









-
The plant root is an important organ throughout the life cycle of almost all terrestrial plant species. Plant roots not only anchor plants to the soil but also facilitate water and nutrient uptake (Benfey and Scheres, 2000; Luo et al., 2020b). Root exudates engineer rhizosphere microbiota. In turn, microbiota play pivotal roles in plant nutrient use efficiency and combat harmful organisms (Hopkins et al., 2013). Therefore, root development and its system architecture (RSA) play an important role in crop productivity, and illustrating the mechanism of root development and architecture adapting to the ever-changing environment provides meaningful information for breeding and cultivation.
RSA is a post-embryonic development process. For instance in Arabidopsis, the most acknowledged model plant: its primary root elongation is coordinated by the cell division activity of the root apical meristem, and subsequent elongation of the cells when they enter the elongation zone. Its lateral roots (LRs) develop along the primary root with regular spacing. The LRs development initiates from a patch of xylem pole pericycle cells that undergo nuclear migration, asymmetric cell division, followed by cell wall remodeling of the overlaying cells, and after several subsequent rounds of cell division in the developing primordium, finally result in emergence of a LR (Dubrovsky et al., 2008; de Smet et al., 2008; de Rybel et al., 2010; Vermeer et al., 2014). The initiation and development of LRs have been demonstrated to be governed by the plant hormone auxin through its biosynthesis, transport and redistribution in the root (by AUXs and PIN auxin carriers), and signaling pathway (Adamowski and Friml, 2015; Chen et al., 2015; Kircher & Schopfer, 2018). More recently, LR development was found to be triggered by oscillation of auxin related gene expression stimulated in a confined root zone (known as oscillation zone, OZ), where a patch of xylem pole pericycle cells are primed to divide and make a lateral root primordium (LRP) (Moreno-Risueno et al., 2010; van Norman et al., 2013). This oscillatory process is described as the root clock, and is characterized by both the amplitude and the frequency of gene expression in OZ visualized by DR5:Luciferase expression (Moreno-Risueno et al., 2010; Xuan et al., 2015; Duan et al., 2021). It is proposed that the oscillatory behavior of gene expression is dictated by the generation of auxin pulses in the main root derived from a release of auxin from dying lateral root cap (LRC) cells that are undergoing programmed cell death (PCD) (Xuan et al., 2016; Möller et al., 2017).
The development of plant root is continuous and sensitive to the environmental factors such as light, temperature, moisture, soil pH and ion concentration (van Gelderen et al., 2018; Kim et al., 2020; Luo et al., 2020a; Msimbira & Smith, 2020; Wang et al., 2022; Liu et al., 2022). For instance, lateral roots are found to emerge towards water sources under water-deficit (termed as hydro-patterning), and reduce initiation under drought stress (xero-branching response); primary root can adjust growth direction to avoid salt stress (halo-tropism) (Galvan-Ampudia et al., 2013; Bao et al., 2014; Giehl & von Wiren, 2014; Feng et al., 2016; Dietrich et al., 2017; Orman-Ligeza et al., 2018). Soil pH is a pivotal parameter shaping root architecture. Both soil acidification and alkalization have negative effects on plant root development (Msimbira & Smith, 2020), and is looming worldwide. For instance, according to the soil survey results, the area of acidified soil in China is as high as 2.04 × 108 hm2, mainly distributed in the subtropical and tropical areas south of the Yangtze River, Yunnan, Guizhou and Sichuan (Guo et al., 2010). Acidified soil environments cause a decrease in plant root cellular pH. When the pH value of soil solution is below 5.0, the plant cell biological activities are gradually inhibited as the concentration of Al3+ and Mn2+ ions reaching a toxic level, and interfer with the absorption of essential macronutrients (Guo et al., 2010). A high acidic environment is also harmful to root development because the outside proton over inflow into cytoplasm is destructive to root development. Soil alkalization means that Ca2+ and Mg2+ adsorbed in the soil colloid are replaced by a large amount of Na+ during soil desalination, resulting in soil pH ≥ 8.5 (Lagerwerff & Brower, 1972; Zhang et al., 2021). The salt content of the alkaline soil is usually below 0.5%, but the soil solution contains a large amount of Na2CO3 and NaHCO3. Alkaline soil is easy to harden, mainly due to the changes of physical and chemical properties, resulting in poor water retention and air permeability making it difficult for roots to breathe and absorb water. The High-affinity K+ transporter (HKT) family genes, Na+/H+ exchanger (NHX) family genes, and Salt overly sensitive 1 (SOS1) are overexpressed in plant roots, stems and leaves when plants roots are subjected to alkaline stress (Choudhury et al., 2017).
As shown above, it has been reported that plant root development could be largely affected by environmental pH conditions (Cross et al., 2021). Recent studies also showed both LR formation and PR elongation are restricted under low environmental pH (acidic) conditions (Ojeda-Rivera et al., 2020; Friml et al., 2022). In this study, gradient pH setting treatments showed that the seedlings cultured in slight alkaline conditions (pH = 7.5) had better primary and lateral root growth. We found that slightly alkaline conditions are essential for maintaining the oscillation periodicity, amplitude in the oscillation zone (OZ), and promoting periodic LR branching. Further experiments showed that slightly alkaline promoting root development were mainly mediated by polar auxin transport (PAT), and auxin signaling genes: TIR1, AFB2, ARF7 and ARF19 were triggered for lateral root development. Therefore, slightly alkaline is a key factor in coordinating the effects on root development via modulating auxin homeostasis in Arabidopsis.
-
Arabidopsis thaliana (Columbia) wild type lines were used in this study. Seed surfaces were sterilized with 70% (v/v) ethanol for 2 min followed by 30% (v/v) bleach for 15 min. Subsequently, the seeds were rinsed at least five times with sterile water, and kept at 4 °C in the refrigerator. After 3 d of vernalization, the Arabidopsis seeds were sown on Petri dishes containing sterile half-strength Murashige and Skoog (1/2MS) medium (0.5× MS salts, 1% sucrose, 10 mg/L myo-inositol, 0.5 g·L−1 2-(N-morpholino) ethanesulfonic acid, and 1% w/v agar) in a growth chamber (AR1200, Wuhan Ruihua Instrument & Equipment, China) at 22 °C under continuous light (100 μmol·m−2·s−1). The medium pH was adjusted with KOH. Arabidopsis seedlings were directly germinated on the 1/2MS medium with indicated pH, and their root phenotype was analyzed after 10 d.
Chemical preparation and treatments
-
1-N-Naphtyl Phtalamic Acid (NPA) and 2-[4-(diethylamino)-2-hydroxybenzoyl] benzoic acid (BUM) (Product No. 202908) was order from Shanghai Macklin Biochemical Co., Ltd Iodacetamide (IAM, ordered from Sigma, Product No. I1149), and Propidium iodide (PI, ordered from Sigma, Product No. P4864) were acquired from Sigma. D-luciferin (Product No. L1349) was acquired from Duchefa Biochemie. PI was dissolved in ddH2O, and D-Luciferin was dissolved in 0.01% Tween 80.
For chemical treatments, the required amount of the stock solutions was added to (50 °C) melted 1/2 MS agar-containing medium, and mixed in 50-mL Falcon tubes before being poured into petri dishes (13 cm × 13 cm). Arabidopsis seeds were transferred to 13 cm square 1/2 MS agar plates containing the compounds after 5 d germination and treated for another 7 d. Primary root length and LR number were subsequently measured.
Primary root and lateral root phenotype analyses
-
To quantify the root phenotype in wild-type plants and mutants, the emerged lateral roots of the 8-d-old seedlings were counted under a microscope. Subsequently, the whole seedlings were scanned with an EPSON XL11000 flatbed scanner and the length of the primary root was measured. For microscopic inspection of primordium stages, root samples were cleared as described previously (Malamy & Benfey, 1997), all samples were analyzed by differential interference contrast microscopy (Leica DM2500).
