








-
Nitrogen is a major component of chlorophyll, nucleic acids, proteins, vitamins, and several other secondary metabolites in plants, which plays an irreplaceable role in the regulation of plant photosynthesis, carbohydrate allocation, root formation, and resistance to abiotic stress[1]. Previous studies have demonstrated that the application of an optimal amount of nitrogen (N) fertilizer can enhance the electron transfer capacity between photosystem I (PSI) and photosystem II (PSII) in wheat, which leads to improved photosynthetic efficiency, higher biomass production, and increased crop yield[2]. However, N deficiency stress is a common nutrient stress in plants. Under conditions of mild nitrogen deficiency stress, plants usually exhibit slow growth and pale or yellow-green leaves, while severe nitrogen deficiency stress can inhibit plant photosynthesis and accelerate leaf senescence, ultimately affecting plant yield and quality. Given the potential negative impact of nitrogen deficiency stress on plant growth and development, understanding the molecular mechanisms underlying plant tolerance to this stress has become an urgent challenge in current research.
Apple (Malus domestica Borkh) is one of the most widely cultivated and important commercial crops worldwide. In 2020, the total output of apples reached 46 metric tons in China, which accounted for over 55% of global apple production (FAOSTAT, 2022,
http://www.fao.org/faostat/en/#home ). However, the quality and yield of apples are usually affected by unfavorable environmental conditions, such as N-deficiency. Studies have shown that in response to nitrogen deficiency stress, the color of apple leaves change from dark green to light yellow-green[3]. There is a also a significant increase in the levels of malondialdehyde (MDA), soluble protein, and proline content, while the chlorophyll content significantly decreases. These changes are adaptive responses of the plant to nitrogen deficiency stress[3]. At the same time, the molecular regulation mechanism of apple trees in response to nitrogen deficiency stress conditions has also been discovered. Many genes involved in this process have been identified, including Apple MdBT2 (BTB and TAZ DOMAIN PROTEIN 2), which was repressed by nitrate starvation. Overexpression of MdBT2 negatively regulates apple response to tolerate N deficiency stress[4]. MdMYB10, an important MYB (v-Myb avian myeloblastosis viral oncogene homolog) transcription factor in apples, was also involved in regulating nitrate uptake and usage. Overexpression of the MdMYB10 gene can enhance the absorption of nitrate by apple roots, promote the transport and redistribution of nitrogen in the plant, thus increasing the nitrogen content and accumulation of anthocyanins in apple fruits[5]. However, research on apple N deficiency stress response is still insufficient. Understanding the functions of the genes involved in this process will be crucial for developing strategies to improve the growth and productivity of apple trees under conditions of N deficiency.SUMOylation is a crucial post-translation modification (PTM) that is the covalent attachment of small ubiquitin-associated modifier (SUMO) molecules to target proteins catalyzed by enzyme cascade, which regulates numerous cellular and physiological processes of plants, particularly in response to abiotic stress[6,7]. Currently, SUMOylation has been proven to involved in the physiological processes of many plants, including Arabidopsis, rice, soybean, maize, potato, tomato, peanut, apple, poplar, and so on[6,8−10]. In Arabidopsis, the SUMO E3 ligase SIZ1 (SAP and MIZ1 protein interacting zinc finger protein 1) functions in abiotic stress responses, phytohormone signaling, nitrogen assimilation, development, and photomorphogenesis[11]. Additionally, rice OsSIZ1 is involved in the phosphate and nitrogen-dependent responses[12]. Apple MdSIZ1 is induced by the transcription factor MdMYB2, which has been found to regulate cold tolerance and anthocyanin accumulation[13]. However, to date, how SUMOylation participates in the underlying N deficiency stress response in apple trees is largely unknown.
In this study, we found that interfering SUMOs expression could facilitate tolerance of the transgenic apple plants to N deficiency stress conditions. The MdSUMO2 RNAi transgenic apple plants exhibited better growth phenotype under N deficient conditions compared to the wild type, as well as the developed root system. The activated nitrate reductase NR and nitrate transporters NRT1.1/1.2/2.5 in the root system of MdSUMO2 RNAi plants might partly contribute to the tolerance of MdSUMO2 RNAi in response to N deficiency stress conditions. Our findings highlight the important role of apple MdSUMOs during N deficiency response and provide a foundation for improving the efficiency of nitrogen use in apple.
-
It was reported that apple small ubiquitin modifiers (SUMO) - MdSUMO2A, MdSUMO2B, and MdSUMO2C were induced by various abiotic stress[6,7]. To understand the expression pattern of apple MdSUMO2s in response to N deficient conditions, we examined the relative expression levels of MdSUMO2A/B/C by qRT-PCR (qRT-PCR). The results showed that MdSUMO2A, MdSUMO2B, and MdSUMO2C were all induced by N deficiency stress conditions (Fig. 1), indicating that apple MdSUMO2A/B/C were N deficiency responsive genes.
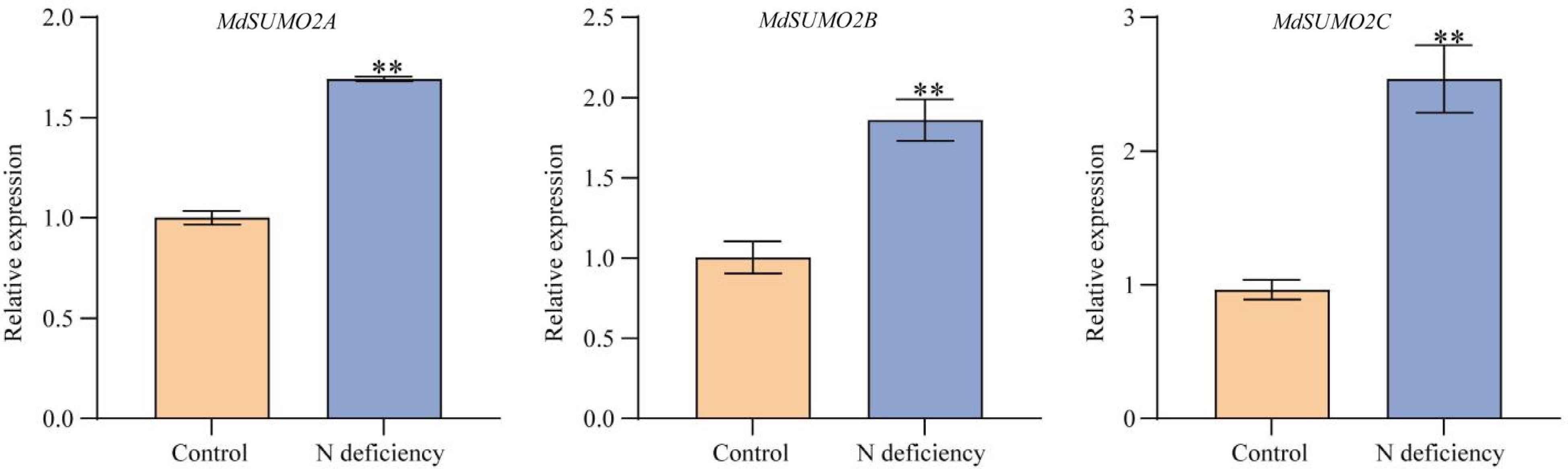
Figure 1.
