









-
With the ever-changing environment and climate, soil salinization has become a major environmental issue. High salinity stress expands the area of crop damage and threatens both crop quality and yield[1]. Salt stress is a limiting factor in crop growth and development. Generally, plants under salt stress have a larger proportion of roots and thus favor the retention of toxic ions[2]. Additionally, salt-tolerant species accumulate Pro and GB for osmotic regulation[3], but accelerate starch consumption to cope with salt stress[4]. It is also well known that salt induces oxidative stress in plants; in response, salt-tolerant plants exhibit an upregulation of antioxidant defences[5].
Alfalfa (Medicago sativa L.) is a widely planted perennial forage crop with a well-developed root system, rich nutrition, and a certain level of stress resistance. Alfalfa can be used as a raw biological material for ethanol production and has great potential for the future energy revolution. It is also a soil and water conservation plant with important economic and ecological functions[6−8]. Although alfalfa is rich in nutrients and has high ecological adaptability, it has some limitations with respect to agricultural production. At this stage, it is important to use molecular breeding technology to breed highly resistant dominant varieties.
CYP450 monooxygenases (CYP450s) are enzymes that contain heme-thiolate domains and play important roles in plant growth, flavonoid synthesis, and other metabolic pathways[9]. CYP450s in plants constitute the largest family of enzymes related to plant metabolism, containing 127 subfamilies and accounting for approximately 1% of the total genes in the plant genome[10,11]. CYP450s have a conserved heme domain sequence, FxxGxRxCxG — usually located in the endoplasmic reticulum, mitochondria, Golgi apparatus, and other organelle membrane systems — that combines with different substrates to catalyze reactions. Based on their evolutionary relationships, plant CYP450s are divided into 11 clans (CYP51, CYP74, CYP97, CYP710, CYP711, CYP727, CYP746, CYP71, CYP72, CYP85, and CYP86); however, new families are still being discovered[9,10]. Since the discovery of CYP450s, members of the CYP450 protein family of many plants, including Arabidopsis, rice, corn, and thistle alfalfa (Medicago truncatula), have been isolated and identified[10,12]. Numerous studies have shown that CYP450s in plants participate in the synthesis of a variety of primary and secondary metabolites, such as phenylpropanes, terpenes, flavonoids, alkaloids, fatty acids, and plant hormones. CYP450s also participate in the synthesis of cell wall structural components, protection against pests and diseases, and the decomposition of toxic substances, such as herbicides and pesticides[13]. CYP51G, CYP85A, CYP90B, CYP710A, CYP724B, and CYP736A of the CYP450 family are relatively conserved in the plant kingdom, and are mainly involved in primary metabolism related to the biosynthesis of sterols, steroid hormones, saponins, phenylpropanes, and auxins, as well as in terpene metabolism[14,15]. CYP716, CYP72, CYP88, and other CYP450s play important roles in the structural diversification and functionalization of terpenoids.
In M. truncatula, CYP716A12 has a catalytic effect on β-vanilla, converting it to oleanolic acid. CYP93B10 and CYP93B11 play important roles in flavonoid synthesis, Hansen et al.[16] demonstrated that CYP716A47 regulates ginsenoside synthesis. Arabidopsis AtCYP79B2 and AtCYP79B3 catalyze the tryptophan synthesis of indole acetaldoxime, an auxin precursor, and Arabidopsis AtCYP85A2 participates in brassinosteroid synthesis. Transgenic plants with ectopic AtCYP79B2 overexpression exhibited traits such as dwarfing and sterility. Plant CYP71, CYP72, CYP76, and other subfamily members exhibit enhanced resistance to harmful foreign substances, while the overexpression of CYP71A10 in soybeans and heterologous expression of ginseng CYP736A12 in Arabidopsis enhanced plant tolerance to phenylurea herbicides. Under drought stress, the expression of the tobacco ABA hydroxylase genes CYP707A1, CYP94C1, and CYP94B3 significantly increased.
Although CYP450s represent a large gene family in plants, the functions of most CYP450s remain unidentified; additionally, few CYP450s have a high similarity in amino acid sequence. There are few studies on the role of CYP450s in plant tolerance to abiotic stresses, such as high temperatures, drought, and salt. Previous studies have shown that most expressed CYP450s contain cis-acting elements — such as the MYB-binding site, ACGT core sequence, or TGA-box — involved in plant resistance. Despite previous research on the structure and function of CYP450s, most studies on CYP450s have focused on the secondary metabolites of models and medicinal plants, and their effects on pest and disease resistance. Few studies have been conducted on the regulatory effects of CYP450s on plant resistance, especially regarding salt tolerance in forage plants. In a previous study, using transcriptomic data analysis, we found that many CYP450 genes in alfalfa responded positively to salt stress. It was thus indicated that these CYP450 genes play important roles in salt stress and regulate plant adaptability to coercion.
CYP450s play a crucial role in the regulation of flavonoid synthesis and plant growth; however, members of the CYP450 family in alfalfa have not been analyzed and identified at the genomic level. We therefore aimed to identify the MsCYP (M. sativa CYP450) genes involved in the salt stress response and quality of alfalfa via genomic sequencing, as well as analyze a CYP450 gene model, phylogenetic relationships, chromosome locations, and other structural features. The expression patterns of key CYP450 genes were analyzed using RNA-seq (RNA sequencing) and RT-qPCR (Real time quantitative PCR). This study therefore lays the foundation for the exploration of CYP450 gene function, and provides valuable information for improving alfalfa varieties under high stress.
