










-
After-ripening is a continuation of the seed maturation process. It is a common method used for dormancy release by regulating a series of physiological changes in after-ripened seeds[1,2]. Seeds that have undergone after-ripening could germinate rapidly and establish robust seedlings after sowing in fields. The status of germination and seedling in sowing under the fields has been significantly associated with the levels of dormancy release. The dormancy release during seed after-ripening mainly depends on the characteristics of the species, time, and environmental conditions of seed storage[3,4]. Rice (Oryza sativa L.) is one of the important food crops in China. An understanding of the physiological and molecular characteristics of rice after-ripening is important in evaluating seed vigor for sowing in practical agricultural production.
ABA and GA play crucial roles in seed dormancy and germination. ABA promotes seed dormancy and inhibits germination. Endogenous ABA content is modulated by ABA biosynthesis genes, such as 9-cis-epoxy-carotenoid dioxygenase (NCED), and catabolism genes, such as ABA-8’-hydroxylase[5]. GA breaks seed dormancy to promote germination, and its content is regulated by GA biosynthesis genes (GA20ox and GA3ox) and catabolism genes (GA2ox)[6]. Some reports showed that after-ripening-mediated dormancy release was tightly associated with the contents of ABA and GA[4,7−9]. It is well known that the levels of ABA decline (and GA increase) with the transition from dormant to non-dormant state during after-ripening in seeds of various species, such as tobacco, Arabidopsis, and barley[10−14]. In barley seeds, after-ripening promotes the expression of ABA catabolism gene HvABA8’OH1 and GA biosynthetic gene HvGA3ox2 following imbibition[15]. In rice, GA1/ABA, GA7/ABA, GA12/ABA and GA20/ABA ratios significantly increased in Jiucaiqing imbibed seeds following 3 months of after-ripening[16]. However, the precise changes of ABA and GA content in after-ripened rice seeds remains unclear.
Reactive oxygen species (ROS), including H2O2, regulate seed development, dormancy release, and germination as the redox signaling molecules in several plants[17,18]. It has been demonstrated that exogenous H2O2 could break barley seed dormancy[19]. H2O2 accumulation in seeds has been closely associated with the breaking of sunflower embryo dormancy during after-ripening[18]. The accumulation of H2O2 was also observed in seed after-ripening in Fagus sylvatica, wheat, and Arabidopsis[4,20−22]. ROS levels in plants undergo strict control to fulfill their roles as cellular messengers, a process which involves an efficient antioxidant system[23], e.g., SOD, catalase (CAT), and APX. SOD converts superoxide radicals into H2O2, and then H2O2 is degraded into water and molecular oxygen by two key enzymes, CAT and APX[24,25]. In maize, an increase in the enzymatic activities of SOD in developing and germinated seeds was observed[26]. In rice, a slight increase in the H2O2 level and a significant increase in the activities of SOD, CAT, and APX in imbibed seeds after cold stratification were observed[27]. These indicated that ROS could take a pivotal role in rice seed after-ripening.
Seed dormancy is complex and can be released by after-ripening[28,29]. Recently, some genes in rice have been reported to regulate seed dormancy. Overexpression of two SPL (SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE) genes, SPL12 and IPA1, enhanced seed dormancy and inhibited pre-harvest sprouting (PHS) without noticeable effects on shoot architecture in rice[30]. OsDOG1L-3, encoding a DELAY OF GERMINATION 1 (DOG1) like protein, was reported to regulate rice seed dormancy through the ABA pathway[31]. It was reported that suppression of rice gene GERMIN-LIKE PROTEIN 2-1 (OsGLP2-1) expression resulted in the release of dormancy in immature seeds; and overexpression of OsGLP2-1 led to greater seed dormancy[32]. A 1 bp deletion in Os02g0202400 (OsBT1) was shown to regulate seed dormancy through the glycometabolism pathway in rice[33]. Xu et al. identified 2,169 up-regulated (and 1,271 down-regulated) genes using RNA sequencing (RNA-seq) between the freshly harvested seeds and the after-ripened seeds in peanuts, which were enriched in oxidative phosphorylation, glutathione metabolism, and carbon metabolism[34]. A transcriptome profiling in Arabidopsis identified after-ripening-specific gene sets, including 19 up-regulated genes of after-ripening and 103 down-regulated genes of after-ripening[35]. Nevertheless, there are still few known specific genes related to rice after-ripening, and the same is true of their molecular mechanisms.
In our previous study, we analyzed the physiological and gene expression changes in imbibed seed regulated by after-ripening[16], but the detailed about the changes in the dry seeds controlled by after-ripening remain unknown. Herein, we assessed the characteristics of seed dormancy with seed germination testing during after-ripening in rice. Additionally, we investigated the contents of ABA, GA, and H2O2, as well as antioxidant enzyme and α-amylase activities in Nipponbare dry seeds during after-ripening. We used RNA-seq to investigate transcriptional changes between NPB dry seeds at 0 and 28 DAR, and several differential transcripts (DTs) were identified as rice after-ripening-related genes. Our results helpfully illustrated the role of after-ripening in dormancy release and identified potential genes associated with after-ripening in rice.
-
The japonica Nipponbare (NPB), Nanjing 9108 (NJ 9108), Wuyunjing 7 (WYJ 7), and Zhendao 88 (ZD 88), were used in this study along with indica 9311. All these rice varieties are commercially used in rice production in the Yangtze River basin. The 25-day-old seedlings of ten rows were transplanted into a paddy field at the Experimental Station of Nanjing Agricultural University (Nanjing, Jiangsu Province, China; E 118°509, N 32°029) on June 17th, 2021, with 17 cm between plants within a row and 33 cm between rows. Filed management was carried out according to local common methods[36]. The heading date of five rice varieties, NPB, NJ 9108, WYJ 7, ZD 88 and 9311 were on August 10th, 27th, 27th, 25th, and 21th, respectively. Plants with the same heading date were tagged when the primary panicle emerged from the leaf sheath[37 ]. The tagged panicles were harvested 5 weeks after flowering. All harvested seeds had showed maturing. Freshly harvested seeds (0 DAR) were stored in room conditions at 25 °C for after-ripening. In the preliminary experiment, NPB seeds at 0, 14, 28, 42, and 56 DAR were used for seed germination. It was found that the seed dormancy was gradually released with after-ripening and had strong dormancy (SP ≈ 0%) at 0 DAR, moderate dormancy (SP ≈ 65%) at 28 DAR, and weak or non-dormancy (SP ≈ 98%) at 56 DAR. In order to reflect the regulation of after-ripening, seeds at the 0, 28 and 56 DAR instead of a timecourse were selected and frozen in liquid nitrogen to store in −80 °C refrigerator for other experiments. The water contents of NPB dry seeds at 0, 28, and 56 DAR were 13.97%, 12.07%, and 9.74%, respectively.
Seed germination
-
Thirty seeds of each sample were imbibed in Petri dishes (diameter 9 cm) with 10 mL distilled water at 25 ± 1 °C for 10 d. The number of germinated seeds were recorded every day. Seeds were considered as germinated when the radicle protruded (2 mm) through the seed coat. Seedlings were considered as established when the root length reached seed length and the shoot length reached half of the seed length. Germination percentage (GP) = (Total germinated seeds/30) × 100%; seedling percentage (SP) = (Total seedling seeds/30) × 100%. Germination index (GI) was calculated as follows: GI = ∑(Gt/Dt), where Gt is the number of the germinated seeds on Day t[38]. Three biological replicates were conducted for each sample.
Hormone quantification
-
Ten NPB dry seeds were sampled for hormone quantification at 0, 28, and 56 DAR. The contents of endogenous ABA and GAs were detected by MetWare (Wuhan Metware Biotechnology Co., Ltd., Wuhan, China,
www.metware.cn ) based on the AB Sciex QTRAP 6500 LC-MS/MS platform. The contents of ABA and GAs were expressed as ng·g−1 FW (fresh weight). Three biological replicates were conducted for each sample.Evaluation of H2O2 levels
-
The NPB dry seeds were sampled at 0, 28, and 56 DAR to measure the H2O2 levels using the methods described according to Hydrogen Peroxide Content Assay Kit following the manufacturer’s instructions (Beijing Solarbio Science & Technology Co., Ltd, Beijing, China). Three biological replicates were conducted for each sample.
