








-
In coastal ecosystems, salt marsh habitats are common and consist of diverse halophytes, grasses, herbs, and shrubs, as well as microorganisms [ 1, 2] . These species-rich ecosystems are highly productive, and investigating fungal diversity in these habitats is important while many areas are still being explored [ 2, 3] . Researchers are currently studying the taxonomy of fungi in marine and semi-marine environments and increasing the number of known taxa recorded [ 4− 6] . Ascomycota was identified as the dominant group in world salt marsh ecosystems, including the highest diversity in Pleosporales, Dothideomycetes [ 2] . Coniothyriaceae is a pleosporalean family with a large number of terrestrial taxa, while taxa associated with salt marsh vegetation have rarely been reported [ 2] .
Coniothyriaceae was established by Cooke [ 7] to accommodate Coniothyrium species. The type genus and species of Coniothyriaceae are Coniothyrium Corda and C. palmarum Corda, respectively [ 8− 10] . This family was previously linked to Leptosphaeriaceae [ 11] and this was followed by several authors [ 12− 16] . Subsequently, molecular data analyses for phoma-like asexual morphs were performed by de Gruyter et al. [ 17] based on LSU and ITS sequence data and revealed that C. palmarum is phylogenetically distant from Leptosphaeriaceae and closely related to Coniothyriaceae. de Gruyter et al. [ 17] reinstated Coniothyriaceae as a distinct family in Pleosporales and transferred several Phoma and Pyrenochaeta species into this family. Thus, the morphological variations of Coniothyrium species were expanded by the addition of more characters, such as setose pycnidia and conidiogenesis with elongated conidiophores [ 17] . Several authors later updated the placements of many Coniothyrium species with generic level changes and novel genera placed in different families [ 18, 19] .
Verkley et al. [ 19] studied morphology and phylogenetic relationships of coniothyrium-like and closely related taxa. These species are coelomycetous and characterized in having pycnidial or stromatic conidiomata and small, subhyaline to pigmented, 1- or 2-celled conidia [ 19] . The phenotypic plasticity of these coelomycetous species has made their taxonomic placements uncertain and, thus the majority of them have been placed in Coniothyrium [ 19− 21] . Both Coniothyrium and coniothyrium-like species were identified as polyphyletic within Pleosporales and recent taxonomic treatments were mainly treated with combined morphology and molecular data analyses [ 12, 16, 17, 19, 21− 32] . Coniothyrium sensu stricto is characterized by 1-septate conidia and grouping in Coniothyriaceae [ 17, 19− 21, 33] . Currently, Coniothyriaceae consists of five genera, such as Coniothyrium, Foliophoma Crous , Neoconiothyrium Crous, Ochrocladosporium Crous & U. Braun and Staurosphaeria Rabenh. (≡ Hazslinszkyomyces Crous & R.K. Schumach.) [ 10, 34] .
Coniothyriaceae members have been identified as pathogens that cause necrotrophic and leaf spots on leaves, and saprobes on dead branches [ 10, 17] . The sexual morph is characterized in having cucurbitaria-like, black, globose ascomata, short central ostiole, textura angularis peridium cells, branched, septate, cellular pseudoparaphyses, 8-spored, cylindrical, bitunicate asci and muriform, ellipsoidal ascospores that are initially hyaline and brown at maturity [ 10] . Asexual morphs are coelomycetous and sometimes differentiated with phoma-like, camarosporium-like, coniothyrium-like, or cladosporium-like asexual characters. They are characterized in having dark, globose, pycnidial conidiomata, with central, sometimes papillate ostiole, cells of textura angularis or textura globulosa in the conidiomatal wall, hyaline macroconidiogenous and microconidial cells and conidia. Conidial morphology is varied as macroconidia and microconidia. Macroconidia are ellipsoid, red-brown, and septation is from the central transverse septum to muriformly septate, while microconidia are hyaline, globose to ellipsoid and aseptate [ 9, 10, 24, 27, 35, 36] .
The aim of the study
-
In this study, we aim to expand the taxonomy of fungi associated with dead plant hosts in salt marsh ecosystems. We investigate salt marsh habitats in Thailand to collect fungal specimens and isolate them to find out the taxonomic novelties. Morphological illustrations, comprehensive descriptions, and multi-gene phylogenetic analyses are provided to confirm the placement of new findings.
-
Fungal specimens were collected from salt marsh habitats in Pranburi Province, Thailand, 2021. Samples were preserved in sterile Ziploc bags in the laboratory and incubated at room temperature 25 °C. Rehydrated specimens were observed to identify fungal fruiting bodies and macro-morphology was observed by using a Motic SMZ 168 compound stereomicroscope. Micro-morphologies (e.g., conidiomata, conidiogenous cells, conidia) were examined from hand-sectioned structures using a Nikon ECLIPSE 80i compound stereomicroscope, equipped with a Canon 600D digital camera. The measurements of photomicrographs were obtained using Tarosoft (R) Image Frame Work version 0.9.7. Images were edited with Adobe Photoshop CS6 Extended version 13.0.1 software (Adobe Systems, San Jose, California, USA).