-
The genome sequence was downloaded from the Alfalfa Genome Project website (
https://Figureshare.com/articles/dataset/Medicago_sativa_genome )[17]. The HMM (Hidden Markov Model) of the CYP450 domain (PF00067) was obtained from Pfam (http://pfam.xfam.org/family/PF00067#curationBlock ) and used to search for homologous sequences in the alfalfa protein sequence file using hmmer-3.0-windows. Furthermore, the CYP450 sequences of Arabidopsis were used to blast the alfalfa protein file (e-value 1e−5). The total protein sequences were obtained by combining these two methods. To confirm the reliability of the search, all candidate sequences obtained were checked using Pfam (http://pfam.xfam.org/search#tabview=tab1 ) and NCBI-CDD (https://www.ncbi.nlm.nih.gov/Structure/bwrpsb/bwrpsb.cgi ) for conserved domain identification. Redundant sequences were removed, and nonredundant CYP450 sequences were used for sequence alignment and further analyses, including the determination of chromosomal location, isoelectric point, and subcellular localization.Phylogenetic analysis and gene structure
-
CYP450 protein sequences were extracted from Arabidopsis thaliana and alfalfa. The sequences of CYP450 proteins from all plant species were compared using the MAFFT software. The FastTree software was used to construct a phylogenetic tree using the largest natural control with a bootstrap value of 1,000. The gene and coding sequences of MsCYP genes were used to analyze the gene structure using TBtools[18].
Chromosomal localization and gene replication
-
Duplicate genes in the MsCYP gene family were identified using one-step MCScanX in TBtools v1.098746. Simple Ka/Ks calculators (NG) were employed to calculate Ka and Ks using TBtools v1.098746. The Arabidopsis genome sequence was downloaded from
https://www.ncbi.nlm.nih.gov/genome/?term=Arabidopsis+thaliana ; the rice genome sequence was downloaded fromhttps://data.jgi.doe.gov/refine-download/phytozome?organism=Osativa&expanded=Phytozome-323 ; the Glycine max genome sequence was downloaded fromhttps://ngdc.cncb.ac.cn/search/?dbId=gwh&q=GWHAAEV00000000&page=1 ; and the M. truncatula genome sequence was downloaded fromhttps://www.ncbi.nlm.nih.gov/genome/?term=Medicago+truncatula+ . Pairs of duplicated genes in M. sativa, Arabidopsis, Oryza sativa, G. max, and M. truncatula were analyzed in the same way as described above. The relationships between duplicates in the MsCYP gene family were plotted using Advanced Circo in TBtools v1.098746, and the relationships between duplicates among these species were plotted using multiple synteny plots in TBtools v1.098746.Prediction of cis-acting elements in the upstream promoter of MsCYP genes
-
Promoter sequences (2,000 bp upstream of the MsCYP genes) were obtained from the alfalfa genome using Gtf/Gff3 Sequence Extract and Fasta Extract in TBtools v1.098746. The upstream 1,500 bp sequences were prefetched as promoters and submitted to PlantCARE (
https://bioinformatics.psb.ugent.be/webtools/plantcare/html/ ) to identify cis-acting elements.Plant materials and salt treatment
-
M. sativa (Zhong Mu No. 1) seeds were sown in soil and grown for 10 d, then hydroponically cultivated for 12 d. Tobacco (Nicotiana benthamiana), and A. thaliana were cultivated using nutrient soil in the Plant Growth Laboratory of Ningxia University (China). The culture temperature was 25 °C, and the photoperiod was 16 h light and 8 h dark. The roots of 22-day-old alfalfa seedlings were exposed to 200 mM NaCl for 2 h and recovered using hydroponic experiments. The samples were collected at three time points — before salt treatment (N0), after salt treatment for 2 h (N2), and after rehydration for 3 d (NH) — from three different plants and immediately stored at −80 °C until further use.
RNA extraction and qPCR
-
RNA was extracted from frozen samples using a MiniBEST Plant RNA Extraction Kit (Takara Bio Inc., Shiga, Japan). cDNA was generated with PrimeScript™ RT Master Mix (Takara Bio Inc.), and RT-qPCR was performed in triplicate using a Roche Light Cycler 480 (Roche, Basel, Switzerland) with TB Green® Premix Ex Taq™ II (Tli RNaseH Plus; Takara Bio Inc.) according to the manufacturer’s instructions. Gene-specific primers (Supplemental Table S1) were designed to amplify the nine MsCYP genes using Primer Premier5.0. Actin was used as an internal control gene, and the relative expression levels of these MsCYPs were estimated via the 2−ΔΔCᴛ method.
RNA-Seq sequencing
-
A library was sequenced on an Illumina Novaseg 6000 platform. Fastp was used to remove adapter and low quality reads from raw data. The clean reads were then mapped to the alfalfa genome using Hisat2. Reads counts was obtained with Htseq-count, and differential expression analysis was performed using the DESeq R package. Reads counts were transformed into FPKM with R.
Plasmid construction and genetic transformation
-
The full-length coding sequence of MsCYP273 was amplified from the cDNA of alfalfa, and was inserted upstream of GFP harbored by the pGreen0029 vector via homologous recombination. Relevant negative and positive controls were produced simultaneously. The resulting constructs were then transiently transformed into tobacco leaves. Fluorescence was detected using an SP8 confocal microscope (Leica) at excitation wavelengths of 488 nm, and emission wavelengths of 495–545 nm for GFP, 590–670 nm for chloroplasts. To generate MsCYP273 overexpression lines, the coding sequence of the MsCYP273 protein was inserted into the pCAMBIA1300 plasmid, which was stably transformed into Arabidopsis using A. tumefaciens (GV3101) via the Arabidopsis floral dip method, as previously described[19].
-
To identify CYP450 genes in the alfalfa genome, an HMM search was performed against the alfalfa protein database using BLASTP. A total of 376 MsCYP genes were identified and named MsCYP1–MsCYP376 according to their chromosomal locations. The number of MsCYP amino acid sequences ranged from 120–984. The molecular weight of MsCYP proteins ranged from 13.62–110.08 kDa, and the isoelectric point ranged from 4.53–10.17. The WoLF PSORT tool predicted that approximately 192 (51.06%) and 100 (26.60%) MsCYPs were located in the chloroplast and cytosol, respectively; the remaining MsCYPs were located in the nucleus, mitochondria, and peroxisomes (Supplemental Table S2).
