








-
Sordariomycetes O.E. Erikss. & Winka, is the second largest class in Ascomycota Caval.-Sm., characterized by unitunicate asci and are widely distributed in almost all ecosystems as endophytes, pathogens, or saprobes[1]. Hyde et al.[1], accepted 45 orders, 167 families, 1,499 genera and 308 genera incertae sedis in Sordariomycetes, while in the latest outline by Wijayawardene et al.[2], the numbers were increased to 46 orders, 184 families, 1,594 genera and 346 genera incertae sedis. In Sordariomycetes, there are seven subclasses viz. Diaporthomycetidae Senan., Maharachch. & K.D. Hyde, Hypocreomycetidae O.E. Erikss. & Winka, Lulworthiomycetidae Dayar., E.B.G. Jones & K.D. Hyde, Pisorisporiomycetidae Bundhun, Maharachch. & K.D. Hyde, Savoryellomycetidae Hongsanan, K.D. Hyde & Maharachch., Sordariomycetidae O.E. Erikss & Winka, and Xylariomycetidae O.E. Erikss & Winka[2].
Xylariales Nannf. is a large order and the only order in the subclass Xylariomycetidae which was accepted by Maharachchikumbura et al.[3] and later Amphispheriales and Xylariales were accepted in Xylariomycetidae by Samarakoon et al.[4] and Hongsanan et al.[5] based on phylogenetic analyses and divergence time estimations. Later, Delonicicolales R.H. Perera, Maharachch. & K.D. Hyde was introduced into Xylariomycetidae based on phylogenetic analyses by Perera et al.[6]. According to Wijayawardene et al.[2], currently, 20 families are listed in this order.
Diatrypaceae Nitschke is one important family of higher ascomycetes that belongs to Xylariales[3]. Diatrypaceae was introduced by Nitschke[7] and is typified by Diatrype Fr. Diatrypaceae is characterized by perithecial black stroma, ascomata usually embedded in stroma, cylindric-clavate to clavate asci and allantoid ascospores in its sexual morph[8−12]. The asexual morph is characterized by coelomycetous; acervuli conidiomata, erumpent, with branched conidiophores, conidiogenous cells in dense palisades, cylindrical with filiform conidia, curved, flattened base, blunt apex, and hyaline[1,3]. Zhu et al.[13] showed that Diatrypaceae has high diversity, and the members of this family are usually wood inhabiting fungi in China. According to Wijayawardene et al.[2], currently, 22 genera are listed in this family.
Peroneutypa Berl. was introduced by Berlese[14] to accommodate P. bellula (Desm.) Berl., P. corniculata (Ehrh.) Berl and P. heteracantha (Sacc.), without designating the type species until Rappaz[9] designated P. bellula as the type species and considered Peroneutypa as a synonym of Eutypella (Nitschke) Sacc. Later, based on morphological characteristics and phylogenetic analyses of Acero et al.[15], Carmarán et al.[16] resurrected Peroneutypa as an independent genus, and transferred eight species from Eutypella and Echinomyces Rappaz to Peroneutypa (viz: P. alsophila (Durieu & Mont.) Carmarán & A.I. Romero, P. arecae (Syd. & P. Syd.) Carmarán & A.I. Romero, P. comosa (Speg.) Carmarán & A.I. Romero, P. curvispora (Starbäck) Carmarán & A.I. Romero, P. gliricidiae (Rehm) Carmarán & A.I. Romero, P. kochiana (Rehm) Carmarán & A.I. Romero, P. scoparia (Schwein.) Carmarán & A.I. Romero and P. obesa (Syd. & P. Syd.) Carmarán & A.I. Romero). Later, based on molecular phylogenetic analyses, several studies have clarified that Peroneutypa is a monophyletic group in Diatrypaceae[12,16−20]. Peroneutypa is characterized by poorly developed ascostromata, perithecia ascomata with long prominent necks, 8-spored, clavate asci with sessile to subsessile, and allantoid, hyaline or yellowish ascospores[11,12,16−24]. Recently, a new species (P. polysporae Devadatha, V.V. Sarma & E.B.G. Jones) isolated from decaying wood of Suaeda monoica in India was introduced by Dayarathne et al.[25]. This species has polysporous asci which differ from the generic description (8-spored asci), thus this information shows that Peroneutypa can accommodate taxa with polysporous asci[25]. The members of Peroneutypa are known as saprobes or pathogens and are widely distributed in terrestrial and marine habitats[12,25−28]. Currently, 35 species are recorded in Index Fungorum[29], with only 16 species having molecular data.
