









-
Creeping bentgrass (Agrostis stolonifera L.), one of the major cool-season turfgrass species, is widely planted in temperate and subtropical regions of China[1,2]. It exhibits fine leaf textures, high shoot densities, short internodes, prostrate growing habits and tolerance to extremely close mowing[3]. However, creeping bentgrass is not well adapted to the persistent hot and humid conditions during summer months, which often results in reduced plant vigor and increased susceptibility to diseases[4]. The root disease, summer patch, caused by Magnaporthiopsis poae, was originally isolated from symptomatic roots of Kentucky bluegrass in the USA[5]; it has been documented in creeping bentgrass in China, recently[2,6]. Take-all patch, caused by Gaeumannomyces graminis var. avenae, pythium root rot or dysfunction (Pythium spp.) were common root diseases of creeping bentgrass in the USA[7,8], but have not been documented in China. Among the root associated pathogenic fungi, there are a few fungi that colonize plant roots via darkly pigmented, ectotrophic, runner hyphae, which were named ectotrophic root-infecting (ERI) fungi[9]. Members of the ERI fungi were soil-borne, and some of them were the causal agents of destructive turfgrass diseases[10]. Several well-known genera of ERI fungi have been described on turfgrass, including Gaeumannomyces, Ophiosphaerella, and Magnaporthiopsis[11−13]. Recently, a new genus of ERI fungi, Candidacolonium, was described by Vines[14], and was associated with the decline of ultradwarf bermudagrass (Cynodon dactylon × C. transvaalensis) in late summer and early fall in the Deep South of the USA[14,15]. Virulence tests revealed that C. cynodontis was one of the most destructive and aggressive ERI fungi and the main cause of summer decline observed on ultradwarf bermudagrass[14]. C. cynodontis readily produced round to mitten-shaped hyphopodia, which were frequently observed on roots as well as stolons[14].
In autumn 2020 and summer 2021, a summer patch-like disease emerged on creeping bentgrass greens at two golf courses in East China. Initially, the symptom of the disease showed multiple irregular yellowing spots (3−10 cm in diameter). As the disease progressed, the spots coalesced into larger irregular patches. Roots of the creeping bentgrass were sparse, shortened and dark colored. Dark ectotrophic runner hyphae and slightly lobed or mitten-shaped hyphopodia were observed inside and on the surface of brown or black roots under the microscope. The symptomatology, time of occurrence, as well as presence of slightly lobed or mitten-shaped hyphopodia, suggested that the disease on creeping bentgrass may be caused by an ERI fungal species[9].
Accurate identification of ERI fungi is generally difficult and time consuming by traditional morphological methods[10]. With the exception of the different shapes of hyphopodia and the occasional production of ascocarps and ascospores, the ERI fungi do not produce distinguishable features that allow for precise and timely identification[14,16,17]. In addition, production of ascocarps was often difficult due to attenuated biotypes or incompatible mating types[10]. According to only morphological characters, accurate identification of ERI fungi was often hampered by highly similar structures, especially conidia, among close related species[18,19]. Multi-locus molecular phylogenetic analysis had been successfully used for the identification of ERI fungi, especially for new species[18, 20].
The objective of this study was to employ molecular as well as morphological methods to identify the fungal isolates recovered from diseased creeping bentgrass roots, and determine the main fungal species that cause the summer patch-like disease on creeping bentgrass by pathogenicity tests. The work will provide important references for the management of creeping bentgrass root diseases at golf courses in East China.
-
Samples of symptomatic creeping bentgrass were collected by a soil corer (5 cm in diameter and 15 cm in depth) from the putting greens at Taihu golf course in Wuxi city, Jiangsu province and Jianhu golf course in Shaoxing city, Zhejiang province (China). Root materials were rinsed with distilled water to remove organic materials and soil particles, and microscopically checked for the presence of dark and runner hyphae, a characteristic sign of ERI fungi. Dark colored roots were cut into sections with 2−3 cm in length, surface disinfested with a 0.6% sodium hypochlorite solution for 5 min, rinsed two consecutive times with sterile-distilled water, placed on sterile filter paper, and dried in a laminar flow hood. The dried root sections were transferred onto potato dextrose agar (PDA) medium amended with 50 mg·L−1 each of streptomycin sulfate, ampicillin and tetracycline. Plates (9 cm in diameter) were incubated at 25 °C until hyphal growth was observed from the discoloring root tissues. Two to three consecutive hyphal tip transfers were conducted to obtain pure cultures. Fungal cultures were stored for the long term as described by Hu et al.[21].
