









-
Ginkgo biloba is a relict plant originating from China that possesses high ornamental and medicinal value[1]. G. biloba is abundant in secondary metabolites in leaves and seeds, including flavonoids and terpenoids, which play a crucial role in human disease prevention[2−4]. Notably, G. biloba contains extraordinary terpenoids called ginkgolides and bilobalide, which are exclusive to this plant species and have not been found in any other organism[5]. Terpenoids are primarily five-carbon isoprenoids, classified as monoterpenes, sesquiterpenes, diterpenes, triterpenes, tetraterpenes and polyterpenes based on the number of isoprenoid units[6]. The ginkgolides in G. biloba are mainly diterpenes synthesized through the MEP (methylerythritol) and MVA (mevalonate) pathways[7].
The enzyme geranylgeranyl pyrophosphate synthase (GGPPS or GGPS) is a crucial enzyme in the synthesis of terpenoids via the MEP pathway[7, 8]. It catalyzes the formation of geranylgeranyl pyrophosphate (GGPP)[9, 10]. GGPP is a precursor of terpenoids, photosynthetic pigments (such as chlorophyll and carotenoids) and phytohormones (such as gibberellins)[11, 12]. These compounds play a significant role in plant growth. In Arabidopsis, several genes of the GGPPS family have been identified that are involved in terpenoids synthesis[13, 14]. For example, the AtGGPP11 gene regulates the synthesis of photosynthesis-related isoprenoids through the MEP pathway[15]. The genes SlGGPPS2 and SlGGPPS3 in tomato were found to produce carotenoids and terpenoids by providing GGPP[16]. Overexpression of the snapdragon GGPPS gene in tobacco has been associated with increased production of chlorophyll and gibberellin[17]. Similarly, when GGPPS is overexpressed along with other MEP pathway genes, it enhances the accumulation of taxadiene levels in B. subtilis[18]. Therefore, it is suggested that the GGPPS gene may be a key player in plant growth and terpenoid formation.
Several studies have reported that the GGPPS gene is involved in abiotic stresses. It was found that the GGPPS family gene in cotton can be induced by abiotic stresses and several hormonal signals[19]. In addition, the expression of GbGGPPS can be induced by UV irradiation, which leads to increased accumulation of ginkgolides content in G. biloba[20]. Furthermore, heterologous expression of the sweetpotato geranylgeranyl phyrophosphate synthase (IbGGPS) gene in Arabidopsis enhanced resistance to osmotic stress[21]. Moreover, the activity of LtuGGPPS2 promoter is induced by mechanical damage and hormones, indicating that LtuGGPPS2 plays a key role in stress response[22]. In W. somnifera, the WsGGPPS gene was indicated to be involved in abiotic stress and plant growth metabolic process[12]. The cis-acting elements of the BoGGPPS genes contain abiotic stress factors such as low temperature and drought), indicating that BoGGPPS can respond to abiotic stress[23]. Taken together, these studies suggest that GGPPS may play an important role in the response to abiotic stress.
Members of the GGPPS gene have been identified in G. biloba[24−26], but the function of the GGPPS gene in growth and development and salt stress is unknown. In this study, we performed the isolation, expression, and functional analysis of the GbGGPPS1 gene. The results of this study contribute to the understanding of GbGGPPS1 in growth and development and salt stress, and provide a relevant basis for future research.
-
The seeds of G. biloba variety 'Fozhi' were obtained from Yangzhou University, Jiangsu Province, China. Before planting, the G. biloba seeds were stratified with a sodium hypochlorite solution and then rinsed with water. The G. biloba seeds were planted in black square pots and placed in a light incubator for cultivation at a constant temperature of 25 °C with 16 h of light and 8 h of darkness. The selected samples were immediately frozen in liquid nitrogen and stored in a freezer at −80 °C.
Three-month-old G. biloba seedlings were selected for the NaCl treatment. The leaves of G. biloba were collected at 0, 1, 3, 5, and 7 d, frozen in liquid nitrogen, and stored at −80 °C until they were used.
Identification and bioinformatic analysis of GbGGPPS1
-
The sequence of GbGGPPS1 (evm.model.chr8.380) was obtained from the new G. biloba genome[27]. The physicochemical properties of GbGGPPS1 protein were analyzed using ExPASy online software (
https://web.expasy.org/protparam ). The secondary structure of GbGGPPS1 was analyzed using SOPM website (http://npsa-pbil.ibcp.fr/cgi-bin/npsa_automat.pl?page=npsa_sopma.html ). The promoter of GbGGPPS1 was analyzed using PlantCARE website and visualized using TBtools[28].The domain of GbGGPPS1 protein was analyzed using the NCBI Conserved Domain database (
https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi ) and visualized using the IBS (Illustrator for Biological Sequences) website (http://ibs.biocuckoo.org/online.php )[29]. The phylogenetic tree of the GGPPS protein sequence was generated by blasting from on the NCBI website and constructed using the neighbor-joining method in MEGA11 software.Quantitative real-time PCR (qRT-PCR)
-
Total RNA from G. biloba seedlings was extracted using the FastPure Universal Plant Total RNA Isolation Kit (Vazyme Biotech Co., Nanjing, China). First-strand cDNA was reverse transcribed using the HiScript 1st Strand cDNA Synthesis Kit (Vazyme Biotech Co., Nanjing, China). Quantitative real-time PCR (qRT-PCR) was performed using SYBR qPCR Master Mix reagents (Vazyme Biotech Co., Nanjing, China). The CFX96 Real-Time PCR Detection System (Bio-Rad) was used for qRT-PCR. Each treatment had three biological replicates and the relative expression levels were analyzed using the 2−ΔΔCᴛ method. Primer sequences are listed in Supplemental Table S1.