Single-spore isolation was carried out as described by Senanayake et al. [ 37] . Germinated spores were aseptically transferred into fresh malt extract agar medium (MEA) prepared in 50% or 100% concentrations of sterilized natural seawater [ 38] . Culture plates were incubated at 25 °C for six weeks and inspected every week. Herbarium specimens are preserved at Mae Fah Luang University Herbarium (MFLU) in Chiang Rai, Thailand. All living cultures are deposited at Mae Fah Luang Culture Collection (MFLUCC). Facesoffungi and Index Fungorum numbers for new taxa were obtained [ 39, 40] .
DNA extraction, PCR amplification, sequencing
-
The methodologies for DNA extraction, PCR, gel electrophoresis, and sequencing were followed, as detailed in Dissanayake et al. [ 41] . The genomic DNA was extracted from fresh mycelium using the E.Z.N.A Fungal DNA Mini Kit- D3390-02 (Omega Bio-Tek, USA) following the guidelines of the manufacturer. DNA sequences were obtained for the internal transcribed spacer region (ITS1, 5.8S, ITS2), the small subunit (SSU), and the large subunit (LSU) of the nuclear ribosomal RNA gene. PCR thermal cycle programs for each locus region are presented in Table 1. Purification and sequencing were outsourced to the Bio Genomed Co. LTD laboratory (Biogenomed Co., Thailand). Newly generated sequences were submitted to NCBI GenBank (
www.ncbi.nlm.nih.gov/genbank ).Table 1. Gene regions, primers, and PCR thermal cycle programs used in this study, with corresponding reference(s).
Genes/loci PCR primers (forward/reverse) PCR conditions Reference (s) ITS and LSU ITS5/ITS4 and LR0R/LR5 94 °C; 2 min (95 °C; 30 s, 55 °C; 50 s, 72 °C; 90 s) × 35 thermal cycles, 72 °C; 10 min [ 42− 44] SSU NS1/NS4 95 °C; 3 min (95 °C; 30 s, 55 °C; 50 s, 72 °C; 30 s) × 35 thermal cycles, 72 °C; 10 min [ 42] BioEdit v 7.0.9.0 program [ 45] was used to check the quality of the newly generated sequence chromatograms. For primary identification, contig sequences were checked with BLAST searches in NCBI. Sequences for phylogenetic analyses were downloaded from GenBank ( Table 2) following Hyde et al. [ 25] . Each gene matrix was aligned with MAFFT version 7 [ 46] with default parameters and manually adjusted for improvement where necessary using BioEdit v. 7.2 [ 45] . The trimAl v1.4 software was used for the automated removal of spurious sequences or poorly aligned regions in each single gene alignment, and gappyout was selected as the automated trimming method [ 47] . Two separate phylogenetic analyses were conducted: Maximum Likelihood (ML) and Bayesian Inference (BI). LSU, SSU, and ITS concatenated dataset was analyzed for Coniothyriaceae and selected families in Pleosporales.
Table 2. Taxa used in the phylogenetic analyses and their GenBank accession numbers. Sequences of new taxon generated in this study are in blue-bold and type strains are in black-bold.
Species Strain/voucher number GenBank accession numbers ITS LSU SSU Amarenographium ammophilae MFLUCC 16–0296 KU848196 KU848197 KU848198 Ascochyta pisi CBS 126.54 GU237772 EU754137 EU754038 Bipolaris microstegii CBS 132550 NR_120160 NG_042690 NA Bipolaris victoriae CBS 327.64 NR_147489 NG_069233 NA Comoclathris arrhenatheri MFLUCC 15–0465 NR_165855 NG_068240 NG_068374 Coniothyrioides thailandica MFLUCC 22-0193 OQ023276 OQ023277 OQ025050 Coniothyrium carteri LG1401 MS6E KX359604 KX359604 NA Coniothyrium cereale CBS 157.78 MH861123 JX681080 NA Coniothyrium chiangmaiense MFLUCC 16–0891 KY568987 KY550384 KY550385 Coniothyrium dolichi CBS 124140 JF740183 GQ387611 GQ387550 Coniothyrium glycines CBS 124455 JF740184 GQ387597 GQ387536 Coniothyrium palmarum CBS 400.71 AY720708 EU754153 AY720712 Coniothyrium palmarum CBS 758.73 NA JX681085 EU754055 Coniothyrium