PCR amplification and sanger sequencing
-
Fungal cultures were grown on PDA at 25 °C for 7−10 d to produce actively growing colonies. Mycelia of each strain were scraped without agar medium using a sterilized scalpel and put into a 2-mL micro-centrifuge tube. The genomic DNA (gDNA) was extracted with the previously described method[22]. Concentration and purity of the extracted gDNA was tested by a NanoDrop 2000/2000c device (Thermo Fisher Scientific, Inc., Waltham, MA, USA). DNA samples of the fungal isolates were stored at −20 °C for further use.
PCR amplifications were conducted on a thermal cycler (Applied Biosystems, USA) in a 30-μL reaction volume. The PCR reaction system and cycling condition were the same as described in a previous study[14]. Six genes were used for multi-locus phylogenetic analysis, including the internal transcribed spacer (ITS), 28S large subunit (LSU) rRNA gene and 18S small subunit (SSU) rRNA gene regions of rRNA gene, the largest subunit of RNA polymerase II gene (RPB1), a DNA replication licensing factor gene for mini-chromosome maintenance complex component 7 (MCM7) and the translation elongation factor 1-alpha gene (TEF1). Partial sequences of ITS were used for preliminary identification through BLAST analysis (
https://blast.ncbi.nlm.nih.gov/Blast.cgi ). Isolates were considered as new fungal species and were further used for multi-locus phylogenetic, morphological and biological analyses, if the percent identity of their ITS sequences was less than 97% as compared with the submitted ITS sequences in GenBank database through BLAST analysis. Primers used for PCR amplifications of the six genes were the same as in Vines' research[14]. PCR products were examined electrophoretically in 1% agarose gels. The PCR fragment was purified and sent to Tsingke Biotechnology Co., Ltd. for sequencing. The obtained sequences of the potential new species were deposited in GenBank, and the specific information of the strains representing different species used for constructing the phylogenetic tree is provided in Table 1.Table 1. Species name, isolate IDs, and GenBank accession numbers for fungal isolates used for phylogenetic analyses in this study.
Species name Isolate ID ITS LSU SSU MCM7 RPB1 TEF1 Buergenerula spartinae ATCC 22848a JX134666 DQ341492 DQ341471 JX134706 JX134720 JX134692 Candidacolonium agrostisb 13g11 OM910761 OM910782 OM900029 OM938227 OM938229 OM938231 Candidacolonium agrostis 13g12 OM910762 OM910783 OM900030 OM938228 OM938230 OM938232 Candidacolonium agrostis JH1 OP811262 OP811312 OP811273 OP957424 OP957426 OP957428 Candidacolonium agrostis JH2 OP811263 OP811313 OP811274 OP957425 OP957427 OP957429 Candidacolonium cynodontis HP24-3 KJ855497 KM401637 KP129316 KP007344 KP268919 KP282703 Candidacolonium cynodontis HP38-4 KJ855498 KM401638 KP129317 KP007345 KP268920 KP282704 Gaeumannomyces graminis var. graminis GSGC15-4 KJ855495 KM401635 KP129314 KP007342 KP268917 KP282701 Gaeumannomyces graminis var. avenae CBS 187.65 JX134668 JX134680 JX134655 JX134708 JX134722 JX134694 Gaeumannomyces graminis var. tritici M55 JF414850 JF414900 JF414875 JF710395 JF710445 JF710420 Gaeumannomyces paulograminis RS7-1 KJ855507 KM401647 KP129326 KP007350 KP268929 KP282713 Magnaporthiopsis cynodontis RS7-2 KJ855508 KM401648 KP129327 KP007351 KP268930 KP282714 Magnaporthiopsis cynodontis KR10-6 KJ855499 KM401639 KP129318 KP007357 KP268921 KP282705 Magnaporthiopsis cynodontis TPC5-3 KJ855514 KM401654 KP129333 KP007356 KP268936 KP282720 Magnaporthiopsis incrustans WW3-5 KJ855515 KM401655 KP129334 KP007332 KP268937 KP282721 Magnaporthiopsis panicorum CM10S2 KF689644 KF689634 KF689594 KF689604 KF689614 KF689624 Magnaporthiopsis poae TAP41 KJ855512 KM401652 KP129331 KP007354 KP268934 