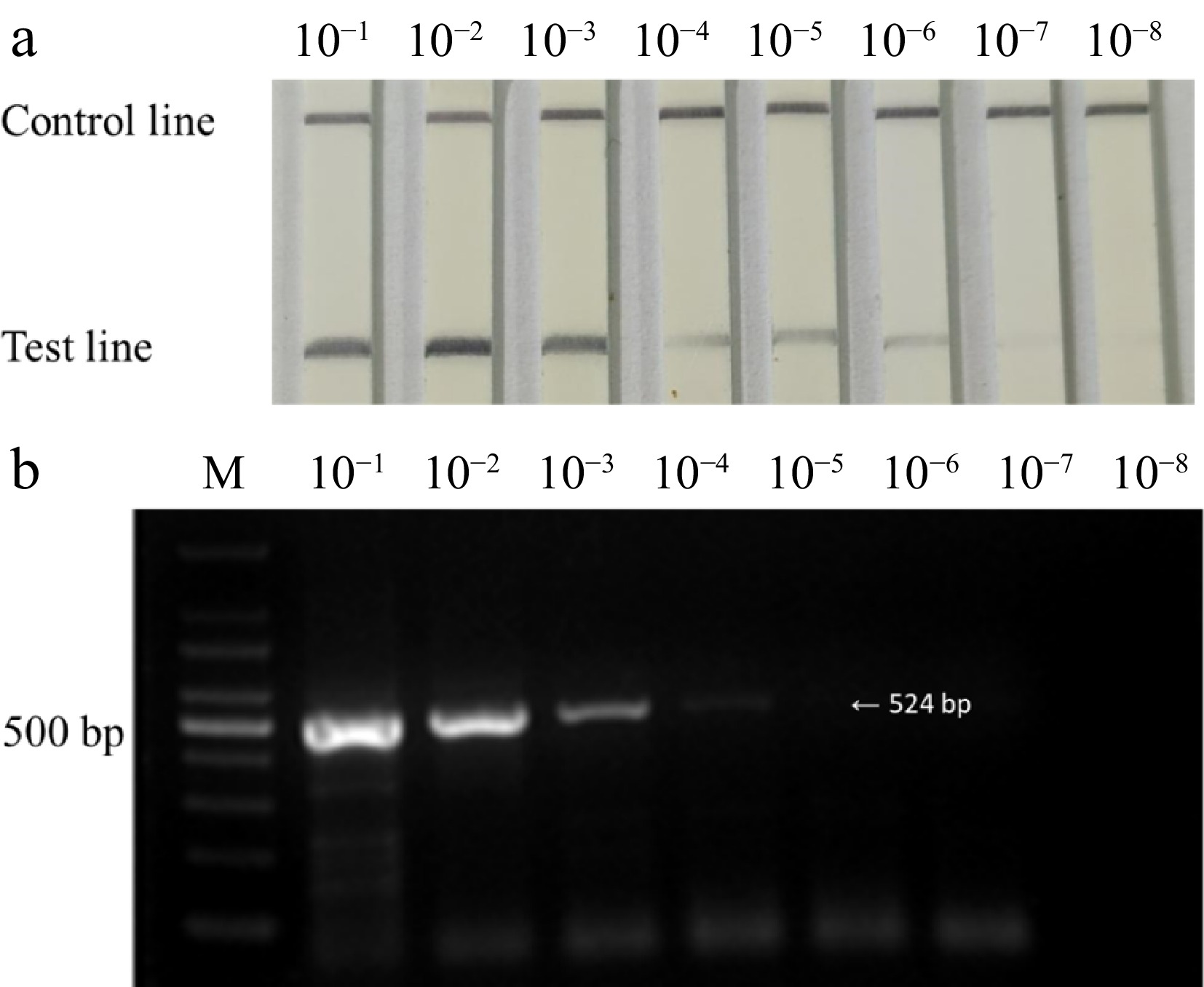
Figure 4.