Evaluation of antioxidant enzyme activities
-
The NPB dry seeds were sampled at 0, 28, and 56 DAR to measure the activities of SOD, CAT, and APX using commercial assay kits, following the manufacturer’s instructions (Beijing Solarbio Science & Technology Co., Ltd, Beijing, China). The kits included Superoxide Dismutase Activity Assay Kit, Catalase Activity Assay Kit, and Ascorbate Peroxidase Activity Assay Kit. Three biological replicates were conducted for each sample.
Determination of α-amylase activity
-
The NPB dry seeds were sampled at 0, 28, and 56 DAR to measure the activities of α-amylase using the methods described, according to α-amylase Activity Assay Kit, following the manufacturer's instructions (Beijing Solarbio Science & Technology Company, Beijing, China). Three biological replicates were conducted for each sample.
RNA-seq analysis
-
The NPB dry seeds were collected at 0 and 28 DAR. The extraction and examination of total RNA, library construction, and Illumina sequencing were conducted by Personal Biotech (Shanghai, China) using the Illumina novaseq pe150. The clean reads were aligned with reference sequences of rice in IRGSP-1.0 (
http://rapdb.dna.affrc.go.jp/download/irgsp1.html ). The transcript levels were calculated with Cufflinks and normalized by the fragments per kilobase of transcript per million mapped reads (FPKM) method. DETs were defined by fold change ≥ 2 and P-adjust < 0.05. The top DETs were further selected as FPKM ≥ 10 in NPB seeds at 0 or 28 DAR. Annotation of gene function was performed by comparison with the information in the NCBI database (www.ncbi.nlm.nih.gov ). Three biological replicates were conducted for each sample.Analysis of transcript levels
-
Total RNA in the whole dry seeds was extracted using the RNAprep Pure Plant Kit (Tiangen Biotech Co., Ltd, Beijing, China). Approximately 400 ng of total RNA was transcribed into cDNA using the HiScript® II Reverse Transcriptase system (Vazyme Biotech Co., Ltd, Nanjing, China). The real-time quantitative PCR (RT-qPCR) was carried out in a total volume of 10 μL (template cDNA: 1 μL, forward primer/reverse primer: 0.5 μL/0.5 μL, SYBR Green qPCR Master Mix: 5 μL, ddH2O: 3 μL) using the Roche LightCycler480 Real-time System (Roche, Swiss Confederation) with the rice OsActin (LOC_Os03g50885) as an internal control. The PCR conditions were as follows: 95 °C for 5 min, 40 cycles of 95 °C for 10 s and 60 °C for 30 s, 95 °C for 15 s, 60 °C for 60 s, 95 °C for 15 s. The transcript levels of ten randomly selected DTs (five up-regulated and five down-regulated transcripts) in NPB seeds at 0 and 28 DAR, and of the four top DTs (the decreased OsOxO1 and OsNCED4 and the increased RSOsPR10, and OsPR10a) and OsNECE4 in all the five varieties of seeds at 0, 28, and 56 DAR, were measured. Primers for RT-qPCR were designed using PrimerQuest Tool[39] and are listed in Supplemental Table S1. Levels of gene expression were calculated using the comparative CT method[40]. Three biological replications were performed for each sample.
Data analysis
-
Experimental data were analyzed using the SAS software (Cary, NC, USA). The significant differences were tested using Tukey’s test at 5% levels of probability.
-
To analyze the characteristics of after-ripening, the seed germination and seedling emergence were evaluated by using NPB seeds at 0, 28, and 56 DAR. Seed germination results showed that there were extremely low germination and seedling emergence levels in freshly harvested seeds (0 DAR), with 10% of germination percentage (GP), 6% of seedling percentage (SP), and 0.3 of germination index (GI) at 10 d of imbibition (Fig. 1). This suggested that there was a deep dormancy in freshly harvested seeds of NPB. The GP, SP and GI of NPB seeds at 28 DAR were significantly increased, with 92%, 67%, and 4.0 at 10 d of imbibition respectively, indicating there was a partial released of seed dormancy levels (Fig. 1). The GP, SP, and GI of the NPB seeds at 56 DAR were 98%, 98%, and 5.2 at 10 d of imbibition respectively (Fig. 1). The results suggested that the seed dormancy releases of NPB seeds were nearly completed at 56 DAR.
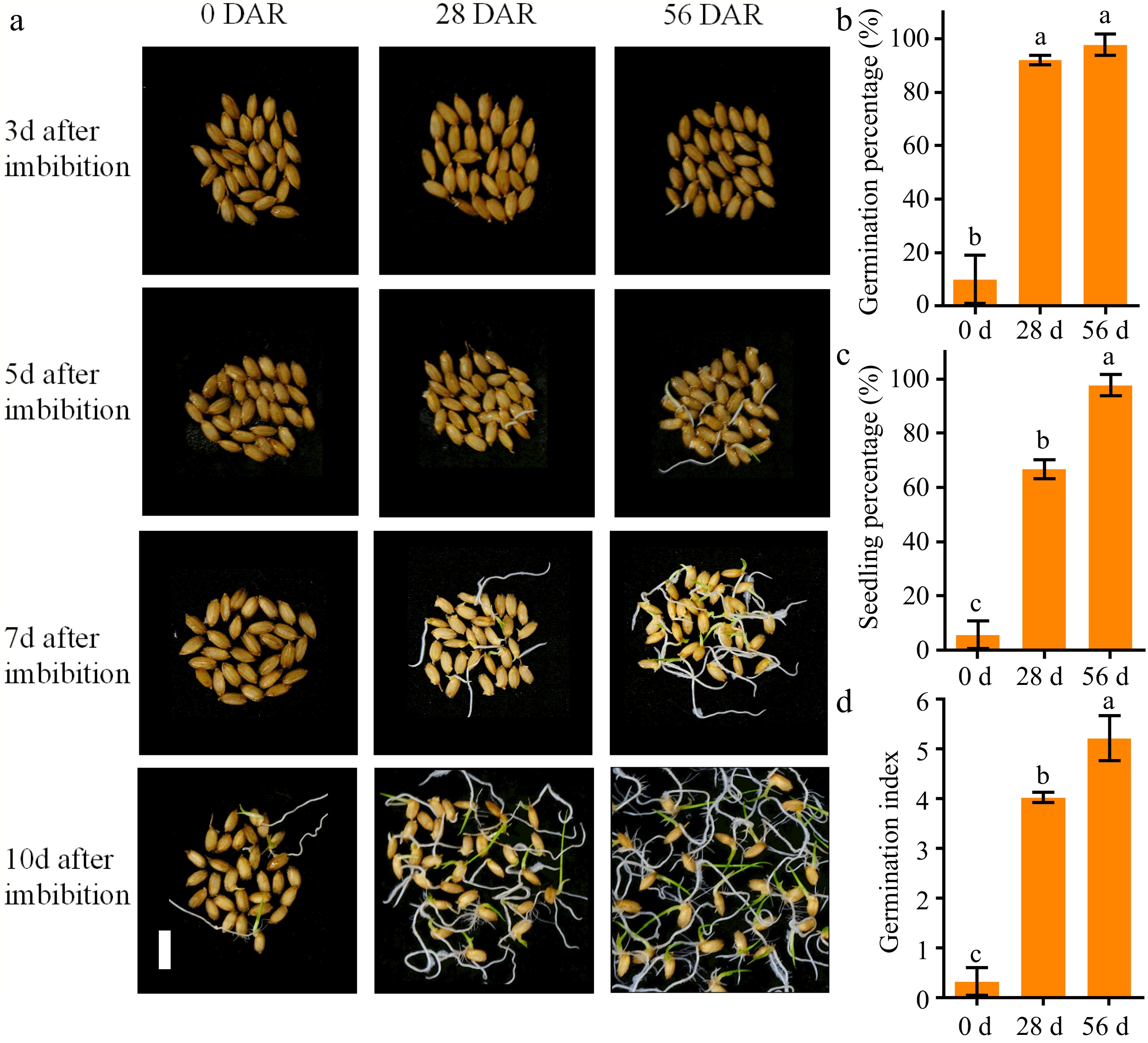
Figure 1.