KP282718 Magnaporthiopsis rhizophila M23 JF414834 JF414883 JF414858 JF710384 JF710432 JF710408 Nakataea oryzae M21 JF414838 JF414887 JF414862 JF710382 JF710441 JF710406 Nakataea oryzae M69 JX134672 JX134684 JX134658 JX134712 JX134726 JX134698 Nakataea oryzae M71 JX134673 JX134685 JX134659 JX134713 JX134727 JX134699 Ommidemptus affinis ATCC 200212 JX134674 JX134686 JX134660 JX134714 JX134728 JX134700 Pseudophialophora cynodontis RW3-4 KJ855509 KM401649 KP129328 KP007352 KP268931 KP282715 Pseudophialophora eragrostis CM12M9 KF689648 KF689638 KF689598 KF689608 KF689618 KF689628 Pseudophialophora eragrostis CM20M5-2 KF689647 KF689637 KF689597 KF689607 KF689617 KF689627 Pseudophialophora panicorum CM9S6 KF689651 KF689641 KF689601 KF689611 KF689621 KF689631 Pseudophialophora schizachyii AL2M1 KF689649 KF689639 KF689599 KF689609 KF689619 KF689629 Pyricularia grisea M83 JX134671 JX134683 JX134657 JX134711 JX134725 JX134697 Slopeiomyces cylindrosporus CBS 610.75 JX134667 DQ341494 DQ341473 JX134707 JX134721 JX134693 a ATCC = American Type Culture Collection, Manassas, Virginia; CBS = Centraalbureau voor Schimmelcultures, Utrecht, the Netherlands. b Species names in boldface indicate newly submitted sequences from this study. Sequence data and phylogenetic analyses
-
Phylogenetic analyses were implemented in PhyloSuite software version 1.2.1[23]. The amplified sequences of the six individual genes were aligned respectively with the MAFFT program by the algorithm G-INS-i[24], alignments were checked with characteristics weighted equally. The incongruence length difference test[25] was implemented to determine if data for the six genes could be combined with the software PAUP 4.0a169[26]. A multi-locus dataset was produced by concatenating the aligned sequences of individual genes with the Concatenate Sequence program[23]. The best nucleotide substitution models for individual genes were selected using the Akaike Information Criterion (AIC) with the ModelFinder program[27]. Individual gene trees were generated for each gene with the above predicted model parameters. Phylogenetic analysis was performed with the multi-locus dataset using maximum likelihood (ML) and Bayesian inference (BI) approaches. BI phylogenies were implemented using MrBayes 3.2.6[28] with partition model (2 parallel runs and 10 million generations), in which 25% of the initially sampled data were discarded as burn-in, the remaining trees were applied to calculate posterior probabilities (PP). ML phylogenies were conducted using IQ-TREE[29] with Edge-linked partition model (5000 ultrafast bootstraps)[30] and Shimodaira-Hasegawa-like approximate likelihood-ratio test[31]. Clades with PP ≥ 0.95 and bootstrap values (BS) ≥ 70% were considered as well supported. Phylograms were annotated and viewed with Interactive Tree Of Life (iTOL) version 6[32].
Cultural characteristics
-
Cultural characteristics were recorded from PDA. Mycelial plugs (diameter at 5 mm) were taken from the margins of one-week growing colonies, transferred to new PDA plates (diameter at 9 cm), and incubated in the dark at 25 °C. The diameter of the colony for each culture was measured daily for five days. The color of fungal colony was noted according to Rayner[33]. Microscopic measurements and photographs were conducted from slides of fungi mounted in sterile-distilled water with a stereo-microscope (OLYMPUS SZX16), and with a digital microscope (OLYMPUS DP80). The width of the hyphae and the size of the hyphopodia were measured from 20−30 structures depending on availability. The specimens and ex-type culture were deposited and preserved in Fungarium of Guangdong Institute of Microbiology (GDGM) and Guangdong Microbial Culture Collection Center (GDMCC), respectively.