One- step RT-RPA-LFD and RT-PCR amplicons at total RNA (500 ng/μL) dilutions levels ranging from 10−1 to 10−8 detect ASGV. (a) Detect ASGV by one- step RT-RPA-LFD. (b) Detect ASGV by RT-PCR. Lane M (b): 100 bp DNA marker.
Rapid detection of ASGV by one-step RT-RPA-LFD using crude RNA
-
Based on high sensitivity of RT-RPA-LFD, this study tested the use of crude RNA for rapid detection of ASGV (Fig. 5) using ASGV-infected leaf samples. Clear test line was observed on the LFD while no test line could be found from the negative sample. Comparison between conventional RT-PCR protocol and RT-RPA-LFD using crude RNA extracts showed the same ASGV detection results (Table 1). With the use of RNA crude extracts, the entire assay can be completed within 1 h (Fig. 5).
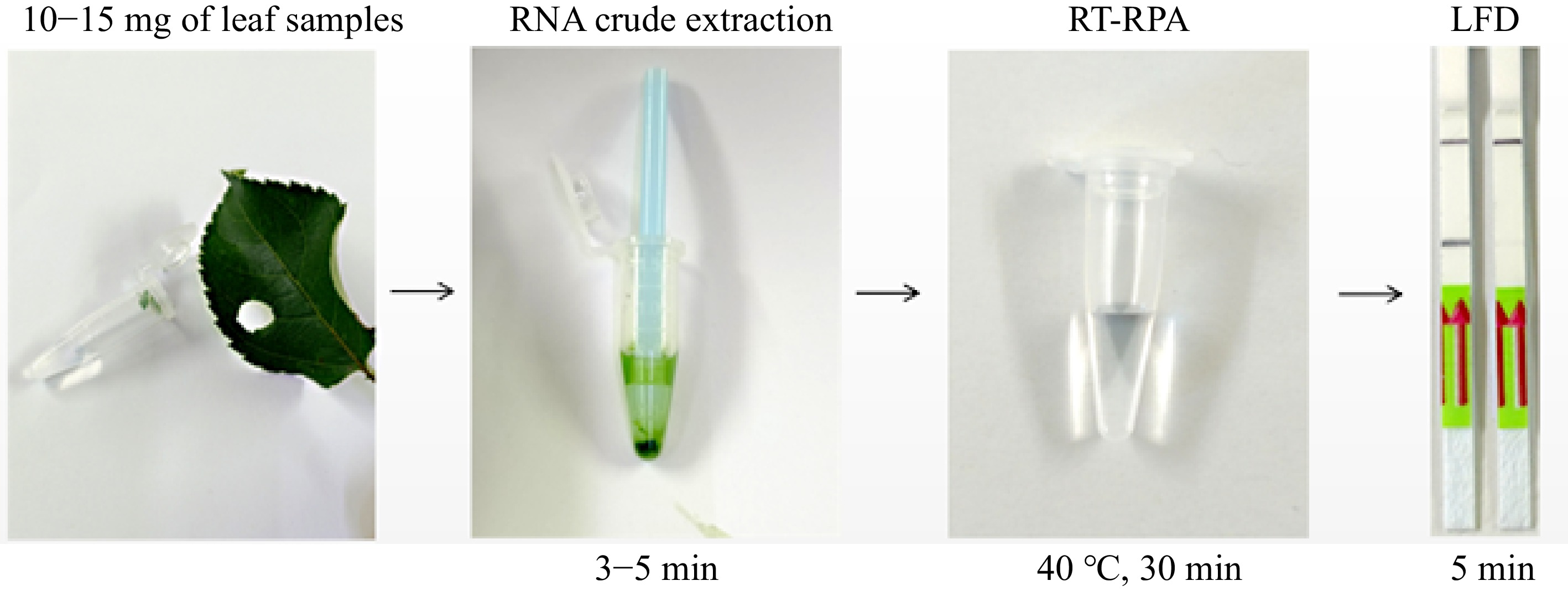
Figure 5.
Rapid detection of ASGV with one-step RT-RPA-LFD using crude RNA.
Table 1. Comparison on the ASGV detection results between conventional RT-PCR protocol and One-step RT-RPA-LFD using crude RNA extracts.
