









-
Grape (Vitis vinifera L.) is one of the fruits with the largest planting area and most yield worldwide, with a long history of cultivation and about 75% of the grapes are used to produce wines[1]. Style and quality of wines are determined by a combination of many substances. Flavonoid compositions at maturity are the main determinant of the sensory quality of grapes and wines, such as color, astringency and bitterness. Depending on the structures, flavonoid compounds can be classified as anthocyanins, flavonols and flavanols. Anthocyanins are the major color-presenting compounds, and their content, composition and proportion, as well as their aggregation with other flavonoid compounds, affect the color of wines. Flavonols protect berries from UV radiation as photoprotectants and act as co-pigments in wines[2]. Flavanols can also form polymeric pigments with anthocyanins to improve wine color stability[3]. In addition, flavanols in grape skins and seeds exist in the form of flavan-3-ols and proanthocyanidins, where the concentrations of flavan-3-ol positively correlated with wine bitterness, and the degree of polymerization of proanthocyanidins is closely related to wine astringency[4]. Therefore, grapes with appropriate levels of flavonoids are in high demand by cultivators, which can be modified by the rootstocks and several other factors (viticultural practices, climate, cultivar, etc.).
The use of grafting technique began with the spread of phylloxera in Europe. V. vinifera and many hybrid cultivars need to be grafted onto a rootstock that can provide resistance to phylloxera. At present, rootstocks are widely used in viticulture and their resistance to biotic and abiotic stresses in particular has been well documented[5,6]. Aside from these functions, rootstocks have been proven to affect vine growth and yield[7]. In accordance with several studies, rootstocks with different genetic backgrounds modified root dynamics, and consequently, nutrient uptake, canopy biomass, and finally, the composition of grapes[8]. Environmental factors may affect the influence of rootstocks, such as climatic differences in years, soil composition, etc. that may amplify or reduce rootstock variables. Several previous studies also documented that the effects of rootstocks on vine growth were significantly related to soil water availability, acidity, nutrients, etc.[9−11] and soil water availability can also be greatly affected by the presence of cover crops, with significant effects on water consumption by vines and qualitative and quantitative aspects of yield[12,13]. In addition, cultivars are correlated with the effect of rootstocks in a bidirectional action. Keller et al.[14] found that 3309C induced Syrah to exhibit low vine vigor and conversely induced high vigor in Chardonnay. Gutierrez-Gamboa et al.[15] indicated that Merlot grapevines grafted onto SO4 presented a higher concentration of total proanthocyanidin in grape skins and seeds in comparison with seven grafted vines, whereas the Merlot/Gravesac combination showed a lower concentration of proanthocyanidins in grapes. It is widely accepted that the strong effects of rootstocks on the vegetative-reproductive growth of the scion may alter the cluster zone microclimate and the berry ripening process, thus modifying the synthesis of flavonoid compounds in grape berries[16−18]. For example, Nelson et al.[19] revealed that the differences in the anthocyanin compositions between two grafted vines were associated with the variations in the size of the canopy, which could regulate the sunlight exposure received by their clusters. Koundouras et al.[20] found that the rootstock may modify the composition of grapes by altering the fruit ripening rate. Han et al.[21] found that a combined effect between vintages and rootstocks, while rootstock played a key role in accumulation of flavonoids in seasons with more rain and cloudy days. There are complex interactions among rootstocks, scion, soil and climatic conditions, so it is vital that each rootstock should be evaluated for a specific cultivar in the given region.
Vitis vinifera L. cv. Petit Verdot, native to France, has a high content of organic acids and tannins in berries. In recent years, Petit Verdot has been planted in eastern China as a very promising wine grape cultivar. The climate of eastern China during the grape growing season is different from the mild Mediterranean climate with little rain and from the hot and dry climate of western China, which is a region with a lot of rain and low light. Under similar climatic conditions, there are few reports about the effects of different rootstocks on the flavonoid profiles of Petit Verdot grapes. In the present study, we used HLPC-MS to evaluate the effects of five rootstocks on the berry flavonoids of Petit Verdot in eastern China, aiming to provide some reference for the selection and application of rootstocks in practice, as well as to provide new insights for understanding the effects of rootstocks on fruit flavonoid profiles.