Histochemical analysis
-
For the visualization of the expression of DR5:GUS and pCYCB1:GUS in the root tip, the histochemical GUS analysis was performed in 3-d-old seedlings under different pH value conditions, essentially as described previously (Beeckman & Engler, 1994), and all samples were analyzed by differential interference contrast microscopy (Leica DM2500 and OLYMPUS MVX10).
Luciferase imaging and expression analysis
-
A Lumazone imaging system equipped with a charge-coupled device (CCD) camera (Andor iKon-M, DU934P-BV) was used to image the luciferase signal in the vertical growing Arabidopsis root tip according to a previous study (Xuan et al., 2018).
Confocal microscopy and quantification
-
A Leica SP8X laser-scanning microscope were used for fluorescence imaging of roots. For the propidium iodide (PI)-treated root images, the seedlings were stained with 2 mg/mL PI for 5 min, washed with water, then seedlings transferred to slides for confocal imaging. The quantification of fluorescence signal intensity, the cortex cell number, and cell length were performed on the individual root.
RNA-seq analysis
-
Arabidopsis seeds were germinated and cultured as described above in normal medium of pH5.8, and then after transferring them into medium of pH7.5, their roots were sampled after 0, 2 and 6 h, with three independent biological repeats. RNA-seq datasets were then analyzed following a custom protocol previously published (Duan et al., 2021). Briefly, raw data were also cleaned using Fastp v0.20 (Chen et al., 2018) and were then aligned to the Arabidopsis reference genome (TAIR10,
www.arabidopsis.org ) using STAR v2.7.8a with a splicing-aware method and two-pass model (Dobin et al., 2013). The gene expression matrix was quantified and normalized with FPKM (fragments per kilobase of transcript per million fragments mapped) using HTSeq. Only the genes with an FPKM > 1 in at least three samples were used for downstream gene expression analysis (Anders et al. 2015). DESeq2 (Love et al., 2014) was used to perform differential gene expression (DEG) analysis with Log2 (Fold change) ≥ 1 and the adjusted p-value < 0.05. Enrichment analysis based on KEGG database was carried out using PlantGSAD (Ma et al., 2021). The Benjamini–Yekutieli method was used for P-value adjustment.Statistical analysis
-
All the experiments in this study were performed at least three times. In each experiment, at least 10 individual seedlings per genotype were treated with or without slightly alkaline condition and used for further data analysis. To perform the statistical analysis regarding the time interval oscillations, at least eight individual seedlings were imaged and used for analysis. The data were analyzed using routines implemented in Prism 8 software (GraphPad Software). The significant difference among sets of data was determined by one-way ANOVA with post hoc Tukey test and Student's t test (p < 0.05). All the results are presented as the mean ± standard deviation (SD).
-
To investigate the root growth under acidic to alkaline conditions, the Arabidopsis seedlings were grown on the set of medium with pH gradient increasing from the acidic (5.8) to alkaline (9.0) since germination. Interestingly, in comparison with the seedlings grown in the medium of pH 5.8 (as the standard pH), the seedlings in neutral and slightly alkaline pH (7.0−8.0) had longer primary root (PR) and more lateral roots (PR) formation (Fig. 1a, b). This finding suggested the neutral and slight alkaline conditions could promote root development. Present research also showed that the growth of Arabidopsis PR and LR were substantially inhibited when the medium pH was at 8.5 and 9.0 (Fig. 1a, b), which hinted that the severe alkaline condition (pH ≥ 8.5) could be stressful to Arabidopsis. Simultaneously, the seedlings grown in the medium with very acidic pH (5.0) showed even shorter PR and less LRs as compared to the standard condition (pH 5.8) (Supplemental Fig. S1). These results collectively indicate that acidic and alkaline conditions hinder Arabidopsis root development, while neutral and slightly alkaline conditions promote root development.
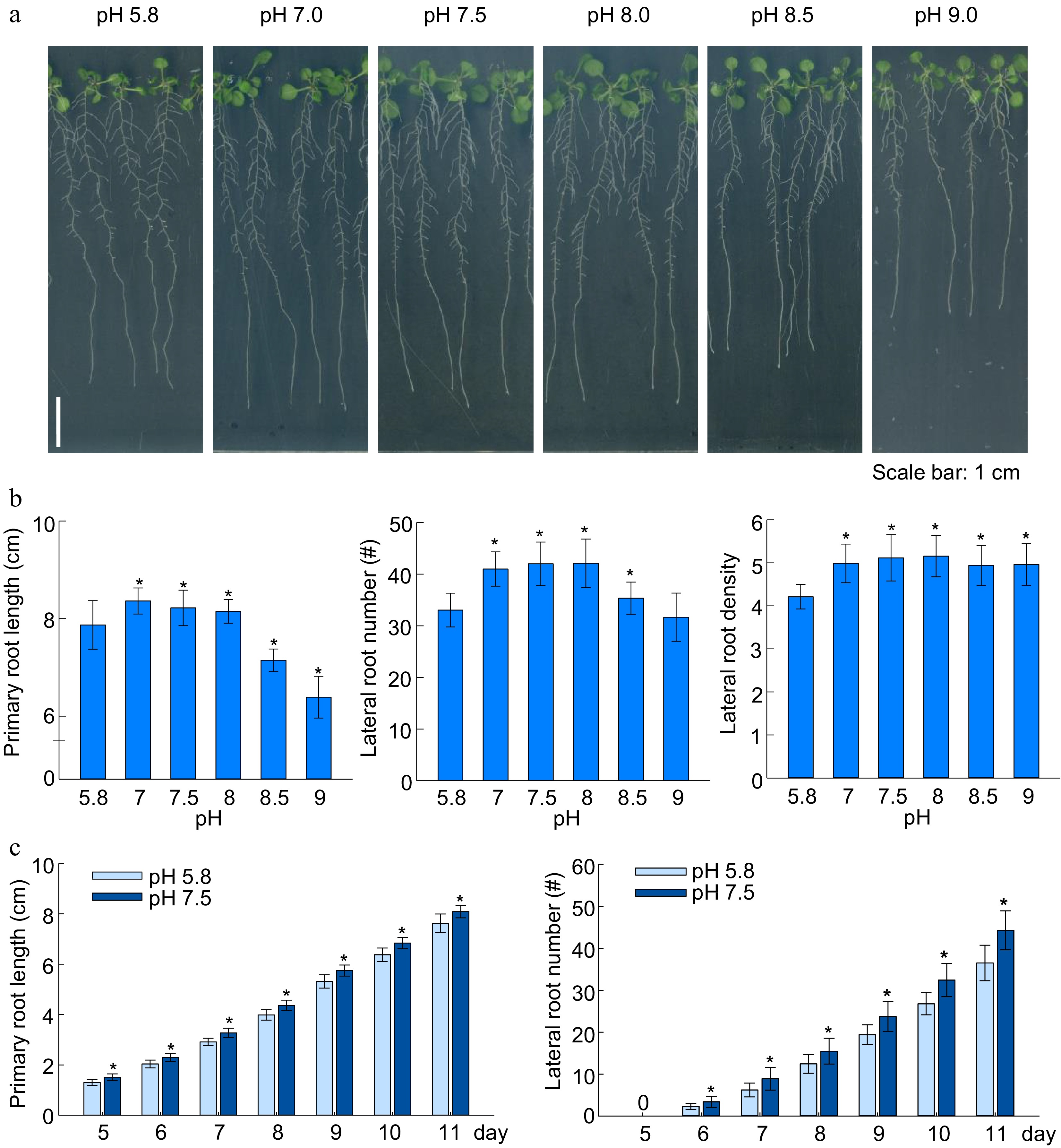
Figure 1.
Effects of alkaline and acidic pH on plant root growth. (a) Plant growth status, (b) primary root length, lateral root number, lateral root density; (c) dynamic changes of root growth of Arabidopsis under slightly alkaline (7.5) and the standard (5.8) pH cultivation. (means ± SD, asterisks denote significant differences compared with control condition pH 5.8, * p < 0.05; ** p < 0.01, n ≥ 8) (Student t-test, Bar = 1 cm).