Sample no. RT-PCR One-step
RT-RPA-LFDASGV ASPV ACLSV ASGV Gala-1 (Positive control) + − − + Gala-2 (Negative control) − − − − Gala-3 + + − + Gala-4 − − + − Gala-5 + + + + Gala-6 + − + + Gala-7 + − − + Gala-8 + − − + Gala-9 + − − + Gala-10 + − + + '+': ASGV positive; '−': ASGV negative. -
In this study, a one-step RT-RPA-LFD method was established and was proved suitable for detection of ASGV from in vitro cultured shoots and leaf sampled from the field.
Compared with traditional molecular biology and immunology detection techniques such as RT-PCR and ELISA assays, RT-RPA has unique advantages and can complete isothermal amplification[21, 24, 25]. As one of the most commonly used detection methods, RT-PCR requires a variable temperature instrument, while the amplification time is 90−180 min, and the gel visualization product can be observed after 20−50 min[9, 26]. At the same time, the entire detection of the viruses by ELISA takes 2 d. It has been reported that RT-LAMP can detect viruses at 58−66 °C for 60 min, but this method requires four pairs of primers[27−30]. However, the RT-RPA can complete amplification in 20−40 min at 25−40 °C, can be carried out with simple experimental equipment[31−33]. In addition, this study used RNA as a template, and by adding M-MLV reverse transcriptase to the RPA system, the results showed that the one-step RT-RPA using a template RNA extract was successful (Fig. 1). Compared with RT-PCR, there is fewer step of reverse transcription of RNA into cDNA, and the product can be visualized in 5 min with LFD, which is not limited to gel electrophoresis detection of products, and also reduces the cost of using chemical reagents.
The design of primers amenable to RPA amplification is a necessity in using RT-RPA technique to detect of RNA viruses. RPA primers are usually longer than PCR primers[21, 25]. In order to obtain RPA primers specific to the ASGV genome, we downloaded and compared the ASGV coat protein (CP) gene in apples and designed an RPA primer from highly conserved regions of the CP gene (Fig. 6). Specificity validation of this study showed that only samples infected with ASGV present positive test lines in RT-RPA-LFD assay, thus confirming the specificity of this detection method.