Phylogenetic classification of members of the MsCYP gene family
-
To investigate the evolutionary relationships of the CYP gene family in alfalfa, the CYP450 sequences of A. thaliana, M. truncatula, and M. sativa were used to construct a phylogenetic tree using FastTree (Fig. 1). The MsCYP proteins are divided into nine major clans: CYP71, CYP51, CYP72, CYP74, CYP85, CYP86, CYP97, CYP710, and CYP711. The CYP71 clan was of the A-type, while the other clans were of the non-A-type. CYP71, CYP86, CYP85, CYP72, CYP97, and CYP74 contained 236, 48, 35, 39, three, and two MsCYP proteins, respectively. The CYP51, CYP710, and CYP711 clans contained only one MsCYP protein (Fig. 1). Many clans, including CYP71, CYP86, CYP72, and CYP74, were significantly more abundant in M. sativa than A. thaliana or M. truncatula. In the CYP711, CYP97, and CYP51 clans, the gene phylogeny roughly followed the species phylogeny, with the genes of M. sativa displaying a sister-group relationship with M. truncatula, and one CYP450 gene in Arabidopsis closely related to alfalfa homologs. In particular, the phylogeny of the CYP85 clan was more complex, suggesting multiple duplication events during the evolution of the dicot CYP450 gene family.
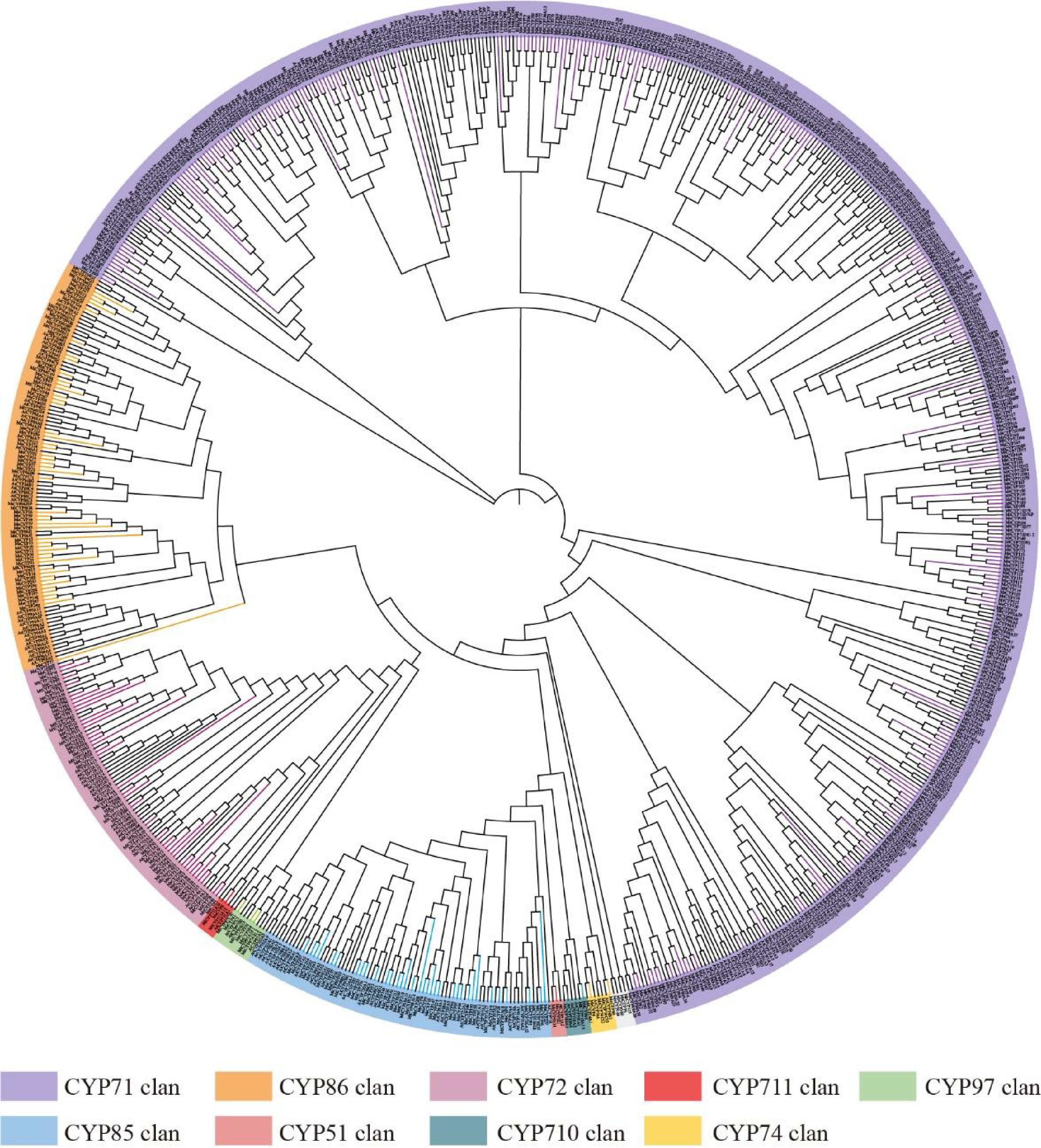
Figure 1.
Phylogenetic analysis of CYP genes from alfalfa, thistle alfalfa, and Arabidopsis. Different colors represent different groups.
Sequence analysis of the MsCYP gene family
-
To understand their functional regions, conserved motifs of the MsCYPs were analyzed. The motif distribution of each MsCYP protein was analyzed using MEME, and most members of the MsCYP gene family contained motifs 5, 3, 7, 10, and 11. Additionally, the arrangement and composition of the motifs were consistent with the phylogenetic tree results. For example, most CYP71 clan members contain motifs 8, 9, 2, 5, 3, 7, 4, 10, 1, and 6; however, these motifs were not identified in CYP76, CYP200, CYP208, or CYP259. All MsCYP proteins contain conserved CYP450 or P450 superfamily domains, and each MsCYP coding sequence is separated by introns ranging from 0–16. Most MsCYP proteins in each sample contained similar numbers of introns; most members of the CYP97, CYP72, and CYP85 clans have more introns, whereas members of the other clans have fewer introns (Fig. 2). Generally, the motif composition of the members within each clan was similar, indicating that the protein structure was highly conserved and further validating the reliability of the phylogenetic tree (Fig. 2).
