








-
The rising sea-levels caused by global warming poses a major threat to coastal regions due to salt-water intrusion and damage to agricultural land[1]. Over the past 20 years, the sea level has risen by about 8 cm[2]. Based on a new digital elevation model (CoastalDEM) utilizing neural networks, Kulp & Strauss found that the rising sea levels and coastal flooding have tripled as compared to the previous estimate by Shuttle Radar Topography Mission (SRTM)[3]. About 1 billion people occupy land lying less than 10 m above current high tide lines, most of them living in the Asia and Pacific region, mainly in China, Bangladesh, India, Vietnam, Indonesia, Thailand, Philippines and Japan[3]. The latest global soil salinity map of 2016 showed that the total area of salt affected land is around 1 billion hectares, an increase of more than 100 Mha from 1986[4]. Tidal flats occupy an area of about 124,286−131,821 km2 globally, with about 11.27% in Indonesia, 9.42% in China, 6.93% in Australia, 4.52% in India and 2.59% in Myanmar[5].
Increased area of saline soils including large areas of tidal flats have significant impacts on the natural environment and ecosystems as well as socio-economic impacts[6]. High salinity levels in the soil harm plant growth and limit crop yields. Currently, most of the salt-affected soils, especially coastal saline soils, have been deserted by many farmers leaving extensive areas of idle and unproductive land. This has posed a big challenge to researchers and farmers on how best to fully utilize these saline areas.
Among the various techniques to reclaim saline soil, phytoremediation is generally preferred compared to e.g. hydro or electro reclamation as it is sustainable and economically viable[7]. Phytoremediation involves the use of plants and associated soil micro-organisms to reduce the harmful effects of contaminants in the environment[8,9]. For instance, legumes make net nitrogen inputs into ecosystems, and have other multiple environmental benefits by improving soil structure and microbial activity of coastal saline land, and fundamentally improve saline soil[10]. Salinity tolerant legumes offer the double advantage of reclaiming degraded coastal agroecosystems and providing a local source of livestock feed. The latter is particularly relevant in highly populated countries mainly depending on feed imports, which is often the case in the Asia-Pacific region[11]. In terms of livestock feed, salinity tolerant legumes improve the quantity and quality of available feed, and have a high nutritional value with high protein content. When grown in areas that are not suitable for crops, they allow farmers to make the most of available land, provide increased income and diversify livelihoods of coastal farmers[12].
Salinity tolerant legumes grow naturally in many coastal countries, and are receiving increasing attention from researchers and developers due to their adaptation capacities in a range of salinity and drought conditions, and their potential economic benefits[13,14]. Fully mining and using these plants to improve coastal saline areas and tidal flats is an effective means to overcome impacts of seawater encroachment and increase the livelihoods of 1 billion coastal smallhold farmers[3].
However, there has been no attempt so far to synthesize knowledge on salinity tolerant legumes and promote their use in coastal ecosystems. Collections and selections have been scattered, mainly in Colombia, China, Australia, India etc.[15,16]. International Center for Tropical Agriculture (CIAT) (
www.tropicalforages.info/text/intro/index.html ), Chinese Academy of Tropical Agricultural Sciences (CATAS)[17], Commonwealth Scientific and Industrial Research Organization (CSIRO) and The International Livestock Research Institute (ILRI) have engaged in the collection and preservation of forage germplasm resources from 1970[18], but the quantity of salinity tolerant legumes are still low. There is an urgent need to systematically review available coastal forage legume plant resources in order to provide theoretical guidelines for the improvement of coastal saline land. Therefore, the objectives of this study were to (i) review currently available salinity tolerant forage legume species, (ii) discuss their potential for integration into smallholder mixed coastal systems of the Asia-Pacific region, and (iii) present the most promising species in more detail. -
Great breakthroughs have been made in the research on the mechanism of plant salinity tolerance, mainly focusing on model plants Arabidopsis thaliana and crops[19]. It's clear that salinity tolerance mechanisms of plants are complex traits involving multiple phases and genes[20]. At the cellular level, salinity tolerant plants maintain this ion balance by excreting Na+ out of the cell or compartmentalizing Na+ into the vacuole and accumulate osmolytes (such as K+, Ca2+, proline, soluble sugar, polyamines etc.) in the cytoplasm. Glycosyl inositol phosphorylceramide (GIPC) sphingolipids in the plasma membrane act as Na+ receptors for sensing Na+ in the apoplastic space and then gate Ca2+ influx channels in plants[21]. With the increase of Ca2+, the salt overly sensitive (SOS) signaling pathways (SOS1, SOS2, SOS3) will be activated and play a leading role in mediating the excretion of Na+ in plants[19]. Many other pathways such as MAPK and salinity tolerance related genes (including transcription factors) will also be activated to deal with salinity stress through osmoregulation, ion transport and radical-scavenging. At the tissue or organ level, some salinity tolerant plants usually store Na+ in the roots instead of above ground, or old leaves instead of young leaves[22].
For the salinity tolerant legumes, many of them are belonging to halophytes (plants that survive to complete their life cycle in at least 200 mM salinity)[23−25], besides the common salinity tolerant mechanisms similar to Arabidopsis thaliana and crops, many of them have special salinity tolerant mechanisms due to their different morphological structure[26]. Big gaps on the study of salinity tolerant mechanisms based on the whole genome sequencing and lack of studies about the function of special salinity tolerant genes.
Rhizobia level
-
Rhizobia is a special group of microbes with the unique ability to form symbiotic association with legumes to fix atmospheric nitrogen, a process known as Biological Nitrogen Fixation (BNF)[27]. Rhizobia-legume association takes place in specialized root structures called nodules, where rhizobia convert atmospheric gaseous dinitrogen (N2) into ammonia, a form that can be assimilated by the plants; in exchange, the host plant provides carbon compounds to rhizobia[28−30] . For centuries, this symbiotic association has been used in cropping systems as the most important N-fixing agent[31,32].