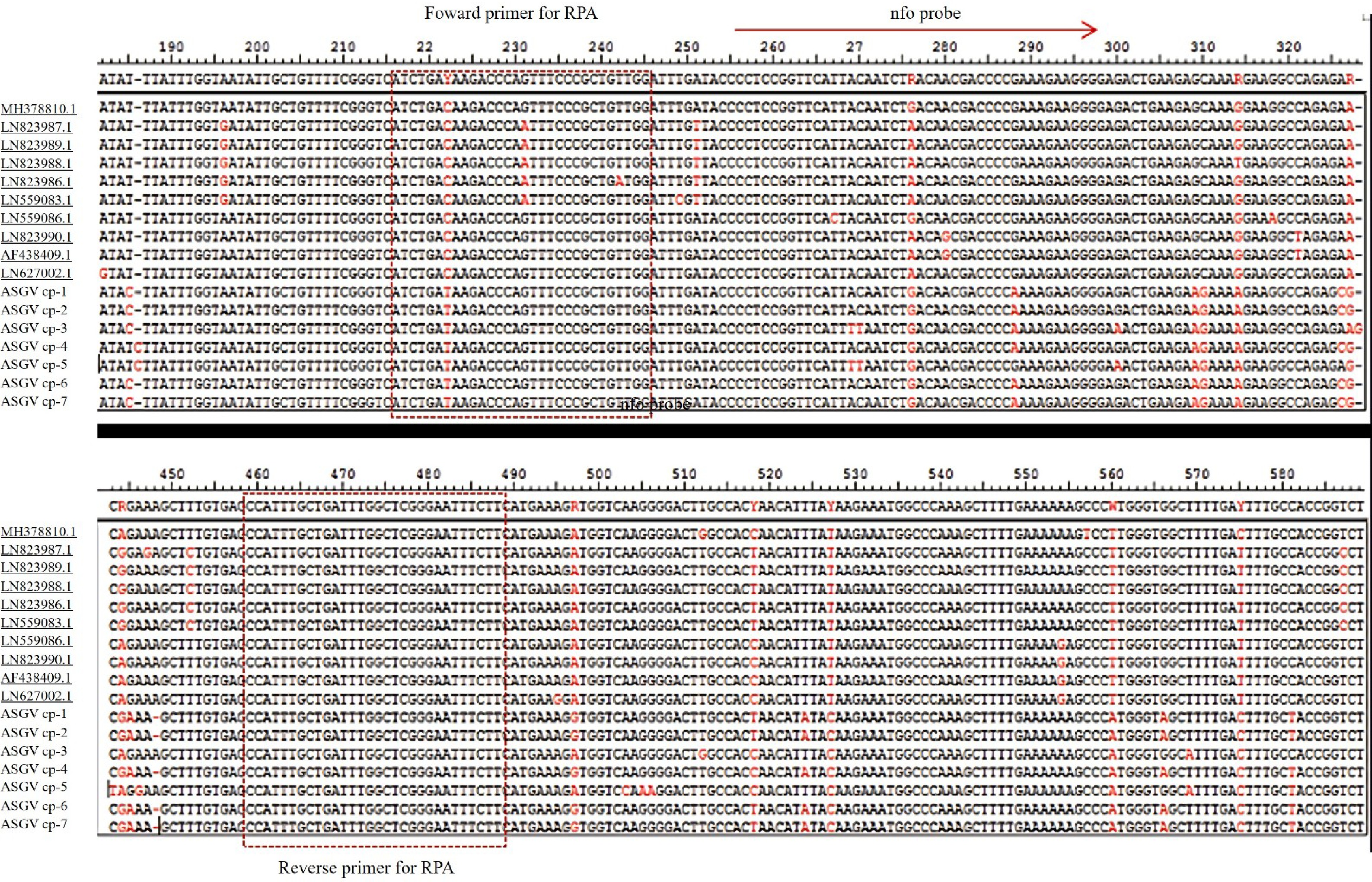
Figure 6.
Primer and probe design position within the coat protein of ASGV.
Sensitivity is also an important factor affecting the usability of pathogen detection methods. In this study, we found that RT-PCR detected the ASGV amplicon diluted samples to 10−4 whereas RT-RPA could detect the ASGV amplicon diluted samples to 10−6 (Fig. 4). One-step RPA detection is 100 times more sensitive than RT-PCR. Kim et al. reported that the level of sensitivity of the RT-RPA assay for ASPV detection was 1000-fold higher than that of the RT-PCR assay[17]. Qin et al. reported the sensitivity of the RPA-LFD assay for BCMV was 1000 times higher than the PCR assay[18]. RT-RPA is also more sensitive than RT-PCR in the detection of potato virus X and potato virus Y[21, 24, 34]. Our finding agrees with these studies highlighting the higher sensitivity of RT-RPA technique for virus detection.
Requires a high concentration of total RNA during the normal detection process, which makes all methods limited in the field detection process. Our study also showed that RNA crude extracts could be used in the detection of ASGV by RT-RPA-LFD with high level of specificity. Our finding agrees with this study with fast, sensitive, and robust direct RT-RPA assay of potyviruses[22, 35]. In the RT-PCR method for virus eradication, RNA extraction and purification are often required before cDNA synthesis. All these procedures should be performed in the laboratory, thus inhibiting the broad use of virus detection to manage viral diseases. Therefore, the one-step RT-RPA-LFD established in this study can overcome these constraints and enable the rapid on-site diagnosis of ASGV in the field.
-
Crude RNA extracts can be used for the rapid detection of ASGV using one-step RT-RPA-LFD, and the detection can be completed within 1 h with an incubator set at 40 °C.