Optimum temperature for in vitro growth
-
Two isolates (13g11 and JH1) of the potential new species were used to estimate the optimal temperature for their growth. The isolates were grown on PDA at 25 °C for 5 d. Mycelial plugs with 5-mm in diameter from the margin of the colony were obtained and transferred onto the center of new PDA plates. The plates were sealed and incubated in the dark at 4, 20, 25, 29, and 37 °C for 5 d. Each isolate had three replications at each temperature, and mycelial growth was measured at the end of the incubation. The effects of temperature treatment were identified by analysis of variance using the general linear model procedure implemented in SAS 9.4 (SAS Institute, Cary, North Carolina, USA). The mean differences were compared with Fisher's protected least significance difference (LSD) test at α = 0.05.
Pathogenicity tests
-
Pathogenicity experiments were conducted by artificial inoculations with the type strain 13g11. Inoculum was prepared through sterilizing oat (Avena sativa) seeds in a 50 mL flask. Ten actively growing mycelial plugs with 5-mm in diameter of the strain were added to the sterilized oat substrate, which was shaken and incubated at 28 °C in the dark for one month. Inoculations were conducted in plastic pots (15 cm top diameter × 10 cm bottom diameter × 15 cm height) by placing eight inoculated oat seeds over 100 g of sterilized Pindstrup substrate (Pindstrup Mosebrug, Pindstrup, Denmark) and sand mixture (1:1, v/v), and healthy creeping bentgrass were transplanted to the pots from the Baima research station of Nanjing Agricultural University. The plastic pots inoculated with eight noninfected and sterilized oat seeds served as the control. The inoculated pots were placed in a growth chamber with 95% relative humidity and a 12-h day/night cycle at 26/22 °C. All pots were covered with plastic bags for the first 5 d and watered once daily to favor disease development. The inoculated plants were monitored for development of symptoms weekly, and a final assessment was conducted one month after the inoculations. Re-isolations and identifications were performed as described above to fulfill Koch's postulates.
Genealogical Concordance Phylogenetic Species Recognition analysis
-
Recombination event between the potential new species and its phylogenetically related species was characterized with the Genealogical Concordance Phylogenetic Species Recognition (GCPSR) model by performing a pairwise homoplasy index (Φw) (PHI) test[34]. The test was performed in SplitsTree4 v.4.16.1. (
http://uni-tuebingen.de ),[35] and the recombination level within phylogenetically related species was determined. Recombination was considered when the PHI value was below 0.05 (Φw < 0.05). The relationships between closely related species could be visualized by constructing a phylogenetic network with the LogDet transformation and the NeighborNet method, and the network was displayed with the EqualAngle algorithm and bootstrap analysis (1,000 replicates)[36]. -
Partial sequences of ITS, LSU, SSU, MCM7, RPBI and TEF1 genes were sequenced in four isolates (13g11, 13g12, JH1 and JH2). The incongruence length difference test generated a p value of 0.01 of the concatenated dataset, which indicated that some incongruence was shown in the datasets for the six regions. These regions were still combined, because studies revealed that the threshold of significance at 0.05 might be too conservative, and it had been shown that concatenating incongruent datasets improved the accuracy of phylogeny[37]. The single-locus BI phylogenetic trees for ITS, rDNA (ITS + LSU + SSU), MCM7, RPB1, and TEF1 were shown in Supplemental Figs S1−S5. The multilocus dataset comprised 3,778 characters, there were 504 nucleotide characters (including gaps) in the SSU alignment, 510 in ITS, 819 in LSU, 500 in MCM7, 612 in RPB1, and 833 in TEF1. A total of 2,833 characters were conserved, and 689 were parsimony informative.
GTR+F+I+G4 was estimated as the best nucleotide substitution model for ITS, rDNA, RPB1, MCM7 and TEF1. The multi-locus phylogenetic tree constructed was similar to those constructed from the datasets of individual markers. The BI and ML phylogenetic trees were constructed similarly in topology (Fig. 1 and Supplemental Fig. S6), and only the BI tree is shown in Fig. 1.