-
A two-year field experiment (2016−2017) was conducted at the Shangzhuang experimental station (40°14′ N, 116°20′ E, 49 m altitude) of China Agricultural University, Beijing, China. In the current study, the Petit Verdot (PV) grapevines were planted and green-grafted onto Paulsen 1103 (1103P), Selection Oppenheim (SO4), Kober 5BB (5BB), Millardet et de Grasset 101-14MGt (101-14MGt) and Beta in 2012, spaced at 2.5 m × 1.2 m with rows oriented south to north. The own-rooted PV was the control. A modified vertical shoot positioning (M-VSP) training system[22] was used in the vineyard, which was spur-pruned and retained 12−15 nodes per row meter. Additionally, drip irrigation, pest and nutrition management were applied in accordance with the local industry standards. The meteorological data (mean monthly temperature, sunshine duration, and rainfall) of this vineyard during grape development were provided by the China Meteorological Data Sharing Service System (
http://cdc.cma.gov.cn/ ).In 2016, a randomized block design with three biological repetitions of each graft combination was conducted in the experiment, and each biological repetition consisted of ten vines. The experiment in 2017 was conducted on the same vines. The harvests in 2016 and 2017 were on the 25th and 24th of September, respectively. At harvest, six berries on the top, bottom, left, right, front and center of each cluster were collected, a total of 300 berries were randomly collected for each biological repetition. The sampling time was before 10:00 am to avoid the high temperature at midday. The physicochemical characteristics of 100 berries were quickly determined after sampling, and the rest of berries were used to determine flavonoid and volatile composition.
Determination of berry physiochemical parameter
-
Berry physiochemical parameters were determined according to the National Standard of People's Republic of China (GB/T15038-2006). For each replicate, 100 berries were randomly selected for weighting, and then manually squeezed. The must was centrifuged at 8,000× g for 5 min at room temperature to extract the supernatant, and the clear juice was determined for total soluble solids (TSS), pH and titratable acidity (TiA). The TSS was measured with a PAL-1 digital portable refractometer (Atago, Japan). The determination of pH was carried out using a PB-10 desktop pH meter (Sartorius, Germany). Juice TiA was titrated with NaOH (0.05 M) to the endpoint of pH 8.2 and represented as tartaric acid (g/L).
Extraction of flavonoids compounds in grapes
-
Mature berry skins and seeds were manually peeled off in liquid nitrogen. Then the skins and seeds were freeze-dried at −40 °C and ground. The extraction of flavonols and anthocyanins was consistent with the method described by Downey et al.[23]. The dried skin powder was accurately weighed (0.100 g) into a 2 mL centrifuge tube mixed with 50% (v/v) methanol in water (1.0 mL), and then sonicated for 20 min. The extraction was centrifuged for 10 min at 12,000 rpm. The supernatant was collected and the residue was extracted twice. The flavan-3-ols in skins and seeds were extracted in accordance with the method described by Liang et al[24]. The extraction of free flavan-3-ols is as follows: the dried powder (0.100 g) was mixed with 0.005 g ascorbic acid and a solution (1.0 mL) of 70% (v/v) acetone in water, and then shaken for 15 min. The mixture was centrifuged for 15 min at 8,000 rpm. The supernatant was collected and the extraction of residue was conducted twice. The supernatant was combined and shaken well, 400 μL pooled supernatants was rapidly blown dry at 30 °C and darkness using nitrogen gas. The dried samples were dissolved in 200 μL of methanol containing 1% HCl, then neutralized with 200 μL aqueous sodium acetate (200 mM). The extraction of proanthocyanins is as follows: 0.100 g dried powder was mixed with a phloroglucinol buffer (0.5% ascorbate, 300 mM HCl, and 50 g/L phloroglucinol in methanol), and then incubated for 20 min at 50 °C, with a final combination of 1.0 mL aqueous sodium acetate (200 mM). The extract was centrifuged at 8,000 rpm for 15 min. The residue was extracted twice. All the supernatants were collected and stored at −40 °C.