Generation of transgenic 35s::GbGGPPS1 Arabidopsis
-
The wild-type (WT) Arabidopsis accession Col-0 was used to generate transgenic plants. The coding sequence (CDS) of GbGGPPS1 was amplified and inserted into the pRI-GFP vector to generate the GbGGPPS1-pRI vector construct, following previously described methods[30]. To generate GbGGPPS1 overexpression Arabidopsis plants, the GbGGPPS1-pRI construct was transformed into Arabidopsis Col-0 through inflorescence dip to obtain 35s::GbGGPPS1 transgenic Arabidopsis. The transformed 35s::GbGGPPS1 Arabidopsis seeds were screened on Murashige and Skoog (MS) medium containing 50 μg/mL of kanamycin. The successfully screened Arabidopsis plants were then identified by PCR and qRT-PCR. Three transgenic plant lines of the T3 generation were used for further study. The number of days from snowing to the first flower bud formation was used to calculate the flowering time, and the number of rosette leaves at that time was counted (10 plants with three biological replicates).
Measurement of chlorophyll
-
Fresh plant samples weighing 0.5 g were immersed in 50 ml of 95% ethanol in the absence of light until the leaves showed no green color (approximately 16−24 h). After centrifugation, the absorbance of the supernatant was measured at wavelengths of 665 nm, 649 nm, and 470 nm, and the chlorophyll content was calculated[31].
Assessment of NaCl treatment for 35s::GbGGPPS1 Arabidopsis seeds
-
Seeds of both WT and transgenic Arabidopsis were placed on either MS medium or MS medium supplemented with 100 mM NaCl or 150 mM NaCl (30 plants with three biological replicates). The germination rates of the WT and transgenic Arabidopsis seeds were observed and counted daily for 8 d.
-
The full-length cDNA sequence of GbGGPPS was cloned from the G. biloba genome (Fig. 1a). Since it is the same protein as the previously amplified GGPPS protein (GenBank: AAQ72786.1)[24], it was named GbGGPPS1. The nucleic acid sequence of GbGGPPS1 is 1,176 bp, encoding 391 amino acids. The estimated molecular weight of the GbGGPPS1 protein is 42.51 kD and its theoretical pI is 5.98. Additionally, the GbGGPPS1 protein contains 51 negatively charged residues (Asp + Glu) and 45 positively charged residues (Arg + Lys). The instability index (II) was calculated to be 46.74, indicating that GbGGPPS1 is an unstable protein. Secondary structure analysis of the GbGGPPS1 protein revealed that it consists of 53.71% α-helices, 5.88% β-turns, 30.43% random coils and 9.97% extended strands (Supplemental Fig. S1). The conserved domain of GbGGPPS1 contains a Trans_IPPS_HT domain at positions 123–389 aa, indicating its function in isoprene biosynthesis (Fig. 1b). In addition, protein sequence analysis revealed that the GbGGPPS1 protein contains CxxxC, FARM, and SARM sequences (Fig. 1c), which are capable of binding allylic substrates and IPPs. As is shown in Supplemental Fig. S2, the GbGGPPS1 promoter can respond to salicylic acid (SA), MeJA, abscisic acid (ABA), and auxin.
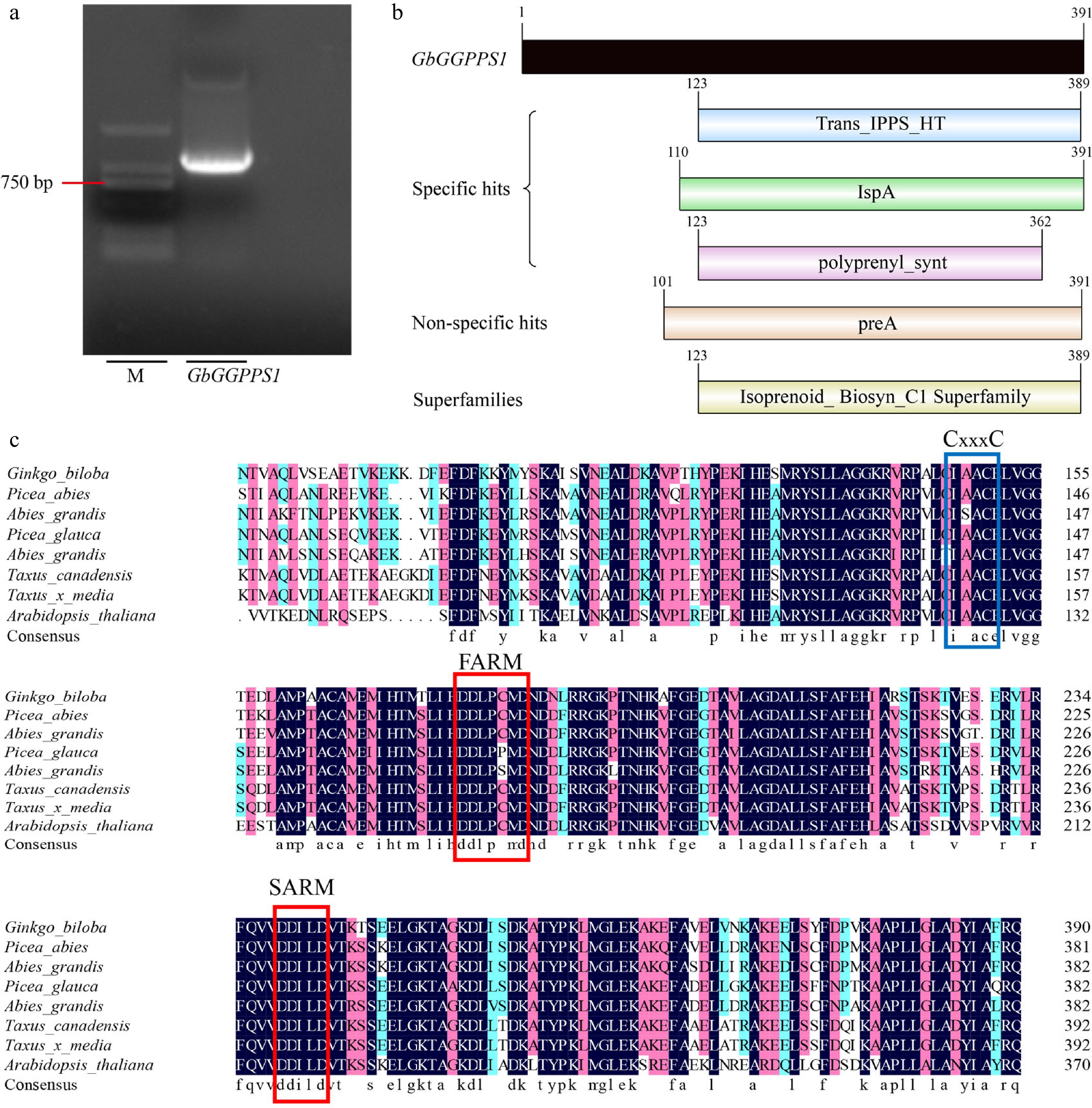
Figure 1.
