









-
The epidermis cuticle of the plant, composed of cutin and wax, is a hydrophobic barrier that covers the surface of all land plants. Plant epidermal wax has the functions of preventing non-stomatal water loss, preventing ultraviolet damage, maintaining surface cleaning and waterproofing, resisting the invasion of diseases and pests, etc., which plays a very important role in adapting to the external environment [ 1− 3] . At present, the metabolic pathway of plant wax biosynthesis has been basically clarified through the studies of various plant wax biosynthesis including Arabidopsis, rice and tomato. However, the transport mechanism of epidermis wax is not clear.
The synthesized wax is transported from the endoplasmic reticulum (ER) to the plasma membrane (PM), then to the apoplast, and then from the apoplast to the epidermal cuticle through trans-cell wall transport, where it is assembled into epidermal wax crystals. Previous studies have shown that ABCG transporter is necessary for the normal wax transport of plants. The absence of ABCG7 in moss ( Physcomitrella patens) exhibited severely deficient in cuticular wax and less tolerant to drought stress [ 4] . AtABCG12/ AtCER5, the first cloned ABCG transporter from plants, was proved to be involved in the cuticle wax transport [ 5] . Its absence leads to the deposition of wax components in the cytoplasm and a decrease in the amount of wax in the epidermis. AtABCG13 is essential for the transport of flower cuticular lipids [ 6] . OsABCG26 and OsABCG15 genes cooperate to regulate the transport of wax components in the outer wall of rice pollen [ 7] . AtABCG32 and its homologous genes, barley HvABCG31 and maize Glossy13, are involved in the extracellular transport of various wax components [ 8− 10] . However, it is unknown whether ABCG transporters have the function of wax transport in apple.
In this study, we isolated an ABCG transporter MdABCG25 from apple, and obtained transgenic Arabidopsis plants stably expressing MdABCG25 and MdABCG25 transgenic apple calli. Subsequently, the epidermal ultrastructure and permeability, amounts of cuticular wax and resistance to abiotic stresses of transgenic lines were examined compared with wild type. Ultimately, we found that MdABCG25 was involved in the transport of cuticular wax, reduced water loss, improved the response to drought and salt conditions.
Our study reveals a new molecular regulation mechanism of cuticular wax transport in apple, providing a candidate gene for studying the formation of apple wax layer and improving apple gloss quality.
-
All kinds of apple tissues, including root, leaf, flower, fruit, and pericarp, were collected from eight-year-old 'Royal Gala' apple trees.
Tissue-cultured 'Royal Gala' apple seedlings were subcultured at 30-d intervals on Murashige and Skoog medium (4.43 g·L −1 MS and 30 g·L −1 sucrose) with 0.3 mg·L −1 6-benzylaminopurine (6-BA), 0.2 mg·L −1 3-indoleacetic acid (IAA), 0.1 mg·L −1 gibberellin A3 (GA3) and 0.8% agar under a 16 h light/8 h dark photoperiod at 26 °C with 60% relative humidity. The seedlings were treated with 10% polyethylene glycol (PEG) 6000, 100 mmol·L −1 NaCl for 0, 1, 2, 3, 6, or 12 h, or 100 μmol·L −1 ABA for 0, 3, 6, 9, 12, or 24 h.
'Orin' apple calli were subcultured on MS medium with 0.4 mg·L −1 6-benzylaminopurine (6-BA), 1.5 mg·L −1 2, 4-dichlorophenoxyacetic acid (2,4-D) and 0.8% agar for 20 d in the dark at 26 °C.
Arabidopsis, including ecotype Columbia (Col-0) and MdABCG25 ectopic expression transgenic lines ( MdABCG25-OE-1, MdABCG25-OE-2, MdABCG25-OE-3), and tobacco ( Nicotiana benthamiana) seedings were grown on 1/2 MS medium under 16 h light/8 h dark photoperiod at 26 °C and 60% relative humidity. Transgenic Arabidopsis seeds were screened in 1/2 MS medium containing 50 mg·L −1 Kanamycin. Positive transgenic plants were detected by polymerase chain reaction (PCR) and real-time quantitative PCR (RT-qPCR). The phenotypic analysis was conducted using the homozygous T3 plants obtained after three consecutive generations of screening. Seeds of the Col-0 and MdABCG25-OE transgenic lines were vernalized with low temperature (4 °C) for 3 d after sowing on 1/2 MS medium. Then, the 4-day-old seedlings with uniform sizes were transferred to 1/2 MS, 1/2 MS with 6% PEG 6000, 1/2 MS with 100 mmol·L −1 NaCl or 1/2 MS with 30 µmol·L −1 ABA medium. The root lengths and fresh weight of the seedlings were measured using ImageJ-win64 software and one in 10,000 electronic balance after 2 weeks.
Bioinformatics analysis of the MdABCG25 gene
-
Basic information about the MdABCG25 sequence were obtained from the NCBI database (
www.ncbi.nlm.nih.gov/ ), namely number of base pairs (bp), number of amino acids (aa), number of exons and introns, chromosome location, and analysis of conserved domains. The molecular weight, isoelectric point (pI), instability index, and aliphatic index were predicted using ProtParam website (https://web.expasy.org/protparam/ ). The hydropathicity were predicted using ProtScale website (http://web.expasy.org/protscale ). The secondary and tertiary structures were predicted using Phyre2 website (www.sbg.bio.ic.ac.uk/phyre2/html/page.cgi?id=index ), PSIPRED website (http://bioinf.cs.ucl.ac.uk/psipred/ ) and SWISS-MODEL website (https://swissmodel.expasy.org/interactive ). The transmembrane domains were predicted using TMHMM2.0 website (www.cbs.dtu.dk/services/TMHMM/ ). The signal peptides were predicted using SignalP - 5.0 website (https://services.healthtech.dtu.dk/service.php?SignalP-5.0 ). The serine, threonine and tyrosine phosphorylation sites were predicted using NetPhos 3.1 website (https://services.healthtech.dtu.dk/service.php?NetPhos-3.1 ).Phylogenetic analysis, multiple sequence alignment of MdABCG25 proteins and conserved domain analysis
-
ABCG25 proteins of 17 different plants were obtained from the NCBI database (
www.ncbi.nlm.nih.gov/ ) and constructed genetic evolutionary trees using MEGA_64 software. DNAMAN were used for sequence alignment. The conserved domains were predicted using SMART (http://smart.embl-heidelberg.de/ ). The bootstrap method was used to identify the evolutionary pathway, and 1,000 repeats were generated to verify the results.Analysis of the MdABCG25 promoter
-
The cis-acting elements in the MdABCG25 promoter (2,000 bp upstream of the transcription start site) were identified using PlantCARE website (
http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ).Construction of the MdABCG25 expression vector and genetic transformation into apple calli and Arabidopsis
-
The full-length MdABCG25 cDNA sequences were cloned from 'Royal Gala' apple tissue-cultured seedlings into the pRI-101-GFP plant expression plasmid downstream of the cauliflower mosaic virus (CaMV) 35S promoter using homologous recombination method. The primers are shown in Supplemental Table S1. MdABCG25 overexpressed and RNAi vectors were transformed into Agrobacterium tumefaciens LBA4404 and GV3101. Transgenic Arabidopsis with MdABCG25 ectopically expressed and transgenic apple calli were obtained by agrobacterium-mediated method [ 11] .
RNA extraction and real-time quantitative RT-qPCR analysis
-
The total RNA of plant materials (including tissues of 'Royal Gala' apple trees, tissue-cultured apple seedlings, apple calli, and leaves of Arabidopsis) were extracted using the OminiPlant RNA Kit (Dnase I, ComWin Biotech Co., Ltd., Beijing, China) under RNase-free conditions. The cDNA as the template of RT-qPCR was obtained using the MonScript TM RTⅢ All-in-One Mix with dsDNase (Monad Biotech Co., Ltd., Wuhan, China). 18s rRNA and AtACTIN rRNA were used as internal reference genes of apple and Arabidopsis, respectively. The RT-qPCR was performed by UltraSYBR Mixture (Low ROX) (ComWin Biotech Co., Ltd., Beijing, China) and 2 −ΔΔCᴛ calculation method. Quantitative correlation primers are shown in Supplemental Table S1.
Toluidine blue (TB) staining, water loss assays, and chlorophyll leaching assays
-
All experiment were performed with the five-week-old Arabidopsis. The inflorescences, stems and rosette leaves of Arabidopsis were stained in the 0.05% TB staining solution for 10 h at room temperature. Rinse all samples with deionized water at least three times [ 12] .
The rosette leaves of Arabidopsis were cut and weighed every 30 min until 150 min, and then the water loss rate was calculated.
The rosette leaves of Arabidopsis were weighed and placed into 50 mL tubes containing 40 mL 80% ethanol solution at 25 °C in the dark. One mL was taken from the sample tubes at 10, 30, 60, 120, and 180 min for spectrophotometer measurement [ 13] .
The absorbance of each sample was measured at 649 nm and 665 nm respectively, and the formula is as follows:
Chlorophyll concentration in the extracting solution (C) = 6.63 A 665+ 18.08 A 649
Chlorophyll content (mg·g −1 FW) = (C * V)/FW
Extraction of cuticular waxes and component analysis by gas chromatography–mass spectrometry (GC-MS)
-
The stems and rosette leaves of Arabidopsis at five weeks of age with cauline leaves and siliques removed were cut off and weighed. Then their areas were measured, and their cuticular waxes were extracted with chloroform.
Each sample was replicated three times. The stems and rosette leaves were immersed in 20 mL chloroform for 30 s and repeated three times until all the wax was dissolved [ 14− 16] .