Salt stress can inhibit rhizobia-legume symbiosis by reducing rhizobia growth, nodule formation and BNF due to osmotic stress and high toxic levels of Na+ and Cl- in the cells[33]. Different rhizobia species exhibit varying levels of tolerance to salt stress. Fast-growing rhizobia species are categorized as more salinity-tolerant e.g. Sinorhizobium meliloti[34], S. fredii[35], Mesorhizobium huakuii and Rhizobium tropici[36] as compared to slow-growing rhizobia such as R. leguminosarum bv. viciae[37]. Some rhizobia strains have one or several high tolerant abilities to abiotic stress such as high temperature, pH, salinity and drought[38].
Previous studies have reported rhizobia to be more tolerant to salinity as compared to the host legumes plants, and can enhance tolerance of legumes to salt stress and yield[39−41]. Salinity-adaptive responses in rhizobia include accumulation of osmoprotectants, enhanced production of exopolysaccharides, expression of stress genes and ROS-scavenging enzymes such as superoxide dismutase[42,43]. Intracellular accumulation of osmoprotectants such as glycine betaine, glutamate, choline and N-acetyl glutaminyl glutamine amide (NAGGN) have been correlated with salt stress tolerance in Rhizobium meliloti[44,45]. Salinity tolerance of rhizobia is also related to hormones and protein changes. For example, increased salinity tolerance of Bradyrhizobium strain RJS9-2 may be achieved by production of indole-3-acetic acid (IAA), protein expression and osmoprotectant accumulation[46]. Salinity tolerant rhizobia may be due to a plasmid-mediated resistance since salinity resistance can be rapidly transferred from tolerant to sensitive rhizobia[38].
Biofertilizers containing rhizobia are an environmentally friendly approach to enhance soil fertility and legume productivity. Establishment of salinity-tolerant legumes combined with inoculation using salinity-tolerant rhizobia strains is a promising strategy for forage legume production and reclamation of saline soils. Generally, farmers can purchase commercial rhizobia inoculants to apply to the legumes. However, most countries in Asia rely on imported biofertilizers as only few rhizobia inoculants are registered or available in the market, the majority being of poor quality. For instance, by 2018, only 1% of biofertilizers/inoculants registered in China contained rhizobia, mainly produced for Chinese milkvetch (Astragalus sinicus), soybean and peanuts[33]. In Vietnam, there are very few rhizobia inoculants available in the market[47], while in Cambodia and Lao PDR, low availability and adoption of rhizobia inoculants have been reported[33]. This shows a gap and a huge need to develop and promote the use of rhizobia inoculants including salinity-tolerant rhizobia inoculants in this context. Selection of high salinity tolerant legumes and the application of legume-specific, high salinity tolerant rhizobia inoculants has immense potential for successful rhizobium-legume symbiosis and increased N inputs in saline soils.
-
The current known salinity tolerant forage legume species and potentially salinity tolerant species in the tropical regions are summarized in Table 1. Only one species Melilotus indicus (L.) All. is an annual plant, five species are annual or perennial plants, 38 species are perennial plants. Twenty five percent of the 44 species are shrubs, 52% of the 44 species are herbs, 6.8% of the 44 species are trees, the rest have more than one growth habit. No variety was available in the 21 species (47.7% of the total species), indicate that breeding of salinity tolerant legumes needs to be strengthened in the future (Table 1).
Table 1. General information of current salinity tolerant forage legume species.
No. Scientific name Common name Variety Distribution* Life cycle Growth habit 1 Abrus precatorius L. Rosary pea ― Tropical zones Perennial Shrub 2 Acacia dealbata Link Mimosa Dealbata Link, 1846; Mackayana Seem. World wide Perennial Shrub or tree 3 Acacia nilotica (L.) Willd. ex Delile Gum arabic tree cupressiformis Africa Perennial Tree 4 Alysicarpus vaginalis (L.) DC. Alyce Clover, Buffalo clove Nummularifolius (DC.) Miq., Parvifolius Verdc., stocksii Baker taiwanianus S.S. Ying, Vaginalis, venosa (Blat.