Further investigation was carried out using two different medium cultivation of pH 5.8 (as standard) and 7.5 (standing for neutral to slightly alkaline pH) from 5-d to 11-d after germination, and the primary root length and LR number were monitored daily. As shown in Fig. 1c, throughout the whole procedure, the root development at pH 7.5 remained superior to the standard conditions. To validate and visualize the apoplastic alkalinization in root induced by the slight alkaline pH (7.5), we analyzed the apoplastic pH using the apo-pHusion marker line (Gjetting et al., 2012). In the CK (pH 5.8), the apoplastic pH remained relatively acidic, while apoplastic pH raised and promoted root growth in the slightly alkaline conditions (Fig. 2a, b).
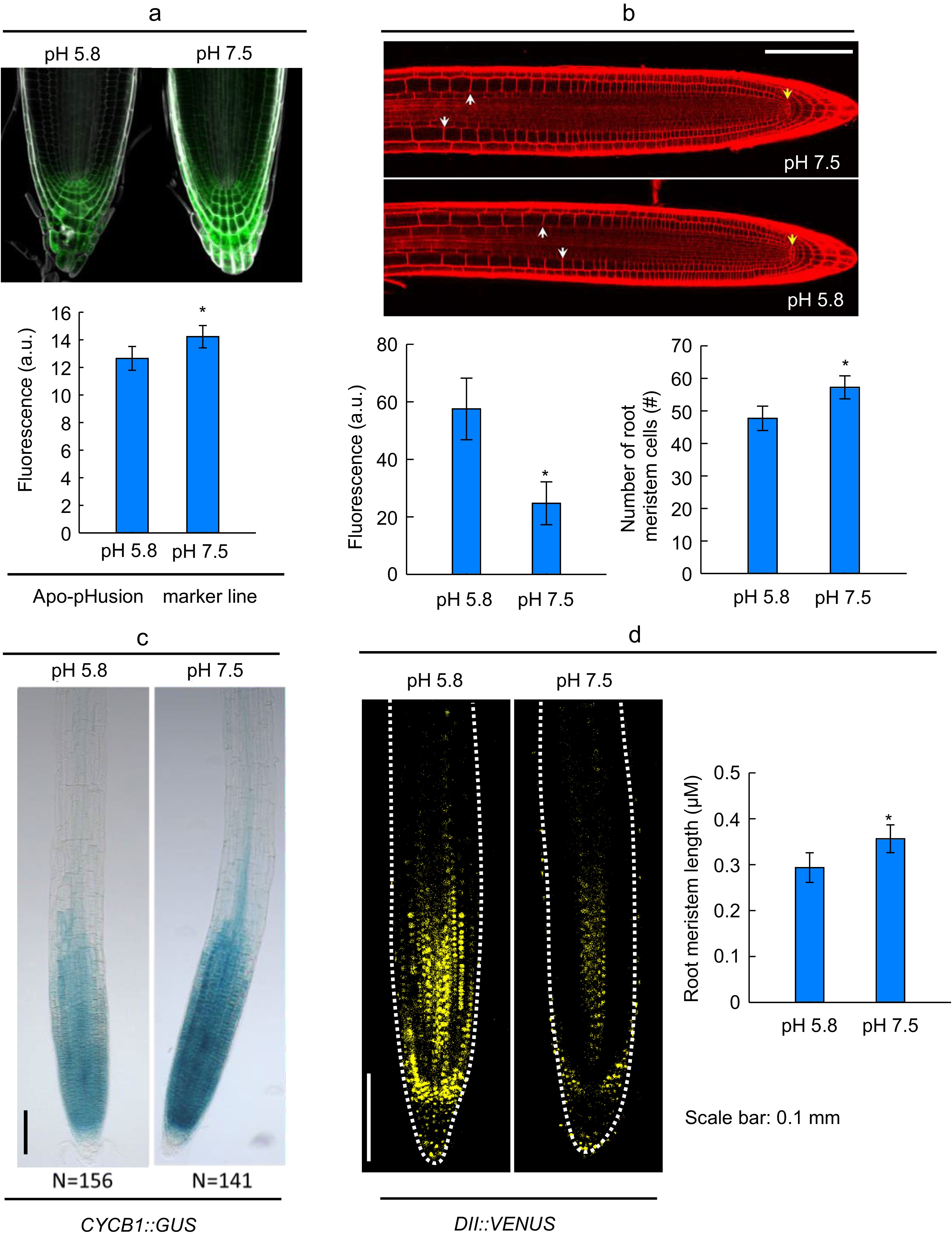
Figure 2.
Effect of slightly alkaline pH on apoplast alkalinization, auxin signal, number of stem cells, and cell cycle in primary root apex. (a), (b) Localization and quantifications of apoplastic pH in the apo-pHusion marker line in the root meristem zone of light-grown Col-0 seedlings. (c), (d) the status of the meristem of primary root apex, yellow arrows indicate quiescent center, white arrows indicate the boundary between meristem zone and elongation zone, and the column charts showed the number of root meristem cells, root meristem length. (e) GUS staining showed the expression of CYCB1 in primary root apex. (f) Fluorescent protein in root apex of DII:VENUS. Bar = 0.1 mm, and the quantitative fluorescence strength were showed in the chart, n ≥ 8. (Means ± SD, asterisks denote significant differences between columns at * p < 0.05; ** p < 0.01, n ≥ 8). (Student t-test, Bar = 0.1 mm).
Root meristem activity is triggered by slightly alkaline pH
-
The elongation of PR is determined by the cell division activity of root meristem at the root tip (Fujinami et al., 2017). We thereby checked the cell division activity of the Arabidopsis root in slightly alkaline pH (7.5) and the standard acidic (5.8). The developing roots in pH 7.5 had increased meristem-cell number and longer meristem length than those in pH 5.8 (Fig. 2c, d). The root tips in pH 7.5 had 19.98% increase in its meristem-cell number, and 21.42% increase in the meristem cell length than those in pH 5.8. Arabidopsis pCYCB1::GUS transgenic lines were cultivated in pH 7.5 and 5.8. The plant roots in the slightly alkaline medium showed stronger GUS blue color (Fig. 2e), which indicated higher cell division activity. As cell cycle progression is regulated by plant hormone auxin, we further investigated the auxin signaling in the meristem by using a high auxin sensitive reporter DII:VENUS, which is fast degraded by auxin (Brunoud et al., 2012). As shown in Fig. 2f, the Arabidopsis seedlings grown in the pH 7.5 medium showed less DII:VENUS signal (57.05% descending) in the root meristem than those in pH 5.8, indicating a higher intracellular auxin accumulation in the root meristem. Thus, the promotion on PR elongation by slightly alkaline pH might be due to the induced auxin signaling and cell division activity in the root meristem.
Slightly alkaline pH facilitates the pre-branch site formation
-
Potential effects induced by the slightly alkaline pH on LR development stage was further analyzed. The cultivation of pH 7.5 medium enhanced the LR emergence, and more LR primordia were detected, than in the standard. Furthermore, the DR5:LUC reporter gene was engaged to illustrate the early LR formation event. The transgenic Arabidopsis line DR5:LUC showed an oscillatory expression pattern in oscillation zone and triggered the formation of pre-branch sites (Fig. 3a). Pre-branch site hallmarks LR founder cells and LR primordium, which subsequently develops into LRs. The DR5:LUC expression sets along the root in pH 7.5 had a 37.99% increase compared with those in the standard (Fig. 3b). Remarkably, by quantifying the DR5:LUC signal along the primary root, we also detected a stronger DR5:LUC signal in the seedling subjected to pH 7.5 than 5.8 (Fig. 3b). These results provided evidence that the LR primordium (LRP) of the plants from pH 7.5 cultivation had higher level auxin accumulation and it is consistent with the results presented above. Time-course monitoring the DR5 oscillations demonstrated shorter oscillation course in the plant of pH 7.5 cultivation and in consequence of a higher frequency, than that in the standard (Fig. 3c, d). On the other hand, the shape of LRP was not altered by different medium pH, implying that the accumulated auxin signaling in LRPs might affect the development of LR primordium (Supplemental Fig. S2). These data strongly suggest the increased pre-branch site and LR number upon slightly alkaline pH could be due to a high frequency of pre-branch site formation.