Changes in seed germination and seedling emergence in Nipponbare seeds at 0, 28, and 56 DAR. (a) Phenotypic graphics of seed germination at 3, 5, 7 and 10 d of imbibition in NPB seeds at 0, 28 and 56 DAR. (b) Comparison of germination percentage, (c) seedling percentage and (d) germination index at 10 d of imbibition in NPB seeds at 0, 28, and 56 DAR. NPB represents Nipponbare. DAR represents day after-ripening. Bars = 1 cm. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
Changes of GA and ABA levels during after-ripening
-
Several studies revealed that ABA and GA contents were vital for after-ripening of seeds[5−7]. We measured the contents of ABA and GA in the dry seeds of NPB at 0, 28, and 56 DAR. The results showed ABA content in NPB seeds at both 28 and 56 DAR significantly decreased, compared to freshly harvested seeds (0 DAR), while there was no significant difference between 28 DAR and 56 DAR seeds (Fig. 2a). Additionally, there were four kinds of GAs identified in NPB seeds at 0, 28, and 56 DAR, including bioactive GA1 and inactive GA9, GA24, and GA53. Compared to 0 DAR seeds, the contents of GA9 and GA24 in 28 and 56 DAR seeds significantly increased, while GA1 and GA53 in 28 and 56 DAR seeds, respectively, did not display significant differences (Fig. 2b). GA1, GA9, and GA53 showed no significant changes between in 28 and 56 DAR seeds, except GA24 (Fig. 2b). Meanwhile, we analyzed GA/ABA ratios of after-ripened seeds and found that GA1/ABA, GA9/ABA, GA24/ABA, and GA53/ABA significantly increased with the time of after-ripening (Table 1). These results suggest that ABA content was degraded in rice after-ripened seeds, bioactive GA1 content did not accumulate.
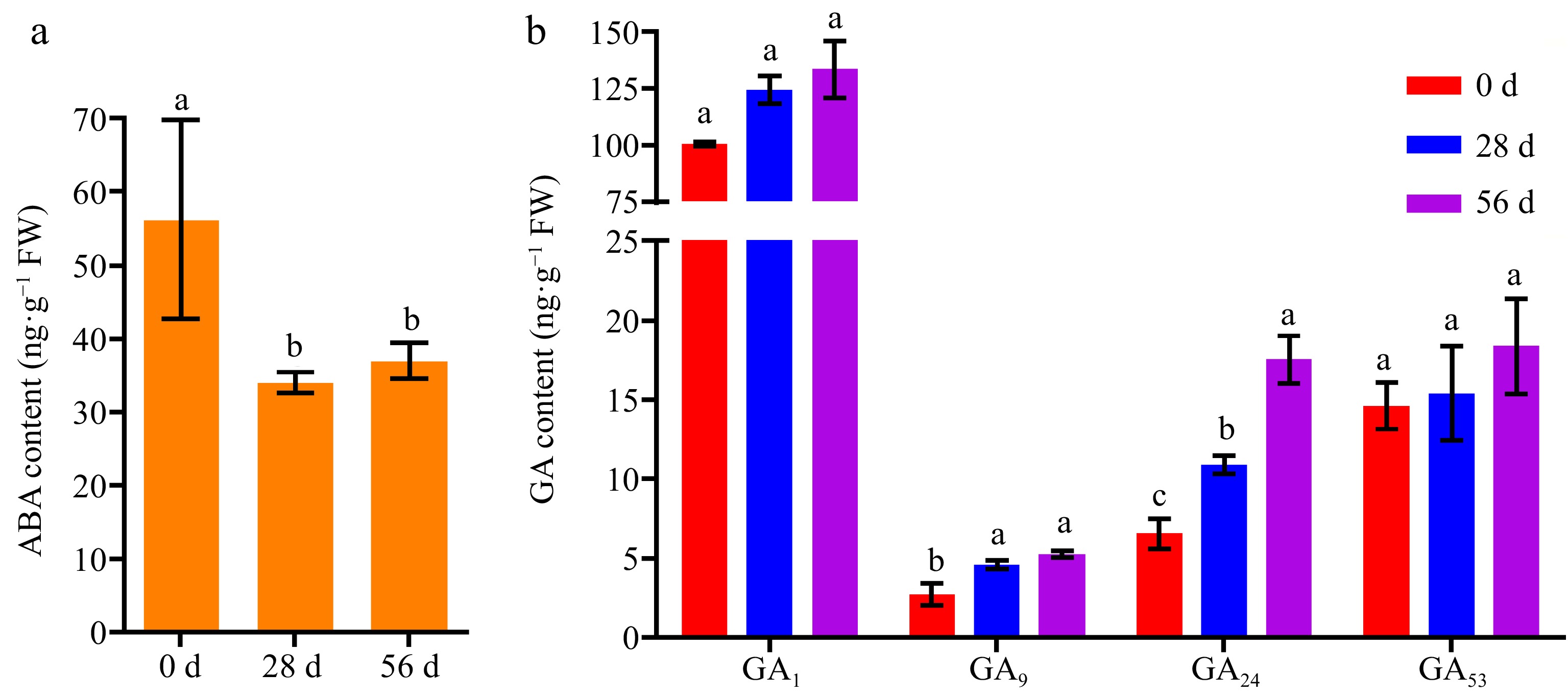
Figure 2.
Contents of ABA and GAs in Nipponbare seeds at 0, 28, and 56 DAR. (a) Content of ABA in Nipponbare seeds at 0, 28, and 56 DAR. (b) Content of GAs in NPB seeds at 0, 28, and 56 DAR. NPB represents Nipponbare. Different lower case letters above columns indicate statistical differences at P < 0.05.
Table 1. GA/ABA ratios in 0, 28, and 56 DAR NPB seeds.
After-ripening (d) GA1/ABA GA9/ABA GA24/ABA GA53/ABA 0 1.9085c 0.0584c 0.1166c 0.2603c 28 3.2926b 0.1353b 0.3206b 0.4531b 56 3.4208a 0.1430a 0.4191a 0.4966a Different lower case letters above columns indicate statistical differences at P < 0.05. Changes in α-amylase activity during after-ripening
-
Amylase is involved in starch degradation and regulation of seed germination[ 41]. We measured α-amylase activities in NPB dry seeds at 0, 28, and 56 DAR. The results showed that the activity of α-amylase significantly increased, from ~3 U·g−1 in 0 DAR seeds to ~8 U·g−1 in 28 DAR seeds, and then slightly decreased to ~6 U·g−1 in 56 DAR seeds (Fig. 3). There were no significant differences in the activity of α-amylase between 28 DAR seeds and 56 DAR seeds.