N, O-bis (trimethylsilyl) trifluoroacetamide (BSTFA, Sigma) was added and the derivatization reaction was carried out at 70 °C for 60 min with oscillation. Then BSTFA was dried by N 2. The sample was redissolved in chloroform and N-tetracosane (C24 alkane) was added as internal standard for gas chromatography-mass spectrometry (Shimadzu, Japan) analysis [ 17] .
Scanning electron microscopy (SEM)
-
The five-week-old Arabidopsis rosette leaves and stems were rapidly frozen with liquid nitrogen and freeze-dried in vacuum for 24 h (FDU-1110, TOKYO RIKAKIKAI CO LTD, Japan, 50/60 Hz, 1.7kVA). Subsequently, they were fixed to metal plates and sprayed with gold following the procedure described in Qi et al [ 18] . The epidermal wax crystals were observed using a scanning electron microscope (SEM, JSM-6610LV, JEOL, Tokyo, Japan) at 5 kV.
Physiological measurements of apple calli and Arabidopsis under PEG 6000, NaCl and ABA treatments
-
'Orin' apple calli were treated with 5% PEG 6000, 150 mmol·L −1 NaCl or 50 μmol·L −1 ABA for 20 d. Fresh weights, malondialdehyde (MDA) contents and relative conductivity were measured [ 19] .
Arabidopsis seeds, including ecotype Columbia (Col-0) and MdABCG25 ectopic expression transgenic lines, were sterilized with 75% ethanol and 15% NaClO and germinated on 1/2 MS medium after 2-d vernalization at 4 °C.
After 4 d, the seedings were transferred to 1/2 MS, 1/2 MS with 5% PEG 6000, 1/2 MS with 30 μmol·L −1 ABA or 1/2 MS with 150 mmol·L −1 NaCl. The root length and fresh weight of the seedlings were measured using ImageJ-win64 software and one in 10,000 electronic balance after 2 weeks.
After 7 d, the seedlings were grown in pots containing 80% medium vermiculite and 20% substrate of 7 cm × 7 cm × 9 cm. All pots were watered with the same amount to ensure the accuracy of the experiments. Seedlings were grown under normal moist environment for 15 d. Then, they were treated with drought for 20 d or 150 mmol·L −1 NaCl for 15 d, respectively. After 20 d, the group treated with drought stress was resumed watering for 2 d. Chlorophyll content, MDA content, H 2O 2 content and
${\text{O}^-_2} $ Protein network relationship predictions of the AtABCG25 protein and MdABCG25 protein
-
The main interaction networks of the AtABCG25 and MdABCG25 protein were predicted using the online STRING database (
http://string-db.org ).Data presentation and statistical analysis
-
Each result was based on three parallel experiments with at least three technical replicates to ensure the accuracy of the results. Error bars show the standard deviation (SD) of three replicates. All data were analyzed by IBM SPSS Statistics 20 software and compared using the single factor Duncan method. Treatment means with different lowercase letters are significantly different at p < 0.05.
-
The apple MdABCG25 (Md05G1042200) gene was identified in the apple genome by blasting the Arabidopsis ABCG25 (AT1G71960) gene against the NCBI database. The full-length cDNA of MdABCG25 was 1,956 bp in length, encoding 651 amino acids and containing three exons and two introns ( Fig. 1a). Figure 1b showed the properties of each amino acid of MdABCG25 protein. MdABCG25 gene was mapped to the 5 th chromosome. Its predicted molecular weight is 71.78 kDa, and its predicted isoelectric point (pI) was 8.93. A maximum hydropathicity value of 3.233 and a minimum value of −3.100 were calculated by ProtScale, and the general average of hydropathicity of the protein was 0.135 ( Supplemental Fig. S1a). The instability index was calculated at 36.45, which classified the protein as stable. Its aliphatic index was 99.62. MdABCG25 protein consisted of 44.85% alpha helices, 35.79% random coils, 14.59% extended strands and 4.76% beta turns ( Fig. 1c, Supplemental Fig. S2). Based on the folding of the secondary structure, the tertiary structure of the AtABCG15 ( Fig. 1d) and MdABCG25 protein ( Fig. 1e) was predicted. There are six transmembrane domains, but no signal peptides were found ( Supplemental Fig. S1b & c). The phosphorylation sites were predicted as follows: 41 serines, 19 threonines and six tyrosines ( Supplemental Fig. S1d).
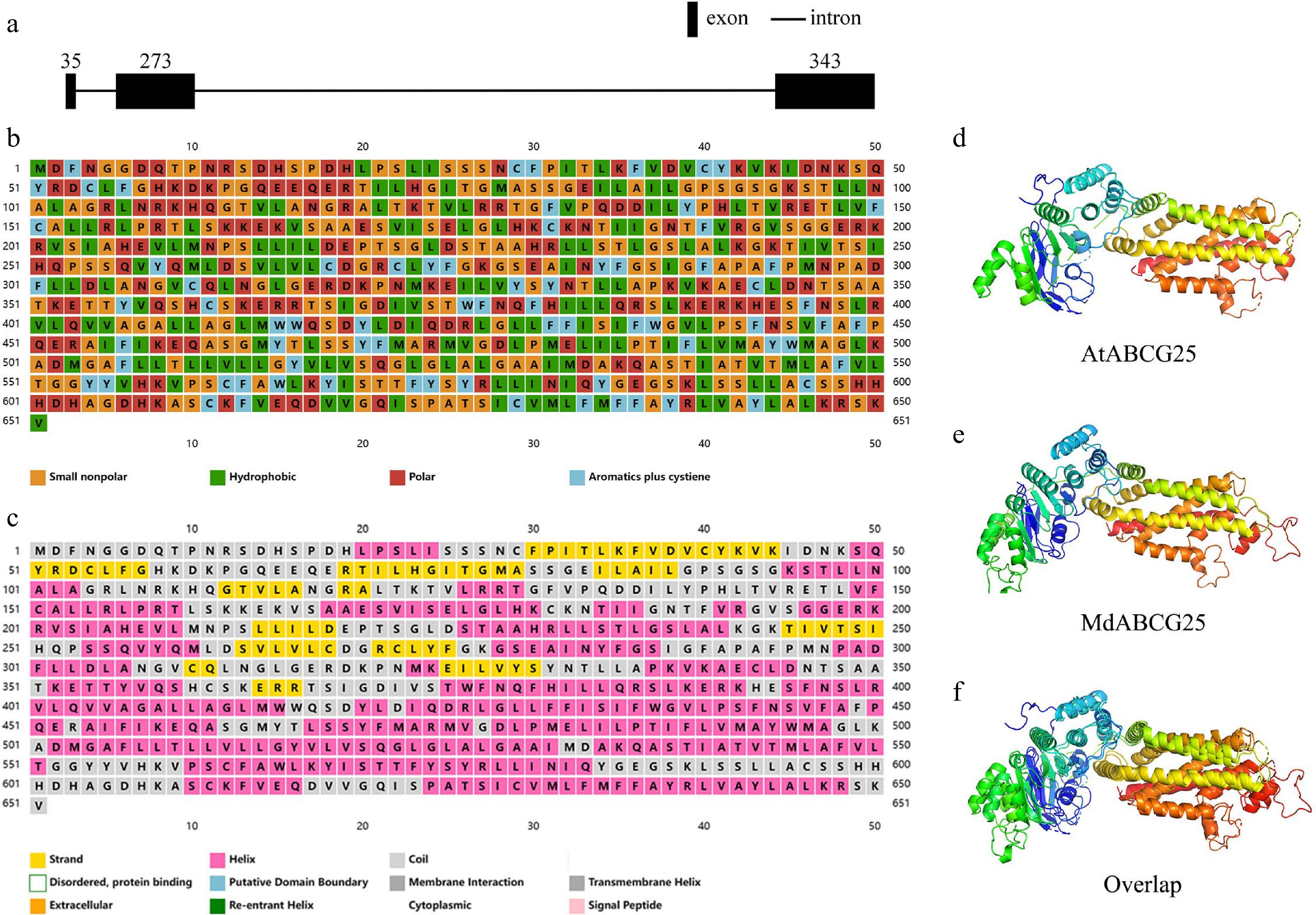
Figure 1.
Basic bioinformatic information about AtABCG25 and MdABCG25 sequence. (a) Genomic structure of MdABCG25 sequence. The black rectangles denote exons, and the black lines denote introns. (b) Properties of each amino acid of MdABCG25 protein; orange for small nonpolar, green for hydrophobic, red for polar, and blue for aromatics plus cystiene. (c) Predicted secondary structures of MdABCG25 protein, each represented by a different color; yellow for strands, pink for helices, and gray for coils. The predicted tertiary structure of (d) AtABCG25 protein, (e) MdABCG25 protein, and their (f) overlaps.
Phylogenetic analysis and conserved domain analysis of ABCG25 proteins from different species
-
To determine the phylogenetic relationship between MdABCG25 protein and ABCG25 in other species, we obtained 17 ABCG25 protein sequences, and constructed a phylogenetic tree by MEGA_64 software in order to compare these 17 protein sequences ( Fig. 2a). The MdABCG25 protein ( Malus × domestica) exhibited the closest evolutionary relationship with Pyrus ussuriensis × Pyrus communis, Prunus persica, and Prunus dulcis. The multiple sequence alignment showed that the homology of MdABCG25 to Pyrus ussuriensis × Pyrus communis, Prunus persica, and Prunus dulcis was 94.78%, 80.60%, and 80.15%, respectively ( Fig. 2b). In addition, ZmABCG25 proteins ( Zea mays) was most distant from MdABCG25.