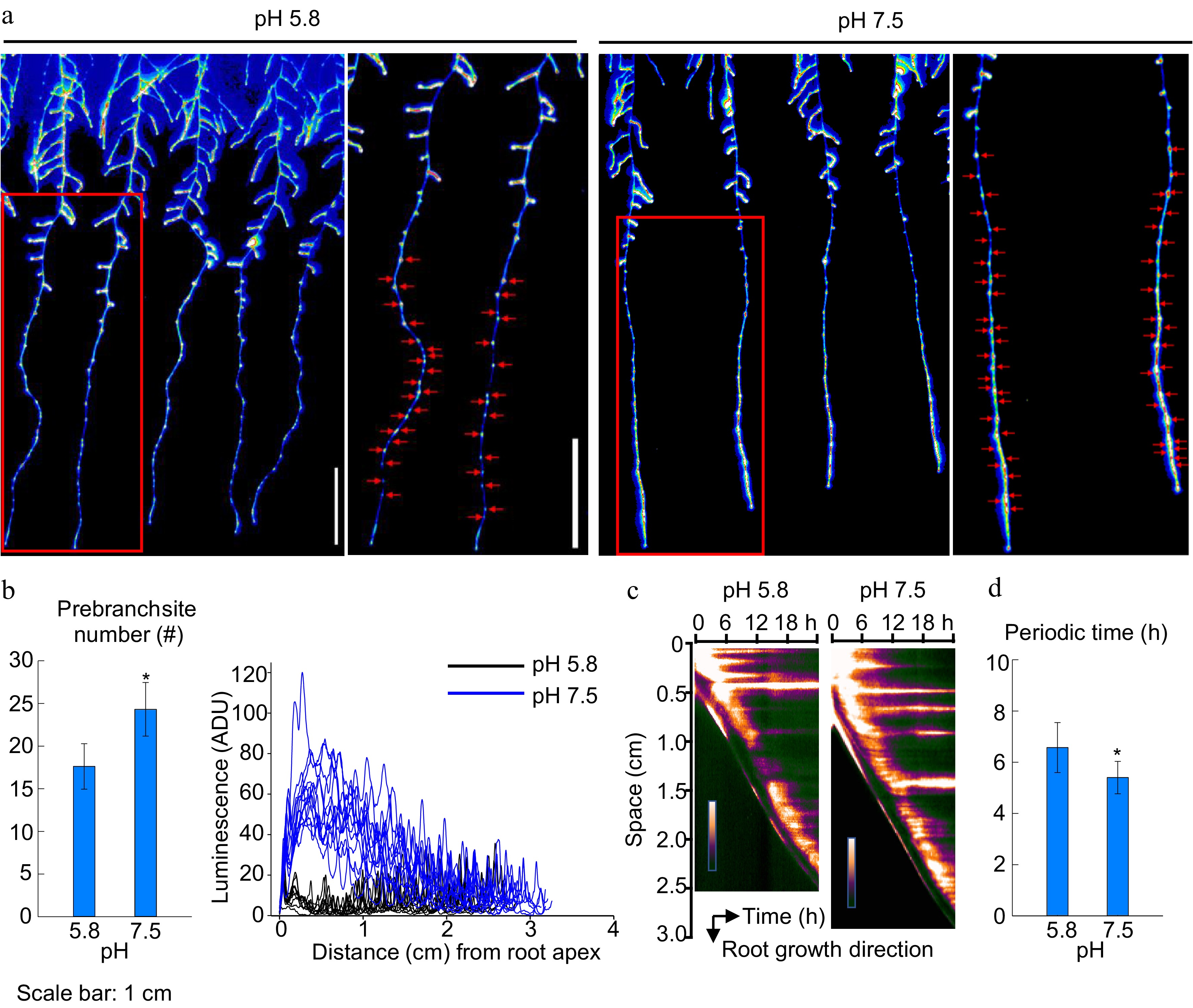
Figure 3.
Expression of Luciferase, prebranch sites and signal frequency in Arabidopsis DR5:Luciferase root. (a) Expression of Luciferase, red arrows indicate prebranch sites, (b) prebranch site number and luminescence, (c), (d) position of oscillation region signal and the cycle of biological clock signals. (Means ± SD, asterisks denote significant differences between columns at * p < 0.05; ** p < 0.01, n ≥ 8). (Student t-test, Bar = 1 cm).
PLT-restricted lateral root formation and primary root elongation is alleviated under slightly alkaline pH
-
PLETHORA (PLT)-like AP2 domain transcription factors are key regulators on both primary root elongation and LR formation in Arabidopsis. Previous research on PLT mutants showed the PLT mutant plants had shorter primary root with less LRs than wildtype (WT) (Galinha et al., 2007; Du & Scheres, 2017). Thus, plt1-4 and plt2-2 single mutants, and plt1-4plt2-2 double mutant, as well as the WT were engaged to approach the mechanism of which slightly alkaline condition promoting root development. In comparison with WT, the plt1-4 mutant and plt1-4plt2-2 double mutant, had shorter roots than WT, when plt2-2 mutant showed no visible difference (Supplemental Fig. S3a). On the other hand, in all the mutants, pH 7.5 cultivation improved root development, based on PR length and LR number. Compared with standard pH cultivation, the PR elongation was 10.41%, 12.05%, 16.84% and 127.74% raise in WT, plt1-4, plt2-2 and the double mutant, respectively; while the increase in LR number was 25.65%, 26.27%, 23.13% and 56.64% (Supplemental Fig. S3b). These data thus suggested that the activation of root growth by slightly alkaline pH might be independent from PLT signaling pathway.
Slightly alkaline pH induced LR formation is mediated by canonical auxin signaling cascades
-
Auxin is the key regulator of LR formation and primary root elongation. The knock-out mutants of auxin receptor TIR1 or its downstream signaling cascades auxin response factor ARF7 and ARF19 severely reduces LR initiation (Du & Scheres, 2017; Cavallari et al., 2021). The results described above also showed auxin might be involved in the signal pathway regulation which led to the root phenotype. Hence, the root development test on double mutants of tir1afb2 and arf7arf19 were conducted as depicted above. As shown in Fig. 4a and b, the LR formation was substantially reduced in tir1afb2 mutant. However, the LRs were not observed in arf7arf19 mutant in pH 5.8 and 7.5. Furthermore, the LR primordium was also not detected under both conditions. This phenomenon thus implicated auxin signaling ought to be essential in the process of slightly alkaline pH promoting LR formation. On the other hand, the PR elongation of arf7arf19 and tir1afb2 mutants were still promoted by slightly alkaline pH cultivation. Accordingly, we concluded that ARF7, 19 and TIR1, AFB2 should be required for LR formation rather than PR elongation in the promotion. Hence, mutant germplasms null in auxin transport and biosynthesis were engaged in further experiments.