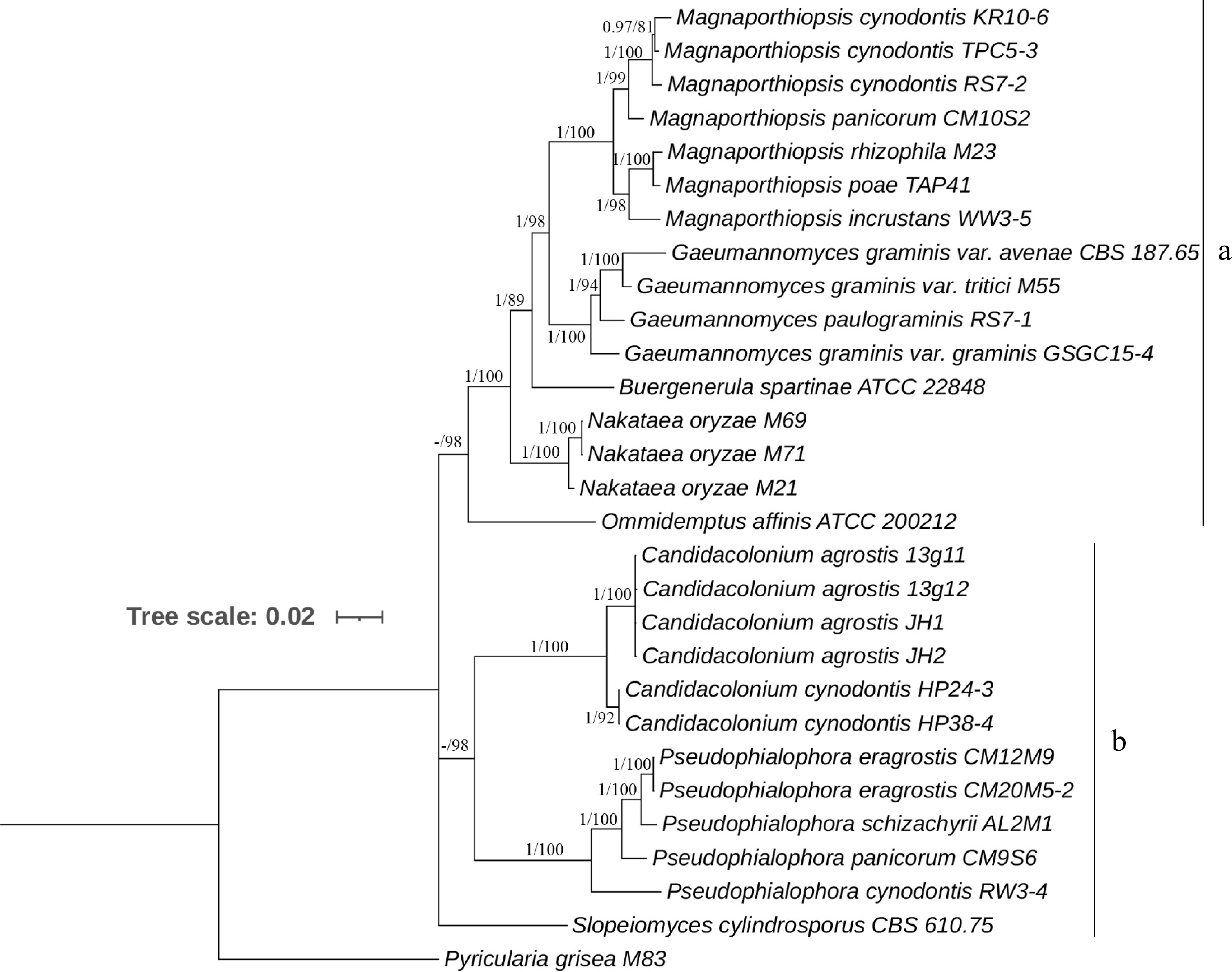
Figure 1.
Majority rule Bayesian phylogenetic tree derived from the combined ITS, LSU, SSU, MCM7, RPB1, and TEF1 DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥ 0.95 and maximum likelihood bootstrap values ≥ 75% are shown. The tree is rooted with Pyricularia grisea M83.
Our four fungal isolates were placed with confidence into the subclade of Candidacolonium (PP = 1, BS = 100%), from which a new species in Candidacolonium was identified, and found to be distinct from C. cynodontis in this genus. A new species of Candidacolonium is thereby proposed based on the phylogeny analysis and morphological characteristics (see Taxonomy; Fig. 2).