Expression patterns of MdSUMO2s in apple under N deficient conditions. Hydroponically-cultured GL-3 were treated with normal (6 mM) or low nitrate (0.2 mM) for 4 d. Root samples were collected and used for gene expression analysis. Error bars indicate SD, n = 3. Asterisks indicate statistically significant differences. Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
MdSUMO2 RNAi transgenic apple plants exhibit enhanced tolerance to N deficiency
-
The induced expression levels of MdSUMO2A, MdSUMO2B, and MdSUMO2C suggested these genes might function in the N deficiency stress response. To examine this hypothesis, MdSUMO2 RNAi transgenic apple seedlings which interfering all SUMOs expression and the wild type (GL-3) were used as experimental materials. Under normal hydroponic culture conditions, no significant difference was observed between the GL-3 and MdSUMO2 RNAi plants. However, under N deficient treatment conditions, MdSUMO2 RNAi transgenic apple plants exhibited a more vigorous phenotype than GL-3 (Fig. 2a). A significant reduction of new leaves was found in GL-3 under N deficient conditions, but the MdSUMO2 RNAi transgenic apple plants were less affected (Fig. 2b). These results showed that GL-3 ceased to produce new leaves under N deficiency stress, while the growth of MdSUMO2 RNAi transgenic apple seedlings was not significantly affected (Fig. 2c & e). Under nitrogen deficient conditions, the MdSUMO2 RNAi apple plants exhibited significantly increased height and fresh weight in comparison to the GL-3 plants, which was the opposite of the control conditions (Fig. 2c & e). All apple seedlings displayed a noticeable reduction in stem diameter when exposed to nitrogen deficiency, suggesting a general impact on stem development under limited nitrogen availability (Fig. 2d). Interestingly, in contrast to the control conditions, the MdSUMO2 RNAi plants demonstrated a strikingly thicker stem than the GL-3 plants under nitrogen deficiency (Fig. 2d). Together, these findings indicate that knock-down of the MdSUMO2s in apple enhanced the tolerance to N deficiency.
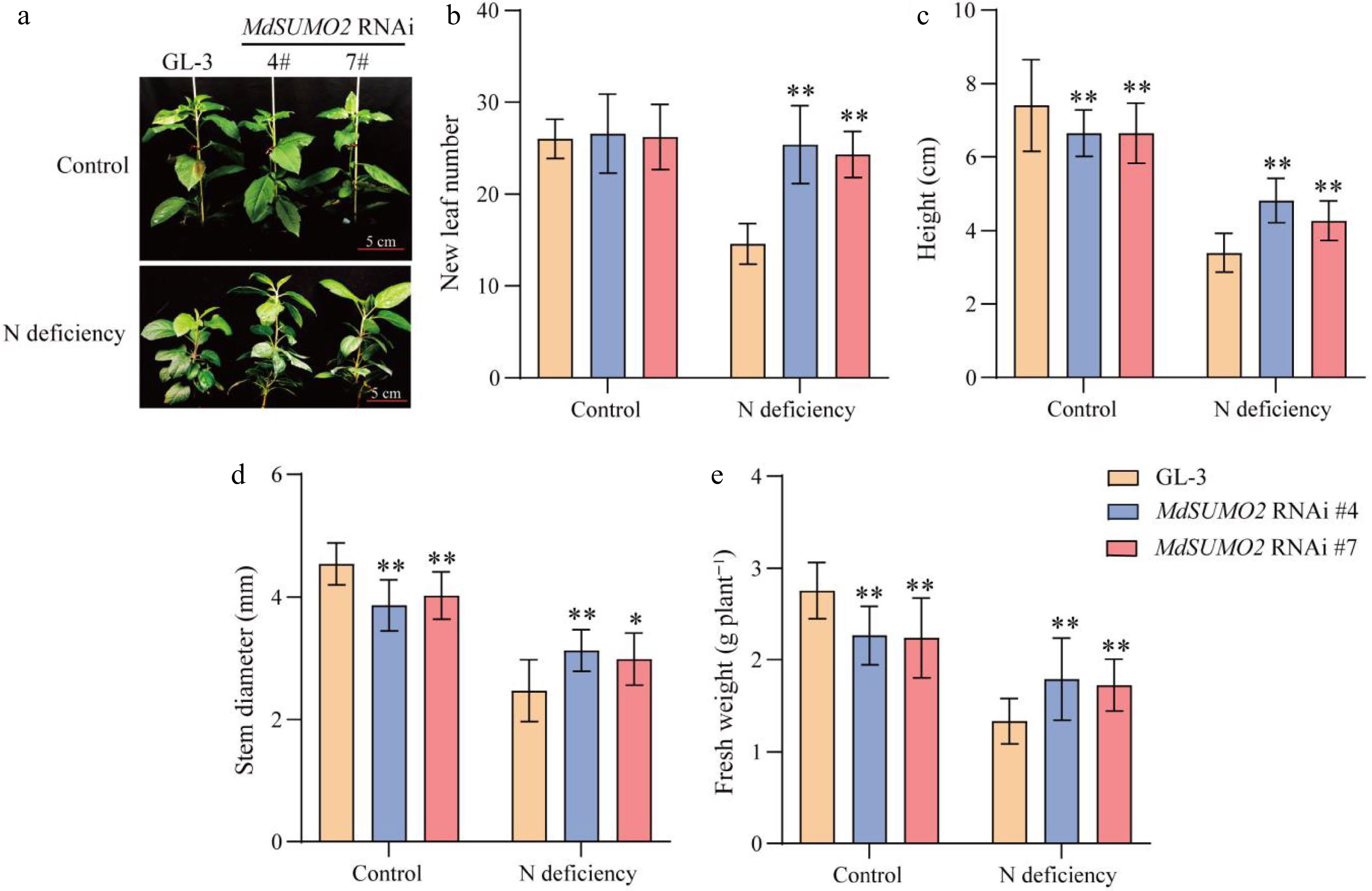
Figure 2.