In this study, a new species, Peroneutypa aquilariae is introduced based on morphological characteristics and phylogenetic analyses of combined ITS and tub2 gene sequences. To our knowledge, this is the first Peroneutypa species from an Aquilaria host. In addition, a synoptic table of
morphological characteristics in Peroneutypa is also provided. -
The dead twigs of Aquilaria sinensis with fungal fruiting bodies were collected from Xishuangbanna, Yunnan Province, China in autumn. The specimens were placed in plastic bags and brought to the mycology laboratory. Morphological structures were examined under an OPTEC SZ650 dissecting stereo-microscope. Fruiting bodies were picked up with needles, hand sectioned by razor blades and mounted on a glass slide with water. OLYMPUS optical microscope (Japan) was used to observe microscopic fungal structures and an OLYMPUS DP74 (Japan) digital camera fitted to the microscope was used to take photographs. All micromorphological structures were measured with Tarosoft® Image Framework program and photo plates were made using Adobe Photoshop CS3 Extended version 10.0 software (Adobe Systems, USA).
Senanayake et al.[30] was followed for the single spore isolation under sterile conditions. Once the spores were germinated, clear photographs were taken using the camera microscope and the germinated spores were transferred to a new potato dextrose agar (PDA) medium with a sterilized needle, and then incubated at 28 °C. Culture characteristics on PDA were observed after one month. The fresh mycelia of the fungus were used for DNA extraction.
The herbarium specimen was deposited in Kunming Institute of Botany Academia Sinica (HKAS), China, while the living cultures were deposited in Kunming Institute of Botany Culture Collection (KUNCC), China. Facesoffungi (FoF) was registered as described in Jayasiri et al.[31], and MycoBank number (MB) was registered as outlined in MycoBank (
www.MycoBank.org ).DNA extraction, PCR amplification, and sequencing
-
Genomic DNA was extracted from fungal mycelium (from one-month old cultures) by using Biospin Fungus Genomic DNA Extraction Kit–BSC14S1 (BioFlux®, China) following the manufacturer's instructions. Polymerase chain reaction (PCR) was used to amplify two gene regions using primers following Shang et al.[12] and Dayarathne et al.[25], with some modifications (Table 1). The total volume of PCR mixtures for amplifications following Du et al.[32], 25 μL containing 12.5 μL 2x Master Mix (mixture of Easy Taq TM DNA Polymerase, dNTPs, and optimized buffer (Beijing Trans Gen Biotech Co., Chaoyang District, Beijing, China), 8.5 μL ddH2O, 2 μL of DNA template, 1 μL of each forward and reverse primers (10 pM). Purification and sequencing of PCR products were carried out by Qinke Biotech Co., Yunnan, China.
Table 1. The primers and thermal cycles of PCR used in this study.
Phylogenetic analyses
-
Dissanayake et al.[35] was followed for the phylogenetic analyses. The nucleotide BLAST search in GenBank (
www.ncbi.nlm.nih.gov ) was used to select the most closely related taxa. The sequences of the closely related taxa to the new taxon, which were used in the phylogenetic analyses (Table 2) were obtained from GenBank based on recently published data[19,20]. Multi-gene phylogenetic analyses were carried out based on combined ITS and tub2 genes, including 82 sequences and two outgroup taxa (Xylaria atrosphaerica (Cooke & Massee) Callan & J.D. Rogers (AFTOL-ID 51) and Xylaria hypoxylon (L.) Grev. (CBS 122620)). Multiple alignments of these sequences were automatically made with MAFFT v. 7 at the web server (http://mafft.cbrc.jp/alignment/server ), using default settings[36]. Automatic cutting was done in trimAl.v1.2rev59, and manually combined the multiple sequences in BioEdit v. 7.0.5.2[37]. The multiple sequences were converted using ALTER (www.sing-group.org/ALTER/ )[38].Table 2. Species, strain numbers, and GenBank accession numbers used in the phylogenetic analyses.
Species Strain number ITS tub2 Reference Allocryptovalsa castanea CFCC 52427 MW632944 – [13] Allocryptovalsa castanea CFCC 52428T MW632945 – [13] Allocryptovalsa castanea CFCC 52429 MW632946 – [13] Allocryptovalsa castaneicola CFCC 52432T MW632947 – [13] Allocryptovalsa cryptovalsoidea HVFIG02T HQ692573 HQ692524 [34] Allocryptovalsa cryptovalsoidea HVFIG05 HQ692574 HQ692525 [34] Allocryptovalsa elaeidis MFLUCC 15-0707T MN308410 MN340296 [46] Allocryptovalsa polyspora MFLUCC 17-0364T MF959500 MG334556 [17] Allocryptovalsa rabenhorstii WA07CO HQ692620 HQ692522 [34] Allocryptovalsa rabenhorstii WA08CB HQ692619 HQ692523 [34] Allocryptovalsa sichuanensis HKAS 107017T MW240633 MW775592 [20] Allocryptovalsa truncata PUFNI 