Figure 2.
Disease symptoms, morphological characters and pathogenicity of Candidacolonium agrostis. (a) Summer patch-like disease on creeping bentgrass putting green. (b) Front and back of colony on PDA plate after 14-d incubation. (c) Hyphopodia. (d), (e) Creeping bentgrass top growth one month after inoculation with sterile and 13g11 inoculum, respectively. (f), (g) Creeping bentgrass roots one month after inoculation with sterile and 13g11 inoculum, respectively.
Growth rate
-
The two isolates of the potential new species, 13g11 and JH1, exhibited similar growth rates. They all grew well at 20−29 °C, but did not show growth at 4 °C, and reached maximal growth at 26−29 °C (Fig. 3). The two isolates 13g11 and JH1 grew significantly slower (p < 0.05) than the two M. poae isolates Y15 and S24 at 20, 26 and 29 °C after 5 d of inoculation.

Figure 3.
Total mycelial growth (colony diameter, mm) of Candidacolonium agrostis (13g11 and JH1) and Magnaporthiopsis poae (Y15 and S24) isolates on PDA after 5 d of incubation in the dark at temperatures of 4, 20, 26, 29 and 37 °C. Error bars represent the mean ± standard deviation.
Pathogenicity tests in a growth chamber
-
The inoculation experiment in the growth chamber led to disease symptoms on creeping bentgrass and fulfilled the Koch's postulates. The roots turned dark-colored and rotted one month after inoculation. Runner hyphae with brown color were apparently observed from roots of all grasses with fungal inoculation (Fig. 2). No symptoms could be found on the grass roots with control treatment. The fungal isolates with similar morphology were consistently recovered from all inoculated samples, and a representative isolate was selected for molecular identification by ITS sequencing. The morphology of the recovered fungal isolates was similar with the isolate 13g11 used for inoculation, and the ITS sequence matched that of the inoculated isolate with 100% identity. Foliar and root health were significantly reduced in inoculated samples compared with uninoculated samples.
GCPSR analysis
-
A PHI test using a six-locus concatenated dataset (ITS, LSU, SSU, MCM7, RPBI and TEF1) revealed that the recombination event was not observed between C. agrostis and C. cynodontis (Φw = 1.0), indicating that they were two different species. No recombination event was observed among the four isolates (13g11, 13g12, JH1 and JH2) within the new species C. agrostis (Φw = 1.0).
Taxonomy
-
In this study, phylogenetic analyses revealed an un-described species in the genus Candidacolonium causing summer patch-like disease in East China. The proposed new species was isolated from creeping bentgrass roots and is named Candidacolonium agrostis.
Candidacolonium agrostis J. Hu and J. M. Geng, sp. nov.
MycoBank: MB849256.
Typification: China, Jiangsu province: Wuxi city, 31°24′38.9″N, 120°07′24.4″E. Roots of Agrostis stolonifera, 11 Oct. 2020, J. Hu and J. M. Geng, 13g11 = THAS136. (holotype: not CDGM89573, ex-type culture: GDMCC 3.776.).
Additional specimens examined: China, Jiangsu province: Wuxi city, 31°24′38.9″N, 120°07′24.4″E. Roots of Agrostis stolonifera, 11 Oct. 2020, J. Hu and J. M. Geng, 13g12 = THAS137. China, Zhejiang province: Shaoxing city, 118°80′24.2″N, 32°06′46.5″E. Roots of Agrostis stolonifera, 15 Jul. 2021, J. Hu and Y. L. Dong, JH1. China, Zhejiang province: Shaoxing city, 118°80′24.2″N, 32°06′46.5″E. Roots of Agrostis stolonifera, 15 Jul. 2020, J. Hu and Y. L. Dong, JH2.
Morphological description: Colony diameter was measured approximately 19.0 mm on PDA at 25 °C in the dark after 5 d of incubation. The color of colony was bright white after 7-d growth, and pale grayish buff after 14- to 21-d growth. Reverse of the colony was pigmented dark grayish buff at the center of PDA plate. Hyphae were septate, 2.8-3.6 µm (mean = 3.2; S.D. = 0.3; n = 30) wide, and hyaline to brown in color. Brown stigmatopods measured 5.0-11.8 × 4.6-6.3 (mean = 7.0 × 5.4; S.D. = 1.8, 0.6; n = 30) Brown slightly lobed or mitten-shaped hyphopodia measured 7.9-11.8 × 4.2-8.5 µm (mean = 10.4 × 6.5; S.D. = 1.1, 1.1; n = 30).