The MdSUMO2 RNAi transgenic apple plants are more tolerance than the wild type (GL-3) under N deficient conditions. (a) Morphology of three-monthold MdSUMO2 RNAi transgenic apple plants and GL-3 treated with normal nitrogen (Control, 10 mM KNO3, and low nitrogen (N deficient, 0.2 mM KNO3) under hydroponic culture. Scale bars = 5 cm. (b) New leaf number, (c) height, (d) stem diameter, (e) fresh weight in (a). Error bars indicate SD, n = 12 in (b)−(e). Asterisks indicate significant differences between MdSUMO2 RNAi transgenic apple plants and GL-3 plants in each group (control and N deficiency). Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
MdSUMO2 RNAi transgenic apple plants increased more chlorophyll content in response to N deficient conditions
-
Nitrogen deficient conditions typically result in yellowing of the leaves[14]. Here, we found that the MdSUMO2 RNAi transgenic apple plants tended to keep green leaves compared to GL-3 under N deficient conditions (Fig. 2a). Since the SPAD value has been commonly used as an indicator of nitrogen levels in plants[15], we evaluated the leaves SPAD value of MdSUMO2 RNAi transgenic apple plants and GL-3 in response to N deficiency stress conditions. The results showed that the SPAD values of all the plants decreased significantly upon N deficient conditions, and the MdSUMO2 RNAi plants kept higher SPAD than GL-3 after N deficiency stress treatment (Fig. 3a). We investigated the chlorophyll content of MdSUMO2 RNAi plants and GL-3 in response to N deficiency stress. The results showed that the content of chlorophyll a, chlorophyll b, and total chlorophyll significantly decreased in leaves under N deficiency stress conditions (Fig. 3b, c & d). However, in the presence of N deficiency stress conditions, all the chlorophyll content in MdSUMO2 RNAi transgenic apple plants were higher than in GL-3.
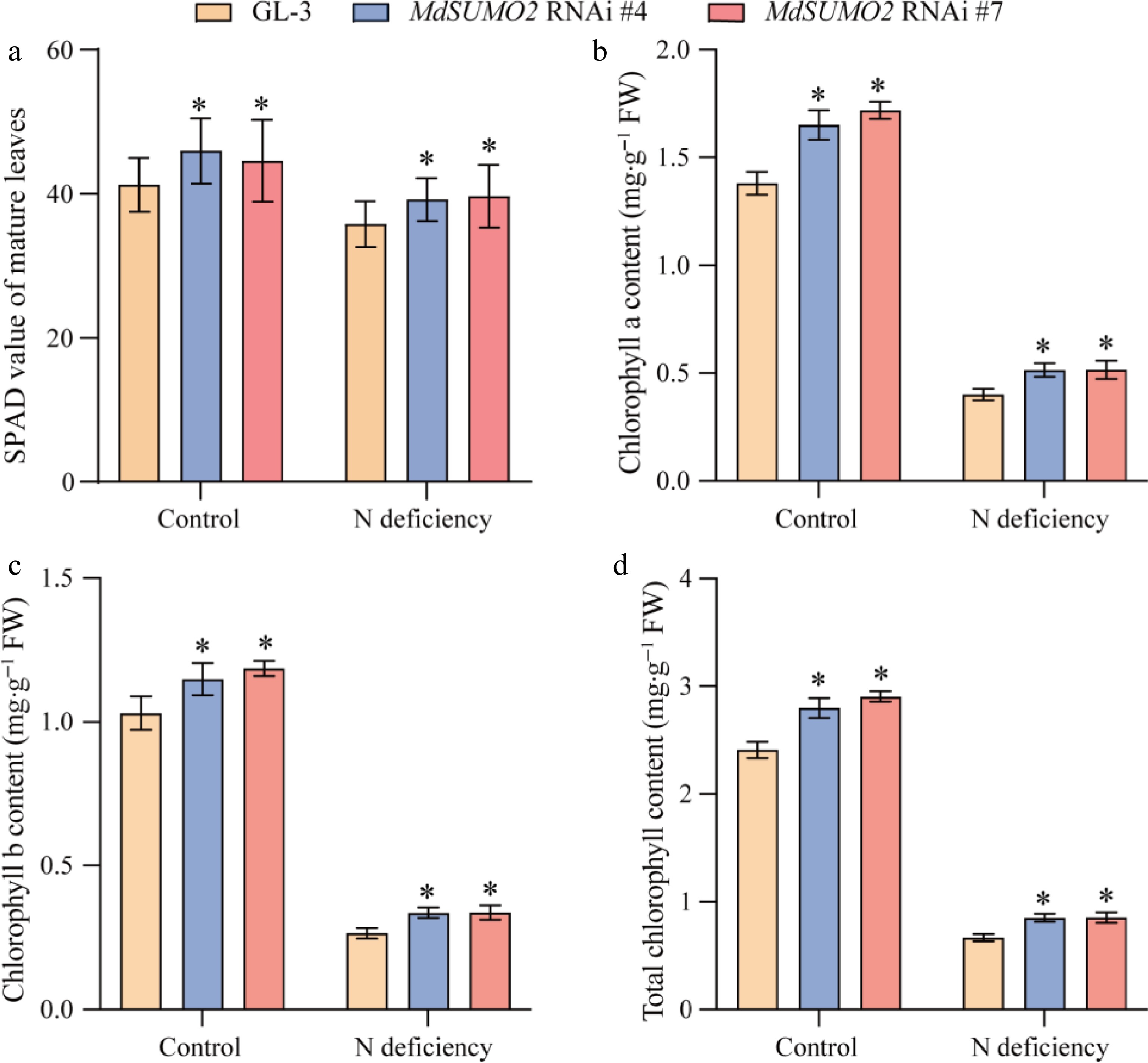
Figure 3.