Analysis of flavonoid compounds in grapes
-
Before sample analysis, all extracts were filtered through 0.22 μm nylon membrane (Membrana, Wuppertal, Germany). Flavonols in mature grape skin were analyzed in accordance with the procedure reported by Sun et al.[25]. Agilent 1200 series HPLC-MSD trap VL (Agilent, Santa Clara, CA, US) was connected simultaneously to a variable wavelength detector and a Zorbax EclipseXDB-C18 column (250 mm × 4.6 mm, 5 μm) was used. The analysis of anthocyanins in mature grape skin was conducted based on the methods developed by Cheng et al.[22]. Anthocyanins were determined using an Agilent 1100 series HPLC-MSD trap VL linked to a diode array detector (DAD) and a Kromasil C18 column (250 mm × 4.6 mm, 5 μm). Flavan-3-ols in mature grape skin and seed were measured with the method described by Li et al.[26]. A DAD and a Poroshell 120 EC-C18 column (150 mm × 2.1 mm, 2.7 μm) were employed on an Agilent 1200 series HPLC system which was coupled with an Agilent 6410 triple quadrupole mass spectrometer (QqQ) with an electrospray ionization source. The detections were carried out in positive ionization modelity. The mobile phases used for elution are 0.1% aqueous formic acid solution as phase A, and 50/50 methanol–acetonitrile solution containing 0.1% formic acid as phase B. Flavonols and anthocyanins were expressed in mg/kg of fresh berry weight (FW) as quercetin-3-O-glucoside and malvidin-3-O-glucoside equivalents, respectively. Flavan-3-ol concentrations were quantified using catechin (C), epicatechin (EC), epicatechin-3-O-gallate (ECG), and epigallocatechin (EGC) as external standards and were measured in mg/kg fresh berry weight (FW).
Statistical analysis
-
Differences of concentrations of compounds were determined by one-way analysis of variance (ANOVA). Differences in physicochemical parameters were determined by two-way ANOVA. The ANOVA was performed using the SPSS 26.0 (IBM, US) by employing Duncan's multiple range test at a level of p < 0.05. The figures were drawn using GraphPad Prism 8.0.2 (GraphPad Software, US). Principal component analysis (PCA) was performed using Simca 14.1 (Umetrics, Sweden).
-
A typical continental monsoon climate prevailed at the experimental vineyard. By analyzing the meteorological conditions for 2 years as indicated by the data in Table 1, it was found that there was no significant difference in the average temperature and sunshine duration from May to September. However, the rainfall displayed great differences between the two growing seasons. Specifically, the precipitation from July to September in 2017 accounted for only 70% of that in 2016, and the rainfall in July and September in 2017 was less than a quarter of that in 2016. Compared to the average data of the last 10 years (2008−2017), there was an increase in sunshine hours and daily temperatures in 2016 and 2017. In the last decade, the average monthly precipitation of 2016 was higher, which was mainly contributed by the rainfall in July. Although the average monthly rainfall in 2017 was consistent with the past decade, its precipitation was higher in August and almost non-existent in September.
Table 1. Meteorological conditions of the experimental vineyard from May to September in 2016−2017.
Year Month Mean May June July August September Mean daily temperature (°C) 2016 21.5 25.9 27.4 27.5 22.2 24.9 2017 23.3 25.6 27.9 26.2 23.0 25.2 Historical average (2008−2017) 21.9 24.5 27.5 26.4 22.1 24.5 Maximum daily temperature (°C) 2016 28.1 31.4 31.8 31.8 27.2 30.1 2017 29.7 31.7 32.3 31.1 28.4 30.7 Historical average (2008−2017) 27.9 30.4 32.1 31.1 27.3 29.8 Minimum daily temperature (°C) 2016 14.8 20.1 23.8 23.0 17.7 19.9 2017 16.3 19.4 24.0 22.2 17.7 19.9 Historical average (2008−2017) 15.7 18.4 23.4 22.3 17.3 19.4 Sunshine duration (h) 2016 281.9 224.4 153.4 218.7 201.5 281.9 2017 298.5 250.9 179 205.9 214.2 298.5 Historical average (2008−2017) 269.7 219.0 180.0 209.8 194.5 269.7 Precipitation (mm) 2016 24.0 72.9 344.3 76.8 59.0 115.4 2017 31.2 119.5 97.4 233.9 2.8 97.0 Historical average (2008−2017) 30.9 83.5 176.4 116.4 60.4 93.5 Effects of rootstocks on berry physicochemical parameters
-
The berry weight, total soluble solids, titratable acidity and pH of mature berries were shown in Table 2. The effects were not the same for different physicochemical parameters and depended on the rootstocks and the vintages. In terms of berry weight, an increase in berry weight was observed for all grafted vines compared to own-rooted vines, but the difference was not significant. For the TSS of berries, there was no significant influence of rootstock combinations when compared to PV. The pH in PV juice was lower relative to that in the grafted vines, although their differences were not significant except for PV/Beta. Juice TiA was not influenced by either rootstock or vintage in this study. Moreover, berry weight, TSS and pH were significantly affected by vintages. The higher berry weight, TSS and juice pH in 2016 may be due to more precipitation and shorter sunshine durations in 2016 compared to 2017, which delayed berry ripening. Two-way ANOVA showed that only the pH of juice were significantly affected by the interaction of rootstock and vintage.