-
In this study, the ASGV-infected in vitro apple of 'Gala', which had been obtained by Wang et al.[23], the three-infected (ASGV, ASPV and ACLSV) 'Yanfu 8' derived from the Laboratory of Fruit Tree Developmental Biology for Northwest Arid Area. All in vitro stock cultures were maintained on the Subculture and Proliferation Medium (SPM), including: Murashige and Skoog (1962)[36] medium (MS) 4.43 g/L, sucrose 30 g/L, 6-BA 0.4 mg/L, IBA 0.01 mg/L and agar 8 g/L, the pH of the medium was adjusted to 5.8 with NaOH prior to autoclaving at 121 °C for 20 min. Subculture were done every month, and stored at a temperature of 22 ± 2 °C with a photoperiod of 16 h and a light intensity of 50 μmol/m2/s. The light source was provided by a white fluorescent tube. The field samples of 'Gala' were collected at the Yangling Subsidiary Center of the National Apple Improvement Center in Shaanxi, China (Yangling, Shaanxi; 34°31′N, 108°05′E).
RNA isolation and virus detection by RT-PCR
-
Total RNA were extracted from fresh in leaf samples (0.5 g) and the Spectrum Plant Total RNA Kit (Sigma-Aldrich, USA) was used to obtain total RNA with the genome removed. cDNA was performed in the following reaction: 2 μL total RNA (500 ng/μL), 2.5 μL of DEPC water, 0.5 μL of Primer-R (10 μM), incubated for 5 min in a 70 °C water bath; 1.75 μL of DEPC water, 2.5 μL dNTPs (10 mM), 2.5 μL M-MLV 5 × Reaction buffer, 0.5 μL M-MLV reverse transcriptase and 0.25 μL RNase inhibitor, incubated at 42 °C for 60 min, kept at 95 °C for 5 min, the reaction was terminated on ice.
PCR operation reaction included 2 μL cDNA template, 12.5 μL 2 × Taq PCR Mix (GS101, Innovagene Biotech, China), 1 μL Primer-F (10 μM), 1 μL Primer-R (10 μM), 8.5 μL DEPC water. The sequences of all PCR detection primers in this study are shown in Table 2. The PCR reaction amplification was as follows: 94 °C for 3 min; followed by repeat for 35 cycles of 94 °C for 30 s, 56 °C for 45 s, and 72 °C for 50 s; finally, an extension at 72 °C for 10 min, ending the reaction and stock at 4 °C. The PCR products were electrophoresed on 2.0% agarose gel (w/v) containing DuRed dye (A168, Biomiga, China), and then photographed and recorded under ultraviolet light.
Table 2. The primer used for RT-PCR in apple virus detection.
Gene name Primer sequence Length (bp) Reference ASGV F: CCCGCTGTTGGATTTGATACACCTC 524 [37] R: CTGCAAGACCGCGACCAAGTTT ASPV F: ATGTCTGGAACCTCATGCTGCAA 370 [38] R: TTGGGATCAACTTTACTAAAAAG
CATAAACLSV F: GAGARTTTCAGTTTGCTMGA 794 [39] R: AGTCTACAGGCTATTTATTATAAGT nfo-ASGV F: ATCTGATAAGACCCAGTTTCCCGCT
GTTGG264 This study R: Biotin-AAGAAATTCCCGAGCCAAAT
CAGCAAATGGP: FAM-GGTTCATTACAATCTGACAA
CGACCCCAAAA(THF)GAAGGGGAGACTGAAG-C3 SpacerDetection by RT-RPA-LFD
-
Using DNAMAN to compare the ASGV coat protein sequences (MH378810.1, LN823987.1, LN823989.1, LN823988.1, LN823986.1, LN559083.1, LN559086.1, LN823990.1, AF438409.1 and LN627002.1) from NCBI to elucidate the conserved region of the virus genome, the ASGV special sequence was cloned by design primers in the conserved region (Fig. 6). The RT-RPA primers and probes are shown in Table 2. The RT-RPA amplification reaction of the TwistAmp® nfo kit (INTANFO, TwistDK, UK) is as follows: 2.1 μL Primer-F (10 μM), 2.1 μL Primer-R (10 μM), 1.2 μL RPA-Probe (10 μM), 29.5 μL Rehydration Buffer, 2 μL M-MLV reverse transcriptase (10 mM), 1 μL RNA template and 9.6 μL ddH2O to 47.5 μL, vortex gently; add the above mixture into the RPA reaction tube which contains the solid product after vacuum freeze-drying, pipette and mix well to rehydrate the lyophilized solid reactant; then add 2.5 μL 280 mM/L MgOAc to start reaction. The RT-RPA reaction amplification for 30 min at 40 °C, and then the RT-RPA product was tested by LFD (MILENIA01, TwistDK, UK). The coat protein of ASGV amplified by GenBank and PCR was sequenced in Seqman; the unfilled box represents the primer region, and the arrow represents the position of the probe used in this study.