17639T MK990279 – [1] Allodiatrype arengae MFLUCC 15-0713T MN308411 MN340297 [46] Allodiatrype elaeidicola MFLUCC 15-0737aT MN308415 MN340299 [46] Anthostoma decipiens JL567 JN975370 JN975407 [47] Anthostoma decipiens IPV-FW349 AM399021 AM920693 Unpublished Cryptosphaeria eunomia CBS 216.87T KT425230 KT425165 [48] Cryptosphaeria eunomia CBS 223.87 KT425231 KT425166 [48] Cryptovalsa ampelina A001 GQ293901 GQ293972 [49] Cryptovalsa ampelina DRO101 GQ293902 GQ293982 [49] Diatrypasimilis australiensis ATCC MYA-3540T NR111369 – [50] Diatrype disciformis GNA14 KR605644 – [23] Diatrype disciformis MFLU 17-1549 MW240629 – [20] Diatrypella elaeidis MFLUCC 15-0279 MN308417 MN340300 [46] Diatrypella verruciformis UCROK1467 JX144793 JX174093 [51] Eutypa lata RGA01 HQ692614 HQ692497 [34] Eutypa lata EP18 HQ692611 HQ692501 [34] Eutypa microasca BAFC: 51550T KF964566 KF964572 [52] Eutypa microasca BAFC: 51551 KF964565 KF964570 [52] Eutypa microasca BAFC: 51556 KF964567 KF964571 [52] Eutypella cerviculata M68 JF340269 – [53] Eutypella semicircularis MP4669 JQ517314 – [54] Halocryptovalsa salicorniae MFLUCC 15-0185T MH304410 MH370274 [25] Halodiatrype avicenniae MFLUCC 15-0953 KX573916 KX573931 [55] Halodiatrype salinicola MFLUCC 15-1277 KX573915 KX573932 [55] Monosporascus cannonballus CMM3646 JX971617 – [56] Monosporascus cannonballus ATCC 26931T FJ430598 – Unpublished Neoeutypella baoshanensis HMAS:255436 MH822887 MH822888 [24] Neoeutypella baoshanensis GMBC0052 MW797106 MW814878 [57] Pedumispora rhizophorae BCC44877 KJ888853 – [58] Pedumispora rhizophorae BCC44878 KJ888854 – [58] Peroneutypa alsophila EL58C AJ302467 – [15] Peroneutypa aquilariae KUNCC 22-10817T OP454038 OP572195 This study Peroneutypa aquilariae KUNCC 22-10818 OP482262 OP572196 This study Peroneutypa comosa BAFC:393 KF964568 – [52] Peroneutypa curvispora HUEFS 136877 KM396641 – [11] Peroneutypa diminutiasca MFLUCC 17-2144T MG873479 MH316765 [18] Peroneutypa diminutispora HUEFS 192196T KM396647 – [11] Peroneutypa indica NFCCI-4393 MN061368 MN431498 [25] Peroneutypa kochiana EL53M AJ302462 – [15] Peroneutypa kunmingensis KUMCC 21-0001T MZ475070 MZ490589 [19] Peroneutypa leucaenae MFLU 18-0816T MW240631 MW775591 [20] Peroneutypa longiasca MFLUCC 17-0371T MF959502 MG334558 [17] Peroneutypa mackenziei MFLUCC 16-0072T KY283083 KY706363 [12] Peroneutypa mangrovei PUFD526T MG844286 MH094409 [24] Peroneutypa polysporae NFCCI-4392 MN061367 MN431497 [25] Peroneutypa scoparia MFLUCC 11-0615 KU940152 – [59] Peroneutypa scoparia MFLUCC 17-2143 MG873478 MH316764 [18] Quaternaria quaternata EL60C AJ302469 – [15] Quaternaria quaternata GNF13 KR605645 – [23] Xylaria atrosphaerica AFTOL-ID 51 DQ491487 – Unpublished Xylaria hypoxylon CBS 122620 AM993141 KX271279 [60] The newly generated sequences are in black bold, superscripted 'T' indicates ex-type, and '–' indicates sequence unavailable. Randomized Accelerated Maximum Likelihood (RAxML) and Bayesian inference analyses (BI) were carried out in the CIPRES Science Gateway (
www.phylo.org/portal2/login!input.action )[39]. The ML trees are performed via RAxML-HPC2 on XSEDE (8.2.12)[40,41] with GTR+I+G model of evolution and bootstrap supports were obtained by running 1000 pseudoreplicates. The BI tree was conducted by Markov Chain Monte Carlo (MCMC) in MrBayes (v3.2.a)[42] to evaluate posterior probabilities[43,44], and the best model was GTR+I+G. Six simultaneous Markov chains were run for one million generations, and trees were sampled every 100th generation. The first 20% of generated trees representing the burn-in phase of the analyses were discarded and the rest were used to calculate posterior probabilities (PP). The phylogenetic tree was visualized with FigTree v1.4.2 (http://tree.bio.ed.ac.uk/software/figtree/ )[45], and bootstrap values were shown at the nodes, and edited by Microsoft Office PowerPoint 2010. The newly obtained alignments and phylogenetic trees were deposited in TreeBASE (https://treebase.org/treebase-web/user/submissionList.html , submission ID: 29758). -
The phylogenetic trees obtained from RAxML and BI gave similar topologies. The RAxML analyses of the combined dataset yielded the best scoring tree (Fig. 1), with a final ML optimization likelihood value of –20,817.822757. The matrix had 1,187 distinct alignment patterns, with 54.28% of undetermined characters or gaps. Parameters for the GTR+I+G model of the combined ITS and tub2 were as follows: estimated base frequencies A = 0.219035, C = 0.277925, G = 0.235904, T = 0.267137; substitution rates AC = 0.957294, AG = 2.875562, AT = 1.254222, CG = 0.889850, CT = 3.867387, GT = 1.000000; proportion of invariable sites I = 0.174656; and gamma distribution shape parameter α = 0.580597.