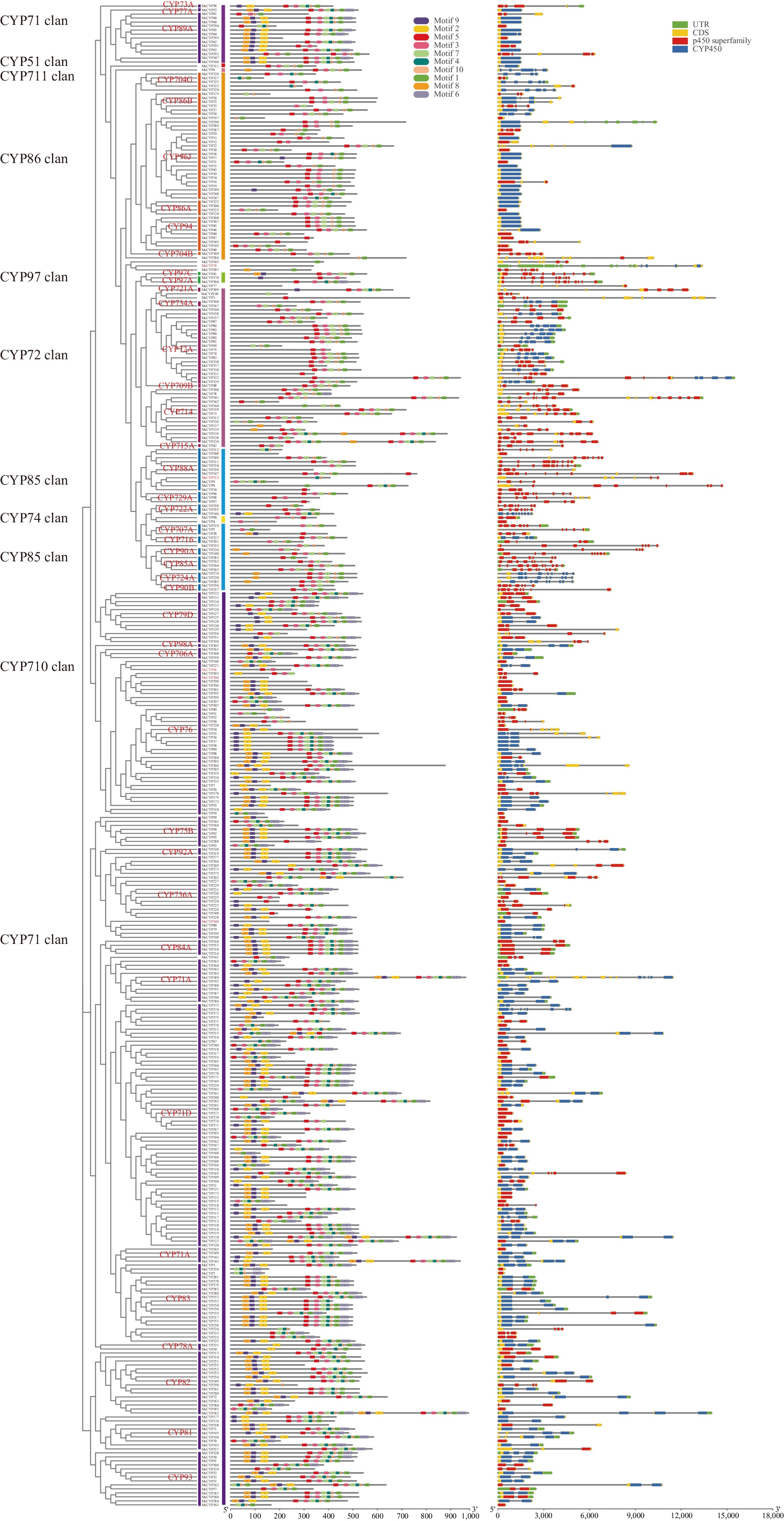
Figure 2.
Phylogeny, conserved motifs, and exon–intron structure of MsCYP genes.
All MsCYP proteins contained either the P450 superfamily or CYP450 domain, indicating that they had a similar nature or function. The MsCYP sequence contained introns 1–13, and both the number and phase of MsCYP genes were well conserved in the same clan. For example, each MsCYP gene in CYP706A contained two phase 0 introns. The number of introns in the MsCYP gene of CYP724A ranged from 9–10 (Fig. 2).
Chromosomal mapping and gene duplication of the MsCYP gene family
-
Based on newly published genome-wide data from alfalfa, the distribution of MsCYPs on chromosomes was analyzed. The 376 MsCYP genes were not evenly distributed on the eight chromosomes, with more genes identified on chromosomes 1 and 8 (59 and 58 genes, respectively). A total of 56 MsCYP genes were located on chromosomes 3 and 4, 44 on chromosome 5, 31 on chromosomes 2 and 6, and 33 on chromosome 7 (Fig. 3).

Figure 3.
Chromosomal locations of MsCYP genes. Tandem duplicates are marked with curved lines.
To explore the relationship between the evolutionary expansion of the MsCYP gene family and whole-genome duplication events in alfalfa, tandem duplication and segmental replication events in MsCYP genes were analyzed using the one-step MCScanX method. A total of 112 MsCYP genes in the MsCYP family were involved in tandem duplication, accounting for 29.8% of all MsCYP genes. A total of 64 tandem duplication pairs were obtained, which were distributed on eight chromosomes; most occurred on chromosomes 3 and 4 (Fig. 3). Only 37 genes (approximately 10%) were involved in segment duplication, and 19 pairs of segment duplication genes were obtained; these were distributed on chromosomes 1, 3, 4, 5, 6, 7, and 8. Chromosome 2 was not involved in segment duplication (Fig. 4). Tandem duplication events are thought to be a major driver of the expansion of the MsCYP family. Notably, six genes (CYP30, CYP106, CYP121, CYP165, CYP191, and CYP203) were involved in both segment and tandem duplications.