Table 2. Physicochemical parameters of mature berries on different rootstocks in the two seasons (2016−2017).
Source of variation Berry weight (g/100 berries) Total soluble solids (oBrix) pH Titratable acidity (g/L) Rootstock (R) PV 135.56 ± 12.30a 18.70 ± 2.69ab 3.00 ± 0.19b 11.14 ± 3.04 PV/101-14MGt 146.47 ± 6.61 19.46 ± 1.57ab 3.06 ± 0.51ab 10.95 ± 0.84 PV/1103P 164.77 ± 11.92 18.98 ± 1.16ab 3.08 ± 0.41ab 11.26 ± 2.76 PV/Beta 168.34 ± 58.71 20.32 ± 1.38a 3.13 ± 0.14a 9.23 ± 1.96 PV/5BB 197.57 ± 77.01 18.28 ± 1.58b 3.09 ± 0.11ab 10.47 ± 1.36 PV/SO4 179.12 ± 65.70 18.43 ± 0.87b 3.05 ± 0.73ab 10.44 ± 1.75 Vintage (V) 2016 182.73 ± 64.24a 19.97 ± 1.16a 3.13 ± 0.10a 10.15 ± 2.23 2017 147.69 ± 15.26b 18.10 ± 1.61b 3.00 ± 0.09b 11.10 ± 1.83 Significanceb R 0.321 0.115 0.171 0.428 V 0.047 0.000 0.000 0.179 R × V 0.611 0.204 0.032 0.255 a Data are expressed as means of three replicates, and different letters within the same column indicate significant differences among the rootstocks or vintages in accordance with Duncan’s test (p < 0.05). b Two-way ANOVA tests for significance of the differences of rootstock × vintage interaction. Effects of rootstocks on berry anthocyanin concentration
-
Over two-successive vintages, anthocyanin concentrations in grape skins were significantly affected by rootstocks (Fig. 1). Regarding the anthocyanin concentrations in mature grapes in this study, it has been shown that grapes from own-rooted vines presented lower concentration of anthocyanins than the grapes from PV/1103P and PV/Beta in two years, except caffeoylated anthocyanins. Besides, no significant differences in anthocyanin concentrations were found between PV/5BB, PV/SO4 and own-rooted vines. Generally, the effects of 101-14MGt rootstock on the concentration of anthocyanins depended on the vintage in this study. In 2016, 101-14MGt slightly reduced the concentration of total anthocyanin while significantly reduced the concentration of several anthocyanins, such as malvidins, anthocyanins in glucoside form and methoxylated anthocyanins (Fig. 1). On the contrary, as in the grapes from PV/1103P and PV/Beta, PV grafted onto 101-14MGt showed significantly positive effects on accumulation of the anthocyanins in 2017, except the caffeoylated anthocyanins. With few exceptions, the rootstocks did not show a consistent effect on the proportions of different types of anthocyanins in the two seasons. 101-14MGt, 1103P and 5BB significantly reduced the proportions of anthocyanins in glucoside form in the two seasons. In 2016, most grafted vines showed higher proportions of acetylated, caffeoylated and 3'-hydroxylated anthocyanins, and lower proportions of coumarylated, methoxylated and 3′5′-hydroxylated anthocyanins. The effects of rootstocks on the proportions of different anthocyanin fractions in 2017 were opposite to those in 2016, except for the methoxylated anthocyanins.