The MdSUMO2 RNAi transgenic apple plants displayed darker green leaves than the wild type (GL-3) under N deficient conditions. (a) The SPAD value of leaves in response to N deficiency in MdSUMO2 RNAi transgenic apple plants and GL-3. (b) Chlorophyll a, (b) chlorophyll b, and (c) total chlorophyll content of leaves in MdSUMO2 RNAi transgenic apple plants and GL-3 under N deficient conditions. Error bars indicate SD, n = 12 in (a)−(d). Asterisks indicate significant differences between MdSUMO2 RNAi transgenic apple plants and GL-3 plants in each group (control and N deficiency). Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
MdSUMO2 regulates root system conformation in response to N deficient conditions
-
The roots of apple trees serve as the primary organs for nutrient absorption, including nitrogen uptake[16]. We observed the root systems of the MdSUMO2 RNAi transgenic apple plants and GL-3. As can be seen from the root morphology results (Fig. 4a), N deficient conditions induced the root system development of both GL-3 and the MdSUMO2 RNAi transgenic apple plants (Fig. 4b, c & d). And the MdSUMO2 RNAi plants showed significantly greater average diameter and larger root volume than GL-3 in response to N deficient conditions (Fig. 4c & d). These results suggested that the improved root system of MdSUMO2 RNAi transgenic apple plants may be responsible for their increased tolerance to nitrogen deficiency.
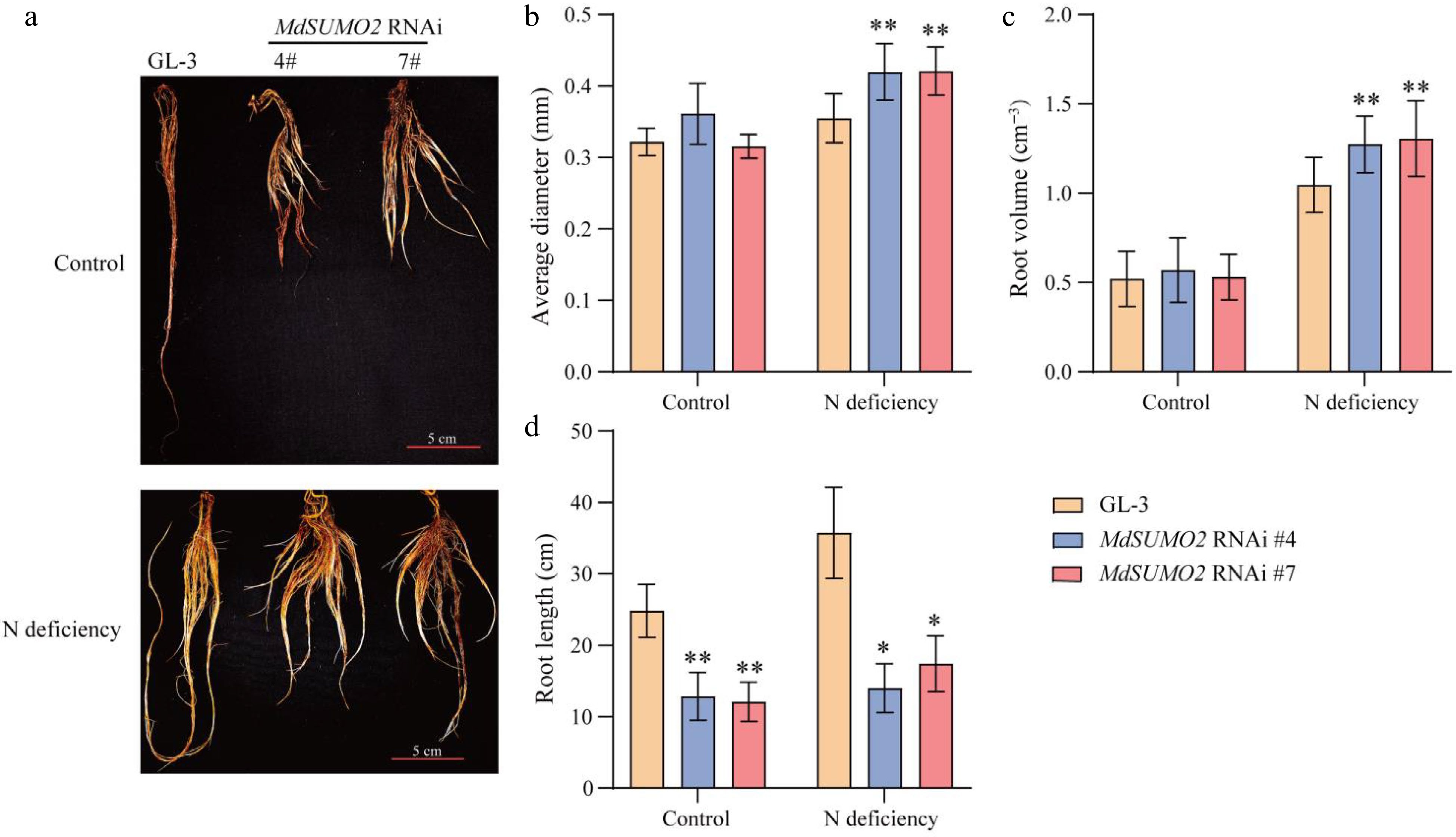
Figure 4.