Sequence analysis of GbGGPPS1. (a) The CDS sequence of GbGGPPS1 cloning. (b) The domain of GbGGPPS1 protein. (c) Homology analysis of GbGGPPS protein in different plants. FARM and SARM motifs are indicated by red boxes. CxxxC motifs are indicated by blue boxes.
To gain insight into the phylogenetic relationships of the GbGGPPS1 protein, we conducted a BLAST search in the NCBI using the GbGGPPS1 protein sequence. The similarity to the GGPPS proteins of Picea abies, Abies grandis, Picea glauca, Picea sitchensis, Abies grandis, Taxus canadensis, and Taxus x media was all greater than 70%. In addition, we used the GGPPS protein sequence of different plants to construct a phylogenetic tree using the neighbor–joining (NJ) method by MEGA (Supplemental Fig. S3). The resulting phylogenetic tree showed that GbGGPPS1 proteins clustered with gymnosperms and were distant from the Arabidopsis GGPPS protein (Fig. 2).
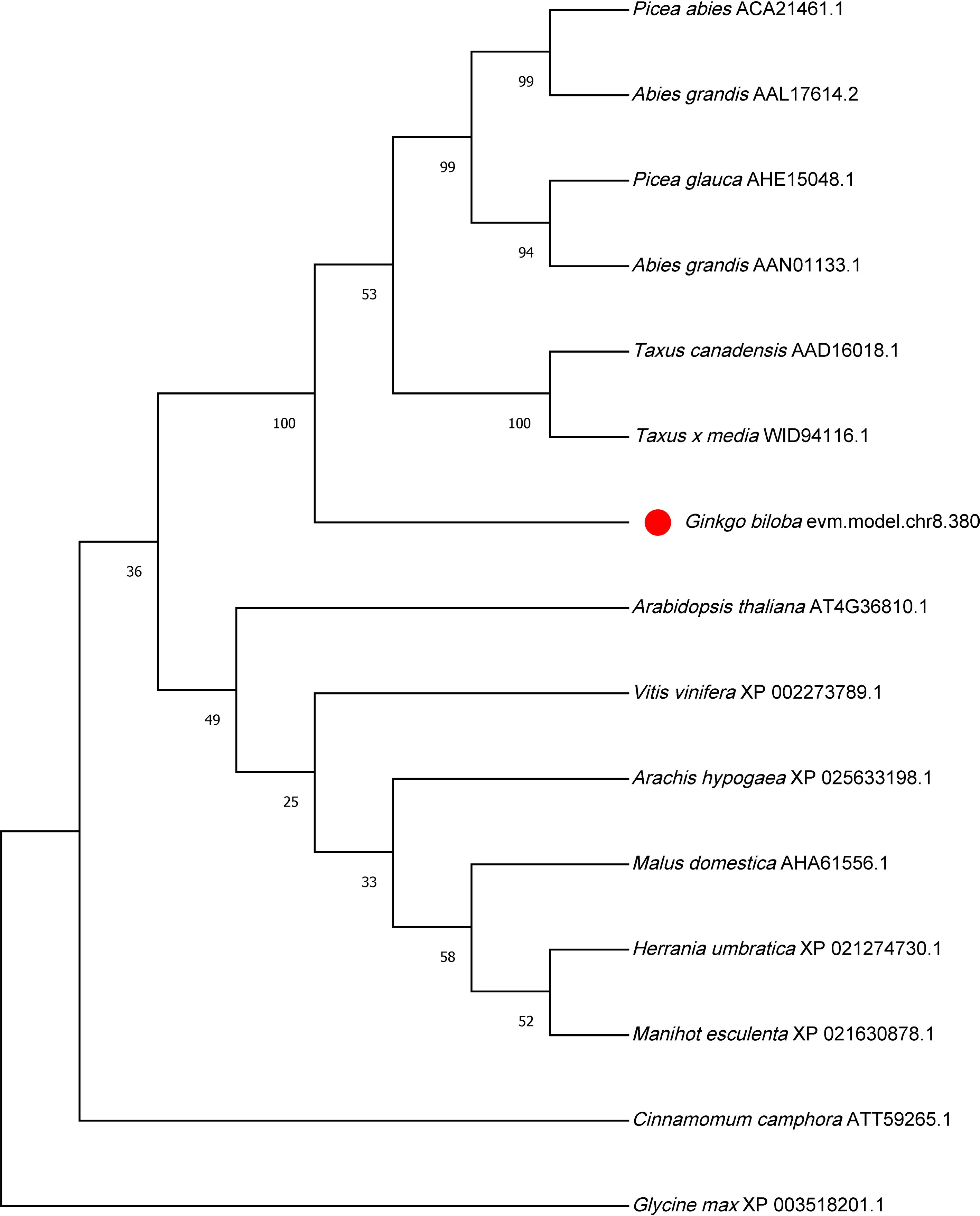
Figure 2.