sp. B9-10-9 MW764153 NA NA Coniothyrium sp. P16-10-4 MW764259 NA NA Coniothyrium telephii CBS 188.71 JF740188 GQ387599 GQ387538 Coniothyrium telephii CBS 856.97 JF740189 GQ387600 GQ387539 Coniothyrium telephii UTHSC:DI16–189 LT796830 LN907332 NA Coniothyrium triseptatum MFLU 19–0758 NR_171948 NG_073674 NA Curvularia heteropogonis CBS 284.91 JN192379 JN600990 NA Didymella azollae A1 MT514913 MT514910 NA Foliophoma camporesii MFLUCC 18–1129 KY929151 KY929181 NA Foliophoma fallens CBS 161.78 KY929147 GU238074 GU238215 Foliophoma fallens CBS 284.70 KY929148 GU238078 GU238218 Libertasomyces myopori CPC 27354 NR_145200 NG_058241 NA Libertasomyces platani CPC 29609 NR_155336 NG_059744 NA Libertasomyces quercus CBS 134.97 NR_155337 DQ377883 NA Melnikia anthoxanthii MFLUCC 14–1010 NA KU848204 KU848205 Neoconiothyrium hakeae CPC 27616 KY173397 KY173490 NA Neoconiothyrium hakeae CPC 27620 KY173398 KY173491 NA Neoconiothyrium multiporum CBS 353.65 NR_111617 JF740268 NA Neoconiothyrium multiporum CBS 501.91 JF740186 GU238109 GU238225 Neoconiothyrium persooniae CBS 143175 NR_156386 NG_058509 NA Neoconiothyrium viticola CPC 36397 NR_165929 NG_068326 NA Neoplatysporoides aloeicola CPC 24435 NR_154230 NG_058160 NA Ochrocladosporium elatum CBS 146.33 EU040233 EU040233 NA Ochrocladosporium frigidarii CBS 103.81 NR_156512 NG_064123 NA Phaeosphaeria chiangraina MFLUCC 13–0231 KM434270 KM434280 KM434289 Phaeosphaeria musae MFLUCC 11–0133 KM434267 KM434277 KM434287 Phaeosphaeria thysanolaenicola MFLUCC 10–0563 NR_155642 NG_069236 NG_063559 Phaeosphaeria oryzae CBS 110110 NR_156557 NG_069025 NG_061080 Phaeosphaeriopsis dracaenicola MFLUCC 11–0157 NR_155644 NG_059532 KM434292 Pleospora herbarum MFLUCC 14-0920 KY659560 KY659563 KY659567 Querciphoma carteri CBS 101633 JF740180 GQ387593 GQ387532 Querciphoma carteri CBS 105.91 JF740181 GQ387594 GQ387533 Querciphoma carteri Gv5 MT819903 NA NA Querciphoma carteri UASWS2031 MN833930 NA NA Shiraia bambusicola NBRC 30753 AB354987 AB354968 NA Shiraia bambusicola NRBC 30771 AB354990 AB354971 NA Shiraia bambusicola NRBC 30772 AB354991 AB354972 NA Staurosphaeria aloes CBS 136437 KF777142 KF777198 NA Staurosphaeria aloes CPC 21572 NR_137821 NG_067283 NA Staurosphaeria aptrootii CBS 483.95 NR_155186 NA NA Staurosphaeria lycii CPC 30998 KY929150 KY929180 NA Staurosphaeria lycii CPC 31014 KY929151 KY929181 NA Staurosphaeria rhamnicola MFLUCC 17–0813 MF434200 MF434288 MF434376 Staurosphaeria rhamnicola MFLUCC 17–0814 NR_154461 MF434289 NG_063659 Stemphylium vesicarium CBS 191.86 MH861935 MH873624 GU238232 NA: Sequences not available in GenBank. Taxon treatments
Phylogenetic analyses
-
In the phylogenetic analyses, maximum likelihood (ML) was executed using IQ-Tree web server (
http://iqtree.cibiv.univie.ac.at/ ) with bootstrap support obtained from 1,000 pseudoreplicates [ 48, 49] . Bayesian Inference (BI) analysis was performed on the CIPRES Science Gateway portal under MrBayes on XSEDE (3.2.7a) [ 50] . Six simultaneous Markov chains were run for 1,000,000 generations, and trees were sampled every 1,000 th generation, ending the run automatically when the standard deviation of split frequencies dropped below 0.01. The best nucleotide substitution models for each genetic marker were evaluated using jModelTest2 on XSEDE in the online CIPRES Portal (www.phylo.org/portal2 ) [ 51, 52] . The best-fit models under the AIC criterion were revealed to be GTR+I+G for ITS and LSU regions while GTR+I for SSU region. Phylogenetic trees were visualized with FigTree version 1.4.0 [ 53] and edited in Microsoft PowerPoint (2019). -
The combined LSU, SSU, and ITS alignment was used to construct the final phylogenetic analysis ( Fig. 1) of maximum likelihood (ML) and Bayesian inference (BI).