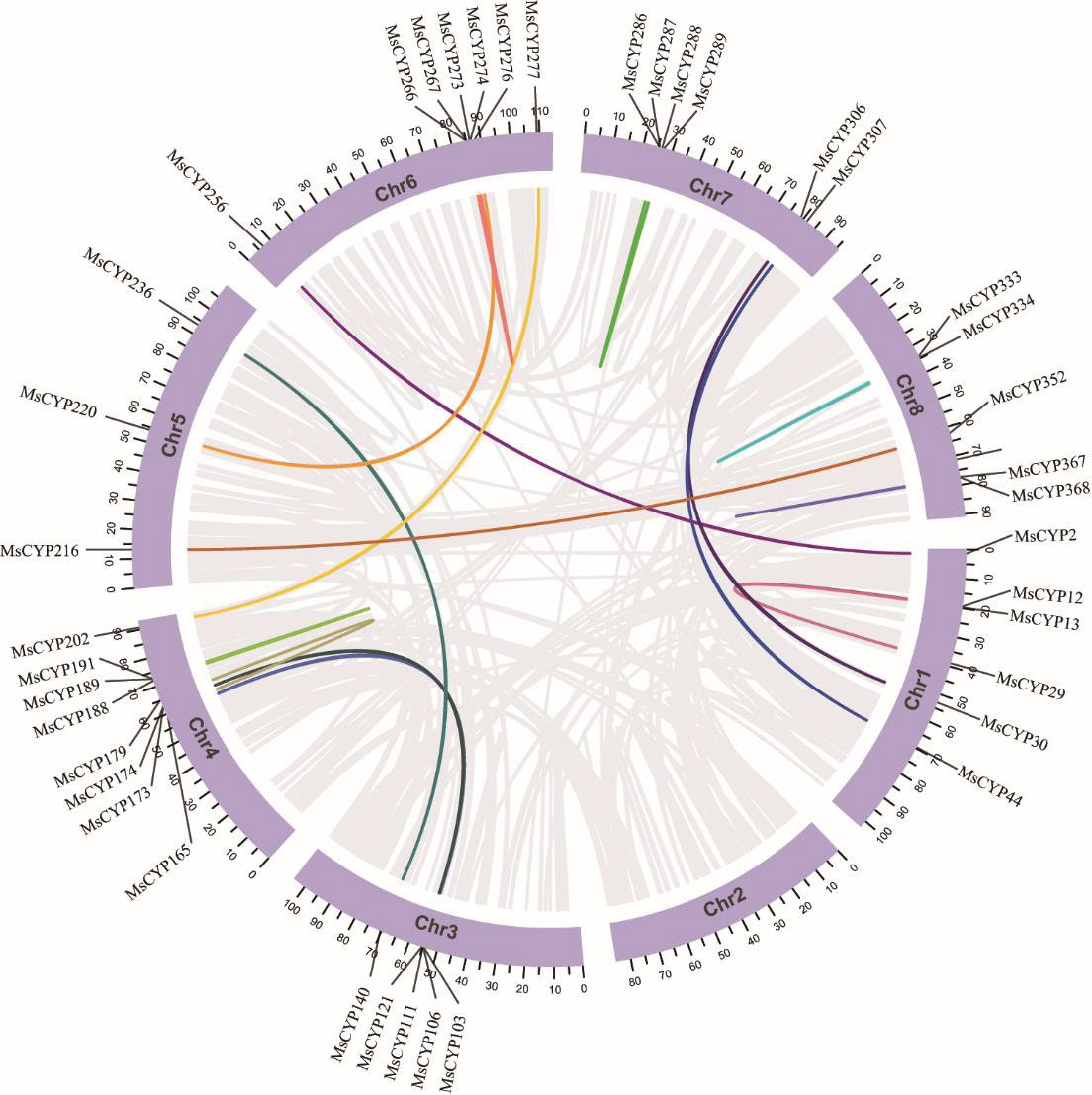
Figure 4.
Synteny analysis of alfalfa CYP genes. Different segmental duplicate pairs are linked with different colors.
To further explore the evolutionary processes of the MsCYP family, four comparative syntenies of alfalfa with rice, Arabidopsis, soybean, and thistle alfalfa were constructed. Alfalfa and thistle alfalfa shared the most orthologous pairs with up to 173 pairs of orthologous CYP450s, followed by alfalfa and soybean with 163 pairs. We also identified 44 pairs of orthologous CYP450s between alfalfa and Arabidopsis, and 15 pairs between alfalfa and rice. A closer relationship has been suggested between alfalfa and thistle alfalfa/soybean than between Arabidopsis and rice (Supplemental Fig. S1).
To explore the selection pressures acting on the CYP450 gene family, Ka, Ks, and Ka/Ks values were calculated for these gene pairs. All Ka/Ks values of the duplicated gene pairs from the alfalfa CYP450 gene family were < 0.8, excluding MsCYP2/MsCYP256. Some orthologous gene pairs had Ka/Ks values < 0.7; however, the remaining orthologous gene pairs were highly divergent between alfalfa and Arabidopsis/soybean/thistle alfalfa. All orthologous gene pairs were highly divergent between alfalfa and rice (Supplemental Fig. S2). These results suggest that the MsCYP gene family underwent strong purifying selection.
Analysis of key MsCYP genes
-
Using RNA-seq and RT-qPCR analyses before and after salt stress and rehydration (N0, N2, and NH, respectively), we identified nine MsCYP genes (MsCYP273, MsCYP272, MsCYP275, MsCYP266, MsCYP271, MsCYP267, MsCYP268, MsCYP89, and MsCYP274) believed to play vital roles in the response to salt stress. These genes were highly expressed in the N2 group and exhibited reduced expression in the N0 and NH groups (Fig. 5).