Phylogenetic tree analysis GGPPS protein in different plants.
Expression of GbGGPPS1 in different tissue and salt stresses
-
To further investigate the specific expression of GbGGPPS1 in different tissues, we analyzed the expression levels in the root, stem, leaf, embryo, microstrobilus, and ovule of G. biloba. GbGGPPS1 expression was detected in all tissues, with the highest expression level in stem, leaf and embryo (Fig. 3a). We also examined whether salt stress affects the expression of the GbGGPPS1 gene. Leaves treated with 200 mM NaCl were used for this analysis. The results showed that the relative expression of GbGGPPS1 increased with the duration of salt treatment, peaked at 1 d, and then decreased (Fig. 3b). This indicates a positive correlation between GbGGPPS1 gene expression and salt stress.
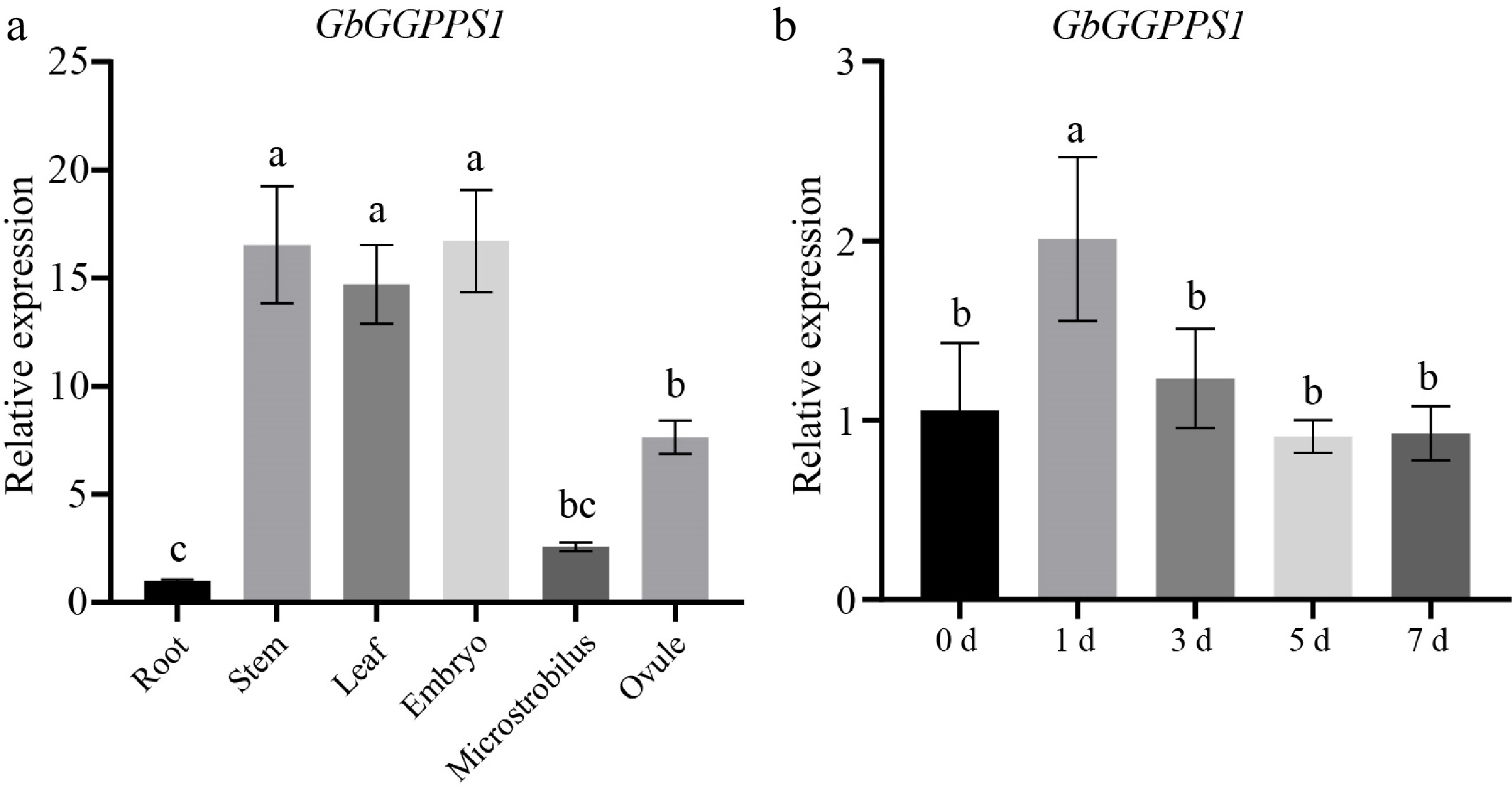
Figure 3.