MdSUMO2 regulates root system in response to N deficient conditions. (a) Morphological characteristics of the root system in three-month-old MdSUMO2 RNAi transgenic apple plants and GL-3 treated with normal nitrogen (Control, 10 mM KNO3), and low nitrogen (N deficiency, 0.2 mM KNO3) under hydroponic culture. Scale bars = 5 cm. (b) Average diameter, (c) root volume, and (d) root length in (a). Error bars indicate SD, n = 12 in (b)−(d). Asterisks indicate significant differences between MdSUMO2 RNAi transgenic apple plants and GL-3 plants in each group (control and N deficiency). Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
Activated glutamine synthetase and nitrate reductase contribute to the tolerance of MdSUMO2 RNAi transgenic apple plants in response to N deficient conditions
-
According to previous studies, both glutamine synthetase (GS) and nitrate reductase (NR) are known as the principal enzymes involved in plant nitrogen conversion[17]. To further explore whether MdSUMOs function under N deficiency stress conditions in apple, the GS and NR activity were determined in roots of MdSUMO2 RNAi transgenic apple plants and GL-3 under control and N deficient conditions. Under control conditions, the MdSUMO2 RNAi transgenic apple plants showed higher GS activity than GL-3 in the root system. The N deficiency treatment produced more significant results—the GS activity markedly increased more in MdSUMO2 RNAi plants than GL-3 (Fig. 5a). With regards to the NR activity, MdSUMO2 RNAi transgenic apple plants were higher than GL-3 under N deficient conditions. Nevertheless, the NR activity was not significantly affected by interfering MdSUMO2 expression in the control (Fig. 5b).
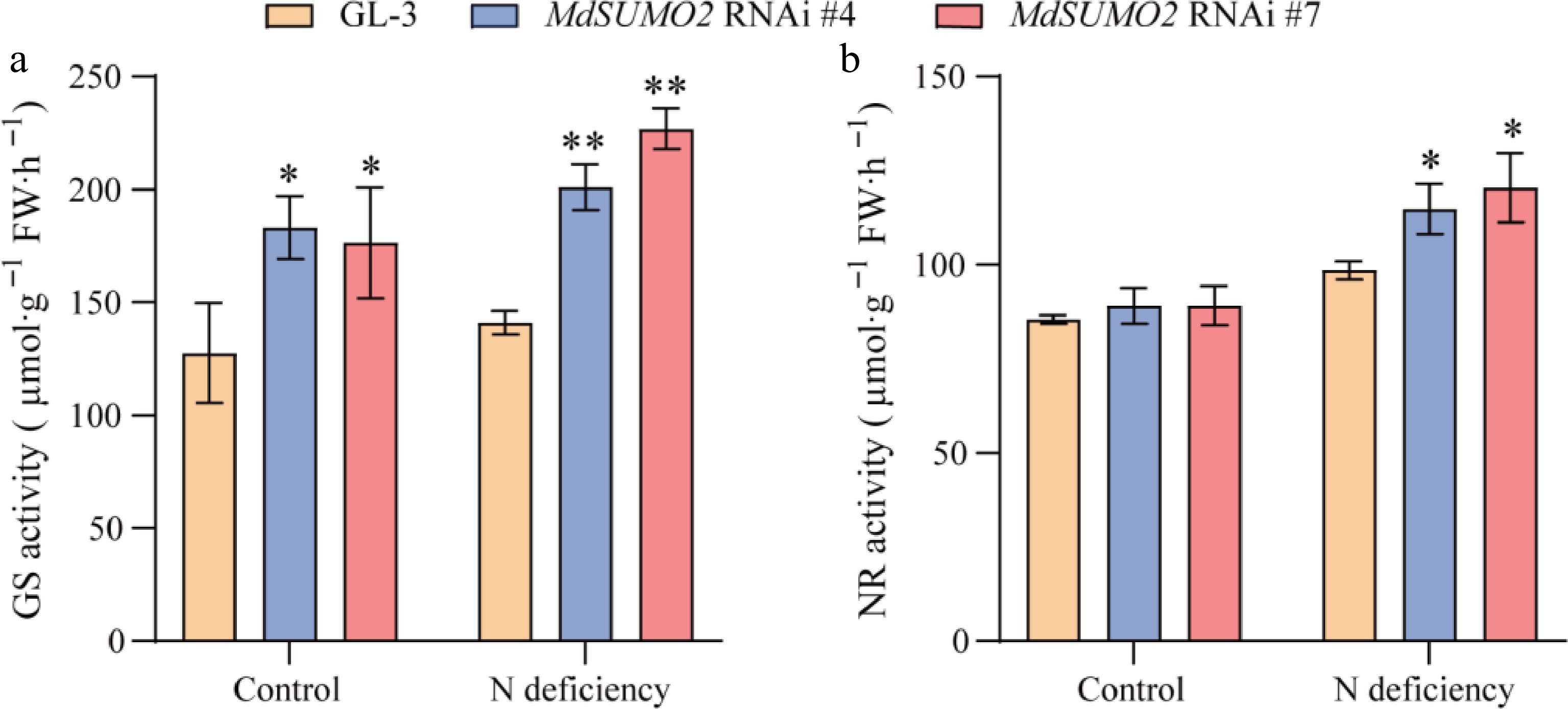
Figure 5.
Activated glutamine synthetase and nitrate reductase in root contribute to the tolerance of MdSUMO2 RNAi transgenic apple plants in response to N deficiency. (a) Glutamine synthetase (GS) activity in MdSUMO2 RNAi transgenic apple plants and GL-3 under control and N deficient conditions. (b) Nitrate reductase (NR) activity in MdSUMO2 RNAi transgenic apple plants and GL-3 under control and N deficient conditions. Error bars indicate SD, n = 3 in (a) and (b). Asterisks indicate significant differences between MdSUMO2 RNAi transgenic apple plants and GL-3 plants in each group (control and N deficiency). Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
The genes responsible for nitrogen assimilation play roles in the tolerance of MdSUMO2 RNAi transgenic apple plants
-
Nitrate uptake and distribution is carried out by nitrate transporters (NRTs)[18]. To determine the mechanism behind the increased tolerance of MdSUMO2 RNAi transgenic apple plants under nitrogen deficient conditions, we further detected the expression levels of genes involved in nitrogen uptake. According to our results, MdNRT1.1, MdNRT1.2 were inhibited in response to N deficiency stress, while MdNRT2.5 is significantly induced and up-regulated. Additionally, the expression levels of MdNRT1.1, MdNRT1.2 and MdNRT2.5 were all higher in MdSUMO2 RNAi transgenic plants than GL-3 under control and N deficient conditions (Fig. 6a−c). These results indicate that MdSUMO2 RNAi transgenic plants have higher levels of NRT gene expression compared to GL-3.