Figure 1.
RAxML tree of Diatrypaceae based on a combined dataset of ITS and tub2 partial sequences. Bootstrap support values for maximum likelihood (ML) equal to or higher than 50% and bayesian posterior probability (BYPP) equal to or higher than 0.90 are indicated above the branches. Newly generated sequences are shown in red, while the type species are shown in bold black and red.
The final RAxML tree is shown in Fig. 1, which shows that Peroneutypa is a monophyletic group in Diatrypaceae, and Peroneutypa aquilariae is well separated from other strains in Peroneutypa. Peroneutypa mackenziei Q.J. Shang, Phook. & K.D. Hyde (MFLUCC 16-0072) is phylogenetically closely related to P. aquilariae. In addition, strains of Eutypa microasca E. Grassi & Carmarán are grouped within Peroneutypa (Fig. 1).
Taxonomy
-
Peroneutypa aquilariae T.Y. Du & Tibpromma, sp. nov. Fig. 2
MycoBank number: MB845438; Facesoffungi number: FoF12744
Etymology: named after the host genus, Aquilaria.
Saprobic on dead twigs of Aquilaria sinensis (Thymelaeaceae). Sexual morph: Ascostromata 0.5–1.5 mm wide, well-developed interior, solitary to gregarious, mostly solitary, immersed, long ostiolar canal raised through host tissue, black, irregular in shape, arranged irregularly, 1–6 locules. Ascomata (excluding necks) 300–570 μm diam., perithecial, immersed in ascostromata, subglobose to globose, dark brown to black. Ostiolar canal 20–40 μm wide, without periphysis, filled with hyaline cells, with 200–300 μm long, cylindrical, straight, dark-brown to black necks. Peridium 35–70 μm wide, composed of outer layer thick-walled, dark brown to pale brown cells of textura angularis, and inner layer thin-walled, hyaline cells of textura prismatica. Paraphyses absent. Asci 15–20 × 5–7 µm (
$\bar{\text {x}} $ $\bar{\text {x}} $ Culture characteristics: Ascospores germinated on PDA within 24 h at a constant temperature incubator (28 °C). Colonies on PDA reaching 6 cm diam., after one week at 28 °C, mycelium white, flossy, circular with entire edge, with filiform margin. After one month, mycelium becomes white to light yellow from above and light brown to brown from below.
Material examined: China, Yunnan Province, Xishuangbanna, on dead twigs of Aquilaria sinensis (Thymelaeaceae), 15 September 2021, Tianye Du, YNA3 (holotype, HKAS 124185; ex-type cultures, KUNCC 22-10817 = KUNCC 22-10818).
Notes: The BLASTn search of ITS sequences of our strains is 87.67% similar to P. mackenziei (MFLUCC 16-0072, NR_154363). In the present phylogenetic analyses, our strains formed a sister branch with P. mackenziei (MFLUCC 16-0072) with a low bootstrap support (63% ML). However, they differ in morphological characteristics i.e. stromata of P. aquilariae have well-developed interior, ostiolar canal without periphyse, peridium inner layer comprises of 3–5 hyaline cell layers of textura prismatica, and paraphyses absent; while P. mackenziei has poorly developed interior stromata, ostiolar canal periphysate, peridium inner layer comprises 8–10 hyaline cell layers of textura angularis, and hamathecium is composed of paraphyses[12]. Furthermore, a comparison of ITS nucleotides between P. aquilariae and P. mackenziei (MFLUCC 16-0072) resulted in 13.8% differences (67/487 bp, without gaps), and 54.1% differences (173/320 bp, without gaps) respectively in tub2. Based on both morphological characteristics and multigene phylogenetic analyses results, we introduce Peroneutypa aquilariae as a distinct new species and this is the first report of Peroneutypa from Aquilaria sinensis. We also provide information about countries, hosts, and a comparison of morphological characteristics of Peroneutypa in Table 3.
Table 3. Morphological characteristics, host, and location information of Peroneutypa species.