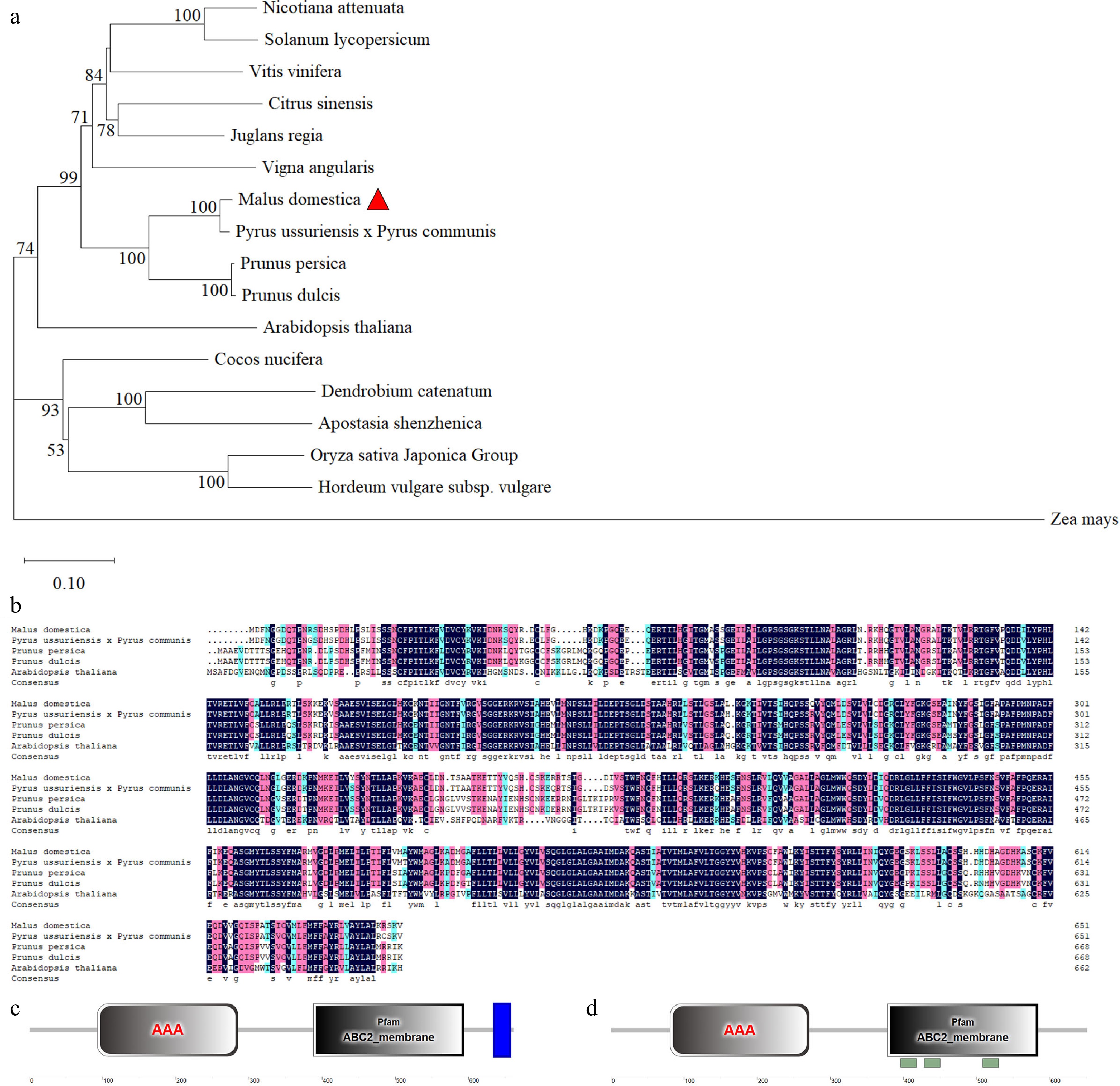
Figure 2.
Phylogenetic analysis, multiple sequence alignment and conserved motif analysis of ABCG25 proteins. (a) Phylogenetic tree assembled from ABCG25 proteins of 17 different species: Nicotiana attenuate, Solanum lycopersicum, Vitis vinifera, Citrus sinensis, Juglans regia, Vigna angularis, Malus domestica, Pyrus ussuriensis × Pyrus communis, Prunus persica, Prunus dulcis, Arabidopsis thaliana, Cocos nucifera, Dendrobium catenatum, Apostasia shenzhenica, Oryza sativa Japonica Group, Hordeum vulgare subsp. Vulgare, Zea mays. (b) Multiple sequence alignment of ABCG25 protein from different species: Malus domestica, Pyrus × bretschneideri, Prunus persica, Prunus dulcis, Arabidopsis thaliana. The conserved structural domains analysis of (c) AtABCG25 protein and (d) MdABCG25 protein sequences. The gray rectangles represent AAA domain. The blue rectangles represent transmembrane domain (TMD).
The structural domains of AtABCG25 and MdABCG25 protein sequences were analyzed using the SMART website. They both contain a conserved AAA domain and an ABC2_membrane domain in a similar location ( Fig. 2c & d). Furthermore, AtABCG25 has one more transmembrane domain (TMD) than MdABCG25, suggesting that they are ABCG half-sized transporters. The tertiary structures of AtABCG25 and MdABCG25 were predicted by the Phyre website. According to their tertiary structure merging results, the two structures strongly overlap ( Fig. 1f). These results all showed that they may have similar functions.
Expression profiles in different apple tissues and promoter cis-acting element analysis of MdABCG25
-
In previous studies, many genes related to cuticular wax were highly expressed in apple pericarp [ 11, 13, 16, 19, 21, 22] . To examine tissue-specific expression of MdABCG25, its transcript levels were analyzed in apple root, leaf, flower, fruit, and pericarp using a quantitative RT-PCR assay. The findings showed that MdABCG25 was expressed at different levels in all measured tissues. As shown in Fig. 3a, MdABCG25 showed the highest expression in apple pericarp and lower expression in leaves and flowers, suggesting a potential specific function in apple fruit development.
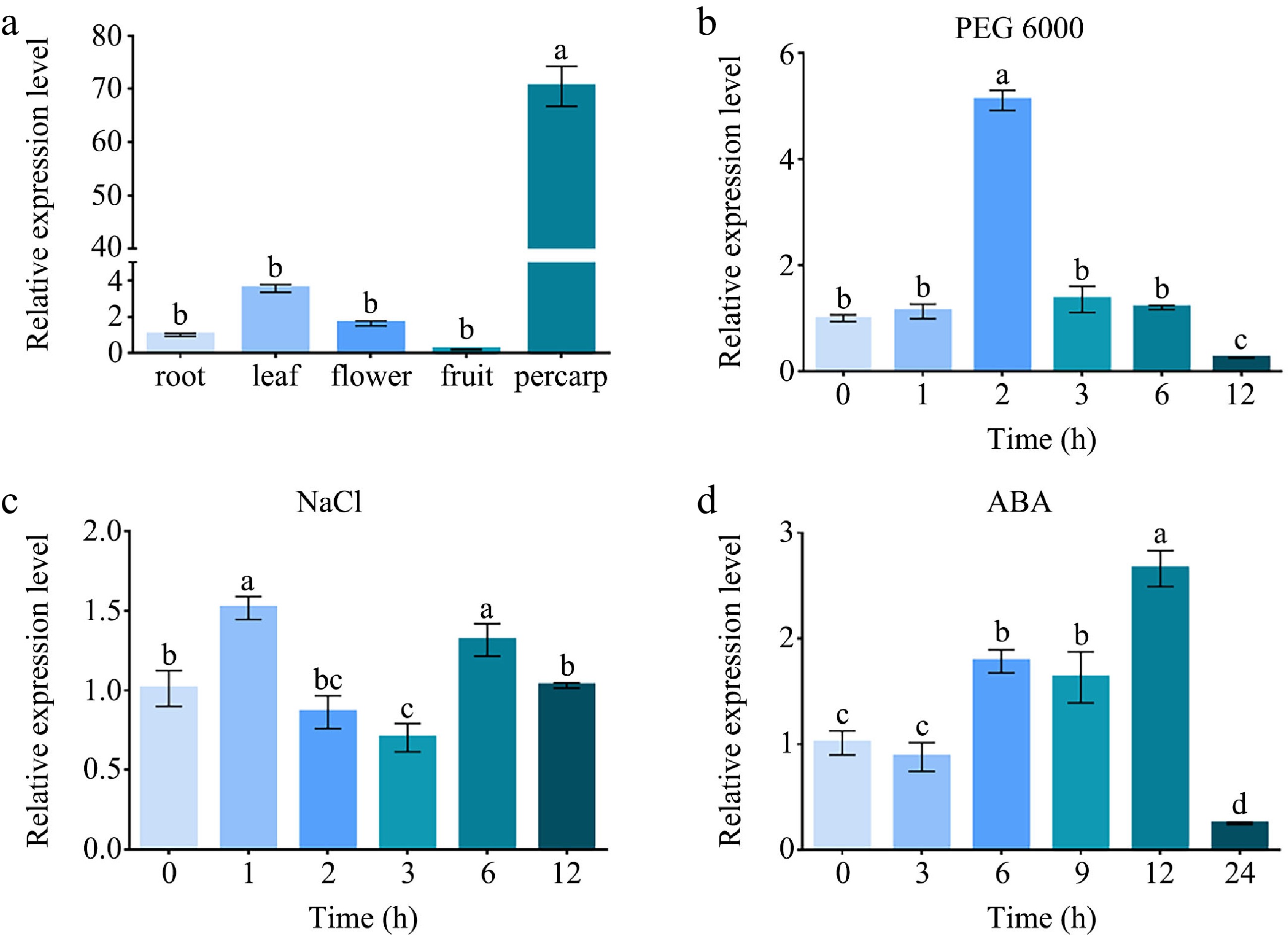
Figure 3.