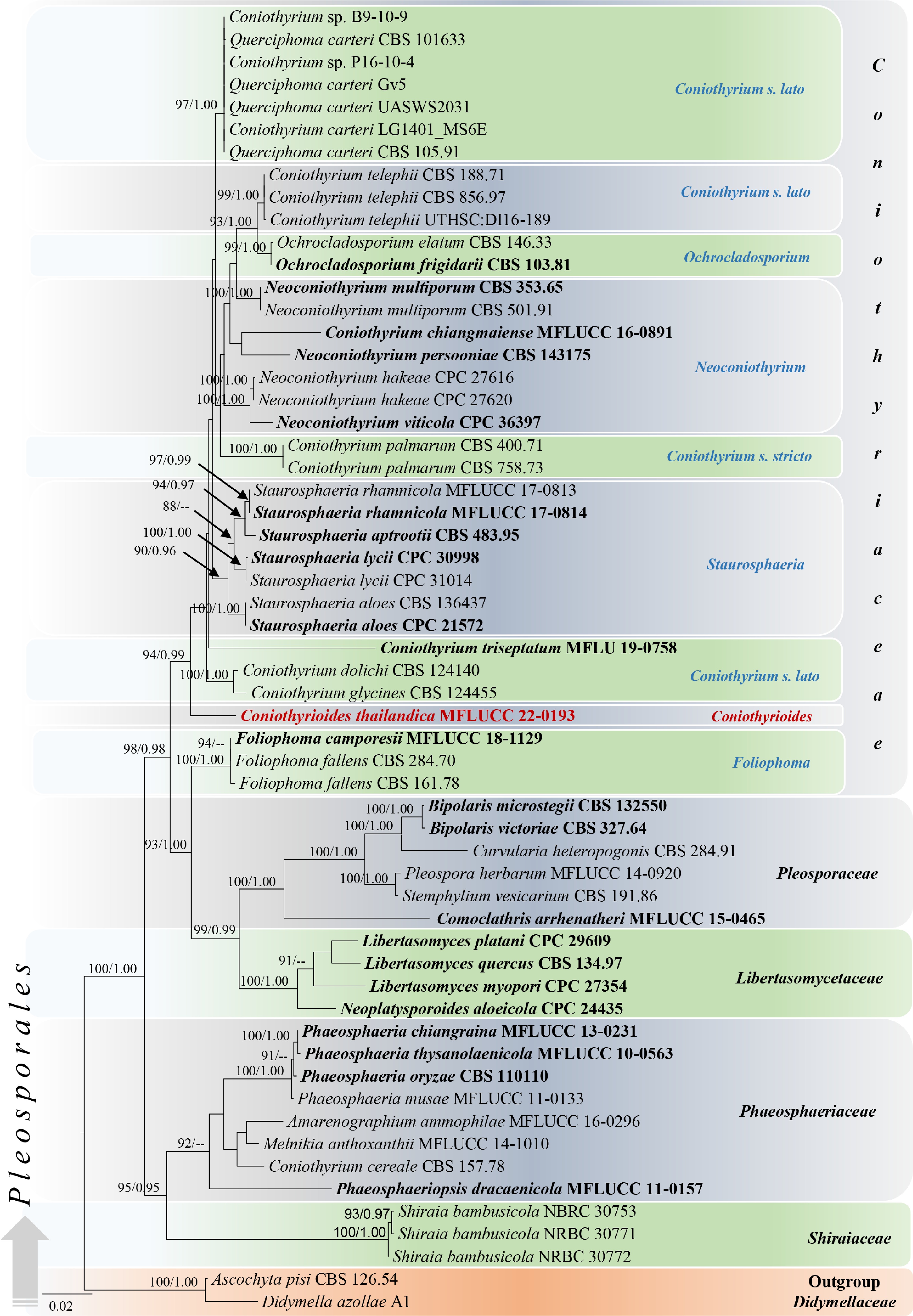
Figure 1.
Phylogram generated from maximum likelihood analysis based on combined LSU, SSU, and ITS sequenced data. Fifty-eight strains were included in the combined sequence analyses, which comprised 2251 characters with gaps (LSU = 800, SSU = 948, ITS = 503). Single gene analyses were also performed, and topology and clade stability were compared from the combined gene analyses. Ascochyta pisi Lib. (CBS 126.54) and Didymella azollae E. Shams, F. Dehghanizadeh, A. Pordel & M. Javan-Nikkhah (A1) were used as the outgroup taxa. The final ML optimization likelihood is -10163.644. The matrix included 494 distinct alignment patterns including undetermined characters. Estimated base frequencies were obtained as follows: A = 0.245, C = 0.219, G = 0.274, T= 0.262; substitution rates AC = 2.73290, AG = 3.93954, AT = 2.73290, CG = 1.0, CT = 7.93321, GT = 1.0 and the gamma distribution shape parameter α = 0.439534. Bootstrap support values for ML (first set) equal to or greater than 75% and BYPP equal to or greater than 0.95 are given above or below the nodes. The strains from the current study are in red bold and the type strains are in black bold. The scale bar represents the expected number of nucleotide substitutions per site.
Morphological analyses
-
Coniothyrioides Wijes., M.S. Calabon, E.B.G. Jones & K.D. Hyde, gen. nov.
Index Fungorum number: 555045; Facesoffungi number: 13901 Fig. 2
Etymology – Resembling Coniothyrium taxa
Saprobic on a submerged decaying wood in salt marsh ecosystems. Sexual morph: Undermined. Asexual morph: Coelomycetous. Forming conspicuous, round to irregular, black pycnidia. Conidiomata semi-immersed, erumpent through the host substrate, globose to subglobose, solitary, scattered to aggregated, uni-loculate, ostiolate, covered in setae, rigid when dehydrated, black. Setae originated from the outermost layers of conidiomatal wall, divergent, brown, with hyaline apex, septate, smooth-walled, uniformly wide from base to apex. Conidiomatal wall composed of several layers, from outer to inner layers black, dark brown, pale brown to hyaline cells of textura angularis. Conidiophores reduced to conidiogenous cells. Conidiogenous cells lining the inner cavity, doliiform to subcylindrical, smooth-walled, hyaline, enteroblastic, phialidic conidiogenesis with periclinal thickening at the apex. Conidia solitary, ellipsoidal to obovoid, rounded at the apex, aseptate, initially hyaline, becoming pale to dark brown at maturity, smooth-walled, sometimes finely verruculose, with smaller guttules at young and indistinct at maturity.