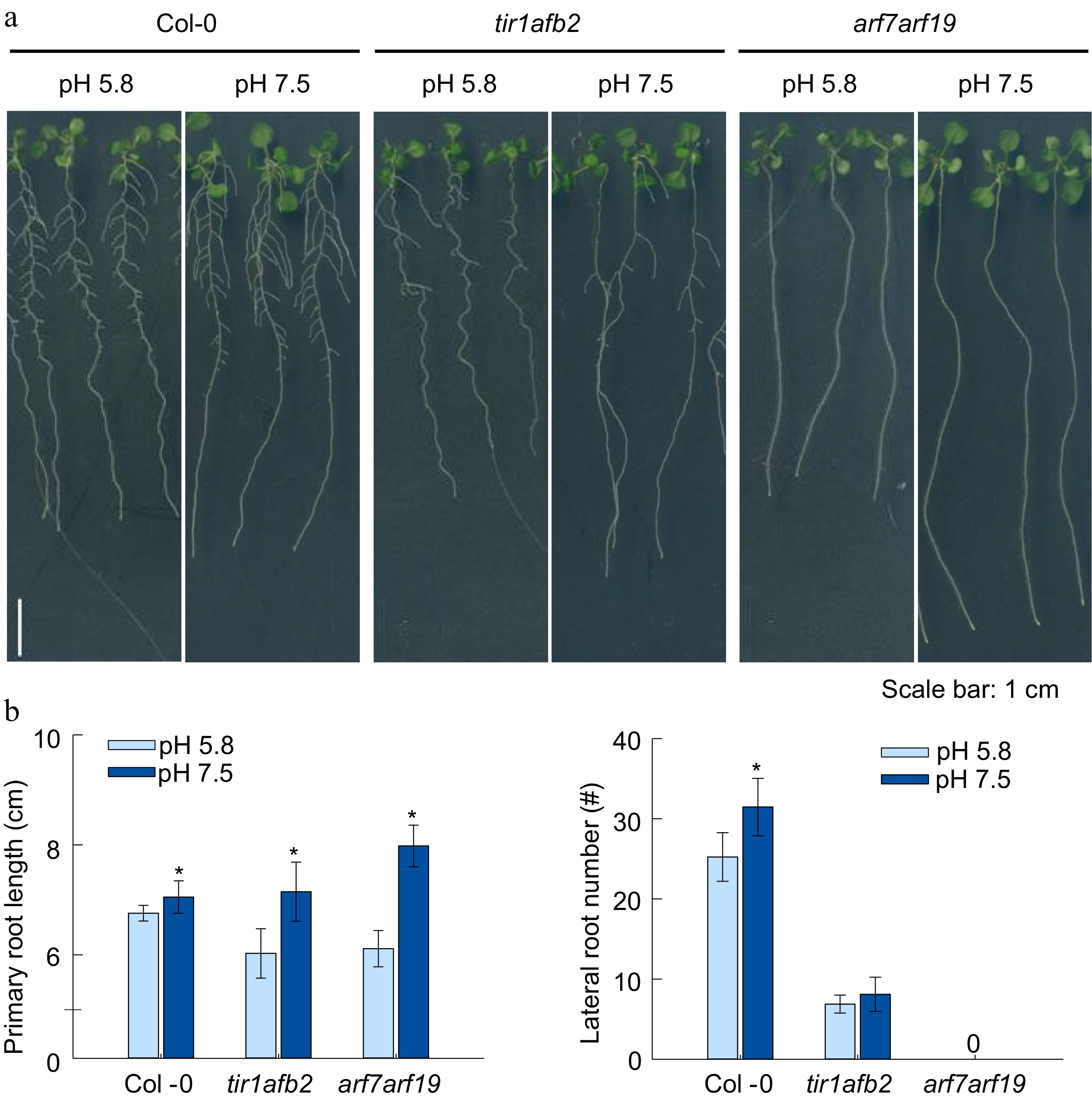
Figure 4.
Effects of alkaline conditions on the root growth of arabidopsis mutants tir1afb2 and arf7arf19. (a) Plant growth status, (b) primary root length, lateral root number. (Means ± SD, asterisks denote significant differences compared with control condition (pH 5.8) at * p < 0.05; ** p < 0.01, n ≥ 8, n.d., not detected). (Student t-test, Bar = 1 cm ).
Polar auxin transport is required for slightly alkaline pH induced LR branching and PR elongation
-
Our results have shown an enhanced auxin signaling in the roots of the seedlings grown under slightly alkaline pH (Fig. 1). In plants, the local auxin accumulation is facilitated by both auxin biosynthesis and polar auxin transport (PAT). Root cap-specific IBA-to-IAA conversion auxin has been suggested to contribute to the auxin accumulation in the meristem and oscillation zone to activate the establishment of pre-branch site (Xuan et al., 2015). The seedlings of ibr1ibr3ibr3 and ech2ibr1ibr3ibr3 mutants, which are defective on the conversion of IBA to IAA, exhibited less LR number and shorter PR length under pH 5.8 conditions. However, the LR number and PR length were significantly increased in ibr1ibr3ibr3 and ech2ibr1ibr3ibr3 mutants pH 7.5 condition (Supplemental Fig. S4). The results indicated that the root cap-specific auxin biosynthesis is not accounted for the slightly alkaline pH induced LR and PR development.
In Arabidopsis, the PAT is facilitated by both auxin efflux and influx carriers, which directs the auxin flux throughout the cells. The block of PAT results in the defects of both LR formation and primary root elongation (Ďurkovič & Lux, 2010). To investigate the involvement of PAT, we first analysis the root phenotype in the presence of two potent PAT inhibitors 1-N-NaphtylPhtalamic Acid (NPA) and 2-[4-(diethylamino)-2-hydroxybenzoyl] benzoic acid (BUM) (Kim et al., 2010). NPA was reported to block the entire auxin efflux from the cell, whilst BUM specifically dampens the transport activity of ABCB transporters. In agreement with previous reports, the treatment with NPA and BUM at a concentration ranging from 0.1 uM to 1 uM, significantly reduced LR number and primary root elongation in a dose-dependent manner, whereas BUM treatment exhibited more severe inhibitory effects on root growth than NPA treatment in both slightly alkaline (7.5) and standard (5.8) pH (Fig. 5a). Noticeably, slightly alkaline pH induced LR formation which was found to be repressed by 0.3 µM BUM, while 1 µM BUM completely blocked the LR formation and primary root elongation. Similarly, NPA also reduced the slightly alkaline pH induced LR formation and primary root elongation compared to the standard. When the NPA treatment was at 0.3 µM, the pH 7.5 cultivation showed no promotion effect on PR, but remarkably the 1.0 µM NPA treatment had switched the phenomenon of which the plant had even longer primary roots in pH 5.8 than in pH 7.5 (Fig. 5b). These findings suggested that the PAT is required for slightly alkaline-induced LR formation and primary root elongation.
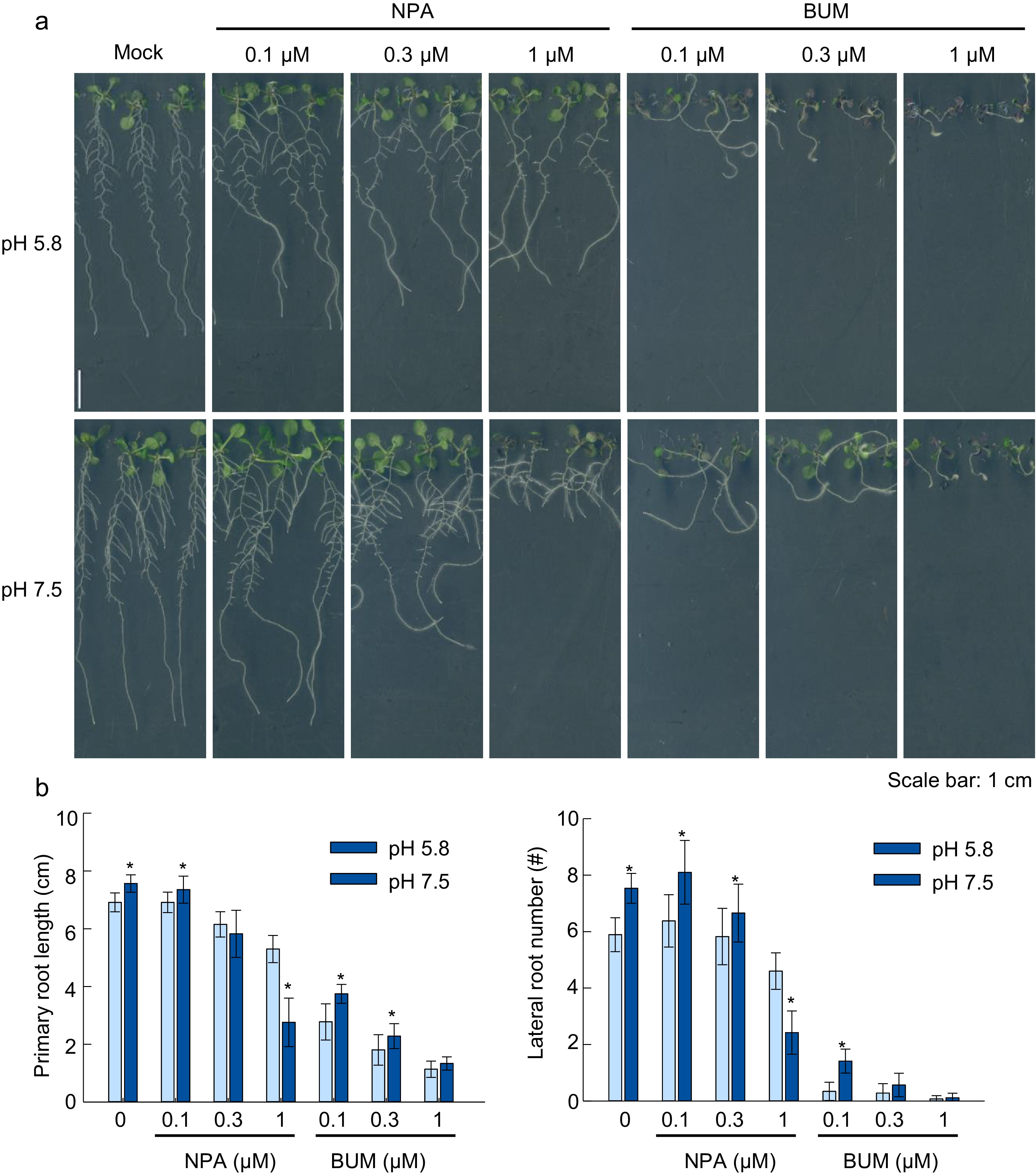
Figure 5.