Expression of the GbGGPPS1 gene in (a) different tissues and (b) salt stress. Data are means (± SD) of the three biological replicates.
Phenotypic and physicochemical of transgenic Arabidopsis
-
To further investigate the biological function of GbGGPPS1, we transferred the overexpression vector GbGGPPS1-pRI into Arabidopsis. We obtained three homozygous transgenic lines (35s::GbGGPPS1-1, 35s::GbGGPPS1-2, and 35s::GbGGPPS1-3) by PCR and qRT-PCR analysis (Fig. 4a, b). Compared to the WT, no significant differences in leaf development were observed in 35s::GbGGPPS1 Arabidopsis (Fig. 4c). Furthermore, the morphology of 35s::GbGGPPS1 Arabidopsis did not show any significant differences compared to WT at early stages (Fig. 4d). However, under long day conditions, the flowering process of transgenic Arabidopsis was accelerated, about 3 d earlier than that of WT (Fig. 4d, e). The number of rosette leaves was also reduced in transgenic Arabidopsis (Fig. 4f).
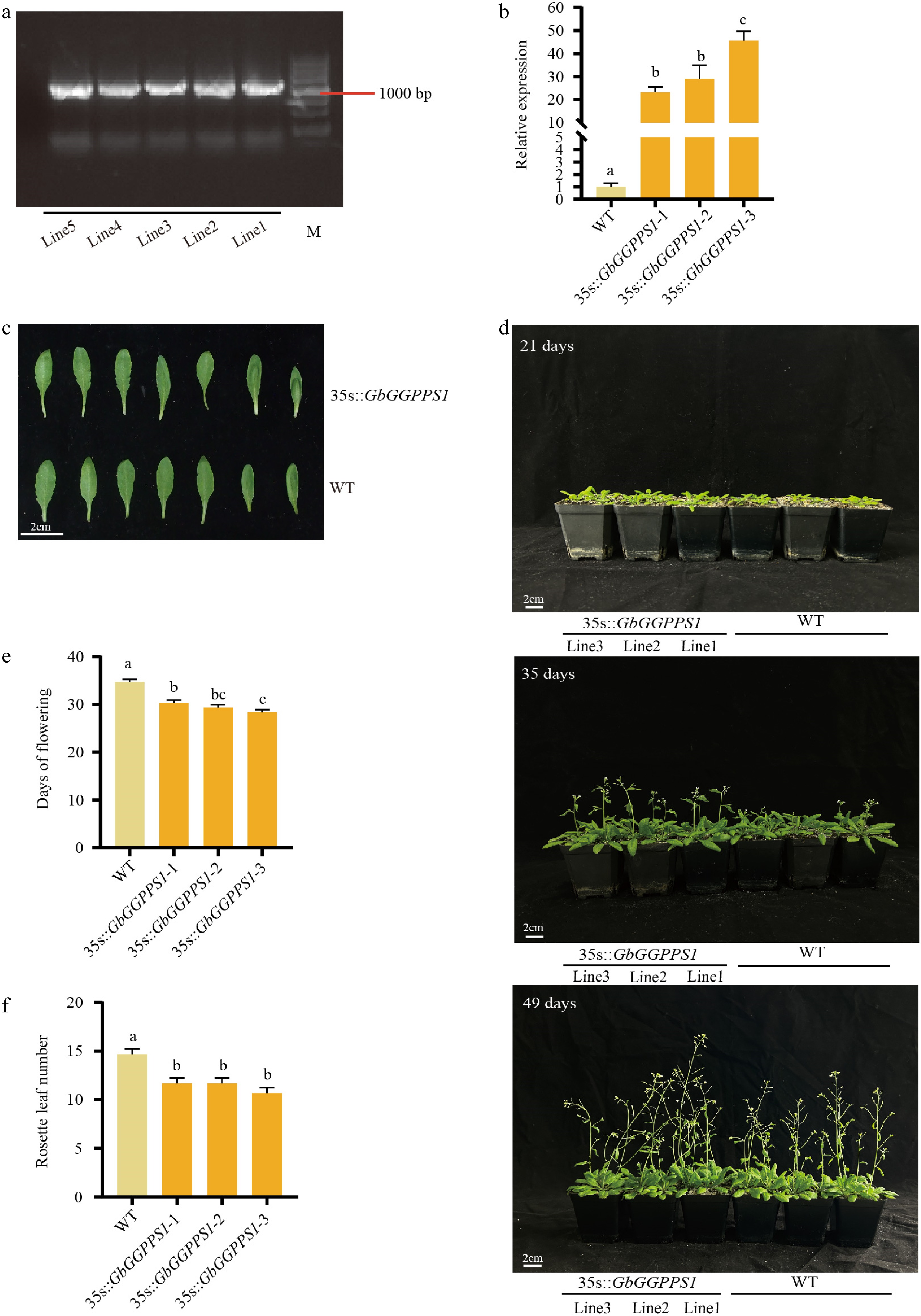
Figure 4.
Phenotypes of WT and transgenic Arabidopsis plants. Identification of transgenic plants by (a) PCR and (b) qRT-PCR. (c) Comparison of leaf phenotypes between WT and 35s::GbGGPPS1 Arabidopsis. (d) Phenotypes of WT and the 35s::GbGGPPS1 Arabidopsis at 21, 35, and 49 d. (e) Days to first flowering of WT and 35s::GbGGPPS1 Arabidopsis. (f) Number of rosette leaves of WT and 35s::GbGGPPS1 Arabidopsis.