Notes: C. agrostis was phylogenetically close to C. cynodontis. However, it clearly formed a single clade with high PP/BS values (1/100%). BLAST analyses showed that the ITS, LSU, MCM7, RPBI and TEF1 sequences of C. agrostis differed from C. cynodontis by 20, 1, 12, 18, 15 bp, respectively. No nucleotide differences were observed in LSU between the two species. C. agrostis grew faster than C. cynodontis on PDA plate in the dark at 25 °C, and its host was Agrostis stolonifera. The two species showed different colony colors[14]. The differences in the sizes of hyphae and hyphopodia were not apparent between the two species[14].
Etymology: The specific epithet refers to its host's generic name.
Distribution: East China
-
In this study, a new species in the genus Candidacolonium was identified, causing a summer patch-like disease on creeping bentgrass in East China. The concatenated six-gene and individual gene datasets were highly homologous in topology. BI and ML analyses were able to separate all the taxa into two main clades (A and B). Multiple plant pathogenic species in the genera Gaeumannomyces, Magnaporthiopsis, Buergenerula, Nakataea and Omnidemptus formed the clade A. Clade B comprised three distinct lineages, the novel species proposed in this study, C. cynodontis and the genus Psedophialophora belonged to the clade B. The concept of new species is well supported not only by the multi-locus phylogenetic analysis, but also by morphological and biological characteristics, and the root habit of the fungi.
Morphological features such as hyphopodial morphology have traditionally been used as identifying features for ERI fungi[18]. Similar to C. cynodontis, C. agrostis also produces slightly lobed or mitten-shaped hyphopodia, which are distinct to lobed or simple hyphopodia produced by the genus Gaeumannomyces[14,38]. It is appears that the slightly lobed or mitten-shaped hyphopodia may be characteristic of Candidacolonium spp. However, other ERI fungi, such as Slopeiomyces cylindrosporus, also produce similarly shaped hyphopodia[39]; therefore, these hyphopodia are not diagnostic for Candidacolonium spp.
The disease severity caused by C. agrostis was significantly different at the two sampled golf courses. This may be because they were sampled at different times, i.e. during a hot summer and a cool autumn. C. agrostis has been observed to cause severe damage on creeping bentgrass putting greens during hot weather. In vitro assays of temperature sensitivity also support that C. agrostis grows better at higher temperatures. Moreover, creeping bentgrass are weakened by many stressed factors during the hot summer months, and become increasingly vulnerable to ERI fungi[9]. Similar pathogenicity results were observed in other ERI fungi, including C. cynodontis, which caused more severe disease at 30 °C than at 20 °C in an in vivo evaluation[15]. The pathogenicity analysis in this study successfully recreated the disease symptoms; however, they were not as severe as observed by the superintendents at golf courses. This is probably because turfgrass plants used for the in vivo evaluation were grown in a growth chamber with 12-h day/night cycle at 26/22 °C, which was at near optimal conditions, and the plants were not stressed. To gain a better understanding of the role C. agrostis play in the summer patch-like disease, field conditions must be considered. A proposed evaluation strategy could be to inoculate C. agrostis isolates into established creeping bentgrass turf, and the research plots be managed according to golf course standards.
C. agrostis is demonstrated to be associated with canopy decline symptoms of creeping bentgrass during summer months. The symptoms of affected creeping bentgrass were similar to summer patch disease, which is a common and very destructive root disease of cool-season turfgrasses[5]. To date, the fungus M. poae is known as the only pathogen associated with root disease on creeping bentgrass in China. This study reports a new fungal species of Candidacolonium associated with summer patch-like symptoms on creeping bentgrass that has a different growth rate, morphology and genetics compared with the causal agent of summer patch M. poae. The results from this study confirmed the pathogenicity of C. agrostis on creeping bentgrass in the controlled environment and its association with creeping bentgrass at two golf courses in East China. By contrast, summer patch of creeping bentgrass in North China, is caused by M. poae. Summer patch-like disease on creeping bentgrass in East China has not been widely studied[2,6]. Further investigation into the distribution and prevalence of C. agrostis populations on creeping bentgrass in East China is warranted, and should lead to better understanding of the overall damage caused by this fungus in this region.