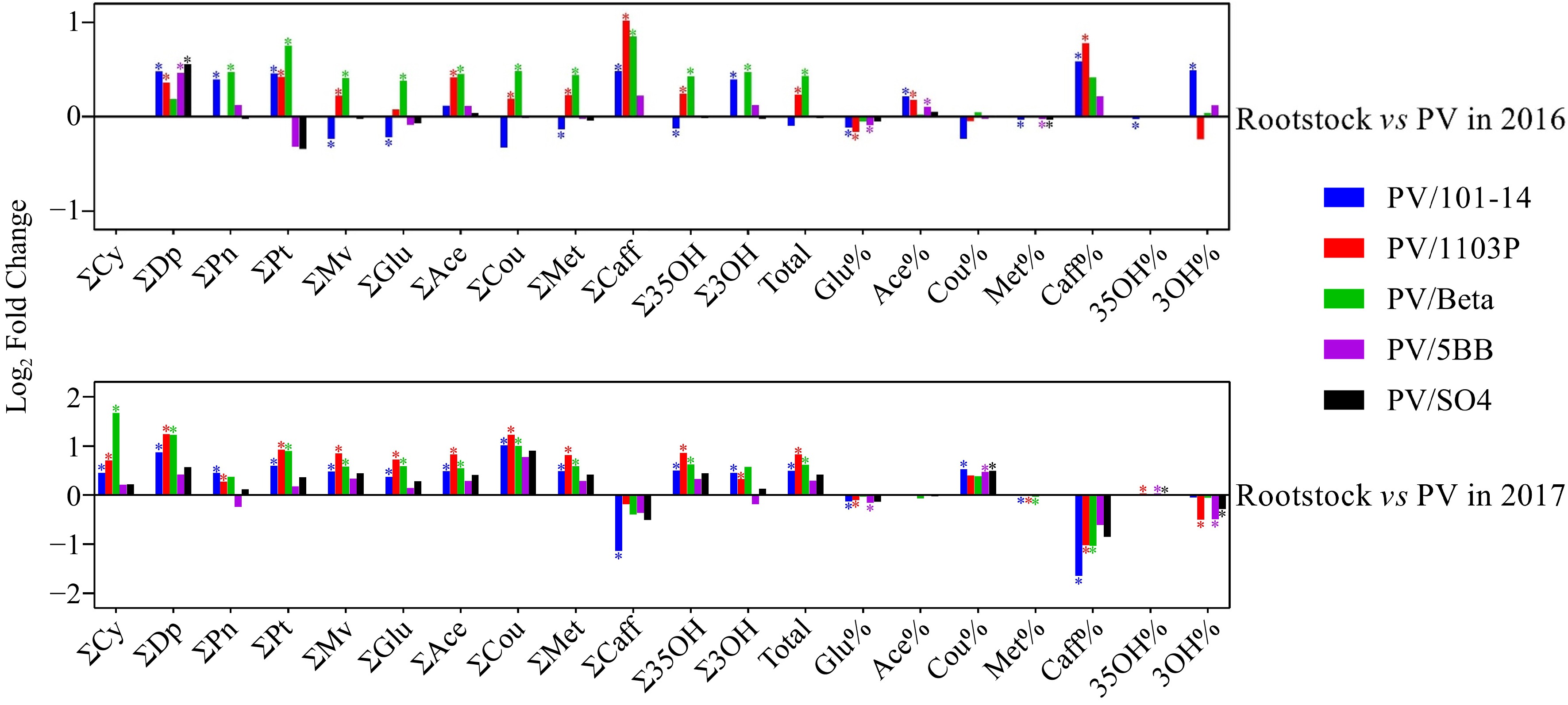
Figure 1.
Effect of rootstocks on anthocyanins in two vintages (2016−2017). Each data represents the log2 fold change in anthocyanin concentration/proportion in mature berries on each rootstock relative to those on own-rooted vines. '∑', the total concentration of different types of anthocyanins; '%', the proportions of different types of anthocyanins; 'Total', the total concentration of anthocyanins; 'Cy', cyanidin; 'Dp', delphinidin; 'Pn', peonidin; 'Pt', petunidin; 'Mv', malvidin; 'Glu', anthocyanins in glucoside form; 'Ace', acetylated anthocyanins; 'Cou', coumarylated anthocyanins; 'Met', methoxylated anthocyanins; 'Caff', caffeoylated anthocyanins; '35OH', 3'5'-hydroxylated anthocyanins; '3OH', 3'-hydroxylated anthocyanins. The * on each column indicates a significant difference between rootstock and own-rooted vines in accordance with Duncan's test (p < 0.05).