Species Ascostromata Ascomata Ostiolar canal Peridium Paraphyses Asci Ascospores Countries Hosts References P. alsophila Slightly or well-developed, less than 1 mm long N/A Necks not prominent N/A N/A Apical apparatus J+ N/A Argelia, France Arthrocnemum fruticosum [15,16] P. aquilariae Slightly or well-developed,
0.5–1.5 mm wide,
1–6 locules300–570 μm (excluding necks) With 200–300 μm long necks, 20–40 μm wide, without periphysis 35–70 μm wide Absent 18.3 × 5.8 μm, short pedicellate or non-pedicellate, J- apical ring, apically rounded to truncate 6.3 × 2 μm,
hyaline to pale
yellow, oblong
to allantoidChina Aquilaria sinensis This study P. bellula Immersed, scattered Globose Long necks N/A N/A Minimum, 5–6-spored, hyaline Oblong, with granules N/A Arundo donax [61] P. coffeae 450–900 × 350–650 μm N/A 700–1,500 × 80–100 μm 20–25 μm wide N/A 23–25 × 4–5 μm, pedicellate 4.5–6 × 1–1.5 μm, allantoid Central African Republic Coffea robusta [62] P. comosa Slightly or well-developed N/A Necks very long, over 1 mm, prominent N/A N/A Apical apparatus J+ / J− N/A Argentina Celtis tala [16,52] P. corniculata Multi-ascoma N/A Prominent N/A N/A N/A N/A America Prunus melanocarpa [9,63] P. curvispora Poorly-developed, 0.6–3 mm wide,
22 ascomata per stroma300–700 μm wide 400–800 μm long, prominent N/A Absent 9–16.5 × 4–6 μm, long pedicellate, apex truncate 3–5 × 1–2 μm, allantoid,
strongly curvedBrazil Unidentified wood [11] P. cylindrica 3–4 mm,
8–10 locules1 mm 1–2.5 mm long N/A Absent 10–20 × 4–5, clavate, pedicellate 4–5 × 2, hyaline, curved South Africa Unidentified wood [64] P. cyphelioides 1–3 mm 300 μm 1–2 mm long N/A N/A 15 × 4–5 μm 4 × 1 μm, allantoid Philippines Streblus asper [65] P. diminutiasca With poorly-developed interior, 1.2–1.4 mm wide, 1–10 locules 147–218 μm (excluding necks) 193 × 48 μm wide, long neck, periphysate 15–32 μm wide 4–7 μm wide, dense, septate 22 × 4 μm, long pedicellate, with a J− subapical ring, apically rounded to truncate 4.2 × 1.7 μm, hyaline to pale
yellowish, allantoidThailand Unidentified wood [18] P. diminutispora Absent or with poorly-developed, isolated or in groups of up to seven 400–700 μm 200–400 μm long N/A Absent Long pedicellate, with a J− apical rings, apex truncate Allantoid, slightly to moderately curved, yellowish in mass Brazil Unidentified wood [11] P. discriminis 1 mm 200 μm 1 mm long N/A N/A 12–14 × 4 μm 5–6 × 1.5–2 μm India, Philippines Ficus racemosa, Macaranga tanarius, Streblus asper [65] P. exigua N/A N/A 500–700 μm long N/A N/A 11–16 × 3–6 μm (sporiferous part) 3–3.5 × 1–1.5 μm Brazil Citrus aurantium [66] P. gliricidiae Interior of the stroma not developed N/A Necks not prominent N/A N/A 10–16 µm, J+ Allantoid Philippines Gliricidia sepium [16] P. heteracanthoides N/A 300 μm N/A N/A Absent 16 × 3.5 μm 3–4 × 1 μm, allantoid, hyaline, with granules Singapore Cassiae sp., Heveae brasiliensis [67] P. indica Well-developed 375 × 202 µm 100–250 µm wide, with moderate neck 100–350 µm long, periphysate 15–35 µm wide 1–2 µm 42 × 3.5 µm,