The GbGGPPS1 expression profile in transgenic Arabidopsis
-
Since GGPPS is responsible for the synthesis of terpenoids, including chlorophyll, we measured the chlorophyll content in transgenic Arabidopsis. The results showed a significant increase in the levels of chlorophyll a, chlorophyll b, and carotenoid in transgenic Arabidopsis when compared to the WT (Fig. 5a−d). Since GGPP is a precursor of phytohormones (such as gibberellins), and to further investigate the potential mechanism of GbGGPPS1 in the flowering process, the GA synthesis and signaling gene and ABA signaling were selected. Compared with WT, the expression of AtRGL2 was significantly downregulated, while the expression levels of AtGID1B and AtGID1C were significantly upregulated. There was no difference in the expression of GA synthesis gene and ABA signaling gene (Fig. 5k, l).
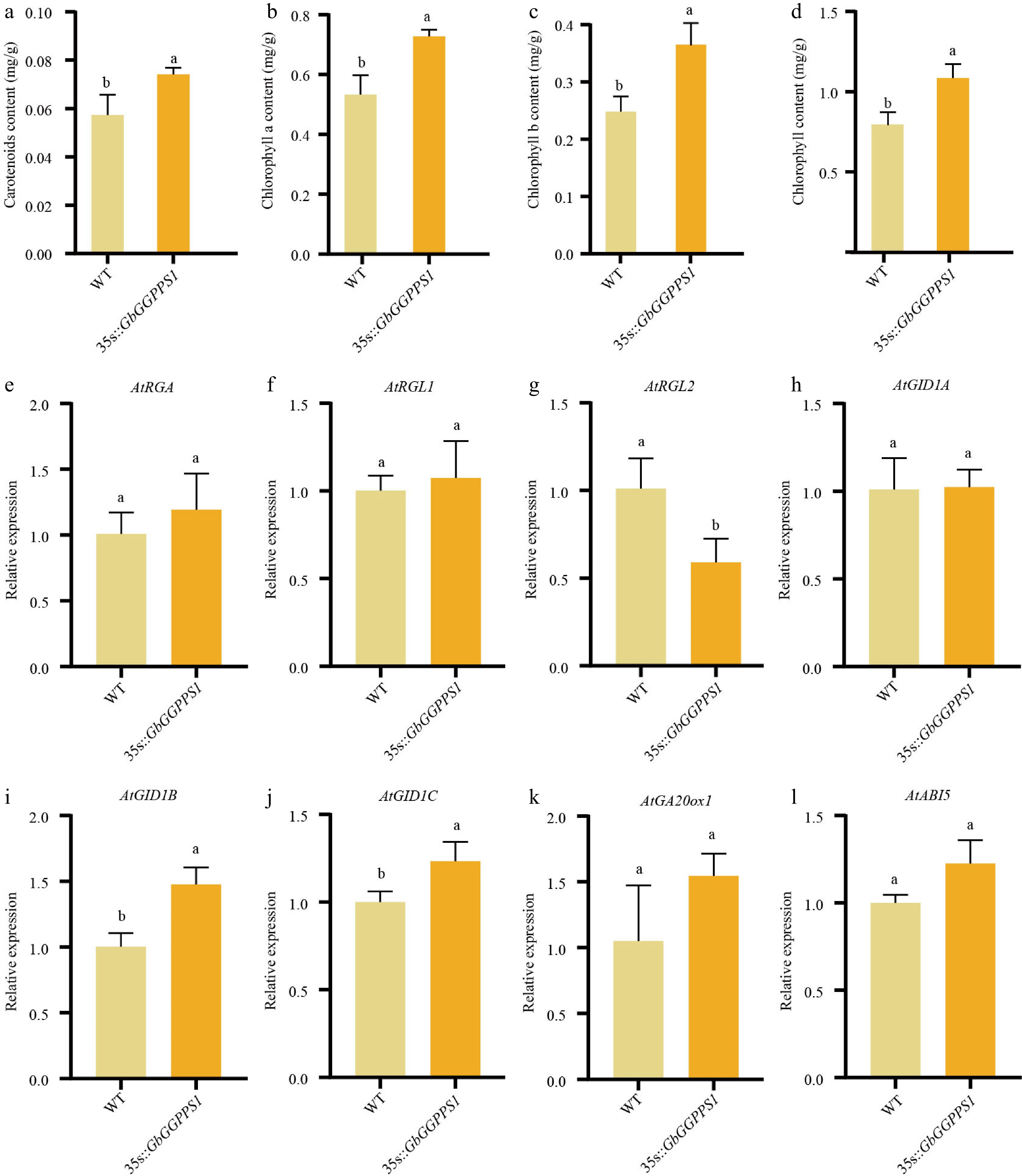
Figure 5.
Functional characterization of WT and transgenic Arabidopsis. (a)−(d) Chlorophyll content in WT and 35S::GbGGPPS1 Arabidopsis. (e)−(l) qRT–PCR analysis of the expression levels of GA- and ABA-associated genes in WT and transgenic Arabidopsis.
Overexpression of GbGGPPS1 can increase the salt tolerance in transgenic Arabidopsis
-
To investigate the function of GbGGPPS1 gene in salt stress, we placed WT and 35s::GbGGPPS1 Arabidopsis on MS medium containg 0 mM, 100 mM, and 150 mM NaCl to determine the germination rate. As shown in Fig. 6a & d, there was no difference in seed germination on MS medium after 8 d. However, when the MS medium contained 100 mM NaCl, the germination rate of 35s::GbGGPPS1 Arabidopsis was higher than that of WT (Fig. 6b, e). Similarly, 150 mM NaCl inhibited seed germination, but the germination rate of 35s::GbGGPPS1 Arabidopsis was over 80%, while that of WT was lower at 50% after 8 d (Fig. 6c, f). These results indicate that overexpression of GbGGPPS1 improves the tolerance of Arabidopsis to salt stress.