Determination of RT-RPA-LFD conditions
-
The RPA reaction temperature was set to 40 °C, and the incubation time was set to 0, 10, 20, 30, and 40 min respectively, based on the optimally set reaction temperatures of 37 °C, 38 °C, 39 °C, 40 °C, and 41 °C. The optimal reaction time and temperature were determined by agarose gel electrophoresis and LFD RPA product analysis.
Specificity and sensitivity of RT-RPA-LFD
-
Testing of the specifics for one-step RT-RPA-LFD assays was conducted, the primer and probe designed in conserved region from coat protein of ASGV, using single ACLSV-infected, ASPV-infected, and ASGV-infected in vitro as the materials, the virus-free 'Gala' as the negative control, and the infected ASGV, ASPV and ACLSV in vitro 'Yanfu 8' as a positive control. For the sensitivity test of one-step RT-RPA-LFD, dilute total RNA (500 ng/μL) with a 10-fold gradient (10−1 to 10−8) as a template for amplification one-step RT-RPA-LFD detection. RNA samples are also used for RT-PCR detection, and 1.5% agarose gel electrophoresis analysis RT-PCR product.
Rapid detection of ASGV by crude RNA
-
Apple leaf (13 mg) were immersed in 300 μL of freshly prepared alkaline polyethylene glycol (PEG) buffer (6 % PEG 200, 20 mM NaOH) and ground, and incubated at 37 °C for < 5 min to obtain a crude extract[22, 35], which was directly used for the RT-RPA assay.
Data analysis and processing
-
Each experiment consisted of three groups, and each experiment was repeated twice.
This work was financially supported by Shaanxi Apple Industry Science and Technology Project (2020zdzx03-01-04), the Science and Technology Activity Fund for Returnees studying abroad of Shaanxi Province (2020-07), the Cyrus Tang Foundation, and the China Apple Research System (CARS-27).
-
The authors declare that they have no conflict of interest. Dong Zhang is the Editorial Board member of Journal Fruit Research. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of the Editorial Board member and his research groups.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Ma X, Bao J, Li J, Cheng X, Tahir MM, et al. 2023. Rapid detection of apple stem grooving virus from reverse transcription recombinase polymerase amplification with a lateral flow dipstick (RT-RPA-LFD). Fruit Research 3:22 doi: 10.48130/FruRes-2023-0022 

Rapid detection of apple stem grooving virus from reverse transcription recombinase polymerase amplification with a lateral flow dipstick (RT-RPA-LFD)
- Received: 21 April 2023
- Accepted: 28 July 2023
- Published online: 05 September 2023
Abstract: Apple stem grooving virus (ASGV) is one of the most widespread and asymptomatic main viruses, that restricts the production of apples worldwide. Establishment of rapid, simple, and effective early detection methods of apple virus is important. In this study, we established and optimized a one-step reverse transcription - recombinase polymerase amplification (RT-RPA) method, using the target-specific primers of ASGV coat protein gene sequence, and M-MLV reverse transcriptase. This method could be completed within 30 min at 40 °C, followed by a visual detection of the results within 5 min by using lateral flow dipstick (LFD). The specificity results showed that only samples infected with ASGV showed a test line, while no test line appeared in the ASGV-negative samples. In addition, when crude extract of leaves was used, the whole detection could be completed within 1 h, which was shortened by 4 to 6 times compared with the RT-PCR method. The detection made on more field samples showed that the RT-RPA-LFD method is of high stability and reliability for ASGV diagnosis, with a great potential in the rapid on-site detection of plant viruses.