Effects of auxin transport inhibitors NPA and BUM on root growth under alkaline conditions. (a) Plant growth status, (b) primary root length, lateral root number. (Means ± SD, asterisks denote significant differences compared with control conditions (pH 5.8) at * p < 0.05; ** p < 0.01, n ≥ 8). (Student t-test, Bar = 1 cm ).
We further analyzed the root phenotype of PAT defective mutants aux1-21, pin2pgp1pgp19, and pin2pin3pin7 (Fig. 6a). The aux1-21 mutant is defective for root auxin uptake (Jia et al., 2020), whereas pin2pgp1pgp19 and pin2pin3pin7 mutants are defective for auxin efflux (Li et al., 2021). To our surprise, the root growth of aux1-21 mutant remains sensitive to slightly alkaline pH as the PR elongation was significantly induced. However, the LR number and gravitropic root response of aux1-21 was not altered under slightly alkaline pH (Fig. 6b). Furthermore, the pin2pgp1pgp19 and pin2pin3pin7 triple mutants completely suppressed the slightly alkaline pH induced LR formation, while the enhancement of primary root elongation was fully repressed in pin2pin3pin7 mutant but partially inhibited in pin2pgp1pgp19 mutant (Fig. 6b). Meanwhile, we also observed that slightly alkaline condition could trigger the accumulation of PGP1-GFP, PGP19-GFP and PIN2-GFP signal in the root meristem. This result further implicated a transcriptional regulation of auxin efflux carrier coding genes triggered by slightly alkaline pH (Fig. 7). These data suggested that auxin efflux carriers were involved in the root development response to environmental pH.
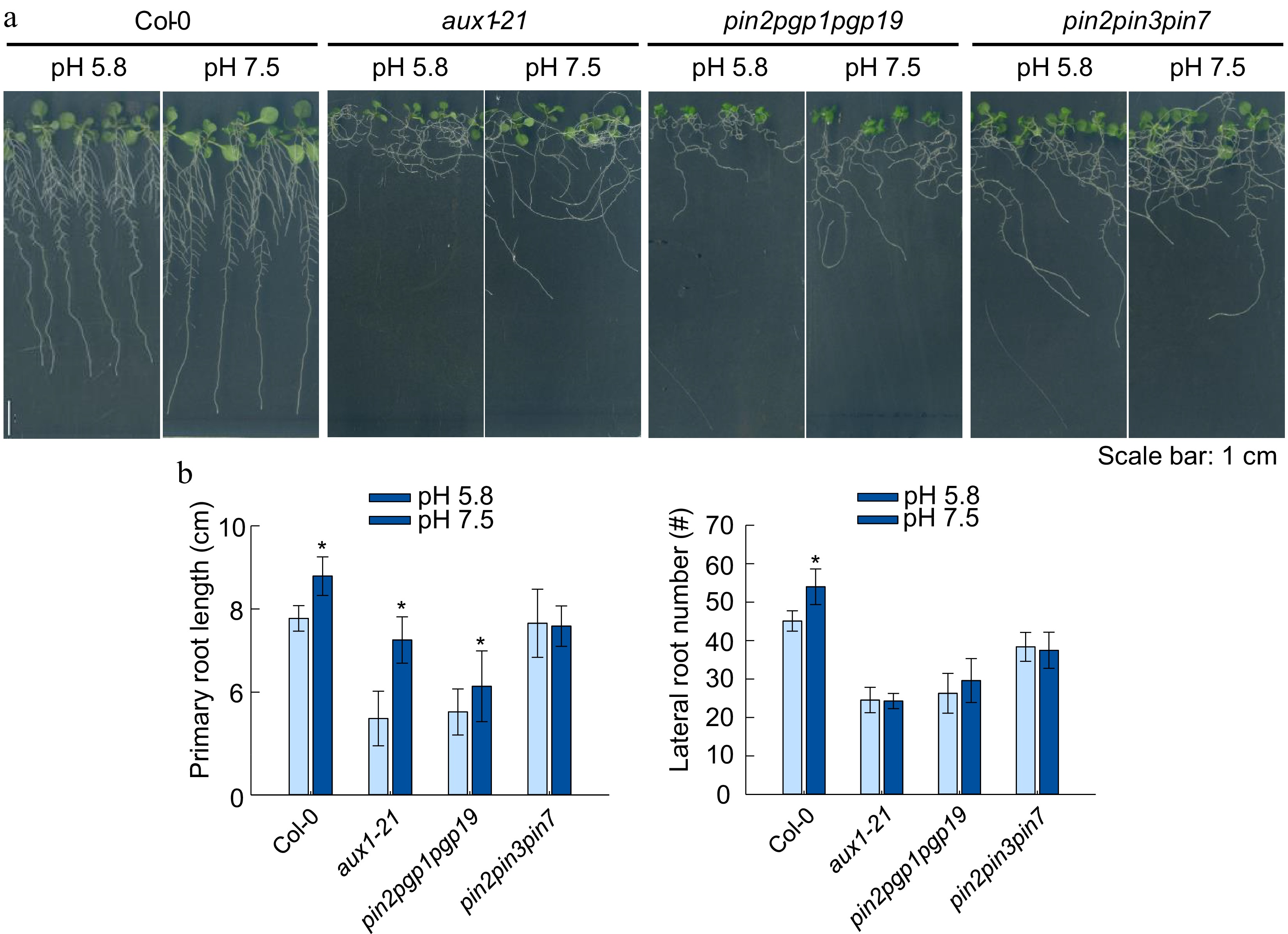
Figure 6.
Effects of alkaline conditions on the root growth of Arabidopsis mutants aux1-21, pin2pgp1pgp19, pin2pin3pin7. (a) Plant growth status, (b) primary root length, lateral root number. (Means ± SD, asterisks denote significant differences compared with control condition (pH 5.8) at * p < 0.05; ** p < 0.01, n ≥ 8). (Student t-test, Bar =1 cm ).