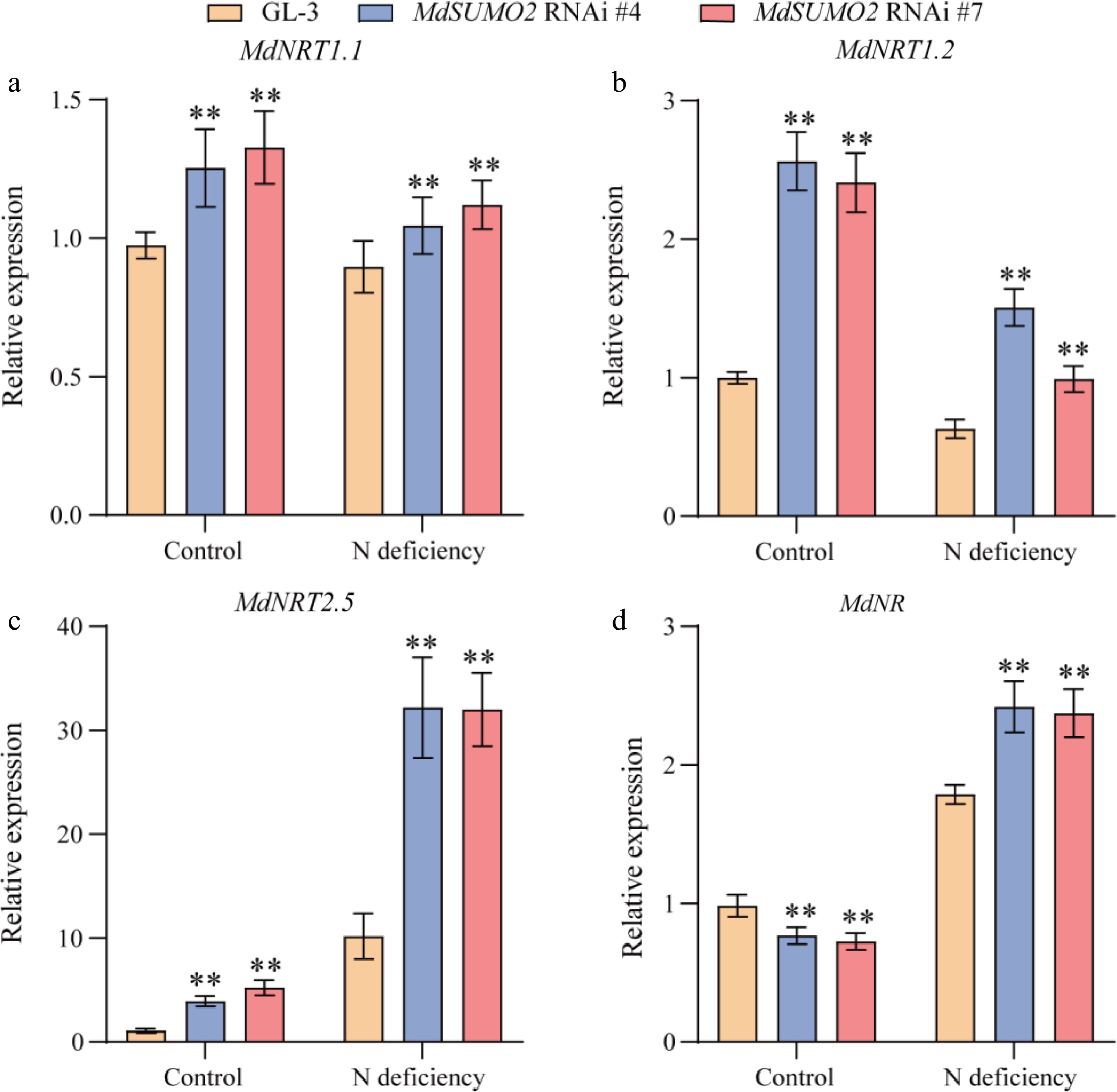
Figure 6.
Genes involved in nitrogen assimilation show a response in the MdSUMO2 RNAi transgenic apple plants under N deficient conditions. Relative expression levels of (a) MdNRT1.1, (b) MdNRT1.2, (c) MdNRT2.5 and (d) MdNR in the root under control and N deficient conditions. Error bars indicate SD, n = 3 in (a)−(d). Asterisks indicate significant differences between MdSUMO2 RNAi transgenic apple plants and GL-3 plants in each group (control and N deficient). Student's t test was performed and statistically significant differences were indicated by *p ≤ 0.05, **p ≤ 0.01.
Nitrate reductase genes (NRs) has been shown to play a pivotal role in nitrogen assimilation and stress tolerance in plants[19]. Thus, the expression levels of the MdNR in MdSUMO2 RNAi transgenic apple plants and GL-3 was examined using qRT-PCR experiment. The result showed that the expression of MdNR was significantly lower in MdSUMO2 RNAi transgenic plants under control conditions compared to the GL-3 (Fig. 6d). N deficient conditions induced the expression of MdNR significantly, and much higher in MdSUMO2 RNAi transgenic plants than GL-3. These results demonstrated that the induced expression of genes involved in nitrogen uptake and assimilation in MdSUMO2 RNAi transgenic plants might contribute to its tolerance to N deficiency stress.
-
The growth and development of plants require various nutrients[20]. Among all necessary nutrients, nitrogen is the primary limiting factor for plant growth and yield formation on a global scale[20]. The application of nitrogen fertilizer is crucial for improving agricultural yields and enhancing crop quality. However, the long-term excessive use of fertilizers has increasingly caused environmental consequences[21]. How to improve the utilization efficiency of nitrogen fertilizer has always been a hot scientific issue in agricultural research. Understanding the molecular mechanisms related to plant perception and response to nitrogen availability can be utilized to improve nitrogen utilization efficiency. While the knowledge of the molecular mechanisms behind improving nitrogen utilization efficiency in model plants has greatly expanded, more extensive research is still needed in woody fruit trees. In this study, we identified that MdSUMO2s were up-regulated in response to N deficiency (Fig. 1), indicating that apple MdSUMO2s might function in the N deficiency stress response. Meanwhile, we found that MdSUMO2 RNAi transgenic apple plants exhibited a more vigorous phenotype compared to GL-3 (Fig. 2a). Under low nitrogen conditions, plants adapt to the limitation by making physiological and morphological adjustments, including new leaf number, height, stem diameter and fresh weight[22]. One such adjustment is increasing leaf production to enhance leaf area for improved nitrogen absorption and utilization efficiency[23]. Due to their higher nitrogen use efficiency and effective utilization of limited nitrogen resources, plants under low nitrogen conditions exhibit a tendency to decelerate the growth of older leaves while allocating increased resources towards the growth of new leaves[24]. By measuring the number of new leaves, we can assess a plant's ability to adapt to low-nitrogen environments and its growth status. Additionally, measuring the number of new leaves provides information about plant growth rate and physiological condition[25]. Changes in the number of new leaves under low-nitrogen treatment reflect both a plant's response level to nitrogen nutrition as well as the extent to which its growth is restricted. Three-month-old MdSUMO2 RNAi transgenic apple plants produce more new leaves compared to GL-3 under N deficiency stress (Fig. 2b). Additionally, Under N deficient conditions, the MdSUMO2 RNAi apple plants showed lower height, stem diameter and fresh weight compared to GL-3 plants (Fig. 2b−e). All these findings indicate that knock-down of the MdSUMO2s in apple enhances the N deficiency tolerance.