The relative expression analysis of MdABCG25. (a) The relative expression levels of MdABCG25 in apple root, leaf, flower, fruit, pericarp measured by RT-qPCR. The relative expression levels of MdABCG25 under different stress treatments: (b) 10% PEG 6000, (c) 100 mmol·L −1 NaCl, and (d) 100 μmol·L −1 ABA measured by RT-qPCR. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
Most ABCG transporters play an important role in response to various abiotic stresses [ 23− 26] . To predict the putative functions of the MdABCG25 genes in response to abiotic stresses, its 2,000 bp upstream of the transcription start site was analyzed. Potential cis-acting elements in MdABCG25 promoter were determined by the PlantCARE website ( Table 1). The promoter region of the MdABCG25 gene contained various stress responsive elements, such as the drought inducibility element (MBS), the low-temperature response element (LTR), and the ABA response element (ABRE). The promoter sequence also contained multiple plant hormone response elements, such as the gibberellin-responsive element (P-box), the MeJA response element (CGTCA-motif), and the auxin-responsive element (TGA-element). These plant hormones are all related to the stress response of plants. Besides, an ACE, a light-responsive element, that can play a crucial part in plant growth and development was found in its promoter region.
Table 1. MdABCG25 promoter cis-acting element analysis.
Cis-element name Sequence Function Start site (bp) Termination site (bp) P-box CCTTTTG Gibberellin-responsive element 271 277 MBS CAACTG MYB binding site involved in drought-inducibility 531 536 CGTCA-motif CGTCA cis-acting regulatory element involved in the MeJA-responsiveness 723 727 LTR CCGAAA cis-acting element involved in low-temperature responsiveness 1,122 1,127 TGA-element AACGAC Auxin-responsive element 1,324 1,329 ACE CTAACGTATT cis-acting element involved in light responsiveness 1,458 1,467 ABRE ACGTG cis-acting element involved in the abscisic acid responsiveness 1,856 1,860 Responses to the ABA, PEG 6000, and NaCl treatments of the MdABCG25
-
To confirm whether MdABCG25 gene responds to abiotic stresses in apple, MdABCG25 transcription levels were measured in the tissue-cultured 'Royal Gala' apple seedlings under PEG 6000, NaCl or ABA treatments using RT-qPCR ( Fig. 3b & d). We found that the expression of MdABCG25 gene showed an increase-decrease trend under three abiotic stress conditions. The relative MdABCG25 expression levels were up-regulated dramatically at 2 h and then decreased in response to the PEG 6000 treatment. The expression of MdABCG25 fluctuated, and peaked at 1 h and 6 h in response to the NaCl treatment. The MdABCG25 expression rose initially, peaked at 12 h, and then declined in response to the ABA treatment. These results suggested that MdABCG25 may be involved in responding to various abiotic stresses.
Moreover, we obtained MdABCG25 overexpressed ( MdABCG25-OE), and suppressed ( MdABCG25-RNAi) apple calli. The relative expression levels of MdABCG25 in transgenic apple calli were measured using RT-qPCR ( Fig. 4b). The 15-day-old apple calli were subcultured on MS medium, 5% PEG 6000, 150 mM NaCl or 50 μM ABA treatments ( Fig. 4a). The results showed there were no significant differences in fresh weight among MdABCG25-OE line, MdABCG25-RNAi line and WT apple calli on MS medium ( Fig. 4c). However, the fresh weight of the MdABCG25-OE line significantly increased compared to that of the WT apple calli, whereas the fresh weight of the MdABCG25-RNAi line significantly decreased compared to that of WT under PEG 6000, NaCl and ABA treatment. MDA content and relative conductivity showed opposite changes, indicating that the damage of apple calli under abiotic stresses was reduced with the increase of MdABCG25 expression level ( Fig. 4d & e). These results suggest that MdABCG25 plays a vital role in the response to abiotic stresses in apple. In addition, overexpression of MdABCG25 decreased the sensitivity of apple calli to ABA.
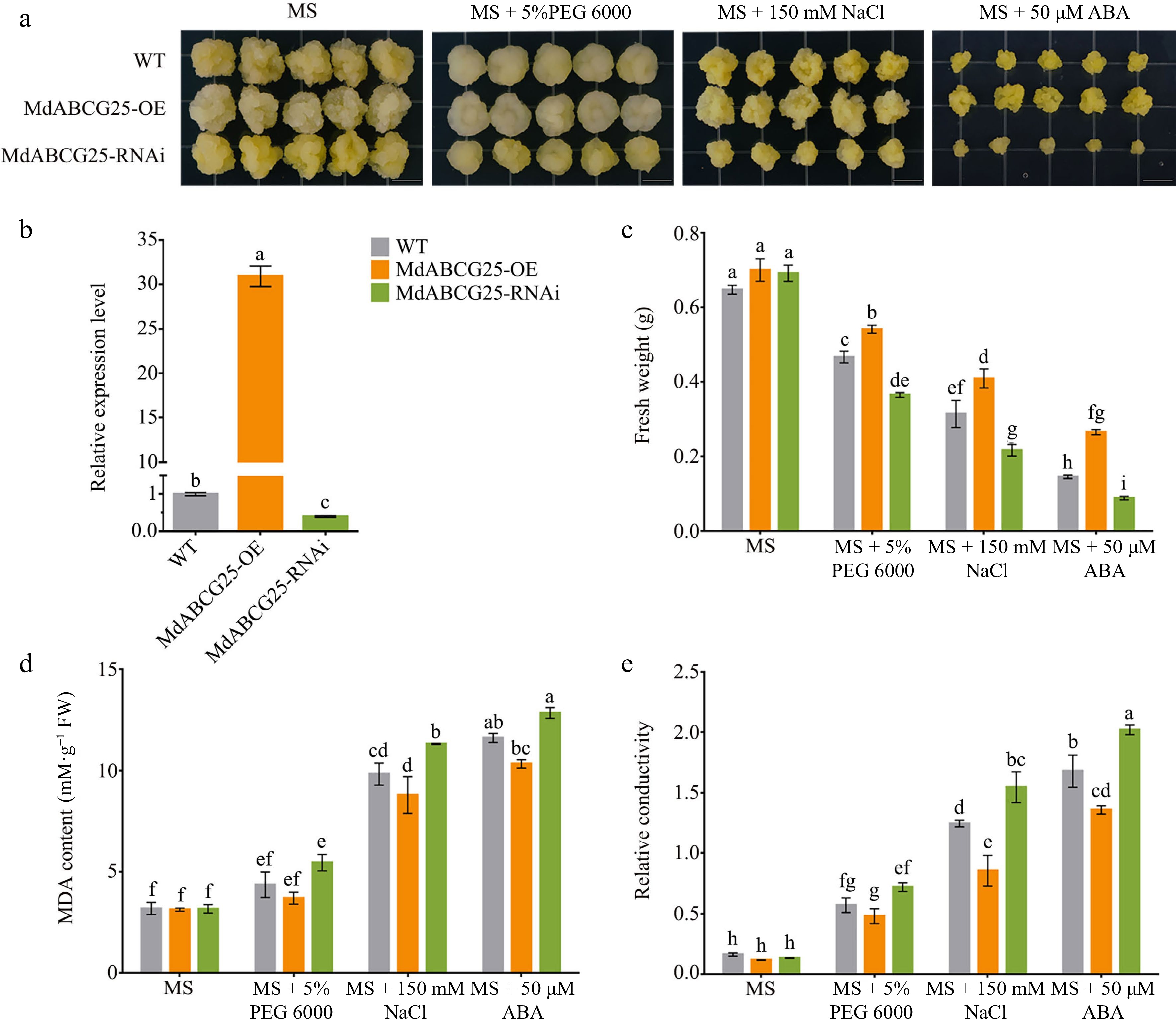
Figure 4.
Phenotypes of MdABCG25 transgenic apple calli under different abiotic stress conditions. (a) Phenotypes of MdABCG25 transgenic lines and wild-type (WT) apple calli treated with MS medium, MS + 5% PEG 6000, MS + 150 mmol·L −1 NaCl and MS + 50 μmol·L −1 ABA. (b) Relative expression levels of MdABCG25 in wild-type (WT) apple calli and MdABCG25 transgenic lines measured by RT-qPCR. (c) Fresh weight, (d) MDA content and (e) relative conductivity of MdABCG25 transgenic lines and WT apple calli after treatments. FW = fresh weight. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
MdABCG25 reduced epidermal permeability in Arabidopsis
-
To investigate the function of MdABCG25 in plants, we obtained three MdABCG25 transgenic Arabidopsis ( MdABCG25-OE-1, MdABCG25-OE-2, MdABCG25-OE-3), and measured MdABCG25 transcription levels using RT-qPCR ( Fig. 5b). To explore whether epidermal permeability was altered in MdABCG25 transgenic Arabidopsis, the inflorescences, stems, and leaves of MdABCG25 transgenic lines and Col-0 was stained with TB ( Fig. 5a). The results of TB staining showed that inflorescence, stems and leaves of Col-0 were more easily stained than those of MdABCG25 transgenic Arabidopsis. The epidermal permeability of MdABCG25 transgenic lines was lower than that of Col-0. Moreover, it was found that the rosette leaves of MdABCG25 transgenic lines had lower water loss rates and chlorophyll extractions compared with Col-0 through water loss and chlorophyll extraction experiment ( Fig. 5c & d). In conclusion, we hypothesized that MdABCG25 may reduce plant epidermal permeability by promoting wax transport, suggesting a potential impact on plant drought resistance.