Figure 2.
Coniothyrioides thailandica sp. nov. (MFLU 22-0276, holotype). (a) & (b) Appearance of conidiomata on a submerged decaying woody substrate. (c) Longitudinal section of conidioma. (d) Conidiomatal wall. (e) The appearance of setae. (f) & (g) Conidiogenous cells with developing conidia. (h) Conidia. Scale bars: a = 200 μm, b = 100 μm, c = 50 μm, d = 20 μm, e, h = 10 μm, f, g = 5 μm.
Type species – Coniothyrioides thailandica
Note – Coniothyrioides gen. nov. is a monotypic genus associated with decaying woody substrates in salt marsh habitats in central Thailand. This genus is characterized in having pycnidial conidiomata with the cells of textura angularis wall surrounded by distinct setae, doliiform to subcylindrical, hyaline conidiogenous cells, and ellipsoidal to obovoid, aseptate and hyaline to brown conidia. Based on some conidial characteristics such as aseptate, hyaline to brown conidia the genus shares similar morphologies to coniothyrium-like taxa [ 19] , by ellipsoidal to subcylindrical conidia sharing similar characters to Coniothyrium and Neoconiothyrium [ 9, 16, 20, 24] . However, other accepted genera in Coniothyriaceae differ from this genus in conidial morphologies: Foliophoma has only hyaline conidia except for F. camporesii D. Pem & K.D. Hyde; Hazslinszkyomyces has muriformly septate conidia [ 27] ; Ochrocladosporium has cladosporium-like conidia [ 35] . Moreover, phylogenetically Coniothyrioides forms a distinct lineage within Coniothyriaceae ( Fig. 1). Coniothyrium carteri (Gruyter & Boerema) Verkley & Gruyter (LG1401_MS6E) was the closest species based on BLAST result of ITS (94.33% similarity) and C. telephii (Allesch.) Verkley & Gruyter (UTHSC:DI16-189) was the closest species LSU sequence data (99.31% similarity) and sequences are lacking for SSU in the GenBank. The genus is known from its asexual morph and the sexual morphology was not observed.
In our phylogenetic analyses, Foliophoma species were grouped outside of Coniothyriaceae with closer to Libertasomycetaceae and Pleosporaceae species. Foliophoma was introduced by Crous & Groenewald [ 27] to accommodate F. fallens (Sacc.) Crous, in Coniothyriaceae based on the parsimony analyses of single LSU and ITS sequence data. Foliophoma camporesii was later introduced based on morphology and maximum likelihood analyses of LSU-SSU- ITS sequence data by Hyde et al. [ 25] . Based on morphology, Foliophoma species share similar characteristics to the species of Coniothyriaceae in having dark brown conidiomata, conidial wall with textura angularis cells, phialidic conidiogenesis sometimes with periclinal thickening or percurrent proliferation, and mainly ellipsoidal shaped conidia. However, the type species of the genus, F. fallens differs other Coniothyriaceae taxa in having eustromatic conidiomata. Based on this taxonomic uncertainty, more fresh collections with additional coding genes are required to clarify the accurate placement of Foliophoma.
Coniothyrioides thailandica Wijes., M.S. Calabon, E.B.G Jones & K.D. Hyde, sp. nov.
Index Fungorum number: 555050; Facesoffungi number: 13902
Etymology – The name reflects the county Thailand, from where the species was isolated.
Saprobic on a submerged and decaying woody substrate. Sexual morph: Undermined. Asexual morph: Coelomycetous. Conidiomata 150–200 μm high, 100–150 μm diam. (x̄ = 160 × 130 µm), pycnidial, semi-immersed, erumpent through the host substrate, globose to subglobose, solitary, scattered to aggregated, uni-loculate, ostiolate, covered by setae, rigid when dehydrated, black. Setae 3–5 µm wide, originating from the outermost layers of conidiomatal wall, divergent, brown, with hyaline apex, septate, smooth-walled, uniformly wide from base to apex. Conidiomatal wall 15–20 µm wide, equally thickened, composed of several layers, outermost layers dark brown to black, towards inside pale brown to hyaline cells of textura angularis, surrounded by setae. Conidiophores reduced to conidiogenous cells. Conidiogenous cells 4–5 μm long × 2.5–3.5 μm wide, lining the inner cavity, doliiform to subcylindrical, smooth-walled, hyaline, enteroblastic, phialidic conidiogenesis with periclinal thickening at the apex. Conidia 3–5 × 2.5–3 μm (
$\overline x $ Culture characteristics – On MEA, colony circular with a filamentous margin, reaching 40–45 mm diam. in 25 d at 25 °C, light gray from above, brown from center becoming light gray in the margin below, surface rough, dry, flat, with dense mycelia, edge filiform.
Material examined – Thailand, Pranburi Province, on a submerged decaying wood, 23 March 2021, Mark S. Calabon, SPAR26 (MFLU 22-0276, holotype), ex-type living cultures, MFLUCC 22-0193.