& Hall.) A. Pramanik & Thoth. Tropical zones Perennial Herb 5 Arachis pintoi Krapov. & W.C.Greg. Pinto Peanut Belomonte, Reyan No.12 Tropical zones of South America Perennial Herb 6 Cajanus cajan (L.) Millsp. Pigeon pea, gungo pea Phule Tur-12, Babati White, bicolor DC., Cajan, flavus DC. Tropical zones Perennial Shrub 7 Calopogonium mucunoides Desv. Calopo, wild ground nut ― Tropical zones Perennial Herb 8 Canavalia ensifomis (L.) DC Jack bean, horse bean Coriacea Domin, Ensiformis, Normalis Kuntze Tropical zones Perennial Herbaceous vine 9 Canavalia rosea (Sw.) DC. Bay bean ― Tropical zones Perennial Herbaceous vine 10 Cassia pumila Lam. ― ― Tropical zones in China, India, Malaysia and Australia Perennial Shrub 11 Centrosema pubescens Benth. Butterfly Pea Centrosema pubescens Benth. Jinjiang (2019, CATAS) Tropical zones Perennial Herbaceous vine 12 Clitoria ternatea Linn. Sp. Pl. Asian pigeonwings, blue clitoria, butterfly pea Angustifolia Hochstetter ex Baker, major Paxton, Pleniflora Fantz, Ternatea Tropical zones Perennial Herbaceous vine 13 Crotalaria albida Heyne ex Roth. Taiwan crotalaria albida, Gengmanensis (Z. Wei & C.Y. Yang), Kangrensis A.A. Ansari Southern Asia Perennial Herbaceous 14 Crotalaria bractaeata Roxb. ex DC. ― ― Southern Asia, America Perennial Herb or shrub 15 Crotalaria ferruginea (Grah.) Benth. Rust-color crotalaria ― Southern Asia, America Perennial Herb 16 Crotalaria retusa Linn. Large yellow rattlebox, rattleweed Indica Nampy & Sibichen, retusa, Tunguensis (Lima) Polhill Tropical zones Perennial Shrub 17 Dendrolobium triangulare(Retz.) Schindl. ― ― Southern Asia Perennial Shrub 18 Desmanthus virgatus (L.) Willd. Wild tantan, Hedge lucerne Glandulosus (L.) Willd.,1806; Depressus (Humb. & Bonpl. ex Willd.) B.L. Turner; Virgatus Tropical zones Perennial Herb or shrub 19 Erythrina corallodendron L. Cutlass Bush Bicolor Krukoff, Connata Krukoff, Corallodendron L,1753 Sporadic spread in tropical zones Perennial Tree 20 Galactia elliptifoliola Merr. ― ― Hainan, China Perennial Herbaceous vine 21 Indigofera chuniana Metc. ― ― Hainan, China Perennial Herb or shrub 22 Indigofera enneaphylla Linn. Mant. ― ― Hainan, China, Indonesia, Papua New Guinea, Australia Annual or perennial Herb 23 Indigofera galegoides DC. ― ― Southern Asia Perennial Shrub 24 Indigofera hirsuta Linn. Hairy Indigo ― Tropical zones Annual or perennial Herb 25 Indigofera litoralis Chun & T.C.Chen ― ― Tropical zones of China Perennial Herb 26 Indigofera suffruticosa Mill. Anil Indigo Canescens (J.A. Schmidt) Lobin, Suffruticosa Tropical zones Perennial Shrub 27 Leucaena leucocephala (Lam.) de Wit. Leucaena, Lead Tree, Cassie Reyan No. 1 Tropical zones Perennial Shrub or tree 28 Macroptilium atropurpureum (L.) Urb. Purple Bean Siratro Tropical zones Annual or perennial Herb 29 Melilotus indicus (L.) All. Annual Melilot, Indian sweet-clover, Indicus, prostratus P.C. Palau, Tommasini (Jord.) O.E. Schulz World wide Annual Herb 30 Melilotus officinalis (L.) Pall. Yellow sweet-clover, common yellow melilot ― World wide Perennial Herb 31 Melilotus siculus (Turra) B.D.Jacks. ― Messina, Neptune Europe and
northern AustraliaAnnual or perennial Herb 32 Pongamia pinnata (L.) Pierre Indian Beech, Pongam tree Hannii Domin, minor (Benth.) Domin, pinnata, Typica Domin Southern Asia and northern Australia Perennial Tree 33 Pycnospora lutescens (Poir) Schindl. ― ― Southern Asia and northern Australia Perennial Herb or shrub 34 Senna bicapsularis (L.) Roxb. Christmasbush Augusti (Harms) H.S. Irwin & Barneby, Bicapsularis Tropical zones of south America and southern Africa Perennial Shrub 35 Sesbania cannabina (Retz.) Pers. Yellow Pea Bush, Dhaincha Sericea Southern Asia and northern Australia Annual or perennial Herb or shrub 36 Sesbania rostrata Bremek. & Oberm ― ― Africa Annual or perennial Herb 37 Sesbania sesban (L.) Merr. Common Sesban Concolor (Wight & Arn.) Baquar; Bicolor (Wight & Arn.) FW. Andrews; Nubica Chiov; sesban (L.) Merr,1912; Zambesiaca J.B. Gillett Africa, India,
southern America
and ChinaPerennial Shrub 38 Stylosanthes guianensis (Aubl.) Sw. Stylo Reyan No. 20, 21, 22, 24, 25 Tropical zones of Africa, Asia-pacific region and south America Perennial Shrub 39 Swainsona formosa Sturt's Desert Pea ― Australia Perennial Herb 40 Tephrosia purpurea (Linn.) Pers. Syn. Pl. Sarphonk, wild indigo Angustissima B.L. Rob., Brevidens Benth., Elongata Craib, Gracilis Tackholm & Boulos, Leptostachya (DC.) Brummitt, Pubescens Baker, Queenslandica Domin, sericea Benth., Yunnanensis Z. Wei Tropical zones Perennial Herb 41 Teramnus labialis (Linn.f.) Spreng. Blue wiss Abyssinicus (Hochst. ex A. Rich.) Verdc., Acutus Verdc., Arabicus Verdc., Labialis, Somalensis Vatke Tropical zones Perennial Herb 42 Trifolium fragiferum L. Strawberry clover Salina Subtropical zones
of north America, Europe, east Asia, southern Australia and New ZealandPerennial Herb 43 Uraria lagopodiodies (Linn.) Desv.ex DC. ― ― Southern Asia and northern Australia Perennial Shrub 44 Vigna marina (Burm.) Merr. Beachpea ― tropical zones Perennial Herbaceous vine * Obtained from www.gbif.org/species There is quite a rich diversity of salinity tolerant species in coastlines of tropical regions, with six different species identified as high salinity tolerant (Table 2, No. 9, 29, 32, 34, 35, 44), 17 as moderate salinity tolerant (Table 2, No. 1−7, 11−12, 16, 30−31, 36−39, 42) and 21 species predicted as salinity tolerant but need to be further proven (Table 2, No. 8, 10, 13−15, 17−28, 33, 40−41, 43).