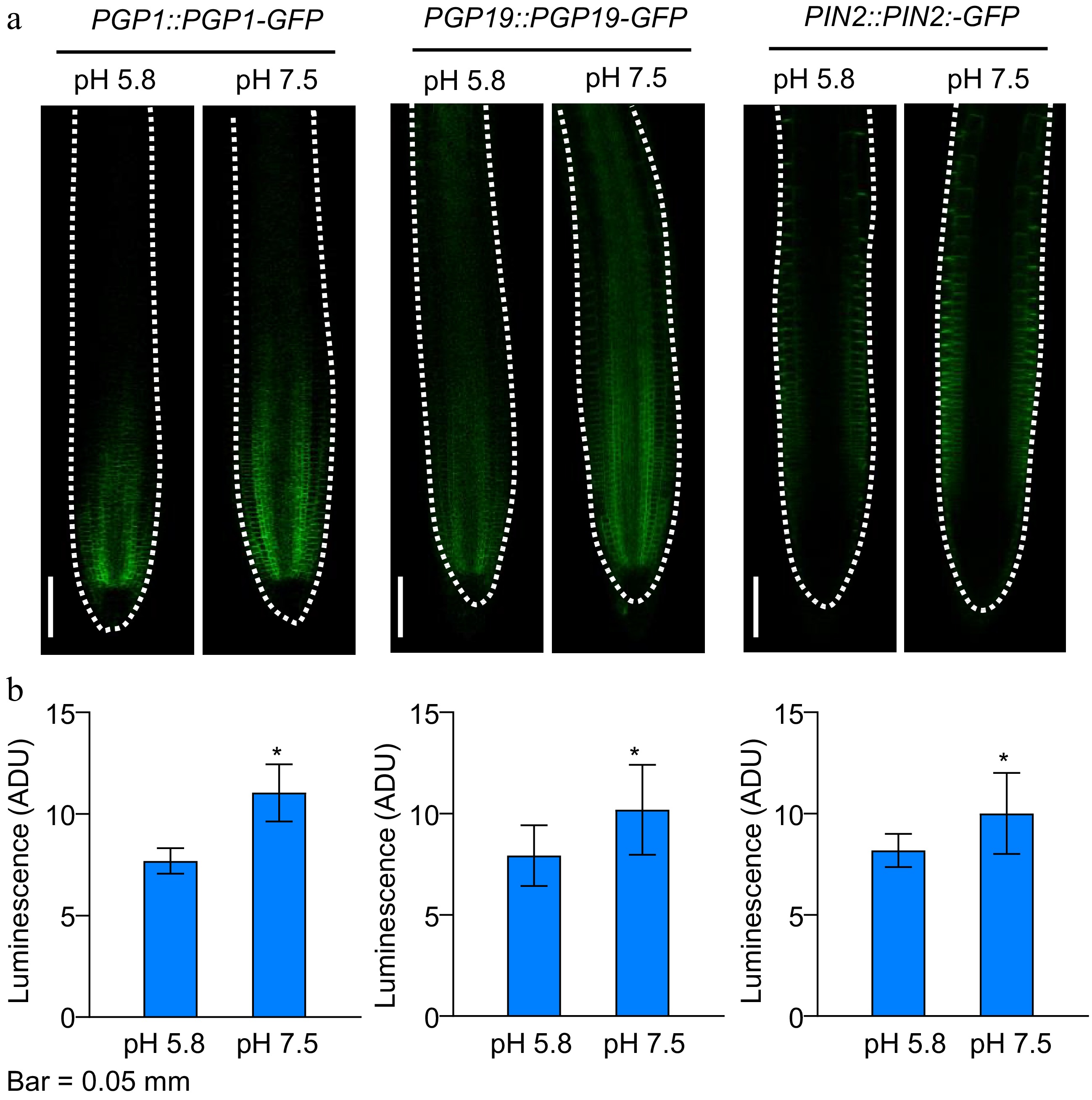
Figure 7.
Effect of slightly alkaline conditions on polar auxin transport in Arabidopsis. (a), (b) Localization and quantifications of PGP1-GFP, PGP19-GFP and PIN2-GFP signal in the root meristem zone of light-grown Col-0 seedlings. (Means ± SD, asterisks denote significant differences compared with control condition pH 5.8, * p < 0.05; ** p < 0.01, n ≥ 8). (Student t-test, Bar = 50 μm).
Transcriptome analysis dissected the mechanism of slightly alkaline pH promoting root development, which also indicated auxin signal pathway cascade and transporters were necessary for the procedure
-
Seedlings growing in standard pH medium were transferred into slightly alkaline conditions, and their root transcriptome dynamics were monitored at 0, 2 and 6 h later. There were 1,297 differentially expressed genes (DEGs) identified in the comparison of 0 h-VS-2 h, and 1,989 DEGs in that of 2 h-VS-6 h. KEGG enrichment analysis showed the DEGs of 0 h-VS-2 h, were mainly involved in 10 pathways as showed in Fig. 8a, and among them the pathway of plant hormone signal transduction was included. The DEGs involved in that pathway were mainly concerned with auxin. On the other hand, nitrogen metabolism, starch and sucrose metabolism were also activated, which also implied the promotion effect of pH 7.5.
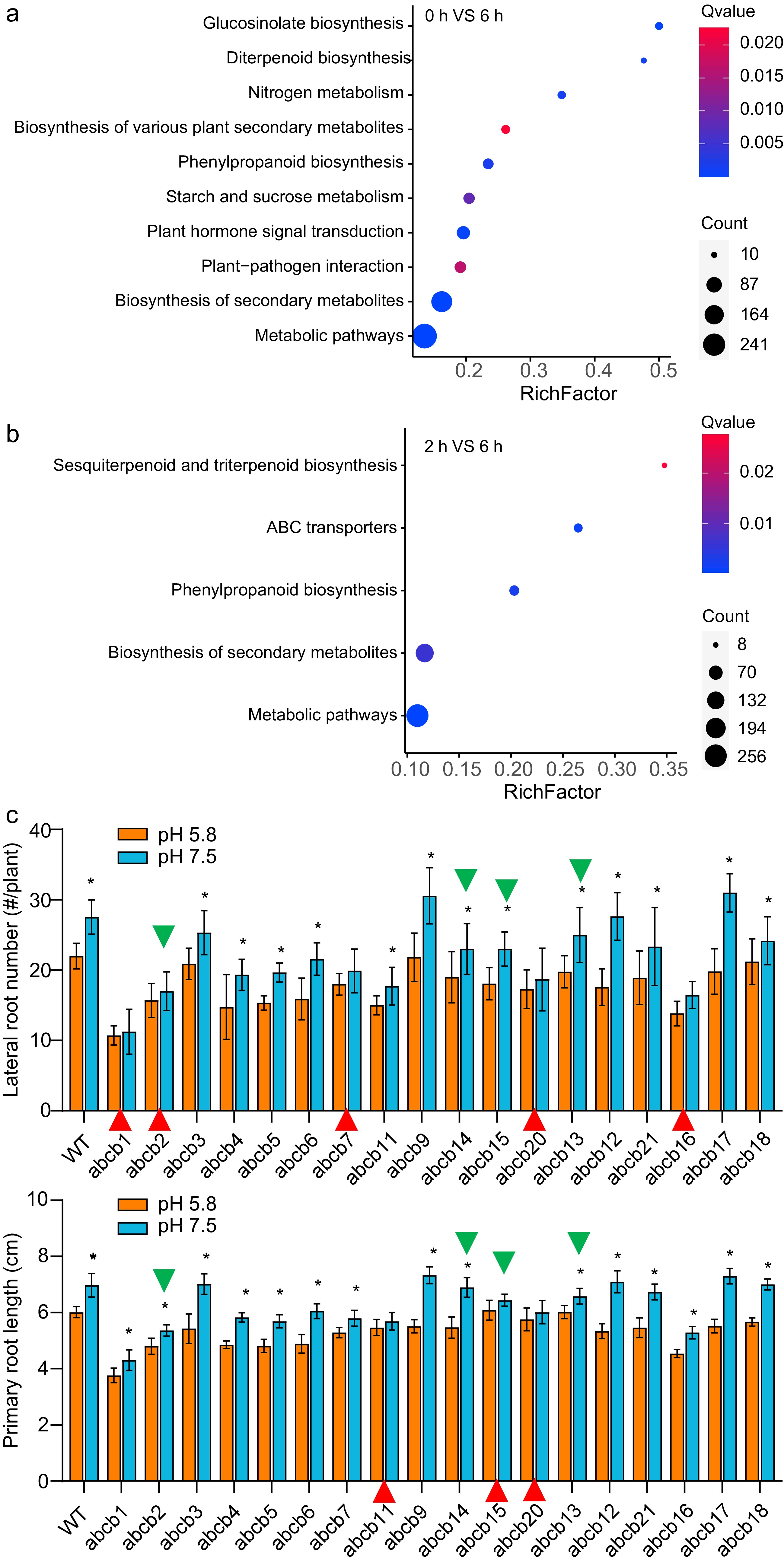
Figure 8.