GenBank numbers – ITS = OQ023276, LSU = OQ023277, SSU = OQ025050.
Notes – Coniothyrioides thailandica sp. nov. shares morphological characters with other representatives in Coniothyriaceae in having pycnidial, globose, uni-locular conidioma with a central ostiole, peridial wall with the cells of textura angularis, and doliiform to subcylindrical conidiogenous cells, phialidic conidiogenesis with a periclinal thickening at the apex. The synopsis of asexual morphological characters for the generic types of the family including their hosts and localities is presented in Table 3. Based on the presence of conidiomatal setae, our species (MFLU 22-0276) resembles Neoconio thyrium [ 24] . In addition, our species resembles Foliophoma camporesii (MFLU 17-1006) in having hyaline to brown and aseptate conidia but differs in having larger conidiomata (150–200 × 100–150 vs 40–47 × 40–69 μm) and the presence of setae on the wall ( Table 3). Phylogenetically, our strain (MFLUCC 22-0193) formed an independent lineage within Coniothyriaceae with 94% ML and 0.99 BI statistical support ( Fig. 1). The base pair differences between our stain and the strains represent type species of other genera in Coniothyriaceae are listed ( Table 4). Thus, the evidence based on both morphology and phylogeny, we establish Coniothyrioides as a new genus in Coniothyriaceae with C. thailandica as the type species.
Table 3. Synopsis of asexual morphological characters of related genera of Coniothyriaceae.
Species Conidiomata (µm) Conidiomata wall (µm) Conidiogenous cells (µm) Conidia (µm) Habitat(s) and host(s) Locality Reference Coniothyrioides thailandica (holotype: MFLU 22-0276) 150–200 high, × 100–150 diam., pycnidial, semi-immersed, erumpent, dark brown to black, globose to subglobose,
uni-locular, ostiolate15–20 wide, black, dark brown to hyaline cells of textura angularis,
Brown, septate setae (3–5 µm wide,) with hyaline apex4–5 long × 2.5–3.5 wide, hyaline, doliiform to subcylindrical, enteroblastic, phialidic conidiogenesis with periclinal thickening 3–5 × 2.5–3, ellipsoidal to obovoid, aseptate, rounded at apex, initially hyaline, becoming pale to dark brown at maturity On decaying wood in salt marsh habitat Thailand This study Coniothyrium
palmarum (CBS 400-71)Immersed, dark brown, globose, pale to uni-locular brown, thick-walled cells of textura angularis hyaline, phialidic conidiogenesis, doliiform to cylindrical Subcylindrical, spherical, ellipsoid or broadly clavate, 0(–1)-septate, apex obtuse, brown, base truncate, sometimes minute marginal frill On Chamaerops humilis ( Arecaceae) Italy [ 16]PP,
[ 20]GNFoliophoma fallens (holotype: CBS 284.70) 120–250 wide, eustromatic, globose, uni-multi locular,
1–3 ostiolate3–6 layers,
brown textura angularis5–7 × 4–5, hyaline, phialidic conidiogenesis with thickening or proliferation at apex, dolliform to subcylindrical,
periclinal(5–)5.5–6(–7) × (3–)4(–5),
broadly ellipsoidal, aseptate, hyaline, guttulate or granular, apex obtuse, base truncate to bluntly roundedLeaf spot on Nerium oleander ( Apocynaceae) Italy [ 27] *Foliophoma camporesii (holotype: MFLU 17-1006) 40–47 × 40–69, pycnidial, immersed to semi-immersed, globose to subglobose, ellipsoidal or irregular, carbonaceous 15–40, 1–2-layered of cells of textura angularis 2–4 × 2–3, hyaline, globose to short cylindrical, phialidic conidiogenesis with periclinal thickening or percurrent proliferation at apex 2–6 × 3–5, ovoid to ellipsoidal, aseptate, hyaline when immature, brown at maturity On dead stems of Maclura pomifera ( Moraceae) Italy [ 25] Hazslinszkyomyces aloes (≡ Camarosporium aloes: ex-type - CPS 21572) 250 diam, pycnidial, erumpent, brown, globose, central ostiolate 3–6 layers of brown textura angularis
5–10 × 4–5, hyaline,
ampulliform to doliiform, apex with several inconspicuous percurrent proliferation,(9–)11–13(–14) × (4–)6–7(–8), ellipsoid, initially hyaline, aseptate, becoming pale brown, subcylindrical
to clavate or obovoid with 3 transverse eusepta, constricted at median septum or not, apex obtuse, base bluntly rounded to truncateDead bark of Aloe dichotoma ( Xanthorrhoeaceae) South Africa [ 27] Neoconiothyrium persooniae (ex-type CPC 32021 = CBS 143175) 100–200 diam, superficial, ellipsoid to obpyriform, 1–2 papillate ostioles, 10–15 diam, with or without setae 3–6 layers, hyaline textura angularis 5–8 × 4–5, hyaline, doliiform to ampulliform, phialidic, with periclinal thickening or percurrent proliferation (5–)6–7(–8) × 3(–4), ellipsoid to subclavate, aseptate, initially hyaline medium brown, becoming cylindrical and at times 1-septate, apex subobtuse, base bluntly rounded On leaves of Persoonia laurina subsp. laurina ( Proteaceae) Australia [ 24] Ochrocladosporium elatum (CBS 146.33) – – Integrated as lateral peg-like loci on hyphal
cells, or erect, subcylindrical, up to 25 µm long, 2.5–4 µm wide,
with 1–3 terminal loci, occasionally lateral, 1–1.5 µm wideRamoconidia, 10–40 × 3–5, subcylindrical to ellipsoid, hyaline to pale brown, 0(–1)-septate, giving rise to branched
chains of conidia that are subcylindrical to ellipsoid, aseptate, (7–)8–10(–14) × (3–)4(–4.5), olivaceous brownWood pulp Sweden [ 35] '–' observed morphologies on cultures, therefore conidiomata and wall characters are not recorded. '*' species which is not represent a generic type. GN- based on the generic description. PP- based on the photographic plate provided. Table 4. The base pair comparisons of our strain (MFLUCC 22-0193) with the strains representing type species of other genera in Coniothyriaceae.