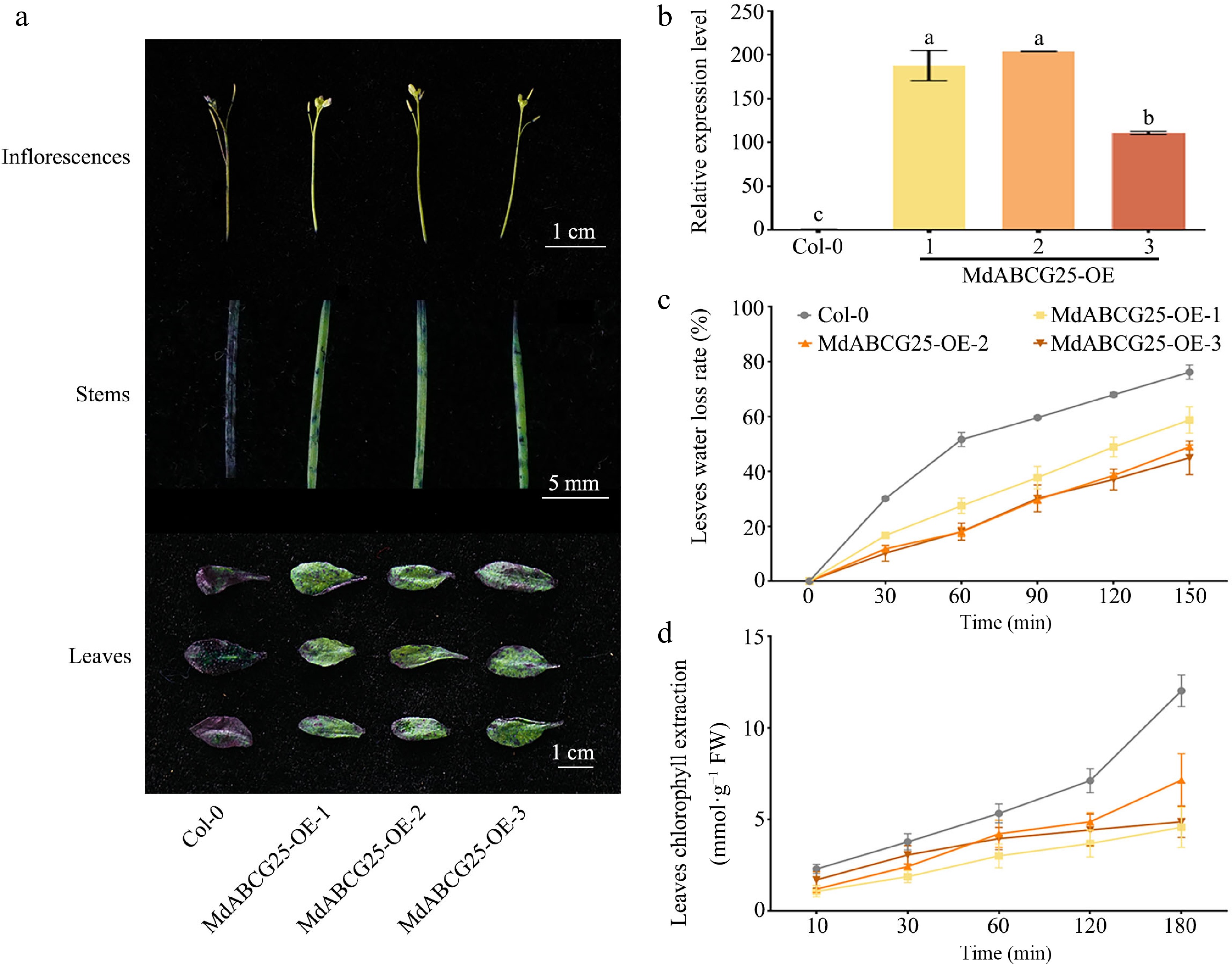
Figure 5.
MdABCG25 can reduce the epidermis permeability of Arabidopsis. (a) TB staining of inflorescences, stems and leaves of the MdABCG25 transgenic lines and Col-0. Scale bars correspond to 1 cm, 5 mm and 1 cm, respectively. (b) Expression levels of MdABCG25 in three MdABCG25 transgenic Arabidopsis and Col-0 using RT-qPCR analysis. (c) Water loss rate of rosette leaves of the MdABCG25 transgenic lines and Col-0. (d) Chlorophyll extraction of rosette leaves of the MdABCG25 transgenic lines and Col-0. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
MdABCG25 promoted cuticular wax accumulation
-
To study the function of MdABCG25 in wax transport, we tested the expression of wax-related genes in MdABCG25 transgenic Arabidopsis and Col-0. Some genes were up-regulated in transgenic Arabidopsis compared with Col-0, including AtCER1, AtACBP1, AtECH and AtKCS1 ( Supplemental Fig. S3). Furthermore, the content and composition of cuticular wax in the Arabidopsis stems were determined ( Fig. 6). There were significant differences in total wax load between MdABCG25 transgenic Arabidopsis and Col-0, both on leaves and stems ( Fig. 6a & b). In addition, the wax components in stems were analyzed by gas chromatography-mass spectrometry (GC-MS). The cuticular wax composition of MdABCG25 transgenic Arabidopsis was significantly different from that of Col-0, and the contents of alkanes, alcohols, aldehydes and ketones were significantly increased ( Fig. 6c− h). Among alkanes, MdABCG25 transgenic Arabidopsis has more C29 alkane and C31 alkane ( Fig. 6f). The contents of C30 aldehyde, C26 and C28 alcohols were much higher in MdABCG25 transgenic lines than in Col-0 ( Fig. 6e & h). However, the levels of fatty acids decreased in MdABCG25 transgenic Arabidopsis, such as palmitic acid and stearic acid ( Fig. 6g).
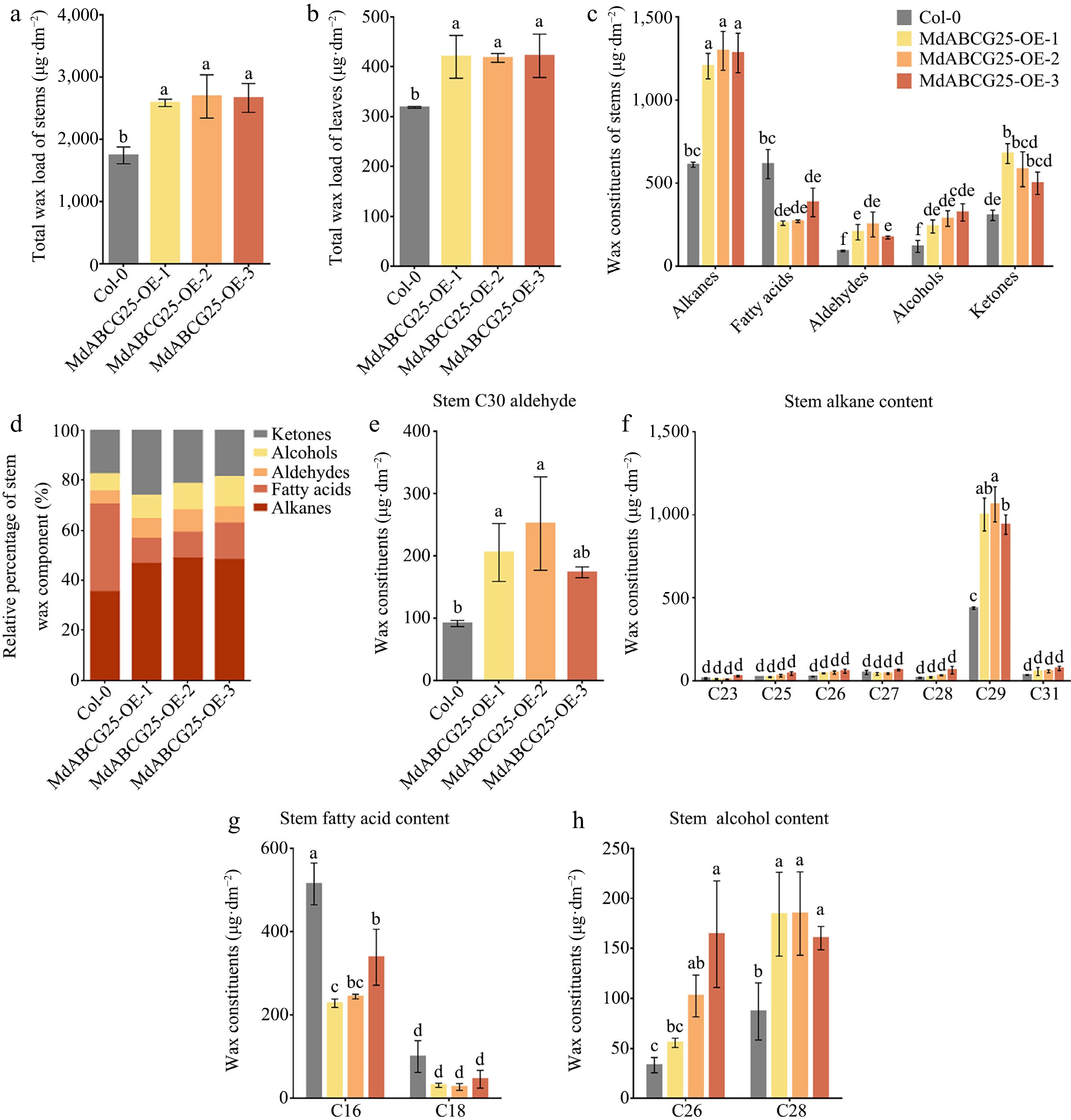
Figure 6.