Dynamic transcriptomic analysis on root tissue cultivated in pH 7.5 conditions. (a), (b), and the phenotype of ABC transporter gene mutants responding to (c) differing pH-value conditions. In (c), the green triangles showed the genes were DEGs of the 2h-VS-6h; the red triangles showed the mutants of which their primary root length or lateral root number did not response to pH 7.5.
KEGG enrichment analysis showed the DEGs of 2h-VS-6h were involved mainly in five pathways, and the pathway of ABC transporters were included. There were 18 ABC transporter family protein coding genes included.
Based on the result of the transcriptome above, all the mutants of the ABC transporter family in our germplasm collection were tested for their root phenotype in pH 5.8 and pH 7.5 cultivation. The primary root length and LR number were recorded in the same way as the experiments performed previously, the data are shown in the column chart of Fig. 8b. A total of 18 mutant germplasms were analyzed as shown in Fig. 8c and Supplemental Fig. S5. Interestingly, several of them showed reduced sensitivity to sightly alkaline conditions than WT. Mutant germplasms of abcb1, abcb2, abcb7, abcb16, and abcb20 did not show significant more LRs generated in pH 7.5 cultivation than the standard; while abcb11, abcd15 and abcb20 did not show significant longer primary roots in pH 7.5 than 5.8. Among them, the root development of abcb20 was null to pH variation. It is remarkable and worthy of note that although mutant of abcb20 block the promoting effect on the root development by slightly alkaline pH, without severe deformity as the double or triple mutants and the chemical inhibitors treatment, like sharp decrease or disappearing LR generation or loss of gravitropism.
-
Soil pH is a vital environmental factor influencing plant growth and development, especially for roots. In the present research, model plant Arabidopsis were cultivated in a standardized system to test a set of continuously altered pH conditions for their corresponding plant root development behaviors. The results showed neutral to slightly alkaline (7.0−8.0) pH could promote root development, in comparison with acidic (pH ≤ 5.8) or severe akaline (pH ≥ 9.0), which is consistent with a previous study (Gujas et al., 2012). Similar phenomena have been reported in crop cultivation in soil, for instance alkaline soil cultivated sugarbeet exhibited better root development in comparison with its cultivation in acidic soil (Geng et al., 2021); Bradutigan et al. have reported that when pea plant were cultivated in soil with a high pH (pH > 9.0), root growth was inhibited compared with the plants grown in pH 7.5 soil (Brautigan et al., 2012).
Our results revealed that slightly alkaline (7.5) pH cultivation could lead to the accumulation of auxin in the root apical meristem by affecting the auxin polar transport. In turn, it enhanced cycle gene expression and triggered the cell division activity at the meristem, leading to the promotion of PR elongation, and this procedure was independent from PLETHORA. Furthermore, the auxin signaling is accumulated in the oscillation zone. The high concentration of auxin thus induced the oscillation frequency and pre-branch site initiation, which resulted in more lateral root emergence. Thus, auxin accumulation in root apical meristem plays a key role in the molecular regulation mechanism of slightly alkaline pH promoting root development. Mutants of auxin transporter, biosynthesis as well as inhibitors were further engaged for the study, and the results showed that slightly alkaline pH promoting root development was mediated mainly by auxin transportation but not by auxin biosynthesis. PINs and ABCBs mediated auxin polar transport were essential for the accumulation of auxin in the root tip. Transcriptomic dynamic analysis showed pH 7.5 cultivation could trigger the changes in the pathways of plant hormones signal transduction and ABC transporter. The mutants of ABC transporter encoding genes were thus tested and the mutants of several ABC transporter members could partially block neutral and slightly alkaline promoting root development, when the root growth of abcb20 mutant was shown to be completely insensitive to neutral and slightly alkaline conditions. In summary, the present research showed neutral and slightly alkaline promoting Arabidopsis root development depended on auxin transportation in which the ABC transporter family were deeply involved, and ABCB20, ABCB1, ABCB2, ABCB7, ABCB11, ABCB15, ABCB16 play very important roles in the procedure.
This work was supported by the Project of Sanya Yazhou Bay Science and Technology City (No. SCKJ-JYRC-2022-21), the Fundamental Research Funds for the Central Universities (KYQN2023049 and KYT2023001), National Natural Science Foundation (No. 32202585), China Postdoctoral Science Foundation (2022M721641), and International Postdoctoral Exchange Fellowship Program (YJ20210263).
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Xingliang Duan, Long Luo
- Supplemental Fig. S1 Effects of acidic conditions on plant root growth. (a) plant growth status, (b) primary root length, lateral root number, lateral root density (means ± SD, asterisks denote significant differences compared with control condition (pH 5.8) at * p < 0.05; ** p < 0.01,n ≥ 8) (Student t-test, Bar = 1 cm ).
- Supplemental Fig. S2 Effect of slightly alkaline conditions on LRP of Arabidopsis. (a) Lateral root primordium of each stage, “×” indicate pericycle, (b) number of LRP+ LR+ emergence (means ± SD, asterisks denote significant differences between columns at * p < 0.05, ** p < 0.01, n ≥ 8) (Student t-test, Bar = 0.02 mm).
- Supplemental Fig. S3 Effects of alkaline conditions on Arabidopsis PLT mutants root growth. (a) plant growth status, (b) primary root length, lateral root number (means ± SD, asterisks denote significant differences compared with control condition (pH 5.8) at * p < 0.05; ** p < 0.01, n ≥ 8) (Student t-test, Bar = 1 cm ).
- Supplemental Fig. S4 Effects of alkaline conditions on the root growth of arabidopsis mutants ibr1ibr3ibr10 and ech2ibr1ibr3ibr10. (a) plant growth status, (b) primary root length, lateral root number (means ± SD, asterisks denote significant differences compared with control condition (pH 5.8) at * p < 0.05; ** p < 0.01, n ≥ 8) (Student t-test, Bar = 1 cm ).
- Supplemental Fig. S5 The mutant seedlings of ABC transporter coding genes cultivated in pH-5.8 and -7.5 conditions.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Duan X, Luo L, Wang Z, Wang W, Ye C, et al. 2023. Promotion of root development by slightly alkaline pH involves an auxin mediated adaption mechanism. Soil Science and Environment 2:6 doi: 10.48130/SSE-2023-0006 

Promotion of root development by slightly alkaline pH involves an auxin mediated adaption mechanism
- Received: 07 April 2023
- Accepted: 02 June 2023
- Published online: 07 July 2023
Abstract: The present study performed experiments to identify the superior pH for Arabidopsis root growth base on its development (radical root length and lateral root density); and then approached the mechanism by which optimized pH primes root development. The results showed that neutral to slightly alkaline pH (7.0−8.0) in medium was the optimum range in which the plant had longer primary roots and denser lateral roots. Auxin reporter DII:VENUS transgenic line cultured in standard (5.8) and slightly alkaline (7.5) pH indicated that in pH 7.5 conditions, there was less fluorescence in the root cap compared to in acidic conditions. Furthermore, the DR5:Luciferase transgenic plant showed pH 7.5 accelerated the auxin oscillation frequency, was 17.83% shorter than that in pH 5.8. Later, a series of mutant germplasms showed slightly alkaline conditions promoting root development were independent from PLT transcript factors, but mainly mediated by auxin transportation. Transcriptomic dynamic analysis showed pH 7.5 conditions could trigger the changes in the pathways of plant hormones signal transduction and ABC transporter. The mutants of the ABC transporter coding genes were thus tested and the results showed that mutant abcb20 could block the slightly alkaline (7.5) pH promoting root development completely, as well as several other mutants blocking it partially.
-
Key words:
- Root development /
- pH value /
- Auxin /
- Auxin transportation /
- ABC transporter