Effects of rootstocks on berry flavonol concentration
-
The flavonol concentrations of mature berries affected by rootstock were shown in Fig. 2. In this study, it was found that the total flavonol concentration of PV/101-14MGt and PV/1103P combination was significantly higher than that of own-rooted vines, which was mainly caused by the increased concentrations of myricetin and quercetin. Besides, these two rootstock combinations had higher concentrations of most flavonols with few exceptions in two-successive seasons. On the contrary, the PV/Beta tended to attenuate the concentration of flavonols in 2016 and 2017, except for the syringetin. In addition, the significant differences of flavonols between PV/5BB and own-rooted vines were shown in 2017. Over two vintages, 5BB did not significantly affect the concentration of flavonols in mature berries, except few exceptions. As with PV/5BB, there were no significant differences in flavonol concentration in PV/SO4 compared with own-rooted vines in 2016. However, SO4 significantly increased the concentration of most flavonols in 2017, except for the kaempferol and laricitrin.
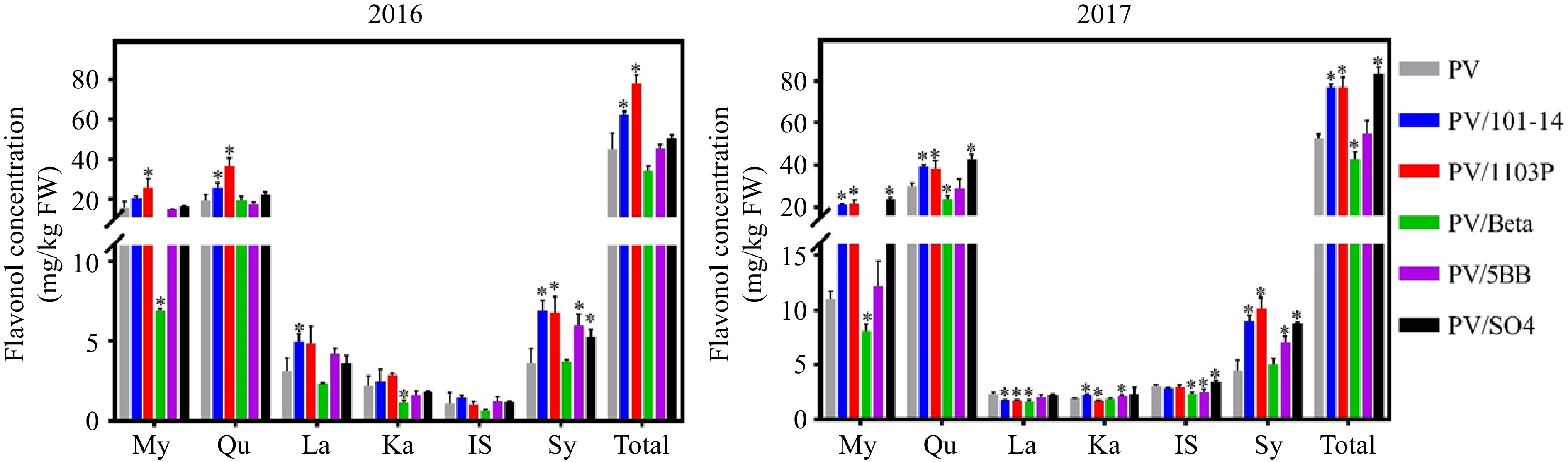
Figure 2.
Effect of rootstocks on flavonols in two vintages (2016−2017). 'My', myricetin; 'Qu', quercetin; 'La', laricitrin; 'Ka', kaempferol; 'IS', isohamnetin; 'Sy', syringetin; 'Total', the total concentration of flavonols; The * on each column indicates significant difference between rootstock and own-rooted vines in accordance with Duncan's test (p < 0.05).
Effects of rootstocks on berry flavan-3-ol concentration
-
Minor differences in flavan-3-ol concentrations of grape skins emerged between grafted and own-rooted vines during the two seasons (Supplemental Table S3). Nevertheless, 101-14MGt and SO4 still tended to promote the accumulation of most flavan-3-ol fractions in grape skins, and the total flavan-3-ol concentration in 2017 was significantly higher than that of own-rooted vines. Contrarily, PV/1103P showed slight negative effects on flavan-3-ol accumulation in skins. Regarding the flavan-3-ol concentration in grape seeds, there were no significant differences among the grafted and own-rooted vines (Supplemental Table S3). In particular, 1103P had almost no influence on the flavan-3-ol concentrations in the two seasons. Besides, 101-14MGt and Beta showed slight positive influences on the content of flavan-3-ols in grape seeds in this study. Regarding the mean degree of polymerization in skins and seeds, no significant differences were found between the own-rooted vines and grafted vines in two-successive seasons.