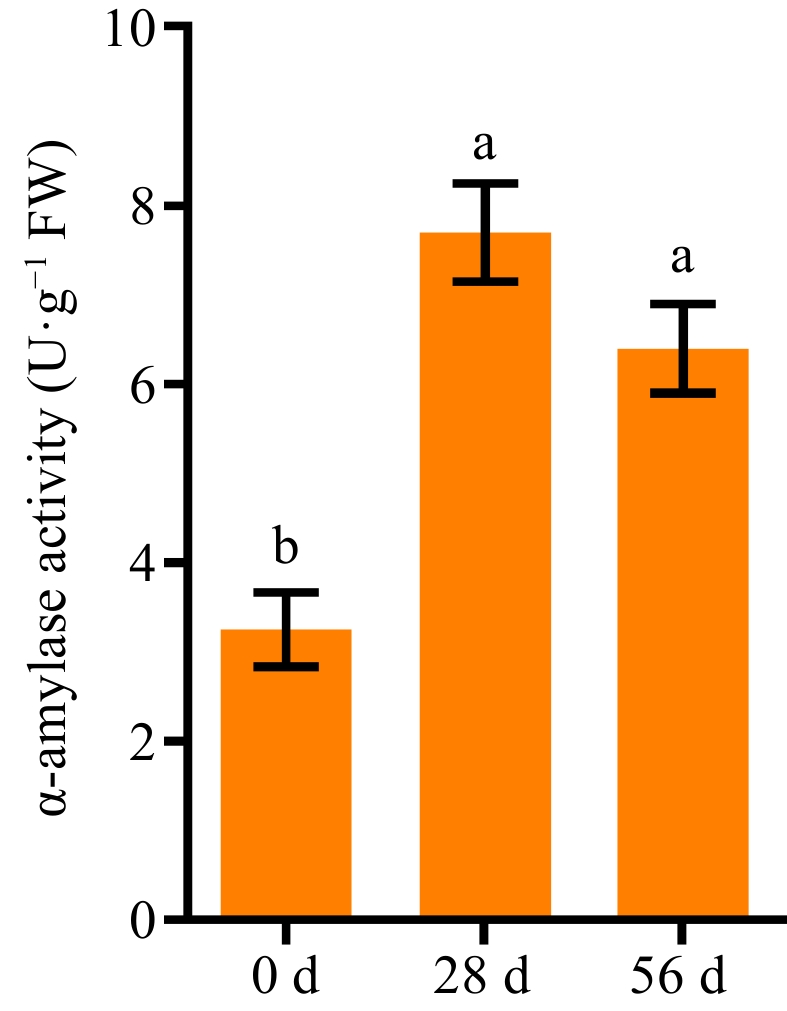
Figure 3.
The activity of α-amylase in Nipponbare seeds at 0, 28, and 56 DAR. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
Changes of H2O2 level and antioxidant enzyme activities during after-ripening
-
As a signaling molecule, H2O2 is well-known to be involved in seed germination and seed vigor[19−21]. We measured the content of H2O2 and its related enzyme activities in NPB dry seeds at 0, 28, and 56 DAR. The results showed that the H2O2 levels significantly increased, from ~3 μmol·g−1 in 0 DAR seeds to ~15 μmol·g−1 in 28 DAR seeds, and then decreased to ~8 μmol·g−1 in 56 DAR seeds (Fig. 4a). The emzymatic activities of SOD significantly increased, from ~261 nmol·min−1·g−1 in 0 DAR seeds to ~413 nmol·min−1·g−1 in 28 DAR seeds, and then decreased to ~209 nmol·min−1·g−1 in 56 DAR seeds (Fig. 4b). This was very similar to the tendencies observed in H2O2 levels during after-ripening of seeds. The activities of CAT significantly increased, from ~94 nmol·min−1·g−1 in 28 DAR seeds to ~116 nmol·min−1·g−1 in 56 DAR seeds, while there were no observed changes between 0 and 28 DAR seeds (Fig. 4c). The activities of APX had no significant changes among 0, 28, and 56 DAR seeds (Fig. 4d). These results suggested that the degradation of H2O2 contents in 56 DAR seeds might be due to the increase in CAT activities.
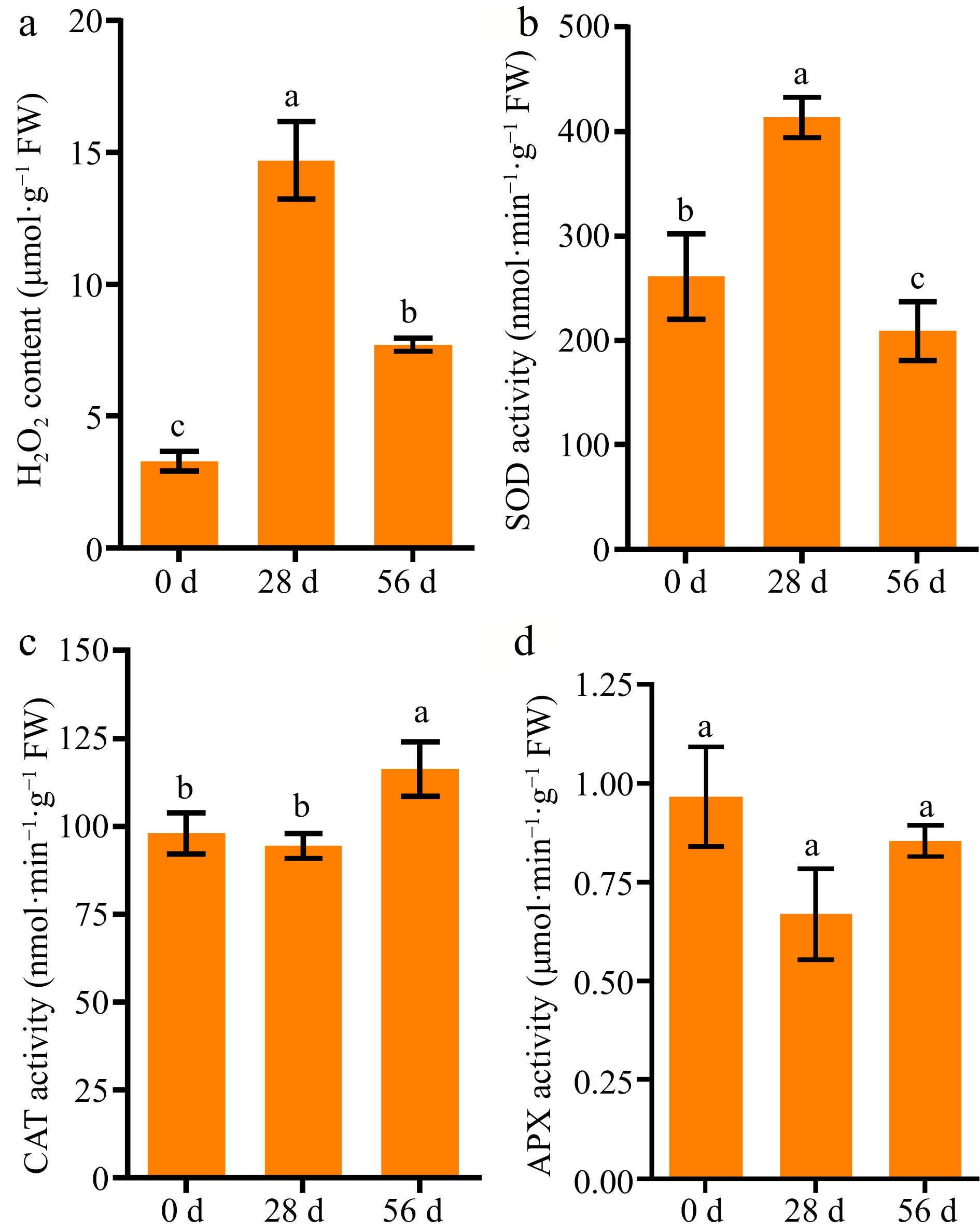
Figure 4.
Comparison of H2O2 level and antioxidant enzyme activities in Nipponbare seeds at 0, 28, and 56 DAR. (a) H2O2 level, (b) SOD, (c) CAT, and (d) APX activities in NPB seeds at 0, 28, and 56 DAR. NPB represents Nipponbare. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
Analysis of differential transcripts by RNA-seq during after-ripening
-
To identify the genes related to seed after-ripening in rice, we performed RNA-seq using 0 and 28 DAR NPB dry seeds. A total of 149 DTs, including 45 up-regulated and 104 down-regulated genes (Supplemental Table S2, Fig. 5a), were identified in 28 DAR seeds, as compared to 0 DAR seeds (Fold Change ≥ 2 and P-adjust < 0.05). Ten DTs, including five up-regulated and five down-regulated transcripts, were randomly selected for their transcript levels by RT-qPCR in NPB seeds at 0 and 28 DAR. It was observed that there were consistent trends for those DTs using RNA-Seq and RT-qPCR in 0 and 28 DAR NPB seeds (Supplemental Fig. S1).