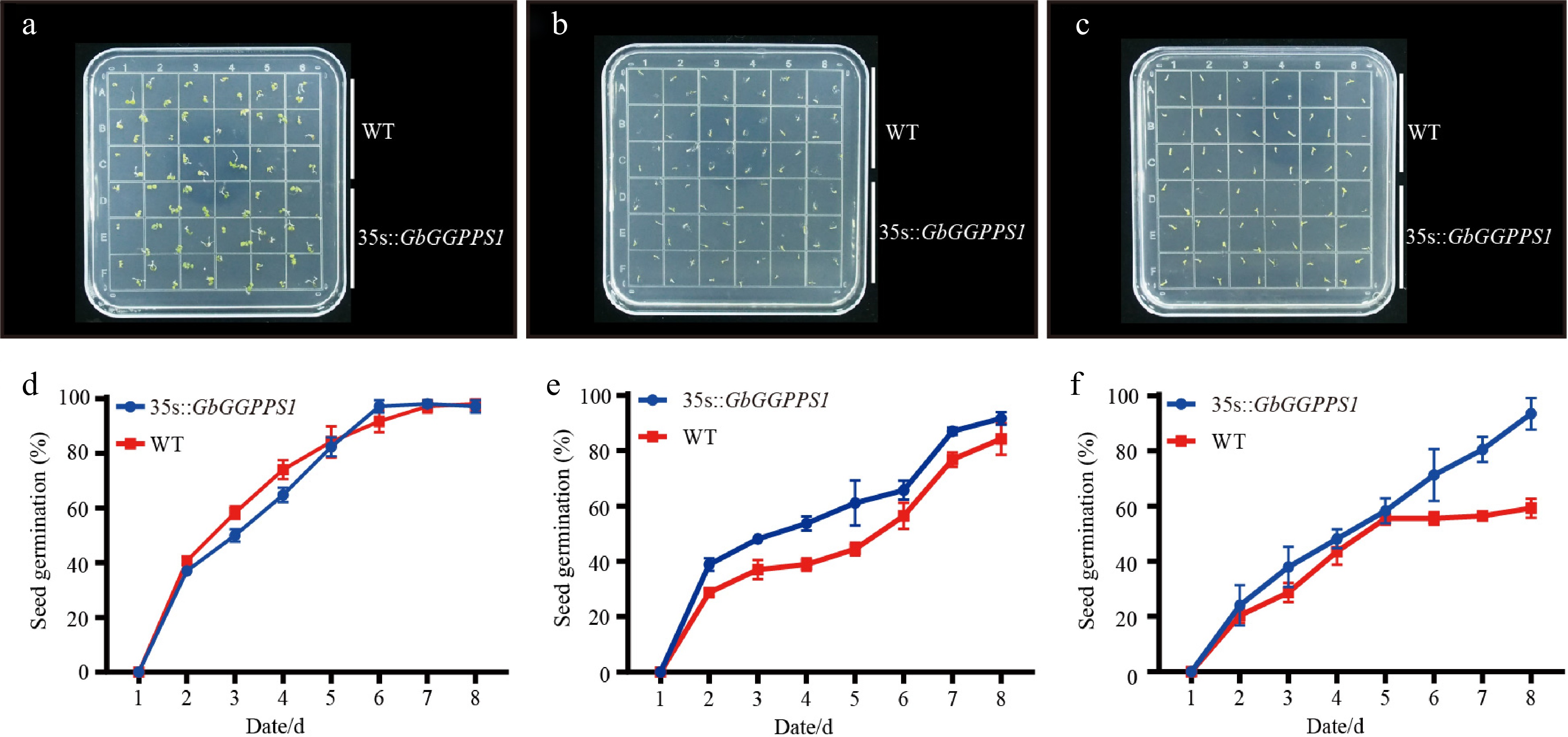
Figure 6.
Germination rates of Arabidopsis seeds treated with (a), (d) 0 mM NaCl, (b), (e) 100 mM NaCl, and (c), (f) 150 mM NaCl.
-
G. biloba is rich in secondary metabolites, including flavonoids and terpenoids, which are beneficial to human health. Terpenoids play an important role in plant growth and development[32]. GGPPS is a key enzyme in the synthesis of GGPP, which is a precursor of terpenoids and chlorophylls[22, 33]. GGPPS proteins contain FARM and SARM motifs that can bind to Mg2+ and pyrophosphate to catalyze the conversion of GGPP and GPP[22, 34]. In this study, we found that the GbGGPPS1 protein also contains the FARM and SARM motifs (Fig. 1c), indicating that it belongs to the GGPPS protein family. The CxxxC sequence is recognized as a necessary domain for protein interaction[22], and GbGGPPS1 also possesses this domain (Fig. 1c). We speculate that GbGGPPS1 functions by interacting with other proteins in G. biloba. The phylogenetic tree shows that GGPPS1 from G. biloba is clustered with the gymnosperm plants GGPPS protein group (Fig. 2). Moreover, previous studies have shown that gymnosperms are capable of producing terpenoids, such as Taxus, which is capable of producing paclitaxel[35]. Therefore, we hypothesize that GbGGPPS1 from G. biloba is involved in terpenoid synthesis.