In conclusion, a novel ERI fungal species in the genus Candidacolonium was obtained and identified from roots of creeping bentgrass that showed summer patch-like disease at golf courses in East China. Prior to this study, M. poae was the only known ERI fungus in the Magnaporthaceae associated with summer patch on creeping bentgrass in China. The finding brings about inquiries concerning the role this fungal species plays in the root disease of creeping bentgrass putting greens during summer months in East China. Future studies may include developing the control methods, determining the host range, identifying the dissemination modes, as well as developing rapid diagnostic methods for this fungal species.
Jiangsu Institute of Botany Talent Fund, grant number: JIBTF202212. We thank Jiangsu Institute of Botany Talent Fund (grant number: JIBTF202212) for financial support, we also thank Zhang Hongliang from Wuxi Taihu Golf Club for helping collect the diseased samples and providing historical information of the disease.
-
The authors declare that they have no conflict of interest.
- Supplemental Fig. S1 Majority rule Bayesian phylogenetic tree derived from ITS DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Fig. S2 Majority rule Bayesian phylogenetic tree derived from rDNA (the combined ITS, LSU and SSU) DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Fig. S3 Majority rule Bayesian phylogenetic tree derived from MCM7 DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Fig. S4 Majority rule Bayesian phylogenetic tree derived from RPB1 DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Fig. S5 Majority rule Bayesian phylogenetic tree derived from TEF1 DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Fig. S6 A maximum likelihood phylogenetic tree derived from the combined ITS, LSU, SSU, MCM7, RPB1, and TEF1 DNA sequence data sets. Nodes having Bayesian inference posterior probabilities ≥0.95 and maximum likelihood bootstrap values ≥75% are shown. The tree is rooted with Pyricularia grisea M83.
- Supplemental Table S1 Species name, isolate IDs, and GenBank accession numbers for fungal isolates used for phylogenetic analyses in this study.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Geng J, Zhou Y, Dong Y, Lamour K, Yang Z, et al. 2023. Candidacolonium agrostis, a novel species associated with summer patch-like disease on Agrostis stolonifera in East China. Grass Research 3:20 doi: 10.48130/GR-2023-0020 

Candidacolonium agrostis, a novel species associated with summer patch-like disease on Agrostis stolonifera in East China
- Received: 30 May 2023
- Accepted: 31 August 2023
- Published online: 08 October 2023
Abstract: Creeping bentgrass (Agrostis stolonifera) is one of the most preferred turfgrass species for putting greens on golf courses throughout China. In autumn 2020 and summer 2021, a summer patch-like disease was observed on creeping bentgrass putting greens at two golf courses in East China. Fungal isolates with similar morphology were consistently isolated from the dark root tissues. Four representative isolates from the two golf courses were identified through morphological, biological, and phylogenetic analyses. Bayesian methods and maximum likelihood were used to construct phylogenetic trees based on both multiple and single loci of partial sequences of the 18S nuc rDNA (SSU), ITS1-5.8S-ITS2 nuc rDNA internal transcribed spacer (ITS), and 28S nuc rDNA (LSU) regions, and of the largest subunit of RNA polymerase II (RPB1), minichromosome maintenance complex 7 (MCM7), and translation elongation factor 1-alpha (TEF1) genes, respectively. The isolates consistently formed a highly supported clade within the genus Candidacolonium, which was further supported by distinctive morphological characters. In pathogenicity tests, the fungus produced slightly lobed or mitten-shaped hyphopodia, colonized roots of creeping bentgrass via ectotrophic, dark runner hyphae, and caused disease symptoms with root discoloration, root and shoot mass reduction, and yellow leaf spots. This pathogenic fungus is described as a new species, Candidacolonium agrostis sp. nov., and is most likely the cause of a summer patch-like disease on creeping bentgrass in East China.
-
Key words:
- Candidacolonium agrostis /
- hyphopodia /
- summer patch-like disease /
- creeping bentgrass /
- East China