The effect of MdABCG25 on cuticular wax content and composition. Total wax load of (a) stems and (b) leaves of the MdABCG25 transgenic Arabidopsis and Col-0. (c) Wax composition of stems of the MdABCG25 transgenic Arabidopsis and Col-0 analyzed by GC-MS. (d) Relative percentage of stems wax component. Stem wax constituents of (e) aldehydes, (f) alkanes, (g) fatty acids and (h) alcohols in MdABCG25 transgenic Arabidopsis and Col-0 analyzed by GC-MS. The compounds were tentatively identified by GC-MS solution software. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
In addition, we observed the epidermis ultrastructure of the MdABCG25 transgenic and Col-0 Arabidopsis stems and leaves using SEM ( Fig. 7). The wax crystals and wax plates of MdABCG25 transgenic Arabidopsis epidermis were significantly larger and more than those of Col-0, especially the long strip wax crystals increased observably ( Fig. 7a & b). These results indicated that the expression of MdABCG25 affected the chemical composition and ultrastructure of plants cuticular wax.
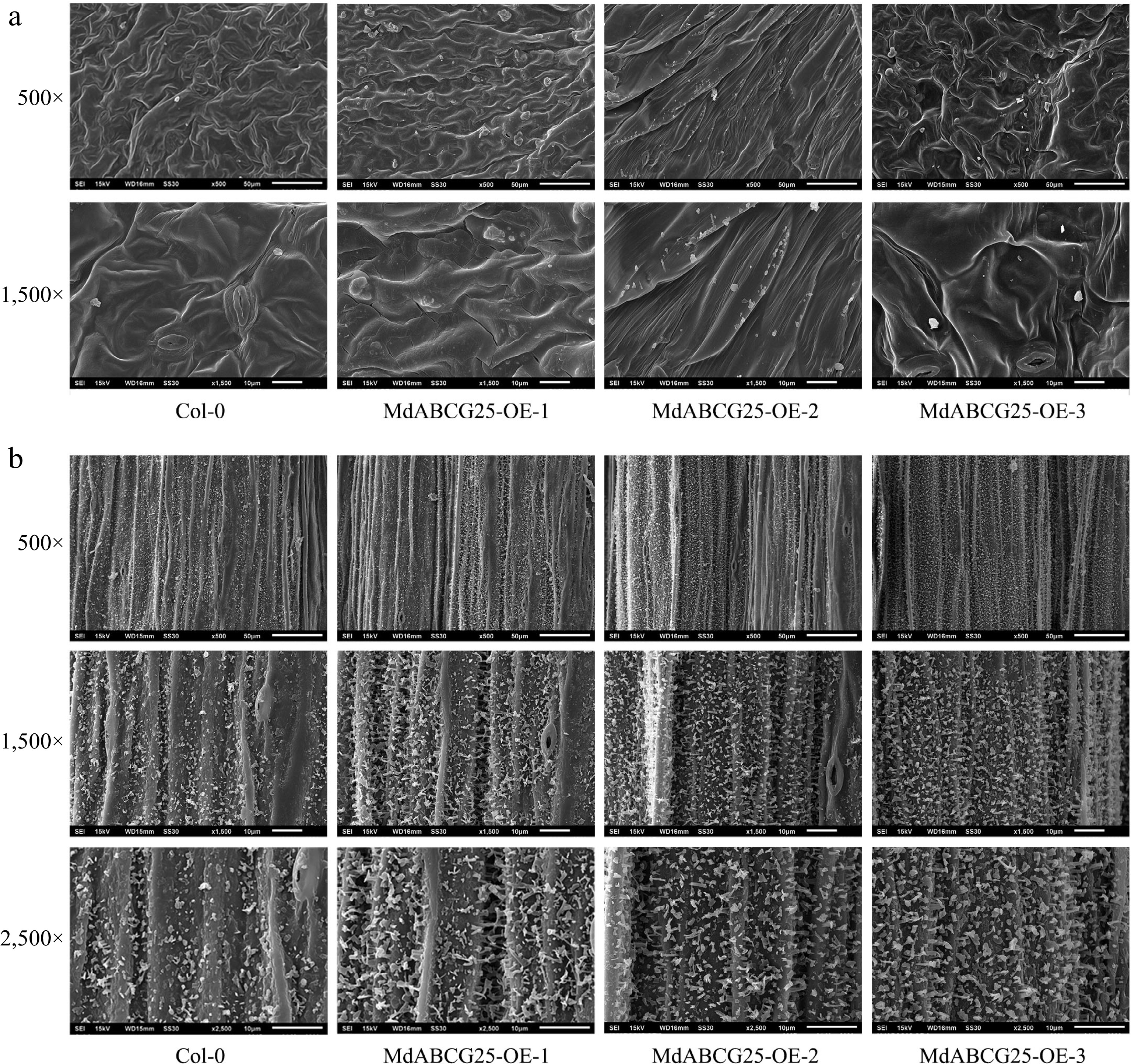
Figure 7.
Cuticle wax crystal morphology of MdABCG25 transgenic and Col-0 Arabidopsis observed by SEM. The epidermal ultrastructure of MdABCG25 transgenic and Col-0 Arabidopsis rosette (a) leaves and (b) stems. Wax crystals were monitored at 500×, 1,500× and 2,500× magnification, and scale bars correspond to 50 μm, 10 μm and 10 μm, respectively.
MdABCG25 increased Arabidopsis resistance to abiotic stress
-
To investigate whether the increase of epidermal wax load caused by MdABCG25 can enhance the resistance plant to abiotic stresses, Col-0 and MdABCG25 transgenic Arabidopsis seeding were transferred to 1/2 MS medium containing 30μmol·L −1 ABA, 5% PEG 6000, or 150 mmol·L −1 NaCl ( Fig. 8a). Under normal conditions, the root length of MdABCG25 transgenic lines was slightly larger than Col-0, but there was no significant difference in fresh weight ( Fig. 8b). Under the PEG 6000 or NaCl treatment, MdABCG25 transgenic lines showed obviously longer primary roots and greater fresh weight than Col-0 ( Fig. 8b & c). Under the ABA treatment, the MdABCG25 transgenic lines showed only a slight increase in root length and fresh weight. In addition, the MdABCG25 transgenic lines showed longer and more lateral roots than Col-0 Arabidopsis under PEG 6000 and NaCl treatments ( Fig. 8a). These results suggest that ectopic expression of MdABCG25 not only enhances the resistance of Arabidopsis to abiotic stresses but decreases plant sensitivity to ABA.
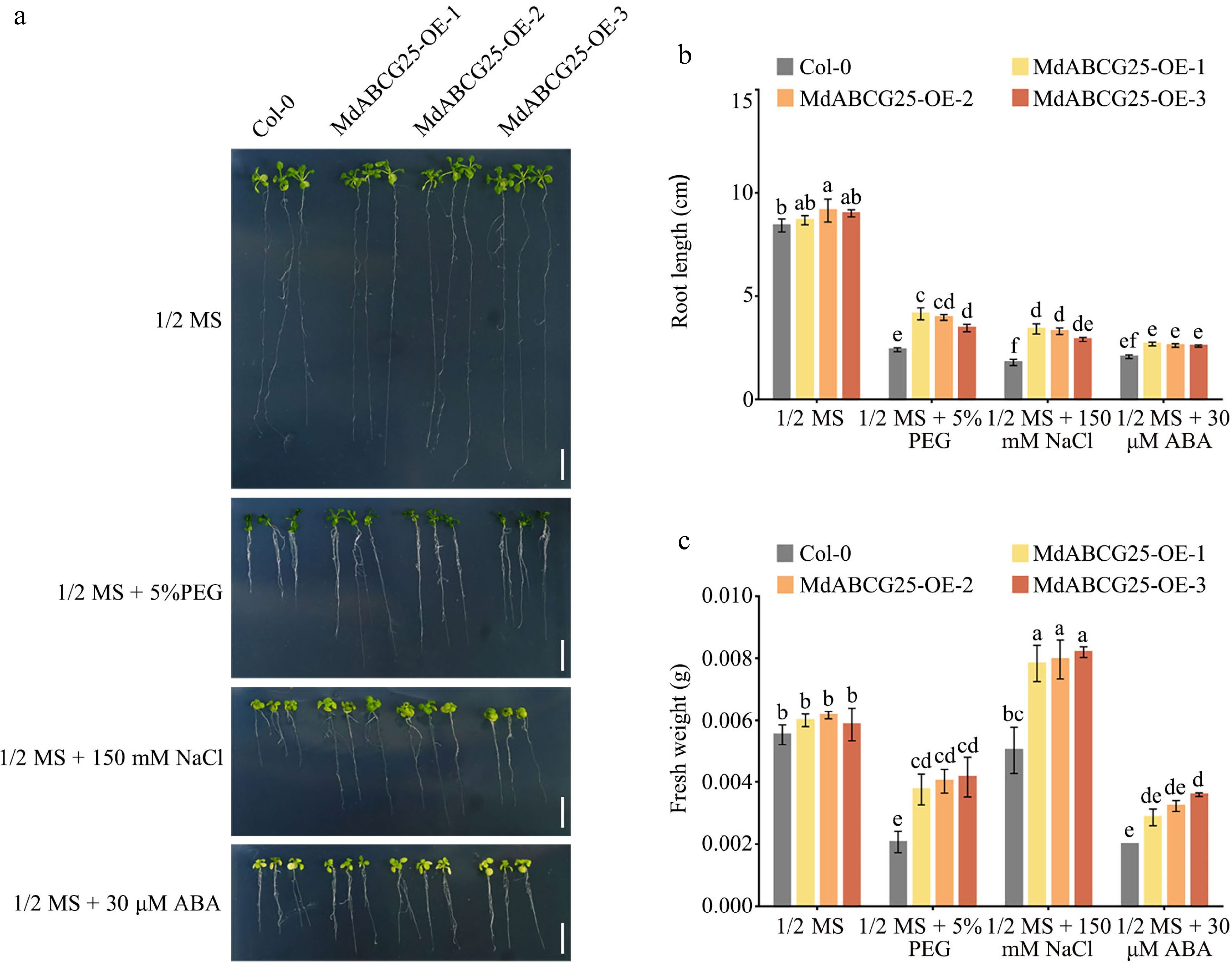
Figure 8.