Principal component analysis of flavonoids compounds in grape
-
Principal component analysis (PCA) was used as the first approach to further clarify the characteristics of flavonoid compounds in the mature grapes of grafted Petit Verdot vines and own-rooted vines, as shown in Fig. 3a. As expected, it was found that the vintage was the major factor for separating the samples, while the grafted vines and own-rooted vines were not clearly distinguished in the PCA model. The first two principal components explained 53.0% and 15.4% of the total variance, respectively. The first principal component (PC1) was classified in 2016 and 2017. The loading plots showed that the PV and grafted vines had higher concentrations of anthocyanins in grapes and higher concentrations of flavan-3-ols in seeds, while the grapes had higher concentrations of flavonols and flavan-3-ols in skins in 2017.
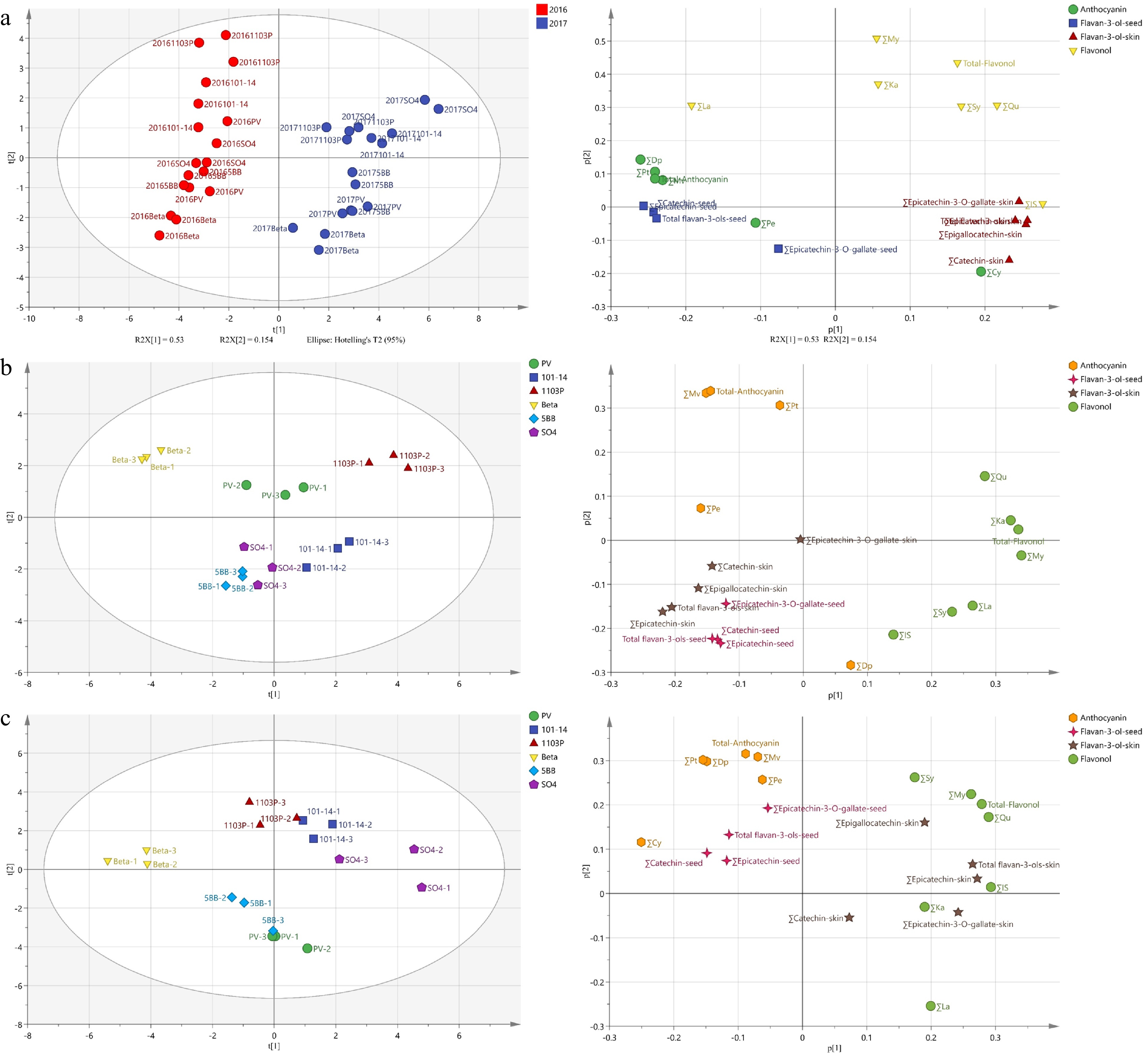
Figure 3.