Species Strain LSU SSU ITS Coniothyrium palmarum CBS 400-71 14/800 (1.75%) 3/948 (0.3%) 69/487 (14.10%) Foliophoma fallens CBS 284.70 8/800 (1%) 2/948 (0.2%) 66/497 (13.27%) Hazslinszkyomyces aloes CPC:21572 6/800 (0.75%) – 52/497 (10.46%) Neoconiothyrium persooniae CBS:143175 20/800 (2.5%) – 49/497 (9.85%) Ochrocladosporium elatum CBS 146.33 13/800 (1.62%) – 53/497 (10.66%) Members of Coniothyriaceae have high morphological plasticity and it is not adequate to use only morphology for identification at the genus level. Coniothyrium dolichi (Mohanty) Verkley & Gruyter (≡ Pyrenochaeta dolichi Mohanty, CBS 124140) and C. glycines (R.B. Stewart) Verkley & Gruyter (≡ P. glycines R.B. Stewart, CBS 124455) form a separate clade within Coniothyriaceae ( Fig. 1). Based on the morphology observed from corn meal agar medium (CMA) by Grondona et al. [ 54] , C. dolichi differs to our species by having two types of conidiogenous cells including discrete, ampulliform conidiogenous cells and integrated, cylindrical conidiogenous cells on filiform, septate conidiophores in the same conidioma, while our species has doliiform to subcylindrical conidiogenous cells and conidiophores are reduced to conidiogenous cells [ 54] . The pycnidial conidiomata and the ostiole of C. dolichi covered by dark brown, septate setae resembles our species and conidia are aseptate, ellipsoid, and hyaline with more or fewer guttules while our species has brown conidia at maturity [ 54] . The original description of C. dolichi, mentioned that conidia were greenish-yellow in mass similar to coniothyrium-like conidia, as well as to our species [ 55]. Also, a monodictys-like synanamorph was reported in C. dolichi based on its dark brown to black, dictyosporous conidia by differs from our species [ 17, 54, 56] . Coniothyrium glycines produces monophialidic, ampulliform, conidiogenous cells and aseptate, ellipsoidal conidia (4–8 × 1–3 µm), while our species has doliiform to subcylindrical conidiogenous cells [ 57, 58] . The unique character of C. glycines is well-defined, dark brown to black, melanized sclerotia covered with setae which differs from our species and other Coniothyrium taxa. Based on the multi-gene phylogeny provided by de Gruyter et al. [ 17] and the results of our study, the placements of these two species were confirmed in Coniothyriaceae. Also, Coniothyrium triseptatum Dayar., Thyagaraja & K.D. Hyde (MFLU 19-0758) creates a separate lineage in Coniothyriaceae ( Fig. 1) and only sexual morph was reported for this fungus. Therefore, we could not compare the morphology of C. triseptatum with our species [ 31] .
-
In this study, we introduced the novel genus Coniothyrioides in Coniothyriaceae, with C. thailandica as the type species, following the guidelines and major criteria for defining generic and species boundaries in Dothideomycetes by Chethana et al. [ 59] and Pem et al. [ 60] . The coelomycetous asexual morph of Coniothyrioides was associated with the decaying and submerged wood in the salt marsh habitats. Traditionally, morphology is used to delimit coelomycetes by considering the characteristics of conidiomata, conidiophores, conidiogenesis, and conidia including host associations [ 20, 32, 61] . However, accurate taxonomy of most coniothyrium-like species is challenging because of their simplicity, plasticity, and morphological variations [ 19] . In our study, genera in Coniothyriaceae differ in some conidial morphologies. For instance, Coniothyrium is characterized by aseptate to 1-septate, ellipsoidal to clavate or cylindrical, brown conidia [ 9, 17, 62] , Foliophoma with aseptate, ovoid ellipsoidal, only hyaline or hyaline-brown conidia, and Hazslinszkyomyces ellipsoidal to obovoid, transversely and muriformly septate, uniformly brown conidia [ 27] . Neoconiothyrium species have aseptate or 1-septate, ellipsoid to subclavate or subcylindrical, hyaline to medium brown conidia [ 24] while Ochrocladosporium species have cladosporium-like pale brown, aseptate or 1-septate conidia that occurring in branched chains [ 35] . Coniothyrioides thailandica is characterized by aseptate, ellipsoidal to obovoid, hyaline to pale or dark brown conidia. However, the phylogenetic analyses in this study reveal these morphological differences are not strong enough for generic delimitation of the family. Some characteristics of Coniothyrioides overlap with those of other accepted genera in the family, such as the conidiogenous cell morphology of Coniothyrium and Foliophoma which have doliiform to cylindrical or subcylindrical, hyaline, phialidic conidiogenesis with periclinal thickening and conidia show aseptate, ellipsoid-associated shapes, and hyaline to brown pigmentation ( Table 3).