short pedicellate, apically rounded to truncate, with a J– apical ring5.5 × 1.3 µm, hyaline, straight to allantoid, light brown in mass India Suaeda monoica [25] P. iranica 0.5–2 mm, 3–16 locules 300–500 μm 40–120 μm, periphysate 30–55 μm wide Septate, hyaline 13.5 × 3.6 μm, with a J+ apical ring 3.6 × 1 μm, hyaline, allantoid Iran Wisteria sinensis [68] P. kochiana Interior of the stroma not developed N/A Necks not prominent N/A N/A 18–28 µm long, J+ Allantoid Russia, Spain Atriplex halimus [15,16] P. komonoensis N/A 250–450 × 200–350 μm 1–1.2 mm long N/A N/A 30–38 × 3.5–4 μm, long pedicellate 3–4 × 1–1.5 μm, allantoid French Equatorial Africa Hevea brasiliensis [69] P. kunmingensis Absent or with poorly-developed interior 260 × 317 μm (excluding neck) 140 × 100 μm wide, long neck, periphysate 30–60 μm wide Septate, hyaline 26 × 4.1 μm, straight or slight curved, hyaline, long pedicel up to 16 μm, with a J– apical ring 4.2 × 1.3 μm, hyaline to pale grey, oblong to allantoid China Unidentified wood [19] P. leucaenae N/A 655 × 525 μm 275–350 μm long neck, periphysate 22–43 μm wide 3.2–7 μm wide, wider at the base, long, septate, 33 × 4.2 μm, pedicel 17–27 μm long, J+ apical ring Yellowish-brown, ellipsoidal to
cylindrical or elongate-allantoidThailand Leucaena leucocephala [20] P. lignicola 609–685 μm high, 1,196–1,246 μm, 4–6 locules 448–662 × 355–445 μm Long neck 62–92 μm wide N/A 16 × 6 μm, sessile, hyaline 6 × 2 μm, elongate-allantoid, straight to slightly curved Thailand On decaying wood submerged in a freshwater stream [28] P. longiasca N/A 180–450 × 170–390 µm 20–50 μm wide, 190–440 μm long 14–47 μm wide N/A Short to long pedicellate, apically rounded to truncate with indistinct J– subapical ring 5.8 × 2 μm, hyaline, oblong to allantoid Thailand Hevea brasiliensis [17] P. mackenziei With poorly-developed interior, 1.2–2.2 mm wide, multi-ascoma 466 × 356 μm 265 × 100 μm, ostiolar canal with long neck, periphysate 45–65 μm wide 2.5–4.5 μm wide, dense, filamentous, aseptate, hyaline 17.7 × 4.2 μm, sessile, apically rounded to truncate, with a J– apical ring 5.6 × 1.6 μm, subhyaline to
pale yellowish,
elongate allantoidThailand Unidentified wood [12] P. macroceras 600–1,500 × 500–650 μm N/A 1,500–2,500 × 100–120 μm N/A N/A 30–35 × 4–4.5 μm, long pedicellate 4–5 × 1.2–1.5 μm, allantoid Central African Republic Coffea robusta [62] P. macrorostrata 1,500–2,000 × 350–500 μm N/A N/A N/A N/A 25–30 × 4–5 μm, pedicellate 4–6 × 1.5–2 μm, allantoid French Equatorial Africa Hevea brasiliensis [69] P. mangrovei Absent or poorly-developed, up to four in groups 375 × 202 μm 50–85 μm wide, with moderate neck length 100–350 μm, periphysate 15–35 µm wide 1–2 μm wide, septate, longer than asci 17 × 3.5 μm, short pedicellate, apically rounded to truncate, with a J– apical ring Hyaline to pale yellow, straight to allantoid India Avicennia marina [24] P. multistromata 400–900 × 200–700 μm 270–450 × 200–300 μm N/A N/A Absent 30–40 × 3–5 μm, (sporiferous part: 14–18 μm), long pedicellate, 8-spored 3–4 × 0.75–1 μm, hyaline, allantoid Cote d'Ivoire Coffea canephora [70] P. obesa Slightly or well-developed, spiny or bristly appearance to the stromatic surface, black line present N/A Necks prominent N/A N/A N/A 3–7 µm long, strongly curved Argentina, Zaire Unidentified