When crops are deficient in nitrogen, protein synthesis is inhibited, leading to a decrease in the amount of protein and enzymes in the crop[26]. The structure of chloroplasts is also damaged due to nitrogen deficiency, and the synthesis of chlorophyll decreases, causing the leaves to turn yellow[27]. These changes slow down the growth process of the plant, which in turn severely affects crop yield and quality. In this study, we investigated the color of apple leaves. The results showed that MdSUMO2 RNAi transgenic apple plants exhibited higher chlorophyll content compared to GL-3 (Fig. 3b−d). Relevant studies have shown that SPAD can be used to accurately predict the chlorophyll content and nitrogen concentration of various crop leaves, and has been applied in the low-nitrogen tolerance evaluation of crops such as rice and corn[15]. Our results showed higher SPAD value in MdSUMO2 RNAi transgenic apple plants than GL-3 (Fig. 2a). So, we speculated the darker green leaves of MdSUMO2 RNAi transgenic apple plants may help assimilate more energy and make the plants more vigorous.
The morphology and physiological state of the root system are closely related to the growth status of the aboveground part, which is crucial for both the movement of nutrients within plants and the absorption of nutrients by plants[28]. Plants can better absorb and store nitrogen when their root systems are properly arranged and distributed, and the availability of nitrogen also has an impact on how well plant roots grow[29]. Therefore, we analyze the root system of MdSUMO2 RNAi transgenic apple plants and GL-3. Although the GL-3 had significantly longer total root lengths compared to MdSUMO2 RNAi transgenic apple plants, the average diameter and root volume were significantly higher in the MdSUMO2 RNAi than in GL-3 plants under N deficiency stress (Fig. 4a−d). The root volume is crucial for nutrient absorption of plants, especially by influencing the root density and root length density to affect their nutrient uptake[30]. So, the larger root volume of MdSUMO2 RNAi might be an important factor in enabling them to perform better under N deficiency. Meanwhile, in plant roots, GS and NR play important roles in the uptake and assimilation of nitrogen from the soil[20]. GS is mainly located in the root cortex, where it assimilates ammonium derived from nitrate reduction or directly absorbed by the roots[26]. NR is mainly located in the root epidermis and cortex, where it reduces nitrate to nitrite[31]. Our results showed higher GS activity and NR activity in MdSUMO2 RNAi transgenic apple plants than GL-3 (Fig. 5a,b). In coincide, the expression of genes involved in nitrogen assimilation in the roots were higher in MdSUMO2 RNAi transgenic apple plants than GL-3 (Fig. 6a−e). Therefore, we speculated that the root system of MdSUMO2 RNAi transgenic apple plants had stronger nitrogen assimilation capabilities, which made their growth less affected under N deficient conditions.
In conclusion, the results of this study showed that interfering small ubiquitin modifiers (SUMO) promoted plant growth and biomass accumulation under N deficient conditions. Meanwhile, MdSUMO2 RNAi transgenic apple plants produced darker green leaves and larger root volume than GL-3 in response to N deficiency stress. In addition, the GS activity, NR activity, and the expression of genes involved in nitrogen assimilation in the root were higher in MdSUMO2 RNAi transgenic apple plants than GL-3. These findings provide theoretical guidance for revealing the mechanism by which MdSUMO2 regulates nitrate uptake and plant growth, as well as candidate genes for future apple breeding to improve nitrogen uptake efficiency.
-
The MdSUMO2 RNAi transgenic apple plants were produced previously[6]. The tissue-cultured apple seedlings were cultured in Murashige and Skoog (MS) medium (4.43 g/L MS salts, 30 g/L sucrose, 0.2 mg/L 6-Benzylaminopurine [6-BA], 0.2 mg/L 3-Indoleacetic acid [IAA] and 7.5 g/L agar, pH 5.8) under long-day conditions (14 h light [cool white, ~100 umol/m2/s, T5 LED batten]: 10 h dark) for 4 weeks at 25 °C[32]. One month-old tissue-cultured apple seedlings were selected for rooting, and rooted on 1/2 MS medium containing 0.5 mg/L IAA and 0.5 mg/L indole butyric acid (IBA). When the tissue-cultured apple seedlings were rooted, the plants were transformed to a nutrient bowl containing substrate (Pindstrup, Denmark), and cultured in a growth chamber (16 h light: 8 h dark, 25 °C, ~55% relative humanity) for 30 d.
RNA extraction and qRT-PCR analysis
-
RNA extraction was performed as described previously[33]. RNase-free DNase Ⅰ (Thermo Scientific, USA) was used to remove genomic DNA. After extracting total RNA, it was detected through agarose gel electrophoresis and UV spectrophotometer (Thermo Nanodrop 2000) for concentration determination. Then, 1 μg of RNA was reverse transcribed to synthesize cDNA by using reverse transcription kit HiScript II Q RT SuperMix for qPCR (+gDNA wiper) (Vazyme, R223-01, China). Quantitative reverse transcription PCR (qRT-PCR) analysis was carried out using a ChamQ Universal SYBR qPCR Master Mix (Vazyme, C601, China) on the CFX96 real-time system (Bio-Rad, USA). Primers used in this study are listed in Table 1.
Table 1. Sequences of primers used in this study.