Phenotypes of MdABCG25 transgenic and Col-0 Arabidopsis seedings under different abiotic stress conditions. (a) Phenotypes of MdABCG25 transgenic and Col-0 Arabidopsis seedings treated with 1/2 MS medium, 1/2 MS + 5% PEG 6000, 1/2 MS + 150 mmol·L −1 NaCl and 1/2 MS + 30 μmol·L −1 ABA. Bar = 1 cm. (b) Root length and (c) fresh weight of MdABCG25 transgenic and Col-0 Arabidopsis. FW = fresh weight. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
To further verify the resistance of MdABCG25 transgenic to drought and salt stress at seedling stage, the healthy 3-week-old MdABCG25 transgenic and Col-0 Arabidopsis seedlings were treated by drought for 20 d or 150 mmol·L −1 NaCl for 15 d, respectively ( Fig. 9a & f). After 20 d, the drought stress treatment group was resumed watering for 2 d. Under the drought or NaCl treatment, the growth of both transgenic lines and Col-0 was inhibited, but the degree of growth inhibition of transgenic lines was significantly lower than that of Col-0. The MdABCG25 transgenic seedings were more robust than Col-0, with significantly fewer dry rosette leaves. The chlorophyll content of MdABCG25 transgenic lines was higher than that of Col-0 Arabidopsis ( Fig. 9b & g). Moreover, MdABCG25 transgenic seedings showed lower MDA, H 2O 2 and
${\text{O}^-_2} $ 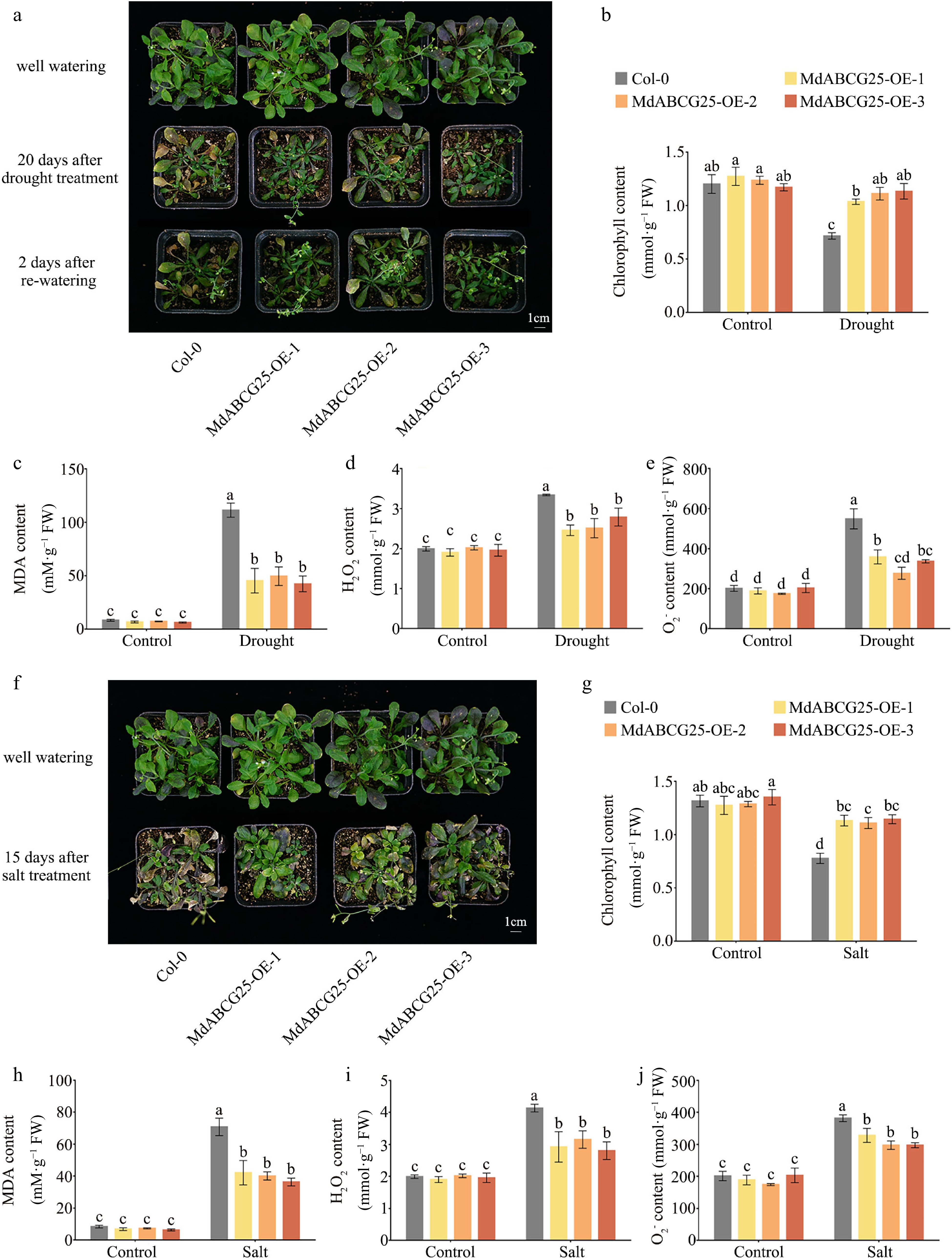
Figure 9.
MdABCG25 can enhance tolerance to drought and salt in Arabidopsis. (a) Phenotypes of MdABCG25 transgenic and Col-0 Arabidopsis in normal conditions or 20 d of drought treatment. Physiological indicators in MdABCG25 transgenic and Col-0 Arabidopsis in normal conditions and drought treatment; (b) chlorophyll content, (c) MDA content, (d) ${\text{O}^-_2} $ content, (e) H 2O 2 content. (f) Phenotypes of MdABCG25 transgenic and Col-0 Arabidopsis in normal conditions or 15 d of 100 mmol·L −1 NaCl treatment. Physiological indicators in MdABCG25 transgenic and Col-0 Arabidopsis in normal conditions and salt treatment; (g) chlorophyll content, (h) MDA content, (i) ${\text{O}^-_2} $ content, (j) H 2O 2 content. Values are mean ± SD of at least three biological replicates. Different lowercase letters indicate a significant difference at p < 0.05.
Protein interaction network predictions for the AtABCG25 protein and MdABCG25 protein
-
In order to further identify the regulating pathway of MdABCG25 in cuticular wax transport and alleviating abiotic stresses, we predicted the protein interaction network of AtABCG25 and MdABCG25 proteins ( Fig. 10). MdABCG25 was predicted to interact with ethylene related proteins, such as ETR1b, ERS, ERS2, and ETR1, indicating that MdABCG25 may be involved in regulating fruit ripening and plant aging. XP_008389536.1, XP_008389530.1, and XP_008339944.1, which were NRT1/ PTR family 4.6-like proteins, were predicted to interact with MdABCG25 ( Fig. 10b). In the network of AtABCG25 protein, ABCD1 is related to transport of lipids and fatty acid. F12K2.4 affects the cell wall structure. ETR1 and ERS1 are related to ethylene. NCED3 and ABA4 are related to abscisic acid. NRT1.1 is related to N absorption and transport. These interactions provide useful resources for further study of function of MdABCG25.
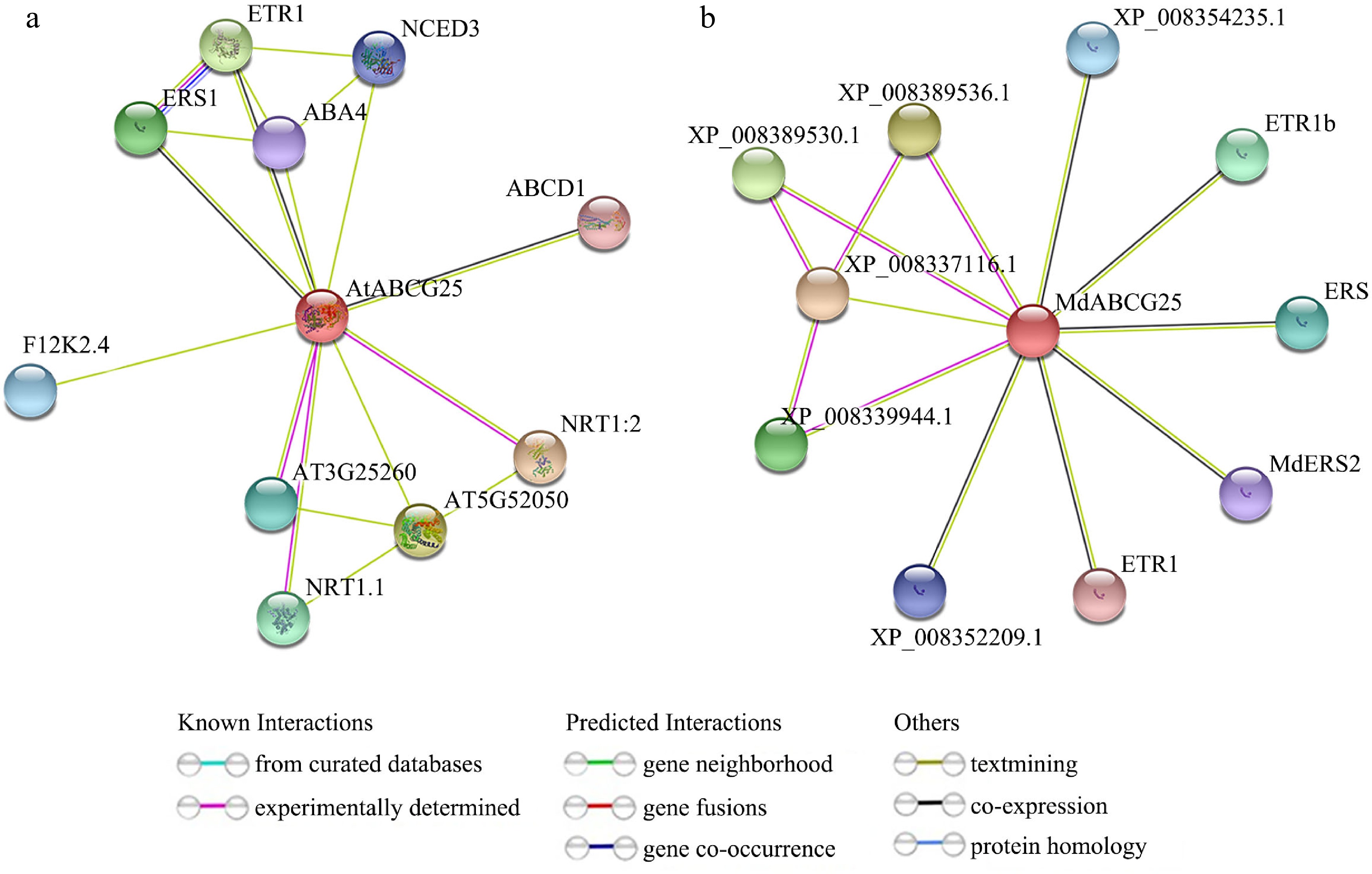
Figure 10.