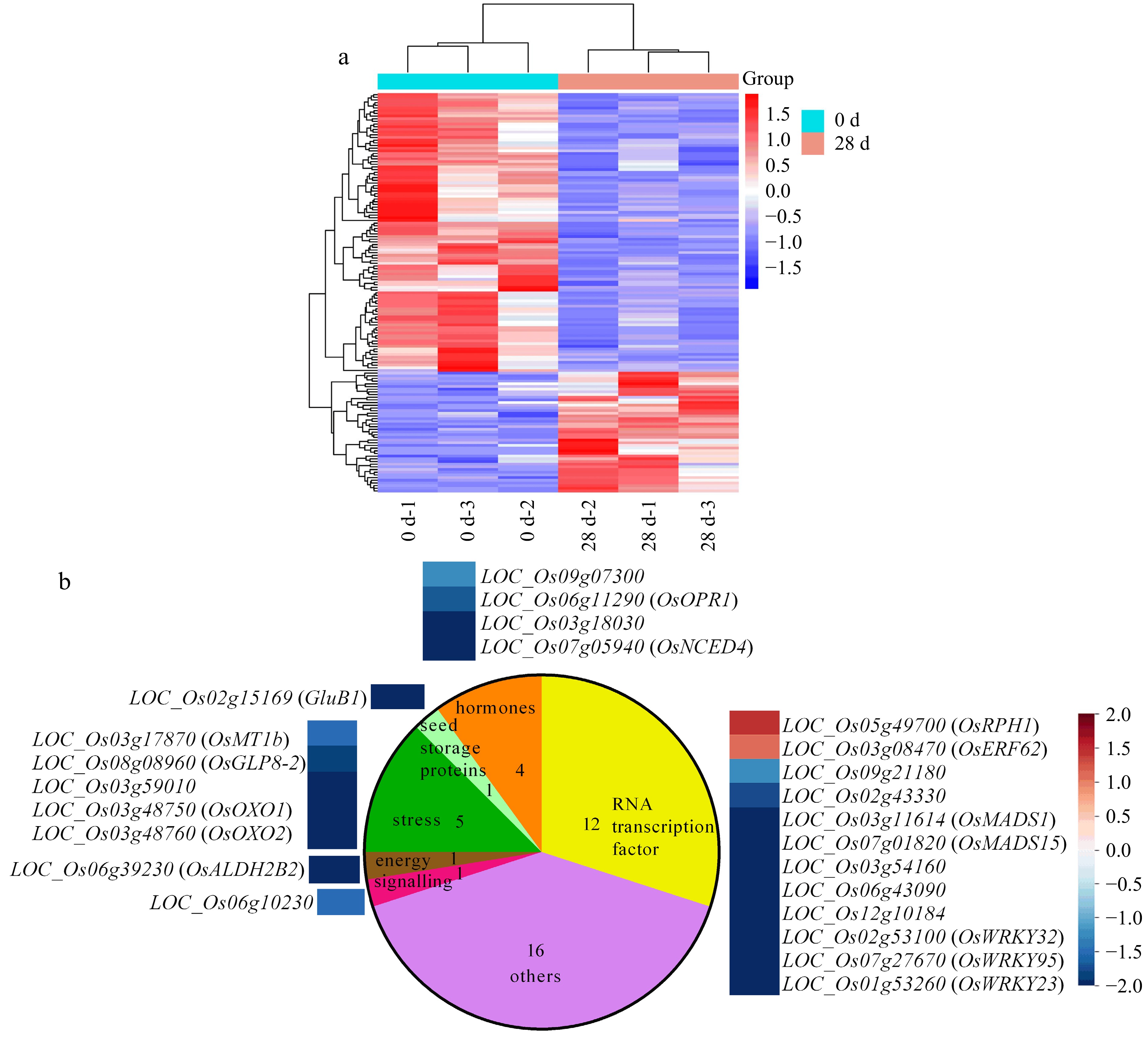
Figure 5.
Identification and analysis of differential transcripts between 0 and 28 DAR Nipponbare dry seeds via RNA-seq. (a) Hierarchical clustering of the DTs between 0 and 28 DAR NPB seeds; scale bar shows fold changes, while the red and the blue colors indicate up-regulation and down-regulation, respectively. (b) Enriched pathways of seed molecular networks, elucidated via Mapman software, including hormones, seed storage proteins, stress, energy, signaling, and RNA transcription factor. Red: up-regulation; blue: down-regulation.
Among 149 DTs identified by the RNA-Seq, 40 DTs were significantly enriched in the seed molecular networks pathway using Mapman software (Fig. 5b), which may be involved in regulation of rice seeds. The pathways involved in seed molecular networks were classified into seven functional groups, including hormones (4), seed storage proteins (1), stress (5), energy (1), signaling (1), RNA transcription factor (12), and others (16) (Fig. 5b). These 40 DTs and their pathways might be tightly associated with seed after-ripening in rice. Interestingly, of the hormones, OsNCED4, encoding ABA biosynthesis enzyme 9-cis-epoxycarotenoid dioxygenase, was also detected.
Validation of the key transcripts of after-ripening seeds
-
To validate the key transcripts involved in seeds after-ripening, we further compared the characteristics of seed dormancy releases of other four rice varieties, Nanjing 9108 (NJ 9108), Wuyunjing 7 (WYJ 7), Zhendao 88 (ZD 88), and 9311 during after-ripening of dry seeds at 0, 28, and 56 DAR. The results showed that the GP and SP (at 10 d of imbibition) and GI, in NJ 9108, gradually and significantly increased from 0 and 56 DAR seeds (Fig. 6a−d), consistent with NPB. In WYJ 7, ZD 88, and 9311, the GP and SP showed no significant differences at 10 d of imbibition between 28 and 56 DAR seeds (Fig. 6e−p), with the exception of GI, suggesting that the levels of seed dormancy of these three varieties were lower than that of NJ9108 and NPB. It could also mean that the seed dormancy of the WYJ 7, ZD 88, and 9311 were released earlier than NJ9108 and NPB during after-ripening.
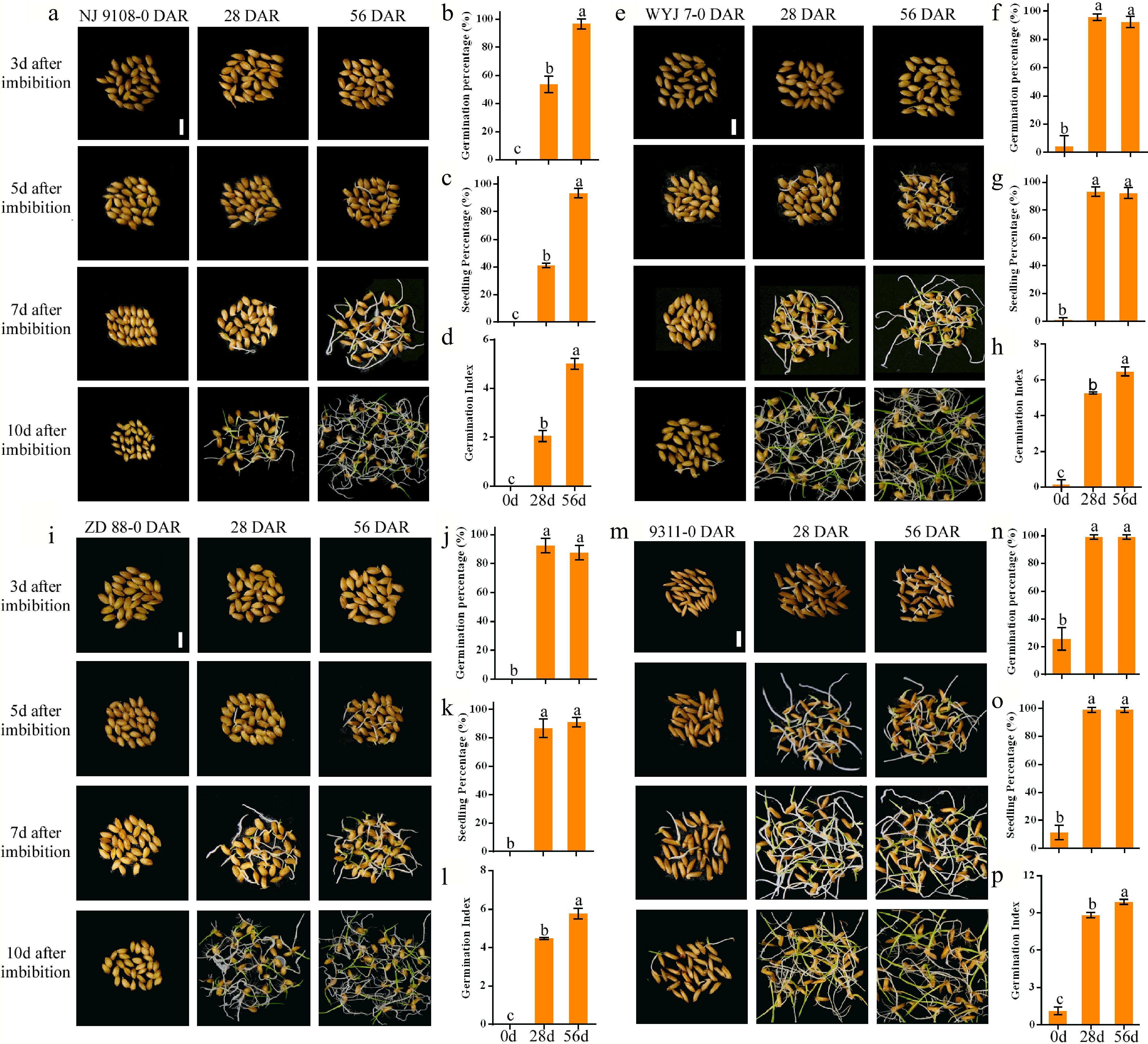
Figure 6.