Gene name Forward primer Reverse primer Purpose MdSUMO2C GCACAGAAGAAACCCCTGGAT CACTTCATTACCATCCTGCCCT qRT-PCR analysis MdSUMO2A GTGTTTGGAAGTGGTGGTAGCTC TCTTGTCCTCCTCCTGGTTCG qRT-PCR analysis MdSUMO2B CGACTCCGCAGCAAGAAGAG CTTCAGCTGAGTGCTTCGC qRT-PCR analysis MdSUMO2-RNAi GGGGACAAGTTTGTACAAAAAAGCAGGCTTC
TCAGGCGTCACGAACCAGGGGGGACCACTTTGTACAAGAAAGCTGGGTCGC
TGGGTACTTCTCTTGATCCTGAPlasmid construction of MdSUMO2 RNAi MdNRT1.1 ACGGGCACCACGCACTTG GGTTGGAGGCTGGGGACTG qRT-PCR analysis MdNRT1.2 TCCCTATCTTCCCCGTACTCTTC GGAGTTGGTCGCTACACGCT qRT-PCR analysis MdNRT2.5 CGTTCCACCTGTCTTGGTTTTC CGATGTCGCTGGCGGTTAG qRT-PCR analysis MdNR TGACGATAGCGTTGTCCAGG AGTGAGGCGGACCATTGAAG qRT-PCR analysis N deficiency treatment
-
N deficiency treatment was performed as described previously[4]. Hydroponically-cultured plants were cultured in 1/2 Hoagland nutrient solution (1.75 mM Ca(NO3)2, 2.5 mM KNO3, 0.5 mM KH2PO4, 0.86 mM MgSO4, 100 μM EDTA-FeNa, 46.26 μM H3PO4, 9.15 μM MnCl2, 0.77 μM ZnSO4, 0.32 μM CuSO4, 0.5 μM H2MoO4). Hydroponic culture was chosen for three-month-old GL-3 and MdSUMO2 RNAi transgenic apple plants with identical growth conditions. After pre-culturing for about 10 d, the plants were in nitrogen starvation for 7 d and then divided into two groups with low and normal nitrogen solution (0.2 mM and 6 mM KNO3) with the same base nutrient medium. The two groups continued to grow in the nutrient solution for 30 d and the solution was refreshed every 3 d. Hydroponically-cultured GL-3 (three-month-old) were treated with normal (6 mM) or low nitrate (0.2 mM) for 4 d. Root samples were used for gene expression analysis[34].
Growth and root architecture measurements
-
Twelve plants were measured for each genotype. The physiological indicators of the above-ground part, including new leaf number, height, stem diameter, and fresh weight. To obtain an accurate assessment of the root systems, the roots were meticulously cleansed without causing any damage. After flattening and spreading the roots, their images were captured using a SNAPSCAN 310 scanner, which allowed for high-resolution imaging. Finally, WinRHIZO image analysis software (V4.1c; Regent Instruments, Quebec City, QC, Canada) was used to examine root system parameters.
Measurements of leaf SPAD
-
For measurements of leaf SPAD, we used a portable chlorophyll meter (SPAD-502; Minolta, Japan) according to the method of Markwell et al[35]. The results obtained from the experiment were measured in SPAD values, which is a unit used to assess the chlorophyll content in plants. The SPAD values ranged from 0 to 100, with higher values indicating that the plants had more chlorophyll content.
Determination of chlorophyll content
-
Chlorophyll content was determined as described previously[36]. To measure the concentration of chlorophyll, samples of leaves were taken from hydroponic experiment at regular intervals throughout the study. The chlorophyll was extracted from the leaves using a solution of 80% ethanol, which was then analyzed using a multi-function enzyme marker meter (Victor Nivo, PerkinElmer, Waltham, MA, USA) at wavelengths of 664 and 647 nm.
Determination of nitrate reductase (NR) activity and glutamine synthetase (GS) activity
-
According to the method described earlier[37], the activity of nitrate reductase was determined in roots. The definition of nitrate reductase activity is the decrease in the amount of
${\text{NO}^-_2} $ Statistical analysis
-
For statistical analysis, Prism 9.4 software (GraphPad software, USA) was used with Student's t-test. Differences were considered significant if p <0.05 or 0.01.
This work was supported by the National Key Research and Development Project (2022YFD1602107) and the Key S&T Special Projects of Shanxi Province, China (202201140601027-6). We thank Dr. Zhihong Zhang from Shenyang Agricultural University for providing tissue-cultured GL-3 plants.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Zeyuan Liu, Ziqing Ma
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Z, Ma Z, Li J, Bian N, Guo Z, et al. 2023. Interfering small ubiquitin modifiers (SUMO) exhibits apple's enhanced tolerance to nitrogen deficiency. Fruit Research 3:24 doi: 10.48130/FruRes-2023-0024 

Interfering small ubiquitin modifiers (SUMO) exhibits apple's enhanced tolerance to nitrogen deficiency
- Received: 25 June 2023
- Accepted: 09 August 2023
- Published online: 08 October 2023
Abstract: Nitrogen is one of the most critical macroelements required for apple biomass accumulation and quality formation. Thus, identification and characterization of candidate genes involved in nitrogen deficiency stress is of great importance to the sustainable development of the apple industry. SUMOylation is a kind of post-translational modification, which is widely involved in plant life processes, including responses to nitrogen (N) deficiency. Here, we identified apple small ubiquitin modifiers (SUMO) that participated in responding to nitrogen deficiency. Firstly, all isoforms of apple SUMOs were induced by N deficiency stress. Under N deficiency stress conditions, the MdSUMO2 RNAi transgenic apple plants showed better growth potential than the wild type GL-3, including more new leaves, increased height, thicker stems, and heavier fresh weight. In addition, the MdSUMO2 RNAi transgenic apple plants exhibited darker green leaves and greater root volume than GL-3 in response to N deficient conditions. Finally, the activated glutamine synthetase (GS) activity, nitrate reductase (NR) activity, as well as the induced expression of genes involved in nitrogen assimilation in the root might contribute to the tolerance of MdSUMO2 RNAi transgenic apple plants in response to nitrogen deficiency. Taken together, our finding provides insight to further study the mechanisms underlying the essential roles of apple MdSUMOs in responding to N deficient conditions.
-
Key words:
- Apple /
- SUMO /
- Nitrogen deficiency