The GGPP generated by GGPPS is involved in the synthesis of gibberellins, chlorophylls, and terpenoids in plants[11, 36]. Additionally, gibberellin signaling plays a partial role in promoting flowering[37]. Here, gene expression analysis revealed the upregulation of AtGID1B and AtGID1C and downregulation of AtRGL2 in the 35s::GbGGPPS1 Arabidopsis (Fig. 5g, i, j), suggesting that GbGGPPS1 regulates flowering by regulating these GA signaling genes. Heterologous expression of GGPS (also known as GGPPS) in tobacco enhances early flowering and growth[38]. In this study, we demonstrated that the heterologous expression of GbGGPPS1 in Arabidopsis enhanced growth, development, and early flowering (Fig. 4d). These phenotypes may be due to the overexpression of GbGGPPS1, which promotes GGPP synthesis and, at least in part, increases gibberellin content, thereby enhancing growth and development and early flowering in transgenic Arabidopsis. Notably, overexpression of the GGPPS gene in Arabidopsis, Nicotiana tabacum, and Nelumbo nucifera has been shown to increase chlorophyll content[33, 39, 40]. Furthermore, the GGPPS family genes in Arabidopsis, with AtGGPPS11 as the only hub isozyme, form an enzyme complex with the downstream protein geranylgeranylgeranyl reductase (GGR), which is involved in chlorophyll biosynthesis[15, 41]. In this study, overexpression of GbGGPPS1 in Arabidopsis was found to increase the chlorophyll content (Fig. 5a−d). In addition, GGPPS was considered as a candidate gene for terpenoid biosynthesis in G. biloba[26, 42]. These results indicated that GbGGPPS1 are responsible for terpenoids and is involved in plant growth and development.
Although GGPPS genes have been reported to be involved in plant growth and development, only a few studies have investigated their function under different abiotic stresses. In cotton, the GGPPS family genes can be induced in response to different stress conditions, and the GhGGPPS8 gene responds specifically to NaCl stress[19]. Similarly, the presence of stress elements in the LtuGGPPS2 promoter suggests that LtuGGPPS2 may play a significant role in the stress response[22]. In tomato, the GGPP synthase gene SlG1 enhances bacterial infection by producing GGPP for the production of defensive diterpenoids in leaves[43]. The GbGGPPS gene from G. biloba was found to be induced by cold, ABA, SA, and MEJA, indicating its important role in stress response[20]. In this study, we found that GbGGPPS1 can be induced by salt treatment (Fig. 3b). Additionally, we found that the transgenic 35s::GbGGPPS1 Arabidopsis exhibited enhanced seed germination under salt treatment (Fig. 6). Furthermore, the GbGGPPS1 promoter contains cis-acting elements associated with SA, MeJA, and ABA (Supplemental Fig. S2), suggesting its involvement in salt stress. Previous studies have shown that GA accumulation can promote salt tolerance[44, 45]. The higher expression of AtGID1B and AtGID1C indicated the accumulation of GA in the transgenic plant, suggesting that GbGGPPS1 could enhance salt stress tolerance through GA signaling. However, the functional mechanism of how GbGGPPS1 enhances salt tolerance needs further investigation.
-
In summary, we cloned and performed a bioinformatics analysis on the G. biloba GbGGPPS1 gene. Our findings indicate that GbGGPPS1 is a protypical GGPPS protein. Overexpression of GbGGPPS1 in Arabidopsi was found to stimulate plant growth and development, increase chlorophyll content, and enhance salt stress resistance.
-
The authors confirm contribution to the paper as follows: designed the experiments, wrote and revised the manuscript: Wang L, Wang Q; performed the experiments, analyzed the experimental data: Wang Q, Zhao X, Jiang Y. All authors reviewed the results and approved the final version of the manuscript.
-
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
This research was supported by National Natural Science Foundation of China (Nos 31971686, 32171838 and 32101558) and Natural Science Foundation of Jiangsu Province (No. BK20210801).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Primers were used in this study.
- Supplemental Fig. S1 Secondary structure of GbGGPPS1 protein.
- Supplemental Fig. S2 Putative cis-acting regulatory elements in the GbGGPPS1 promoter.
- Supplemental Fig. S3 The GGPPS protein sequence in different plant.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wang Q, Zhao X, Jiang Y, Wang L. 2023. Isolation, expression and functional analysis of the GbGGPPS1 gene in Ginkgo biloba. Fruit Research 3:44 doi: 10.48130/frures-0023-0044 

Isolation, expression and functional analysis of the GbGGPPS1 gene in Ginkgo biloba
- Received: 13 September 2023
- Accepted: 13 November 2023
- Published online: 21 December 2023
Abstract: Ginkgo biloba is known for its abundance of secondary metabolites, such as terpenoids. However, the identification and functional analysis of key genes in the terpenoid biosynthetic pathway are limited. Geranylgeranyl pyrophosphate synthase (GGPPS) is a crucial enzyme involved in the production of terpenoids. In this study, we successfully cloned the GbGGPPS1 gene from G. biloba. The length of GbGGPPS1 was 1,176 bp, encoding 391 amino acids. The GbGGPPS1 protein contains the GGPPS protein domain, including the FARM and SAR motifs. Phylogenetic analysis revealed that GbGGPPS1 is closely related to gymnosperms. Furthermore, the expression of GbGGPPS1 was higher in leaves, stems, and ovules compared to the roots. Additionally, overexpression of GbGGPPS1 in Arabidopsis increased chlorophyll content and accelerated flowering process, indicating its role as a terpenoid synthase gene and plant development. Notably, the expression of GbGGPPS1 can be induced by salt stress, and overexpression of GbGGPPS1 in Arabidopsis exhibited enhanced seed germination under salt treatment. These results indicate that GbGGPPS1 plays an important role in plant growth, development, and salt stress resistance.
-
Key words:
- Ginkgo biloba /
- GbGGPPS1 /
- Growth and development /
- Salt stress