Table 2. Tolerances of current salinity tolerant forage legume species.
No. Scientific name Salinity tolerant Drought tolerant Acid
tolerantShade
tolerantWaterlogging
tolerantFrost
tolerantReference 1 Abrus precatorius L. X X X ― ― ― [55] 2 Acacia dealbata Link X ― X ― ― X [7] 3 Acacia nilotica (L.) Willd. ex Delile X X ― ― ― X [25] 4 Alysicarpus vaginalis (L.) DC. X X X ― ― ― https://extension.msstate.edu/content/alyceclover-alysicarpus-vaginalis 5 Arachis pintoi Krapov. & W.C.Greg. X X X X ― ― www.tropseeds.com/arachis-pintoi 6 Cajanus cajan (L.) Millsp. X X X ― ― ― [56] 7 Calopogonium mucunoides Desv. X X ― ― ― ― [57] 8 Canavalia ensifomis (L.) DC potential* X X ― X ― pfaf.org/user/Plant.aspx?LatinName=Centrosema+pubescens 9 Canavalia rosea (Sw.) DC. X X X X ― ― ― [58] 10 Cassia pumila Lam. Potential* X X ― ― ― [59] 11 Centrosema pubescens Benth. X ― ― X X ― [60] 12 Clitoria ternatea Linn. Sp. Pl. X X ― X ― X [61] 13 Crotalaria albida Heyne ex Roth. Potential* X X ― ― ― https://baike.baidu.com/item/%E5%93%8D%E9%93%83%E8%B1%86/4860050?fromModule=lemma-qiyi_sense-lemma 14 Crotalaria bractaeata Roxb. ex DC. Potential* ― ― ― ― ― ― 15 Crotalaria ferruginea (Grah.) Benth. Potential* ― ― ― ― ― ― 16 Crotalaria retusa Linn. X X ― ― ― ― [62,63] 17 Dendrolobium triangulare(Retz.) Schindl. Potential* X X ― ― ― [64] 18 Desmanthus virgatus (L.) Willd. Potential* ― ― ― ― ― ― 19 Erythrina corallodendron L. Potential* ― X X ― X https://baike.baidu.com/item/%E9%BE%99%E7%89%99%E8%8A%B1/3567229?fr=kg_general 20 Galactia elliptifoliola Merr. Potential* ― ― ― ― ― ― 21 Indigofera chuniana Metc. Potential* X ― ― ― ― https://baike.baidu.com/item/%E7%96%8F%E8%8A%B1%E6%9C%A8%E8%93%9D/1483435?fr=kg_general 22 Indigofera enneaphylla Linn. Mant. Potential* X ― ― ― ― https://baike.baidu.com/item/%E4%B9%9D%E5%8F%B6%E6%9C%A8%E8%93%9D/7264010?fr=aladdin 23 Indigofera galegoides DC. Potential* ― ― ― ― ― ― 24 Indigofera hirsuta Linn. Potential* X ― ― X ― https://baike.baidu.com/item/%E7%A1%AC%E6%AF%9B%E6%9C%A8%E8%93%9D?fromModule=lemma_search-box 25 Indigofera litoralis Chun & T.C.Chen Potential* X ― ― X ― https://baike.baidu.com/item/%E6%BB%A8%E6%B5%B7%E6%9C%A8%E8%93%9D/7181556?fr=aladdin 26 Indigofera suffruticosa Mill. Potential* ― X ― ― ― [65] 27 Leucaena leucocephala (Lam.) de Wit. Potential* X ― ― ― ― [25] www.nparks.gov.sg/florafaunaweb/flora/3/4/3471 28 Macroptilium atropurpureum (L.) Urb. Potential* X X ― X ― [60] 29 Melilotus indicus (L.) All. X X X ― ― X ― [66,67] 30 Melilotus officinalis (L.) Pall. X ― X X ― ― [7,68] 31 Melilotus siculus (Turra) B.D.Jacks. X X ― ― X ― [7,53,69] 32 Pongamia pinnata (L.) Pierre X X X X ― X ― [70] 33 Pycnospora lutescens (Poir) Schindl. Potential* ― ― ― ― ― ― 34 Senna bicapsularis (L.) Roxb. X X X X ― ― ― ― Data to be published 35 Sesbania cannabina (Retz.) Pers. X X X X ― ― X ― [14,71,72] 36 Sesbania rostrata Bremek. & Oberm X ― X ― X ― [7] 37 Sesbania sesban (L.) Merr. X ― X ― ― ― [7,25] 38 Stylosanthes guianensis (Aubl.) Sw. X X X ― ― ― [73,74] 39 Swainsona formosa X X X ― ― ― [75] 40 Tephrosia purpurea (Linn.) Pers. Syn. Pl. Potential* X ― ― ― ― https://apps.worldagroforestry.org/treedb/AFTPDFS/Tephrosia_purpurea.PDF 41 Teramnus labialis (Linn.f.) Spreng. Potential* ― ― ― ― ― ― 42 Trifolium fragiferum L. X ― X ― X ― [67] 43 Uraria lagopodiodies (Linn.) Desv. ex DC. Potential* ― ― ― ― ― ― 44 Vigna marina (Burm.) Merr. X X X X X ― ― ― [13,76] Potential*: some germplasms collected from coastal areas with potential salinity tolerance and kept in the seed bank of Prataculturae Research Centre, Tropical Crops Genetic Resources Institute, Chinese Academy of Tropical Agricultural Sciences (CATAS). X means stress tolerant; XX means high stress tolerant. One important additional factor to consider is that some saline soils are marshes, which means that forage legumes integrated in such areas will also need to show waterlogging tolerance. Similar to what was observed for salinity tolerance, many reports have been published to explore legume waterlogging tolerance and legume-rhizobia symbiotic models for waterlogging tolerance[48−52], legume species Canavalia ensifomis, Centrosema pubescens, Indigofera hirsute, Indigofera litoralis, Macroptilium atropurpureum, Melilotus indicus, Melilotus siculus, Pongamia pinnata, Sesbania cannabina, Sesbania rostrata and Trifolium fragiferum may have the combined tolerance of waterlogging and salinity based on the summarized information (Table 2), previous reports also indicated that Melilotus siculus is a very promising fodder for saline and waterlogged soils[53,54]. Researchers should pay more attention to legumes combining the traits of salinity and waterlogging tolerance and their field testing.