Protein interaction networks for the AtABCG25 and MdABCG25 proteins. Predicted interaction networks for the (a) AtABCG25 and (b) MdABCG25 proteins generated using the online software STRING.
-
The cuticular wax on the surface of apples not only protects the fruit by enhancing tolerance to biotic and abiotic stresses [ 16, 27] , but also improves surface gloss and storage capacity after harvest [ 28] . At present, the biosynthesis mechanism of the cuticular wax components is gradually being understood [ 29] . However, the mechanism by which the cuticular wax components are exported from epidermal cells and assembled into complex, three-dimensional, hydrophobic plant surface structures is still unclear. In this work, we cloned and characterized the full-length MdABCG25, encoding a half-sized ABCG subfamily transporter, and studied its properties, aiming to reveal its function in the cuticular wax transport and the adaptation to extreme environments in apple.
In epidermal cells, wax components must be transported from the biosynthesis site of the endoplasmic reticulum to the plasma membrane (PM), through PM, and finally through the cell wall to the stratum corneum. The transport of wax components from the ER to PM can be accomplished through three possible pathways: direct transport at the physical contact sites between the ER and the PM [ 30] , transport via cytoplasmic carrier proteins [ 31] , and vesicle transport [ 31, 32] . After transporting the wax components to the PM, they must pass through the lipid bilayer to enter the apoplast. Studies have found that the ABCG transporter protein was involved in this process. The ABCG subfamily is divided into half-sized transporters (WBC) and full-sized transporters (PDR) [ 33] . WBC must form dimers to exert transport activity [ 33, 34] . AtABCG11 half-sized transporters can form dimers to transport different wax precursors or cutin precursors with itself, AtABCG12, AtABCG5, AtABCG9, or AtABCG14, respectively [ 34− 37] . MdABCG25 showed the highest expression in apple pericarp, suggesting that MdABCG25 may be involved in transport of cuticular wax in apple. Therefore, we suspect that MdABCG25, as a half-sized ABCG transporters, may combine with itself or other half-sized ABCG transporters to form homodimers or heterodimers to export different substrates.
Previous studies have shown that overexpression of TsABCG11 increases cuticular wax accumulation and abiotic stress resistance in Arabidopsis [ 38] . The promoter region of the MdABCG25 gene contained various stress responsive elements, further suggesting that MdABCG25 may also be involved in the plant stress response. The GC-MS analysis of wax showed heterologous expression of MdABCG25 in Arabidopsis can promote cuticular wax transport to resist drought and salt stress. Meanwhile, MdABCG25 transgenic Arabidopsis seedlings also showed stronger resistance to drought and salt stress in the tissue-cultured environment, even if only a small amount of cuticular wax was accumulated, suggesting that MdABCG25 can enhance plant resistance by up-regulating the expression of resistance genes in addition to increasing wax accumulation.
The results of this study indicate that overexpression of MdABCG25 not only improves drought resistance through stomatal-independent pathway, but also through stomatal-dependent pathway. However, these results contradict that MdABCG25 is insensitive to ABA shown by Figs 4 & 8. The plant hormone ABA, as an endogenous messenger in plants responding to biotic and abiotic stresses, plays an important role in the regulation of water loss [ 39] . Drought leads to a strong increase in plant ABA levels, inducing stomatal closure of protective cells and accompanied by major changes in gene expression in order to resist drought stress [ 40] . Similarly, some wax-related genes, MdKCS2 and MdLACS2, are insensitive to ABA and can enhance drought resistance [ 21, 22] . It is speculated that wax-related genes may also participate in improving drought resistance through an ABA-independent pathway. Moreover, overexpression of AtABCG25, ABA exporter, resulted in plant insensitivity to ABA and increased drought resistance [ 24] . NCED3 and ABA4, being related to ABA, are predicted to interact with MdABCG25 ( Fig. 10b). Therefore, MdABCG25 may have the same function. These conjectures need to be confirmed in future studies.
Although ABCG transporters have been proved to be involved in extracellular accumulation of wax molecules, the detailed biochemical processes behind transport events have not been determined [ 41] . The characteristics and functions of ABCG transporters have mostly been reported based on molecular genetic methods, in which the transport mechanism is still unknown [ 42] . Moreover, heterodimerization of half-sized ABCG family members provides great potential to identify a plenty of substrates for transport, but this also affects the specificity of transport substrates.
In conclusion, the process of wax transport in plant epidermis is very complicated, and there are many genes involved in this process, so it is difficult to study the genes involved in wax transport in plant epidermis. In this paper, MdABCG25 gene in apple was cloned and identified, and its characteristics were studied. MdABCG25 gene promotes the accumulation of plant cuticular wax, reduces the permeability of plant epidermis, and improves the drought resistance and salt resistance of plants. It laid a foundation for elucidating the molecular mechanism of plant wax transport in apple, and provided candidate genes for improving abiotic stress tolerance and fruit quality of apple.
-
The authors confirm contribution to the paper as follows: Li YY, Han J, Lv YH designed the idea of the research; Zhou MM performed the experiments, collected data, analyzed and interpreted results, and prepared draft manuscript; Yu ZH, Gao HN, Li MR, Wu YT, Li HY, Wang T assisted in experiments. All authors reviewed the results and approved the final version of the manuscript.
-
All data generated or analyzed during this study are included in this published article and its Supplemental information files.
This work was supported by the National Natural Science Foundation of China (32072539), Natural Science Foundation of Shandong Province (ZR2022JQ14 and ZR2022QC112) and Taishan Scholar Young Expert Program.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers for cDNA synthesis, amplification, and sequencing.
- Supplemental Fig. S1 Characteristics of MdABCG25 protein. (a) The hydropathicity of MdABCG25 protein. (b) The transmembrane domains of MdABCG25 protein. (c) The signal peptides of MdABCG25 protein. (d) The predicted phosphorylation sites of MdABCG25 protein.
- Supplemental Fig. S2 The predicted secondary structure of the MdABCG25 protein.
- Supplemental Fig. S3 The expression levels of genes related to cuticular wax biosynthesis, transport and transcriptional regulation in MdABCG25 transgenic Arabidopsis and Col-0. Data are mean ± SD of three independent replicates. Different lowercase letters indicate a significant difference at p < 0.05.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhou MM, Yu ZH, Gao HN, Li MR, Wu YT, et al. 2023. Ectopic expression of an apple ABCG transporter gene MdABCG25 increases plant cuticle wax accumulation and abiotic stress tolerance. Fruit Research 3:43 doi: 10.48130/FruRes-2023-0043 

Ectopic expression of an apple ABCG transporter gene MdABCG25 increases plant cuticle wax accumulation and abiotic stress tolerance
- Received: 14 September 2023
- Accepted: 17 November 2023
- Published online: 15 December 2023
Abstract: A number of enzymes and transcription factors involved in cuticular wax biosynthesis have been identified in numerous plant species. The pathway of wax biosynthesis is relatively definite. The molecular basis of cuticular wax deposition is still unclear, especially the transport mechanism of cuticular wax from endoplasmic reticulum (ER) to the cell wall. It has been found that the ABCG transporter family is involved in the wax export process in some model plants. However, whether ABCG transporters participate in wax transport in apple is unknown. In this study, MdABCG25, encoding an ABCG transporter, was isolated from apple ( Malus domestica Brokh.). MdABCG25, containing an AAA domain and an ABC2_membrane domain, is highly expressed in apple pericarp and induced by drought, salt and ABA conditions. The heterologous expression of MdABCG25 in Arabidopsis leads to an increasing cuticular wax accumulation of stems and leaves. The epidermal permeability, water loss rate and chlorophyll extraction rate of MdABCG25 transgenic Arabidopsis were decreased. The expression of MdABCG25 improved the drought and salt tolerance of apple calli and Arabidopsis.
-
Key words:
- Apple /
- MdABCG25 /
- Cuticular wax /
- Abiotic stress