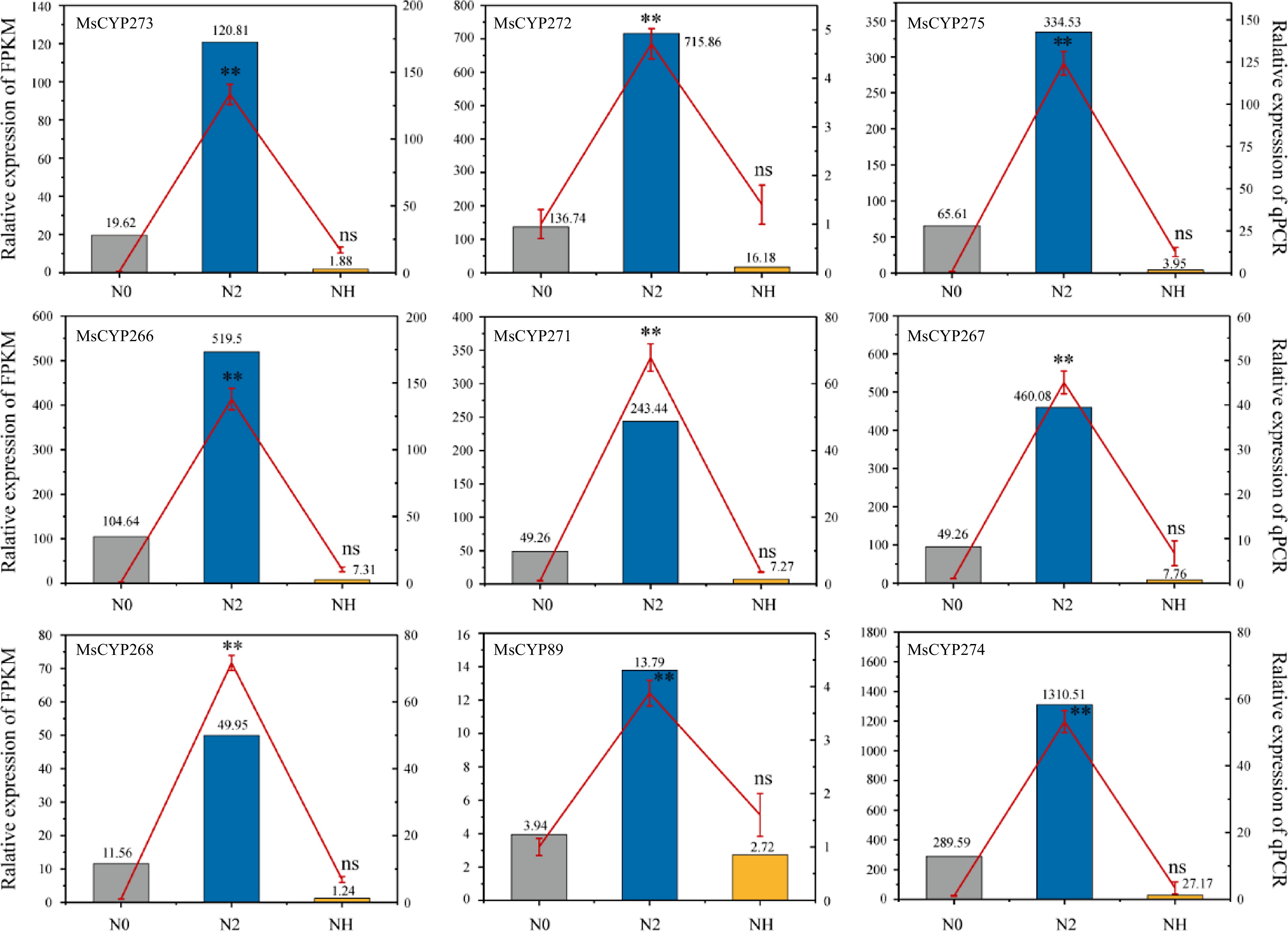
Figure 5.
Expression pattern of nine key MsCYP genes. The column indicates RNA-seq data, the red line indicates RT-qPCR data, and the error bar indicates the standard error of the mean (three biological replicates and three technical replicates). **, p < 0.01. ns, not significant. N0 refers to gene expression before salt treatment; N2 refers to gene expression after salt treatment for 2 h; and NH refers to gene expression after rehydration for 3 d.
To further predict the function of these nine MsCYP genes, cis-elements in the promoters were analyzed, and 72 elements were identified (Supplemental Fig. S3). Most elements were associated with development, especially the light response, including the 3-AF1 binding site, AAAC motif, ACE, and AE box. CAAT and TATA boxes were found in the promoter regions of all MsCYP genes. The MYB-binding site, known for its role in the drought response, was identified in MsCYP273, MsCYP272, MsCYP275, MsCYP266, MsCYP268, and MsCYP89. Among these, MsCYP273 was selected for transient expression in tobacco and heterologous overexpression in Arabidopsis. MsCYP273 was mainly located in the chloroplasts, with some distribution in the cell membrane and cytoplasm (Fig. 6b). In addition, five MsCYP273 overexpression lines were obtained, all of which had an expression of MsCYP273 (Fig. 6a). Compared to the wild type, MsCYP273 overexpression lines demonstrated stronger resistance to 75 mM NaCl stress (Fig. 6c, d).