Changes of seed germination and seedling emergence in NJ 9108, WYJ 7, ZD 88, and 9311 seeds at 0, 28, and 56 DAR. Phenotypic graphics of seed germination at 3, 5, 7, and 10 d of imbibition in (a) NJ 9108, (e) WYJ 7, (i) ZD 88, and (m) 9311 at 0, 28, and 56 DAR. Comparison of germination percentage, seedling percentage, and germination index at 10 d of imbibition in 0, 28, and 56 DAR (b)−(d) NJ 9108, (f)−(h) WYJ 7, (j)−(l) ZD 88, and (n)−(p) 9311 seeds. Bars = 1 cm. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
According to RNA-seq results, the four top DTs, RSOsPR10 (LOC_Os12g36830), OsDTC1 (LOC_Os02g36140), OsPR10a (LOC_Os12g36880), and OsOxO1 (LOC_Os03g48750), were selected as candidate key transcripts for after-ripening, based on the Fold Change and FPKM. Therefore, we measured the transcript levels of the five DTs, including four top DTs and OsNCED4, by RT-qPCR approach in 0, 28, and 56 DAR dry seeds of the five rice varieties. The results showed that the transcript levels of RSOsPR10 and OsPR10a were gradually and significantly up-regulated and the transcript levels of OsOxO1 and OsNCED4 were gradually and significantly down-regulated with after-ripening in NPB and NJ 9108 seeds (Fig. 7a, b), especially from 0 to 28 DAR. The transcript levels of RSOsPR10 and OsPR10a in WYJ 7 and ZD 88 seeds, respectively, as well as RSOsPR10 in 9311 seeds, significantly increased at 28 DAR and then significantly decreased at 56 DAR. Meanwhile, the transcript levels of OsOxO1 in WYJ 7, ZD 88, and 9311 seeds, as well as OsNCED4 in WYJ 7 and ZD 88 seeds, significantly decreased at 28 and 56 DAR (Fig. 7c−e). There were no significant differences in the transcript levels of OsOxO1 between 28 and 56 DAR in the five variety seeds, of OsNCED4 in NJ 9108, WYJ 7, ZD 88, and 9311 seeds, and of OsPR10a in 9311 seeds (Fig. 7). The transcript levels of OsDTC1 were gradually and significantly down-regulated with after-ripening in NPB and ZD 88, significantly increased at 28 DAR and then significantly decreased at 56 DAR in NJ 9108 and 9311, and no changes between 0 and 28 DAR in WYJ 7 (Fig. 7). All these data suggested that RSOsPR10, OsPR10a, OsOxO1, and OsNCED4 transcript levels might be associated with the degree of seed dormancy release in after-ripening and had important roles in rice seed after-ripening.
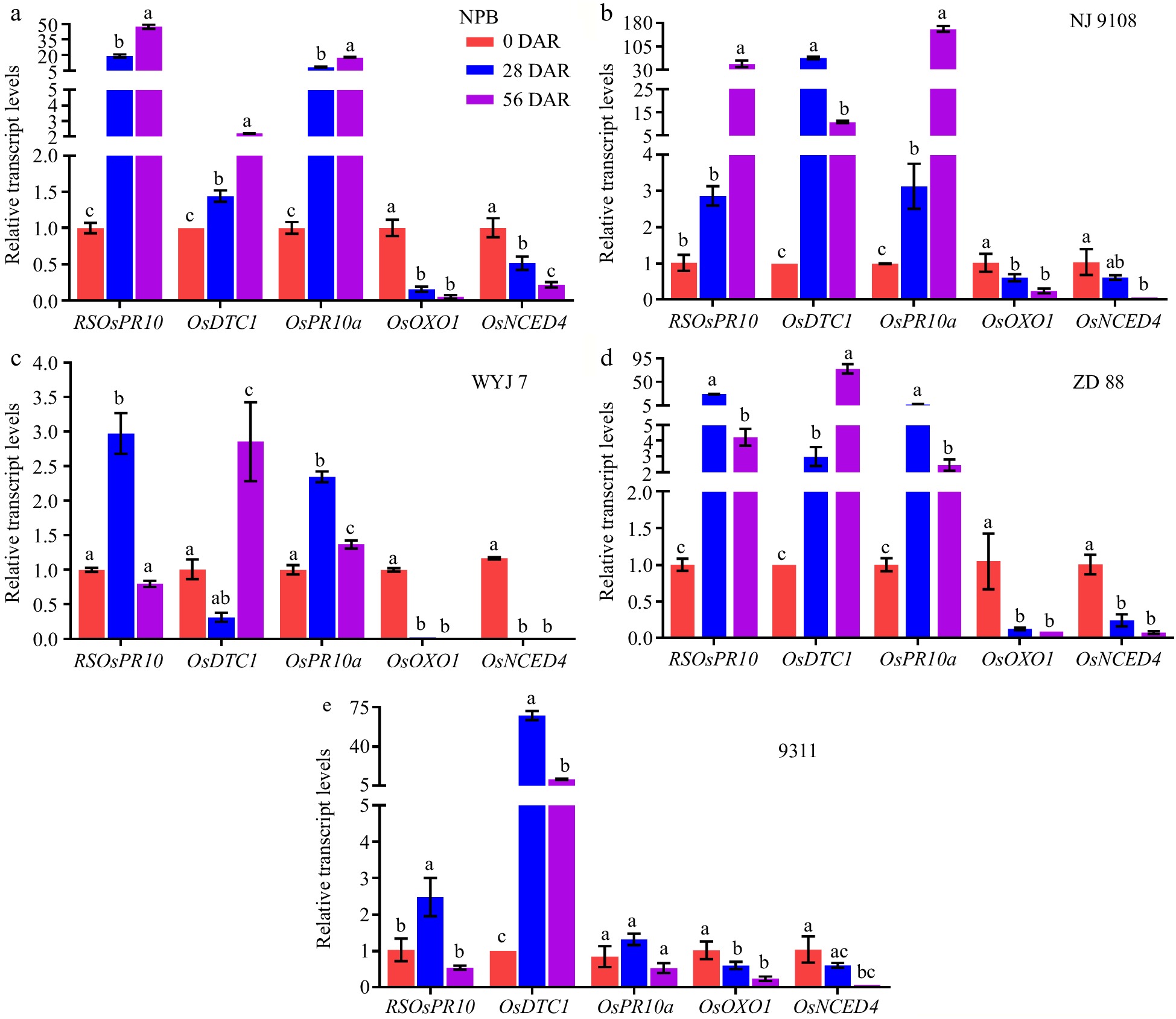
Figure 7.