Some of the listed legume species are tolerant to other abiotic stresses including drought tolerance, acidity tolerance, shade tolerance and frost tolerance (Table 2), drought tolerance is also a very import characteristic compared to salinity and waterlogging.
There is high productivity diversity among the 44 legume species, eight species Alysicarpus vaginalis (4−6 t DW/ha/y), Cajanus cajan (3.175−6.515 t DW/ha/y), Calopogonium mucunoides (4−5 t DW/ha/y), Dendrolobium triangulare (4−5 t DW/ha/y), Indigofera hirsuta (3−6 t DW/ha/y), Macroptilium atropurpureum (2.4−5.5 t DW/ha/y), Stylosanthes guianensis (4−5 t DW/ha/y) and Trifolium fragiferum (4.31−5.19 t DW/ha/y) had relative high productivity compared to other species. The nutritional value of these legumes also varies among species, for instance, the maximum crude protein content of Sesbania sesban, Trifolium fragiferum, Macroptilium atropurpureum and Teramnus labialis are more than 20%. Further evaluation, including other nutrition values, chemical composition and feeding experiments should be conducted to test acceptability to animals and performance of livestock.
Few studies exist regarding the nitrogen fixation efficiency of salinity tolerant legumes even though it is an important indicator for their utilization. Among the 44 species, only 11 species have been reported to have nitrogen fixation efficiency, Sesbania cannabina (949−1,040.25 kg N/ha/y), Arachis pintoi (300 kg/ha/y), Centrosema pubescens (120−270 kg N/ha/y) and Tephrosia purpurea (202.23 kg N/ha/y) had higher nitrogen fixation efficiency[77−79] (
www.tropseeds.com/arachis-pintoi/ ). We also found very few studies on salinity tolerant rhizobia associated with the listed legumes, most reported rhizobia could tolerate to a salinity concentration higher than 300 mM NaCl (Table 3).Table 3. Productivity, nutrition value, BNF and salinity tolerance rhizobia of current tolerant forage legume species.
No. Scientific name Productivity Leaf crude protein Nitrogen fixation efficiency Salt tolerant rhizobia Reference 1 Abrus precatorius L. ― 16.28% ― ― [80] 2 Acacia dealbata Link 1.2−4.0 t DW/ha/y ― 40 kg N/ha/y ― [81, 82] 3 Acacia nilotica (L.) Willd. ex Delile 3.22 t DW/ha/y 13.92% ― ― [83−85] 4 Alysicarpus vaginalis (L.) DC. 4–6 t DW/ha/y 17.91% ― ― https://apps.lucidcentral.org/tropical_forages ,https://extension.msstate.edu/content/alyceclover-alysicarpus-vaginalis 5 Arachis pintoi Krapov. & W.C.Greg. 3−4 t DW t/ha/y 17%-20% 300 kg/ha/y ― www.tropseeds.com/arachis-pintoi/ 6 Cajanus cajan (L.) Millsp. 3.175−6.515 t DM/ha/y ― ― ― [86] 7 Calopogonium mucunoides Desv. 4−5 t DW/ha/y low ― ― [57,87] 8 Canavalia ensifomis (L.) DC ― ― ― ― ― 9 Canavalia rosea (Sw.) DC. 1.0−4.6 t DW/ha/y 15%−20% ― Some strains tolerant to 500−600 mM NaCl [88,89] 10 Cassia pumila Lam. ― ― ― ― ― 11 Centrosema pubescens Benth. 1.6−2.8 t DW /ha/y 21.36%−23.34% 120−270 kg N/ha/y ― [78] 12 Clitoria ternatea Linn. Sp. Pl. 1.2−3.6 t DW /ha/y 14%−20% ― Jd19 Rhizobium strain [90,91] 13 Crotalaria albida Heyne ex Roth. ― ― ― ― ― 14 Crotalaria bractaeata Roxb. ex DC. ― ― ― ― ― 15 Crotalaria ferruginea (Grah.) Benth. ― ― ― ― ― 16 Crotalaria retusa Linn. ― 14.6%−18.0% ― ― [63] 17 Dendrolobium triangulare(Retz.) Schindl. 4−5 t DW/ha/y Average 13.43% ― ― [92] 18 Desmanthus virgatus (L.) Willd. 3.98 t DW/ha/y Average 15.20% ― ― [93] 19 Erythrina corallodendron L. ― ― ― ― ― 20 Galactia elliptifoliola Merr. 2.0−3.5 t DW/ha/y ― ― ― [94] 21 Indigofera chuniana Metc. 2−4 t DW/ha/y ― ― ― Date to be published 22 Indigofera enneaphylla Linn. Mant. 3−4 t DW/ha/y 10.7% ― ― [95] 23 Indigofera galegoides DC. ― Average 3.26% for total N ― ― [96] 24 Indigofera hirsuta Linn. 3−6 t DW/ha/y 1.7−1.9% for total N ― ― [96] 25 Indigofera litoralis Chun & T.C.Chen ― ― ― ― ― 26 Indigofera suffruticosa Mill. 2−4 t DW/ha/y Average 3.77 for total N ― ― [96] 27 Leucaena leucocephala (Lam.) de Wit. 3.4 t DW/ha/y 22.8%−25.9% 76 kg N/ha/y ― [97, 98] 28 Macroptilium atropurpureum (L.) Urb. 2.4–5.5 t DW/ha/y 13.73%−28.2% 62−178 kg N/ha/y ― [99−101] 29 Melilotus indicus (L.) All. 3.8 t DW/ha/y ― ― Some strain tolerant to 6% NaCl [66, 102] 30 Melilotus officinalis (L.) Pall. ― 14.5%−19.4% Add 80–130 pounds/acre of nitrogen to soil ― [103, 104] www.sciencedirect.com/topics/agricultural-and-biological-sciences/melilotus-officinalis 31 Melilotus siculus (Turra) B.D.Jacks. ― ― ― ― ― 32 Pongamia pinnata (L.) Pierre ― ― ― ― ― 33 Pycnospora lutescens (Poir) Schindl. ― ― ― ― ― 34 Senna bicapsularis (L.) Roxb. ― ― ― ― ― 35 Sesbania cannabina (Retz.) Pers. 2.46−3.55 t DW/ha/y ― 949−1,040.25 kg N/ha/y Some strains tolerant to 5.0% (w/v) NaCl [77, 105, 106] www.healthbenefitstimes.com/sesbania 36 Sesbania rostrata Bremek. & Oberm 1.06−2.19 t DW/ha/y Average 19.9% 90−219 kg N/ha/y ― [77,107] 37 Sesbania sesban (L.) Merr. 2.39−2.59 kg DW/ha/y 20%−25% 42.6−109.5 kg N/ha/y ― [108,109] 38 Stylosanthes guianensis (Aubl.) Sw. 4−5 t DW /ha/y 14%−20% 96−122 kg N/ha/y Bradyrhizobium strain RJS9-2 tolerant to 350 mM NaCl [46,110] 39 Swainsona formosa ― ― ― ― ― 40 Tephrosia purpurea (Linn.) Pers. Syn. Pl. ― 16.27% 202.23 kg N/ha/y Some strains tolerant to 2.5%−3% of NaCl [79,111] 41 Teramnus labialis (Linn.f.) Spreng. 3−4 t DM/ha/y 22.86% ― ― [112,113] 42 Trifolium fragiferum L. 4.31−5.19 t DM/ha/y 14.9%−25.7% ― ― [114,115] 43 Uraria lagopodiodies (Linn.) Desv.ex DC. ― ― ― ― ― 44 Vigna marina (Burm.) Merr. 2.0−4.0 t DW/ha/y 13%−20% ― Some strains tolerant to 600 mM NaCl [13,116] From the 44 summarized legumes, 95.5% have potential to be utilized as forage, Abrus precatorius and Crotalaria retusa contain toxic substances such as monocrotaline which is not safe to use as forage. Eighty four point one percent have potential as green manure, the species Abrus precatorius, Acacia dealbata, Acacia nilotica, Leucaena leucocephala and Pongamia pinnata are too big to be green manure. Eleven point four percent of the species may be developed for food, 59.1% as medicine and 40.9% for ornamental purposes (Table 4).
Table 4. Utilization of current salinity tolerant forage legume species.
No. Scientific name Forage Green manure Food Medicine Ornamental Reference 1 Abrus precatorius L. ― ― ― X X [118] 2 Acacia dealbata Link X ― ― X X [119] 3 Acacia nilotica (L.) Willd. ex Delile X ― ― X ― [84] 4 Alysicarpus vaginalis (L.) DC. X X ― X ― https://apps.lucidcentral.org/tropical_forages 5 Arachis pintoi Krapov. & W.C.Greg. X X ― ― X https://apps.lucidcentral.org/tropical_forages ,www.tropseeds.com/ 6 Cajanus cajan (L.) Millsp. X X X X ― [120] 7 Calopogonium mucunoides Desv. X X ― ― ― [121] 8 Canavalia ensifomis (L.) DC X X X X ― [122] 9 Canavalia rosea (Sw.) DC. X X X X X [89] 10 Cassia pumila Lam. X X ― X ― [59] 11 Centrosema pubescens Benth. X X ― ― ― [78] 12 Clitoria ternatea Linn. Sp. Pl. X X ― X X [90,123] 13 Crotalaria albida Heyne ex Roth. ― X ― X ― [124] 14 Crotalaria bractaeata Roxb. ex DC. X X ― X ― https://plants.ces.ncsu.edu/plants/crotalaria-spectabilis/ 15 Crotalaria ferruginea (Grah.) Benth. X X ― X ― [125] 16 Crotalaria retusa Linn. ― X X X ― https://baike.baidu.com/item/%E5%90%8A%E8%A3%99%E8%8D%89?fromModule=lemma_search-box#reference-[1]-3529418-wrap 17 Dendrolobium triangulare(Retz.) Schindl. X X ― X X https://baike.baidu.com/item/%E5%81%87%E6%9C%A8%E8%B1%86/23437879?fromModule=lemma-qiyi_sense-lemma 18 Desmanthus virgatus (L.) Willd. X X ― ― ― [93] 19 Erythrina corallodendron L. X ― ― X X https://baike.baidu.com/item/%E9%BE%99%E7%89%99%E8%8A%B1/3567229?fr=kg_general 20 Galactia elliptifoliola Merr. X X ― ― ― [94] 21 Indigofera chuniana Metc. X X ― ― X https://baike.baidu.com/item/%E7%96%8F%E8%8A%B1%E6%9C%A8%E8%93%9D/1483435?fr=kg_general 22 Indigofera enneaphylla Linn. Mant. X X ― X X [126] 23 Indigofera galegoides DC. X X ― X X [96] 24 Indigofera hirsuta Linn. X X ― ― X [96] 25 Indigofera litoralis Chun & T.C.Chen X X ― X X https://baike.baidu.com/item/%E6%BB%A8%E6%B5%B7%E6%9C%A8%E8%93%9D/7181556?fr=aladdin 26 Indigofera suffruticosa Mill. X X ― X ― [96] 27 Leucaena leucocephala (Lam.) de Wit. X ― X ― X [97] 28 Macroptilium atropurpureum (L.) Urb. X X ― ― ― [99] 29 Melilotus indicus (L.) All. X X ― X ― [66] 30 Melilotus officinalis (L.) Pall. X X ― X X [103] 31 Melilotus siculus (Turra) B.D.Jacks. X X ― ― X [10] 32 Pongamia pinnata (L.) Pierre X ― ― X ― [70] 33 Pycnospora lutescens (Poir) Schindl. X X ― ― ― [127] 34 Senna bicapsularis (L.) Roxb. X X ― X ― [128] 35 Sesbania cannabina (Retz.) Pers. X X ― ― X [14] 36 Sesbania rostrata Bremek. & Oberm X X ― ― ― [107] 37 Sesbania sesban (L.) Merr. X X ― ― ― [109] 38 Stylosanthes guianensis (Aubl.) Sw. [110] 39 Swainsona formosa X X ― ― X [75] 40 Tephrosia purpurea (Linn.) Pers. Syn. Pl. X X ― X ― https://apps.worldagroforestry.org/treedb/AFTPDFS/Tephrosia_purpurea.PDF 41 Teramnus labialis (Linn.f.) Spreng. X X ― X ― [112] 42 Trifolium fragiferum L. X X ― ― ― [115] 43 Uraria lagopodiodies (Linn.) Desv.ex DC. X X ― ― https://powo.science.kew.org/taxon/524403-1 44 Vigna marina (Burm.) Merr. X X X X X [129] X means can be used. When served as forage, these legumes can either be used in a cut and carry system or grazing, depending on the size of the plots, other crops grown in the same area or farmers' preference. However, in the case of forage, the nutrients contained in the plants do not benefit the soil: there is a trade-off between soil rehabilitation and livelihood benefits[117]. A compromise is to alternate use for forage or green manure at different times if space allows. A good strategy would be to return animal manure to the soil, eventually following a composting phase. When used as green manure, these legumes can be intercropped with fruit trees or grown in rotation with salinity tolerant food crops (cereals or tubers). They can then protect the soil (moisture conservation, reduce erosion etc.), decompose and increase soil organic matter and nitrogen content.
Adoption of salinity tolerant legumes in coastal areas is limited. Very few studies have reported the adoption of these crops. In Western Australia, salinity tolerant pasture legumes and grasses have been promoted to improve the productivity and profitability of saline land and salt bush-based pastures (
www.agric.wa.gov.au/soil-salinity/pasture-legumes-and-grasses-saline-land-western-australia ). In south-eastern Tunisia, a study on farmers' willingness to adopt salinity-tolerant forage crops showed that off-farm income availability and flock size significantly affected farmers' willingness to adopt salinity-tolerant forages[130]. Testing of halophytes and salinity-tolerant plants as potential forage for ruminants was carried in the Near East region, Egypt, but few of them are legumes[131]. However, adoption barriers typical for cover crops can be expected such as measures related to soil management, impact on yields and income are not immediate, making it difficult for farmers to invest in the technology[132]. When used as forage, the likelihood of adoption is higher as increases in milk production and weight gain can be quickly observed. One main barrier is the availability of planting material, it's a challenge for the farmers to find planting legumes with high salinity tolerant, high biomass and high nutritive value. -
Salinity tolerant legumes have a great potential to reclaim sea-level-rise affected tropical coastal agroecosystems, support bridging the protein gap in an environmentally-friendly manner and increase smallholder farmers' income (Fig. 1). Although usages and some benefits have been documented, detailed information is lacking for many of them. Tolerance combinations for locations with multiple abiotic stresses like marshes also need to be further explored. Among the species reviewed, Sesbania cannabina (Retz.) Pers., Melilotus indicus (L.) All. and Vigna marina (Burm.) Merr. are good candidates but there are still gaps in the research before it can be promoted at scale. Gaps include the nitrogen fixation efficiency and soil reclamation potential, as well as the impact on farming systems and livelihoods in a holistic way. The selection of salinity-tolerant rhizobia symbiosis, which are more effective than when both legumes and rhizobia are selected separately, is particularly crucial.