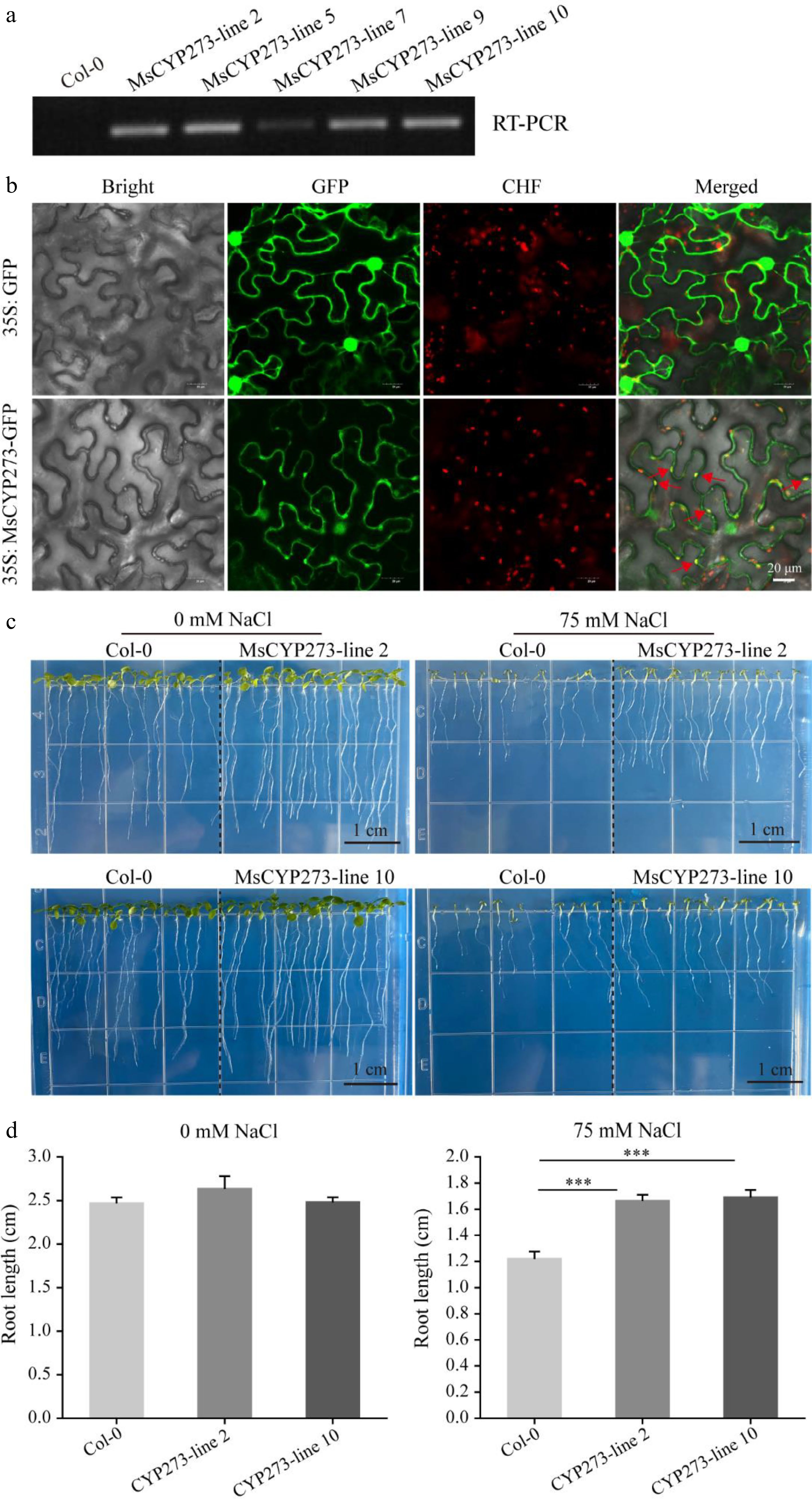
Figure 6.
Analysis of the MsCYP273 protein. (a) RT-PCR results of five MsCYP273 overexpressing lines. (b) Subcellular localization of MsCYP273 protein. Bar = 20 μm. Bright: Bright field; GFP: green fluorescent protein of 495-545 nm; CHF: chloroplast autofluorescence of 590–670 nm. (c) Phenotype of MsCYP273 overexpressing lines (MsCYP273-line2 and MsCYP273-line10) under salt stress. (d) Root length analysis of MsCYP273 overexpressing lines (MsCYP273-line2 and MsCYP273-line10) under salt stress. Bar = 1 cm. ***, p < 0.001.
-
Cyclophilins are involved in several physiological processes, including protein transport, transcriptional regulation, signal transduction, mRNA splicing, cell apoptosis, and stress response. The total number of CYP450 genes in a single plant species is usually higher than in animals[16]. Comprehensive analyses of CYP450 gene families in several plants have been published, and gene duplication has played an important role in the expansion of the gene family. A total of 62 CYP450 genes were identified in soybeans (G. max); 54 were clustered in pairs (27 pairs) in the phylogenetic tree[20]. In the apple (Malus domestica) genome, 30 CYP450 genes were identified; there were no tandem duplicated gene pairs, and 10 segmental duplicated gene pairs. There are 188 CYP450 genes in Panax notoginseng, with eight tandem and 11 segmental duplication events[21]. In the Tartary buckwheat (Fagopyrum tataricum) genome, 285 CYP450 genes have been identified, with 62 pairs of tandemly duplicated and 18 pairs of segmentally duplicated genes. A total of 355 CYP450 genes were identified in rice (O. sativa); more than half of these genes were found in 53 tandem duplicated gene clusters, while 55 OsCYP450s were distributed in segmental duplication blocks[22]. In this study, 376 MsCYP genes were identified in alfalfa; 144 genes were involved in gene duplication, resulting in 64 tandemly duplicated gene clusters and 19 segmentally duplicated gene pairs (Figs 3 & 4). Thus, gene duplication has played a significant role in the evolution of the plant CYP450 gene family. In particular, the Tartary buckwheat, alfalfa, and rice CYP450 gene families are prone to differential expansion due to tandem duplication during evolution.
Many alfalfa CYP450 genes have evolutionarily conserved functions
-
Gene duplication is the main source of redundancy and functional specialization in the evolution of plant genomes. For example, the duplication and neofunctionalization of CYP98 subfamily genes have enabled hydroxylation during rosmarinic acid biosynthesis in some lamiid plants[23]. The CYP450 KLUH/CYP78A5 and its homolog, CYP78A7, promote organ growth via a noncell-autonomous signaling pathway. TaCYP81D5, a tandemly arranged CYP81D gene, confers salinity tolerance by scavenging reactive oxygen species (ROS) in bread wheat. In the present study, we identified nine key MsCYP genes (MsCYP273, MsCYP272, MsCYP275, MsCYP266, MsCYP271, MsCYP267, MsCYP268, MsCYP89, and MsCYP274), all of which were clustered in the CYP71 clan and may play vital roles in the response to salt stress (Fig. 1). Furthermore, MsCYP266, MsCYP273, and MsCYP275 clustered closely in the CYP736A subfamily, whereas MsCYP268, MsCYP272, MsCYP274, MsCYP271, and MsCYP267 clustered closely in the CYP71D subfamily. In particular, MsCYP266/MsCYP273 and MsCYP267/MsCYP274 were segmentally duplicated gene pairs, both of which showed similar expression patterns under salt stress conditions. Thus, further studies are needed to explore the functional relevance of similar duplicated genes to confirm whether neofunctionalization and subfunctionalization of alfalfa CYP450 genes influence the adaptation of alfalfa to diverse conditions.