Transcript levels of key genes related to after-ripening of seeds in five rice cultivars. Changes of transcript levels of RSOsPR10, OsDTC1, OsPR10a, OsOxO1, and OsNCED4 in (a) NPB, (b) NJ 9108, (c) WYJ 7, (d) ZD 88, and (e) 9311 seeds at 0, 28, and 56 DAR. The transcript levels of genes were normalized to that of OsActin. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
Analysis of ABA and GA- related to transcripts during after-ripening
-
Given the ABA biosynthesis OsNCED4 identified by RNA-seq, we further measured the transcript levels of ABA and GA metabolism- related to genes in NPB dry seeds at 0, 28, and 58 DAR using the RT-qPCR. Compared to 0 DAR seeds, the transcript levels of the ABA biosynthesis genes OsNECD1 significantly decreased in NPB seeds at 28 and 56 DAR, similar to that of OsNCED4. Meanwhile, the catabolism genes CYP707A5 significantly increased in 28 and 56 DAR seeds, while CYP707A6 increased in only 56 DAR seeds (Fig. 8a). Compared to 28 DAR seeds, the transcript levels of the OsNECD1 and CYP707A6 significantly increased in 56 DAR seeds (Fig. 8a). The changes in transcript levels of these genes, related to ABA metabolism, were consistent with the changes of ABA contents.
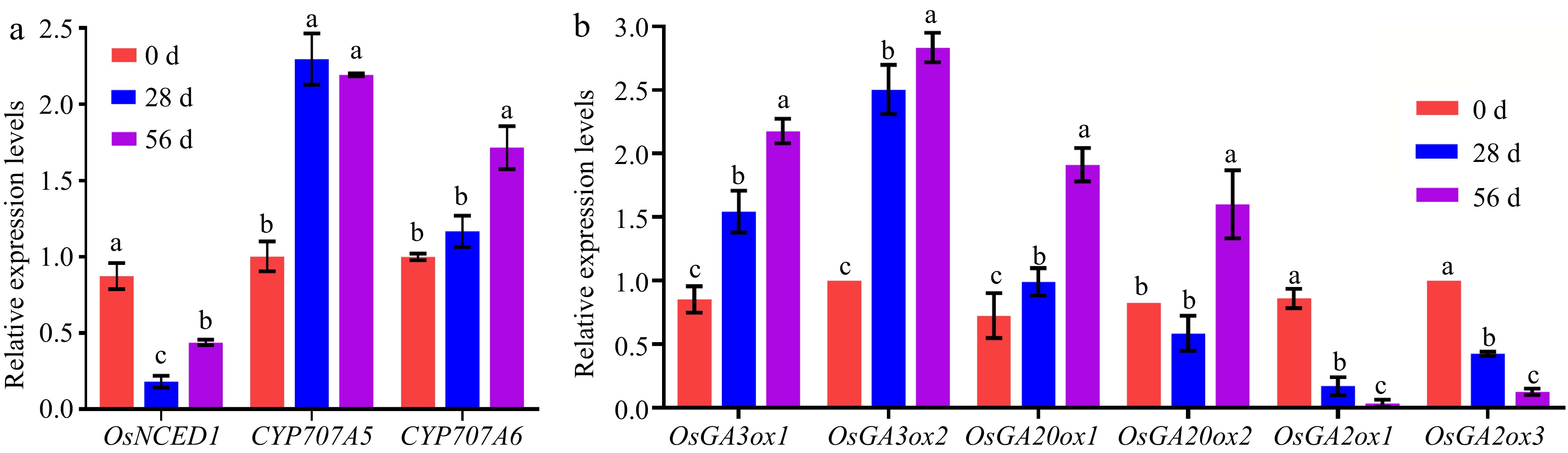
Figure 8.
Transcript levels of of ABA and GAs- related genes in Nipponbare seeds at 0, 28, and 56 DAR. (a) Relative transcript levels of OsNCED1 (ABA biosynthesis) and CYP707A5 and CYP707A6 (ABA catabolism) in NPB seeds at 0, 28, and 56 DAR. (b) Relative transcript levels of OsGA3ox1, OsGA3ox2, OsGA20ox1, and OsGA20ox2 (GA biosynthesis) and OsGA2ox1 and OsGA2ox3 (GA catabolism) in NPB seeds at 0, 28, and 56 DAR. NPB represents Nipponbare. The expression of genes is normalized to that of OsActin. Each column presents the means ± standard deviations of three biological replicates. Different lower case letters above columns indicate statistical differences at P < 0.05.
Compared to 0 DAR seeds, the transcript levels of the GA biosynthesis genes OsGA3ox1 and OsGA3ox2 increased in 28 and 56 DAR seeds, as did OsGA20ox1 and OsGA20ox2 in 56 DAR seeds. The catabolism genes OsGA2ox1 and OsGA2ox3 significantly decreased in 28 and 56 DAR seeds (Fig. 8b). Compared to 28 DAR seeds, the transcript levels of the GA biosynthesis genes OsGA3ox1, OsGA3ox2, OsGA20ox1 and OsGA20ox2 significantly increased in 56 DAR seeds. The catabolism genes OsGA2ox3 significantly decreased in 56 DAR seeds (Fig. 8b). The changes of transcript levels for these genes, related to GA metabolism, might be associated with GA9 and GA24.
-
Complete release of seed dormancy is of great importance when sowing in the field. Seeds with dormancy exhibit low germination and slow seedling establishment, or even an inability to germinate, resulted in a serious decline of crop yields. After-ripening is a common method used to break seed dormancy for dormant seeds in agricultural production. It is widely known that seed dormancy in most rice varieties can be broken through several months of after-ripening under dry room conditions. Consistently, Du et al. found that the seed dormancy of japonica variety Jiucaiqing with strong dormancy was released after 3 months of after-ripening[16]. Herein, we detected the characteristics of seed dormancy release in five rice varieties, including japonica NPB, NJ 9108, WYJ 7, and ZD 88, as well as indica 9311. The seed dormancy of all the five varieties were gradually released through after-ripening. Also, the seed dormancy released in WYJ 7, ZD 88, and 9311 seeds were faster than in NPB and NJ 9108 seeds during after-ripening. This demonstrated that the after-ripening period for release of seed dormancy differed among the different rice varieties. Therefore, farmers should master seed dormancy characteristics of different varieties to better sow their rice fields.
As important plant hormones, ABA and GA are extensively involved in regulating seed dormancy and after-ripening[3,6,42]. In our study, the significant decrease of ABA levels occurred at the early stage of after-ripening (0 to 28 DAR), not the late stage (28 to 56 DAR). The consistent changes of transcript levels of ABA-related genes were observed in two after-ripening stages. It is possible that these results were tightly associated with the water content in seeds at the two stages. Therefore, we measured the water content of NPB dry seeds at different time points of DAR. The results showed that the water contents of the fresh harvested seeds, 28 DAR seeds, and 56 DAR seeds to be 13.97%, 12.07%, and 9.74%, respectively. It is well known that the water content of rice seeds should be below 12% for safe storage. The cytoplasm of seed cells becomes highly viscous and is transformed into a glass after drying and dehydration[43]. In the glassy state, cellular components in dry seeds are stabilized and their mobility is severely restricted, preventing enzymatic reactions with free available water[44,45]. In this study the changes of ABA in seeds are possible that the early stage of after-ripening has a weak glass and the late stage has a deep glass. In addition, we detected the GA content in dry seeds. Bioactive GA1 content showed a slight increase but no significant difference, while inactive GA9 and GA24 content were increased during after-ripening. It is possible that the increase of GA9 and GA24 during after-ripening accelerated bioactive GA1/3/4 biosynthesis to induce the α-amylase activity during seed germination.