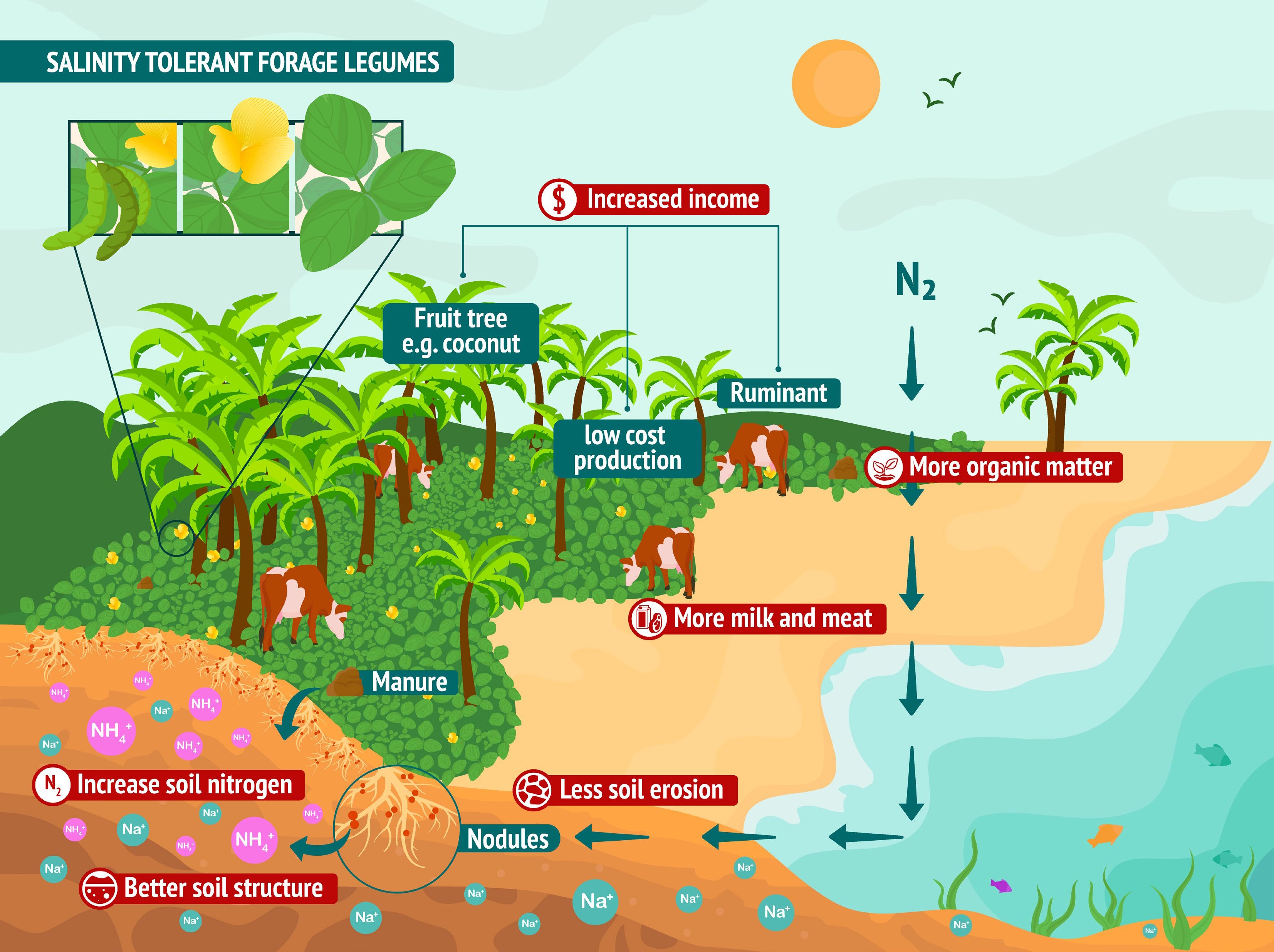
Figure 1.
Inter-cropping system with salinity tolerant legumes on coastal ecosystems.
The integration of salinity tolerant legumes into coastal farming systems will be subject to country-specific adoption barriers and system requirements. To ensure livelihood benefits for millions of smallholder farmers in tropical coastal agroecosystems, commitment and support from government and public services are necessary to address both seed system and extension needs, through the provision of adequate incentives, policies and development efforts.
This work was funded by the National Science and Technology Basic Resources Investigation Project (2017FY100600), Feeds and Forages flagship of the CGIAR Research Program on Livestock, the Key Research and Development Program of Hainan (321RC646), China Agriculture Research System of MOF and MARA (CARS-22). We warmly thank Ms. Andrea Ramírez and Mr. José Luis Urrea Benitez for the design of Fig. 1.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Yiming Liu, Mary Atieno
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liu Y, Atieno M, Cardoso JA, Yang H, Xu Bin, et al. 2022. Mining and utilization of salinity tolerant legumes in tropical coastal agroecosystems: An overview. Grass Research 2:10 doi: 10.48130/GR-2022-0010 

Mining and utilization of salinity tolerant legumes in tropical coastal agroecosystems: An overview
- Received: 18 September 2022
- Accepted: 16 November 2022
- Published online: 21 December 2022
Abstract: Coastal saline soils are increasing year by year caused by climate change and human activities. Most of the coastal saline soils are idle due to their high salinity level and few crops can grow normally. Salinity tolerant legumes are naturally tolerant to salt, which can ecologically cover the coastal saline soil, enhance soil fertility by symbiotic nitrogen fixation and increase the smallholder farmers’ benefits in terms of forage, green manure, food or medicine. However, few reports are available for the systematic evaluation of salinity tolerant legumes. This review summarizes and evaluates currently available salinity tolerant legume species that could potentially be used and discusses their potential for integration into smallholder mixed coastal systems of the Asia-Pacific region. Fourty four salinity tolerant legumes were summarized, six of them showed a high level of salinity tolerance, 17 of them showed a moderate level of salinity tolerance and 21 of them showed potential salinity tolerance but need to be further studied. Many gaps such as combined tolerance evaluation, nitrogen fixation efficiency, animal feeding experiments and salinity tolerant rhizobia screening/inoculants exist. Case studies demonstrate legumes could be used to reclaim coastal saline soils, but commitment and support from government and public services are necessary to address both seed system and extension needs, through the provision of adequate incentives, policies and development efforts.
-
Key words:
- Forage legume /
- Rhizobia /
- Salinity tolerance /
- Intercropping system /
- Coastal smallholder farmers.