wood[16] P. perseae 500–700 × 300–500 μm,
5–6 locules300–600 × 150–200 μm N/A N/A Absent 18–26 × 4–6.5 μm, 8-spored, hyaline 5–6 × 1.5–2 μm, hyaline Morocco Persea americana [71] P. philippinarum 2–3 locules 200 μm 300–1000 μm long N/A N/A 15 × 4–5 μm 4 × 1.5 μm, allantoid Philippines Streblus asper [65] P. polymorpha N/A N/A N/A N/A N/A N/A N/A Cote d'Ivoire Manihot esculenta [72] P. polysporae N/A 286 × 223 µm 113 × 60 µm, periphysate 15–50 µm wide Septate, hyaline 113 × 12 µm, polysporous 9 × 1.8 µm, hyaline to pale yellow, allantoid India Suaeda monoica [25] P. rubiformis With poorly-developed interior, 1–1.4 mm wide, 1–10 locules 109 × 159 μm (excluding necks) 176 × 52 μm, roundish, prominent ostioles in the centre, rubus-like or star-like in shape, periphysate 20–25 μm wide 7–11 μm wide, septate, hyaline 50 × 5 μm, short pedicellate, apically rounded to truncate, with a J- subapical ring 4.3 × 1.6 μm, hyaline to pale yellowish, elongate-allantoid Thailand Unidentified wood [18] P. scoparia Not developed or with poorly-developed interior,1.2–2 mm wide, 4–8 locules 256 × 266 μm 441 × 105 μm,
long necks,
prominent, periphysate20–35 μm wide Septate, hyaline 22 × 4.5 μm, long pedicellate, apically rounded to truncate, with a J– subapical ring 4.1 × 1.5 μm, oblong to allantoid, sometimes slightly curved Argentina, Chile, Czech Republic, France, India, Iran, Italy, Paraguay, Sweden, Thailand Acer pseudoplatanus, Actinidia deliciosa, Avicennia marina, bamboo culms, Gleditsia sp., Juglans regia, Robinia pseudoacacia, Vaccinium corymbosum [16,18,25,59] P. variabilis N/A 400–500 μm 700–2500 × 100–200 μm N/A N/A 12–18 × 3 μm (sporiferous part), tapering pedicel 3–5 × 1.5–2 μm, greenish-hyaline, cylindric, curved Sri Lanka Unidentified herbaccous [73] Host and location information of taxa extracted from the listed references and Farr & Rossman[74]. The symbol “N/A" denotes no information available or not mentioned in given papers, and the new species is in black bold. 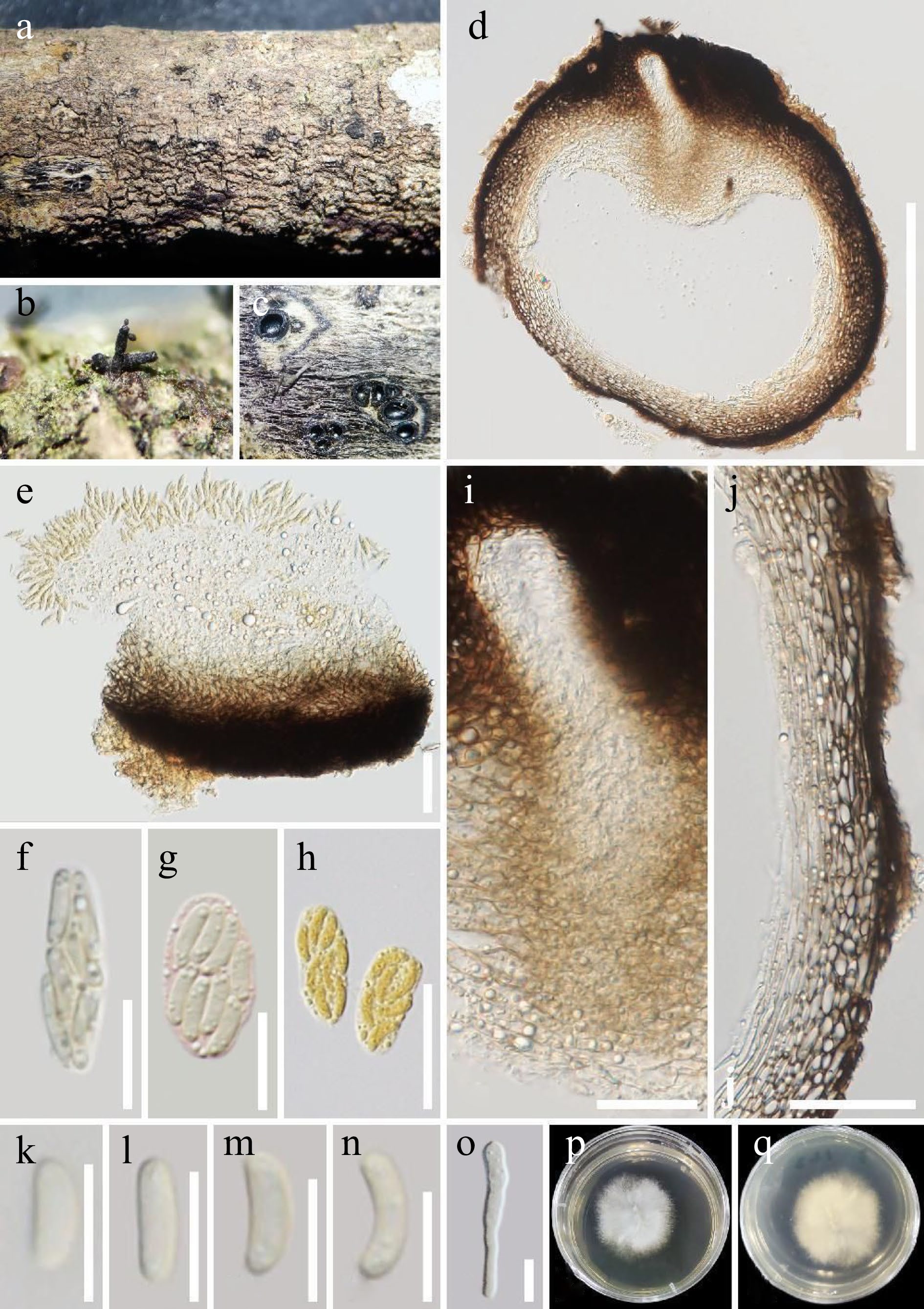
Figure 2.
Peroneutypa aquilariae (HKAS 124185, holotype). (a)–(c) Appearance of ascomata on the substrate. (d) Section through an ascoma. (e)–(h) Asci, (g) ascus stained with Congo red reagent, (h) asci stained with Melzer’s reagent). (i) Ostiole. (j) Peridium. (k)–(n) Ascospores. (o) A germinating ascospore. (p), (q) Colony on PDA medium (after 7 d in culture). Scale bars: (d) = 300 μm, (e), (j) = 50 μm, (f), (g) = 10 μm, (h) = 20 μm, (i) = 30 μm, (k)–(o) = 5 μm.
-
The updated phylogenetic tree of this study shows that Peroneutypa is a monophyletic genus, with E. microasca strains clustered within Peroneutypa, and this also is consistent with the previous research results of de Almeida et al.[11], Senwanna et al.[17], and Phukhamsakda et al.[19]. Senwanna et al.[17] considered that E. microasca should be listed under Peroneutypa. Though we agree with them, fresh specimens of E. microasca need to be recollected in the future and transferred to Peroneutypa based on the evidence of morphological comparison and phylogenetic analyses.