The number of fungi was estimated at between 2.2 to 3.8 million [ 63] , with about 100,000-150,000 known species and fungus-like taxa [ 64 − 66] . There are 151,834 species listed in Species Fungorum [ 67] . An up-to-date online database (
https://coelomycetes.org/ ) for coelomycetes is being implemented [ 21, 32] . As coniothyrium-like taxa are frequently collected and morphologically similar, it is likely that they will remain unidentified. Therefore, it is to be expected that if molecular data are incorporated in morpho-taxonomic studies of these groups, will help identify many more novel taxa. This has occurred in other genera, which are plant pathogens and ecologically more important. According to Bhunjun et al. [ 65] Coniothyrium is one of the most speciose genera listed in Species Fungorum in 2021 and studies of coniothyrium-like taxa may yield more novel species. A few records of Coniothyriaceae taxa have been identified in salt marsh ecosystems, such as Coniothyrium obiones Jaap (India and Portugal) and as unidentified Coniothyrium species (USA) [ 2] . However, Wanasinghe et al. [ 29] referred the placement of C. obiones in Neocamarosporiaceae based on multi-gene phylogeny. In marine habitats, C. cerealis E. Müll. was isolated as an alga-derived fungus in the Baltic Sea by Elsebai et al. [ 68, 69] .In this study, we discussed the morphology and multi-gene phylogenetic analyses results of our new collection to verify its identity and phylogenetic placement in Coniothyriaceae. Based on ecological and geographical data on salt marsh fungi, we noted lack records of Coniothyriaceae worldwide (see Calabon et al. [ 2] ). Thus, we propose that additional collections be conducted in order to identify other Coniothyriaceae taxa and improve our understanding of fungal diversity in salt marsh ecosystems, a topic that is currently understudied.
S.N. Wijesinghe would like to thank Mae Fah Luang University on behalf of the Graduate Studies Support Grant (Grant No. Oh 7702(6)/125) for financial support and Mushroom Research Foundation (MRF), Thailand. Further, S.N. Wijesinghe would like to thank Dr. Udeni Jayalal, Dr. Samantha C. Karunarathna, and Dr. Gothamie Weerakoon for their precious advice during this study. M.S. Calabon is grateful to the Department of Science and Technology – Science Education Institute (DOST-SEI). K.D. Hyde would like to thank the National Research Council of Thailand (NRCT) grant 'Total fungal diversity in a given forest area with implications towards species numbers, chemical diversity and biotechnology' (Grant No. N42A650547).
-
Kevin David Hyde is the Editorial Board member of Journal Studies in Fungi. He is blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of Kevin David Hyde and his research groups.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Wijesinghe SN, Calabon MS, Xiao Y, Jones EBG, Hyde KD. 2023. A novel coniothyrium-like genus in Coniothyriaceae ( Pleosporales) from salt marsh ecosystems in Thailand. Studies in Fungi 8:6 doi: 10.48130/SIF-2023-0006 

a novel coniothyrium-like genus in Coniothyriaceae ( Pleosporales) from salt marsh ecosystems in Thailand
- Received: 14 December 2022
- Accepted: 15 January 2023
- Published online: 27 February 2023
Abstract: In this study, a novel coniothyrium-like genus Coniothyrioides is introduced to Coniothyriaceae based on a fresh fungal collection from salt marsh habitats in Thailand. Coniothyrium-like taxa are taxonomically controversial and have been classified into different families in Pleosporales such as Didymosphaeriaceae ( Alloconiothyrium and Paraconiothyrium), Coniothyriaceae ( Coniothyrium) and Didymellaceae ( Microsphaeropsis). However, our novel genus shares similar morphology to some key characters in Coniothyriaceae in having dark, globose pycnidia, uni-locular conidiomata, a central ostiole, a peridium of textura angularis cells, and doliiform conidiogenous cells with a periclinal thickening at the apex, while conidial morphologies are diverse. The presence of setae arising from the outer peridial wall is the main difference between Coniothyrioides and other closely related Coniothyriaceae genera. Phylogenetically, LSU-SSU-ITS sequence analyses confirm the placement of this novel genus as a distinct lineage within Coniothyriaceae. Species boundaries were defined, based on morphology and multi-gene phylogenetic analyses using maximum likelihood and Bayesian inference analyses. The comprehensive descriptions and micrographs are provided. Our findings expand the taxonomic knowledge of Ascomycota in salt marsh ecosystems.
-
Key words:
- 2-new taxa /
- aquatic fungi /
- Ascomycota /
- Dothideomycetes /
- marine habitats