It is known that H2O2 can accumulate during seed storage under the dry state, and its accumulation has been tightly associated with the breaking of embryo dormancy[18,20−22]. In this study, we observed a significant increase in H2O2 content in the early stage of after-ripening, and then a decrease in the late stage. These observations suggested that seeds could face oxidative stress involved in ROS detoxification during desiccation[17]. Our results showed that SOD activity increased initially in the early stage of after-ripening and then declined in the late stage, while CAT activity increased significantly in the late stage. These indicated that SOD and CAT were likely involved in the regulation of H2O2 content at a precise level in after-ripened seeds. Previous investigation indicated that H2O2 promoted the production of α-amylase mRNA[46]. Similar results of α-amylase activity with H2O2 content were also observed in the two stages of after-ripening in NPB dry seeds, suggesting α-amylase activity was induced by the accumulation of H2O2 during after-ripening.
It is debatable whether there were transcriptional and translational activities in after-ripened seeds[4]. Meimoun et al. concluded that the regulated variation in gene expression did not occur during after-ripening, in agreement with the absence of metabolic activity in dry seeds[47]. Several transcriptome analyses showed that transcript abundances in seeds could be affected by after-ripening, resulting in the selective change of specific transcripts and, ultimately, dormancy release[35,44,48]. There is a relatively reasonable explanation, namely that the relative levels of some transcripts could appear to increase because they are more stable than the overall transcriptome and be influenced by oxidation in the dry seeds[44,49]. Therefore, 149 stored transcripts (mRNA), identified in NPB dry seeds between 0 and 28 DAR by RNA-seq, could be relative stable during after-ripening. To confirm the differential transcripts, we further used four additional rice varieties, and analyzed their dormancy levels and the transcript levels of the four top DTs and ABA biosynthesis OsNCED4 during after-ripening. RT-qPCR results showed that their transcript levels had a similar trend, although there were only slight differences among these rice varieties. The AR-up/down-regulated transcripts mainly occurred between 0 and 28 DAR dry seeds, rather than 28 and 56 DAR dry seeds, which were consistent with their degree of dormancy release. This suggested that these transcripts mainly occurred in the early stage of after-ripening and might be associated with seed dormancy release caused by after-ripening.
The transcripts of OsNCED4, the ABA biosynthesis gene[50], which significantly decreased during after-ripening, especially in the early stage, may be involved in the decrease in ABA in the early after-ripening stage. OsNCED3 is known to regulate seed dormancy[51], but was undetected by RNA-seq in this study. It is probable that OsNCED3 does not regulate the process of after-ripening, or that the expression of OsNCED3 was too low in after-ripened dry seeds. OsOXO1 encodes oxalate oxidase (OxO), which can catalyze the oxidative breakdown of oxalate to H2O2 and CO2[52]. This suggests that the degradation of OsOXO1 in after-ripening might be involved in the changes of H2O2 content. Previous studies have shown that RSOsPR10 and OsPR10a are both pathogenesis-related (PR) protein genes. Bioinformatic analyses showed that RSOsPR10 and OsPR10a were highly similar in protein sequence and structure, and were speculated to have similar functions. RSOsPR10 was a root-specific rice PR10, which could be rapidly induced by salt and drought stresses, and blast fungus infection[53,54]. OsPR10a was reported to play multiple roles in phosphate recycling in phophate-starved cells and improved resistance to pathogen infection[55]. However, the function of RSOsPR10 and OsPR10a in seed dormancy and germination has not been investigated in rice. In our study, the changes of transcript levels of RSOsPR10 and OsPR10a were observed in five rice varieties during after-ripening. Especially, transcript levels of RSOsPR10 had a large increase in the early stage of after-ripening in five rice varieties, suggesting tightly associated with dormancy release during after-ripening. It is deserved that the roles of the RSOsPR10, OsPR10a, OsOxO1, and OsNCED4 would be investigated further in the after-ripening process.
In summary, we proposed a model of the physiological characteristics and transcripts in rice after-ripened seeds, possibly associated with seed dormancy release. At the early stages of after-ripening, when seeds detached from the mother plant, there were some changes in transcription and translation in the dry seeds. ABA content significantly declined, H2O2 levels significantly increased, SOD and α-amylase activities significantly raised, but not bioactive GA1 significantly increased. The 149 differential transcripts were identified by RNA-seq and the changes of transcript levels, e.g. the decreased OsOxO1 and OsNCED4 and the increased RSOsPR10 and OsPR10a, mainly occurred at the early stage of after-ripening through the verification of four other rice varieties in after-ripened seeds. At the late stage of after-ripening, there were only slight changes in transcription and translation, in agreement with the absence of metabolic activity in fully dry seed[56]. This model significantly increased our understanding of the regulatory mechanism of seed after-ripening and the biological process of after-ripening in rice.
-
All data supporting the findings of this study are available within the paper and within its supplementary data published online. All the raw sequence reads of the RNA-Seq have been deposited in
https://pan.baidu.com/s/1onpR7Cd1h1LF4EjnJOW6qw (code: 2022). This research was supported by Hainan Yazhou Bay Seed Laboratory (project of B21HJ1002), the National Natural Science Foundation of China (Grant No. 32272169, 32172037 and 32000377), and the Natural Science Foundation of Jiangsu Province (Grant No. BK20201322).
-
Hongsheng Zhang is the Editorial Board member of journal Seed Biology. He was blinded from reviewing or making decisions on the manuscript. The article was subject to the journal's standard procedures, with peer-review handled independently of this Editorial Board member and his research groups.
-
# These authors contributed equally: Fengzhi Yuan, Yongji Chen
- Supplemental Fig. S1 RT-qPCR verification of DTs selected from RNA-seq. RT-qPCR validation of up-regulated (A) and down-regulated (B) DTs. The transcript levels of genes was normalized to that of OsActin. Each column represents the mean ± standard deviation, n = 3.
- Supplemental Table S1 The primer pairs used in this study.
- Supplemental Table S2 149 DEGs between 0 and 28 DAR Nipponbare seeds via RNA-seq.
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press on behalf of Hainan Yazhou Bay Seed Laboratory. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Yuan F, Chen Y, Chen X, Zhu P, Jiang S, et al. 2023. Preliminary identification of the changes of physiological characteristics and transcripts in rice after-ripened seeds. Seed Biology 2:5 doi: 10.48130/SeedBio-2023-0005 

Preliminary identification of the changes of physiological characteristics and transcripts in rice after-ripened seeds
- Received: 14 November 2022
- Accepted: 28 March 2023
- Published online: 25 April 2023
Abstract: A common way of breaking seed dormancy in cereal crops is by treating seeds with a period of dry storage or after-ripening. In this study, we determined changes in the physiological characteristics and transcripts in after-ripened seeds in rice. Phenotypic data showed that the seed dormancy of five rice varieties was gradually released with after-ripening. Physiological results showed that, at the early stage of after-ripening from the freshly harvested seeds (0 d after-ripening, DAR) to 28 DAR, abscisic acid (ABA) content significantly decreased, hydrogen peroxide (H2O2) levels significantly increased, superoxide dismutase (SOD) and α-amylase activities significantly raised, and bioactive gibberellin (GA1) content and ascorbate peroxidase (APX) activity showed no changes in Nipponbare seeds. RNA-seq showed that the 149 changed transcripts were observed at the early stage of after-ripening. Of these transcripts, transcript levels of OsOxO1, OsNCED4, RSOsPR10, and OsPR10a mainly occurred at early stages by the verification of four other rice varieties in after-ripened seeds, perhaps signifying that those transcripts played pivotal roles in rice after-ripening. At the late stage of after-ripening, from 28 to 56 DAR, there were merely changes in transcription and translation. Our results facilitate an understanding of the biological process of after-ripening and dormancy release in rice seeds.
-
Key words:
- Rice /
- After-ripening /
- Physiological changes /
- Transcripts /
- Dormancy release
{kind=link}