Previous studies have reported that it is difficult to distinguish the members of Diatrypaceae by morphology, and many taxa still lack molecular data, which undoubtedly make the classification of Diatrypaceae more difficult[11,17,18]. There are similar difficulties in Peroneutypa. At present, 35 species of Peroneutypa are listed, and only 16 of them have molecular data (Tables 2 & 3). In particular, P. lignicola Z.L. Luo, K.D. Hyde & H.Y. Su (MFLUCC 14-0040) was not included in the phylogenetic analyses of this study as this species has no ITS and tub2 gene sequences, and therefore, more fresh samples need to be collected in the future in order to supplement the known species with molecular data.
In Peroneutypa, currently, only ten species have tub2 gene sequences. The nucleotide fragments were compared and the results showed that the tub2 sequences of the ten species were quite different. Even among different strains (MFLUCC 17-2143 and NFCCI-4396) of the same species P. scoparia, 9% nucleotide differences (31/344 bp, without gaps) were observed. This may lead to the long branches of most species of this genus in the phylogenetic trees. The reason for the large difference in tub2 sequence might be due to the fact that different primers were selected in different strains, viz. using primers Bt2a/Bt2b: P. indica Devadatha, V.V. Sarma & E.B.G. Jones (NFCCI-4393), P. mackenziei (MFLUCC 16-0072), P. mangrovei Devadatha & V.V. Sarma (PUFD526), P. polysporae (NFCCI-4392), and P. scoparia (MFLUCC 17-2143); using primers T1/Bt2b: P. diminutiasca Q.J. Shang, Phukhamsak & K.D. Hyde (MFLUCC 17-2144), P. kunmingensis (KUMCC 21-0001), P. longiasca Senwanna, Phookamsak & K.D. Hyde (MFLUCC 17-0371), P. rubiformis Q.J. Shang, Phukhamsak & K.D. Hyde (MFLUCC 17-2142), and P. scoparia (NFCCI-4396); and using primers T1/T22: P. leucaenae (MFLU 18-0816). On the other hand, we suppose the difference of tub2 might be the unique characteristics of this genus, thus the tub2 gene can be used as a good marker in phylogeny. However, this study suggests that researchers working on this genus should use the same tub2 primers for PCR amplification.
In this study, no asexual morph of the new taxon was observed, but in Shang et al.[18], the asexual types of P. rubiformis and P. scoparia were made in photo plates and descriptions, with the main characteristics of pycnidial conidiomata, cylindrical conidiogenous cells, with holoblastic and hyaline conidia lunate to filiform, or fusiform, curved at the upper part, flattened at the base and blunt apex. The discovery of the asexual morph will be useful in the taxonomic identification of Peroneutypa in the future.
Our new species is associated with the genus Aquilaria, which belongs to Thymelaeaceae and is famous for producing agarwood[32,75,76]. Agarwood is very expensive and has a wide range of uses worldwide[76]. At present, the production mechanism of agarwood is not clear, but fungi are considered to be involved in producing agarwood[77,78]. Therefore, it is necessary to study the fungi associated with agarwood, including the saprophytic fungi. Saprophytic fungi associated with Aquilaria are poorly studied and this study reports the first Peroneutypa species from an Aquilaria host. In the future, more fresh samples need to be collected for in-depth research on the fungal diversity of Aquilaria.
This research was supported by the National Natural Science Foundation of China (No. NSFC 31760013, 31950410558, 32260004), and High-Level Talent Recruitment Plan of Yunnan Provinces ('Young Talents' Program). Tian-Ye Du thanks Mae Fah Luang University for the award of fee-less scholarship. Ausana Mapook thanks the Mae Fah Luang University Fund (Grant No. 651A16029), entitled 'Taxonomy, phylogeny, risk assessment, and potential impact of fungi on Siam weed in northern Thailand'. Shaun Pennycook is thanked for his assistance in selecting a species epithet for the new species.
-
The authors declare that they have no conflict of interest.
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Du TY, Karunarathna SC, Zhang X, Dai DQ, Gao Y, et al. 2022. Morphology and multigene phylogeny revealed Peroneutypa aquilariae sp. nov. (Diatrypaceae, Xylariales) from Aquilaria sinensis in Yunnan Province, China. Studies in Fungi 7:18 doi: 10.48130/SIF-2022-0018 

Morphology and multigene phylogeny revealed Peroneutypa aquilariae sp. nov. (Diatrypaceae, Xylariales) from Aquilaria sinensis in Yunnan Province, China
- Received: 28 November 2022
- Accepted: 30 November 2022
- Published online: 22 December 2022
Abstract: Dead twigs of Aquilaria sinensis (Thymelaeaceae) with fungal fruiting bodies were collected from Xishuangbanna, Yunnan Province, China. After initial morphological observations, an interesting fungus whose morphologically resembled Peroneutypa was isolated. Molecular phylogeny of combined ITS and tub2 showed our fungal collection
-
Key words:
- Ascomycota /
- Fungi /
- New taxon /
- Saprophyte /
- Sordariomycetes /
- Taxonomy