Some alfalfa CYP450 genes play an important role in the plant response to salt stress
-
Salt stress is a major environmental factor that can adversely alter plant growth and development and reduce global crop production. CYP450 enzymes play significant roles in the salt stress response of plants. In this study, we identified nine MsCYP genes that exhibited increased expression during salt stress, and decreased expression during recovery (Fig. 5); this was consistent with the results of previous studies. For example, AtCYP709B3 is markedly induced by salt stress and remains highly expressed, while mutant Arabidopsis plants are sensitive to ABA and salt during germination[24]. The overexpression of PgCYP736B in Arabidopsis confers enhanced resistance to salt stress by decreasing hydrogen peroxide accumulation, thereby increasing carotenoid levels and abscisic acid biosynthesis gene expression[25]. The heterologous expression of AoCYP94B1 in Arabidopsis and rice seedlings confers salt tolerance by enhancing root suberin deposition[26]. PagWOX11/12a induces the expression of PagCYP736A12 to modulate ROS scavenging, thus enhancing salt tolerance in 84 K poplar (Populus alba × P. glandulosa)[27]. PtCYP714A3 is markedly induced by salt stress, and transgenic rice plants exhibit reduced shoot growth and enhanced salt tolerance[28]. OsCyP20-2, located in the thylakoid, is thought to be involved in photosynthetic acclimation to help plants cope with environmental stress[29]. OsCyP2-P, localized in both the cytosol and nucleus, is upregulated in response to salt stress, and the ectopic expression of OsCyP2-P imparts salt stress tolerance via ROS scavenging and ion homeostasis[30]. GhCyp1 expression was higher in the roots and stems, and overexpression of GhCyp1 conferred higher tolerance to salt stress in Pseudomonas syringae pv. tabaci infection[31]. CcCYP is predominantly localized in the nucleus, and transgenic plants exhibit high tolerance to major abiotic stresses, as evidenced by increased chlorophyll levels, biomass, and plant survival[32]. In particular, transgenic plants display higher Na (+) ion accumulation to maintain ion homeostasis in the roots under salt stress[32].
-
Alfalfa (M. sativa L.) is a perennial forage crop planted worldwide that has a well-developed root system and salt tolerance. In this study, we identified nine MsCYP genes thought to play vital roles in the response to salt stress that were highly expressed during salt stress and decreased during recovery. Additionally, MsCYP273 overexpression plants showed stronger resistance to NaCl stress than wild-type plants. In summary, the identification of these MsCYPs provides a vital foundation for their possible functions in stress breeding of alfalfa. Further studies are needed to explore the functional relevance of similar genes and confirm their functional characteristics under diverse conditions.
-
The authors confirm contribution to the paper as follows: study conception and design: Liu X and Ma D; data collection: Zhang X and Xue L; analysis and interpretation of results: Chen R, Ma Q, Liu X, Zhang X and Xue L; draft manuscript preparation: Liu X, Zhang X and Xue L. All authors have read and agreed to the publication of the manuscript.
-
RNA-seq data generated in this study were deposited in NCBI under a bioproject, accession numbers as follows: the three N0 biological repeats—SRR26104474, SRR26104473, and SRR26104472; the three N2 biological repeats—SRR26104471, SRR26104470, and SRR26104469; and the three NH biological repeats—SRR26104468, SRR26104467, and SRR26104466.
This study was supported by Ningxia Natural Science Foundation (2022AAC03007) and The Key R&D Program of Ningxia (2021BEB04075). Thanks to OmicShare, Guangzhou, China for the RNA-seq analysis. Thanks to Elsevier for language editing assistance during the preparation of this manuscript.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Xinyu Zhang, Li Xue
- Supplemental Table S1 Primer sequences used in RT-qPCR.
- Supplemental Table S2 The nomenclature and physicochemical properties of MsCYP450 genes.
- Supplemental Fig. S1 Synteny analysis of CYP450 genes form alfalfa with rice, Arabidopsis, soybean, and thistle alfalfa.
- Supplemental Fig. S2 Ka and Ks analysis of plant CYP450 duplicated genes. The number (A), Ka (B), Ks (C), and Ka/Ks (D) of CYP450 duplicated genes. Segment, segmental duplicated gene pairs; tandem, tandem duplicated gene pairs; Ms-Os, CYP450 duplicated gene pairs between alfalfa and rice; Ms-At, CYP450 duplicated gene pairs between alfalfa and Arabidopsis; Ms-Gm, CYP450 duplicated gene pairs between alfalfa and soybean; Ms-Mt, duplicated gene pairs between alfalfa and thistle alfalfa.
- Supplemental Fig. S3 Different cis-elements in the promoters of nine MsCYP genes. White squares represent the absence of a cis-element in the promoter of MsCYP genes, and the number in the square represents the number of cis-elements in the promoter.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Zhang X, Xue L, Chen R, Ma Q, Ma D, et al. 2023. Genome-wide identification of the cytochrome P450 family and analysis of CYP regarding salt tolerance in Medicago sativa L. Grass Research 3:21 doi: 10.48130/GR-2023-0021 

Genome-wide identification of the cytochrome P450 family and analysis of CYP regarding salt tolerance in Medicago sativa L.
- Received: 09 May 2023
- Accepted: 17 October 2023
- Published online: 07 November 2023
Abstract: With the ever-changing environment and climate, high salinity has become a major environmental issue that threatens crop quality and yield. Alfalfa (Medicago sativa L.) is a perennial forage crop planted worldwide that has a well-developed root system and salt tolerance. Cytochrome P450 monooxygenase (CYP450) genes play important roles in flavonoid synthesis, plant growth, and development. However, few studies have focused on CYP450s in forage grass, especially the role of CYP450 genes in plant resistance to environmental stresses, such as drought and high salinity. In this study, 376 menbers in MsCYP family genes were identified using hmmsearch and BLASTP in the alfalfa protein database using the AtCYP450 protein sequence. Then by exploring MsCYP gene structures, tandem and segmental duplication events, and evolutionary relationships with CYP450s in other plants, potential MsCYPs that respond to salt stress were screened. Candidate genes were selected for transient expression in tobacco and heterologous overexpression in Arabidopsis for salinity response. This study provides a foundation for verifying the function of MsCYPs in improving the quality of agricultural products.
-
Key words:
- Alfalfa /
- Salt stress /
- Phylogenetic analysis /
- CYP450 gene family /
- Transcriptome