(a) Principal component analysis (PCA) based on flavonoid compound concentrations in grape of 2016 and 2017, (b) grapes of 2016, (c) grapes of 2017. Left side is score plot and the right side is their corresponding loading plot. '∑', the total concentration of different types of flavonoid; 'Cy', cyanidin; 'Dp', delphinidin; 'Pn', peonidin; 'Pt', petunidin; 'Mv', malvidin; 'My', myricetin; 'Qu', quercetin; 'La', laricitrin; 'Ka', kaempferol; 'IS', isohamnetin; 'Sy', syringetin.
To reduce the dominant influence of vintages, the PCA was performed in 2016 and 2017 based on the original concentrations of flavonoids to investigate the biomarkers of different rootstocks (Fig. 3). For 2016 grapes, PC1 accounted for 31.2% of total variance. Samples of PV/1103P and PV/101-14MGt combinations were located at the positive part of PC1, whereas the samples of PV/Beta combination were located in the negative part of PC1. Flavonols contributed positively to the PC1, while the anthocyanins and flavan-3-ols contributed negatively. PC2 explained 19.4% of the total variance, classifying PV/Beta and PV/1103P combinations into one group and other rootstocks into another, with the PV located in between the two groups. Grapes of PV/1103P and PV/Beta contained more anthocyanins, while grapes of the other rootstock combinations included more delphinidins, several flavonols (isohamnetins, syringetins and laricitrins) and more flavan-3-ols in skins and seeds. In 2017, PC1 explained for 32.9% variation and classified samples into three groups. Grapes of PV/SO4 and PV/101-14MGt combinations had a higher concentration of flavonols and flavan-3-ols in skins, whereas grapes of PV/Beta accompanied with more flavan-3-ols in seeds and anthocyanins. PC2 accounted for 26.1% of total variance, distinguishing four rootstock combinations such as PV/1103P from PV and PV/5BB, which was explained positively by most flavonoids and negatively by laricitrins. In combination with consistent results over two years, these results indicated: 1103P promoted the accumulation of anthocyanins and flavonols in PV berry skins; Beta favored the biosynthesis of anthocyanins in skins and flavan-3-ols in seeds of PV grapes and discouraged the biosynthesis of flavonols in skins; 101-14MGt was able to enhance the concentration of flavonols and flavan-3-ols in PV grape skins; and SO4 increased the accumulation of flavanols in the skins of PV grapes.
-
The berry composition at harvest, which is the major determinant for grapes and wines depend on complicated interaction among several factors including environment, rootstocks and viticultural practices[15,27−29]. In the background of climate warming, grapevines need to cope with the increasing water stress, and therefore rootstock and water status are more important among other factors[30] and the application of sensors could help in managing the irrigation . In this study, the rainfall displayed great differences between 2016 and 2017. The precipitation from July to September in 2016 was significantly higher than in 2017, especially and the rainfall in July and September in 2017 was less than a quarter of that in 2016. Notably, veraison starts in July and the harvest commences in September, and the period have a great impact on the ripening and compositions of grapes. Therefore, the more precipitation and less sunshine duration in 2016, which resulted in moderate grape ripening compared with 2017. Delayed phenology may have contributed to higher berry weight, TSS, pH, and lower TiA in 2016.
Generally, the rootstocks could affect the ripening rate of grapes by modifying the vegetative and reproductive growth of the vines. As previously studied, Cabernet Sauvignon vines had higher reproductive growth when grafted onto 1103P, which resulted in a delayed ripening of Cabernet Sauvignon grapes[21,31]. Nevertheless, the effect of the same rootstock may vary on different cultivars[32]. Based on the physicochemical parameters of grapes, no effect of rootstock on berry ripening rate was observed in Petit Verdot. Except for the interaction with vintages which significantly affected pH, this study reported that rootstock had only a minor effect on fruit physicochemical parameters, which was in agreement with our previous study[33]. Besides, the influence of rootstocks on physicochemical parameters of mature grapes was reported recently, which did not consistently alter the berry weigh, TSS, pH and TiA[15,21,27]. Notably, the effect of rootstock on the pH and TiA is probably related to the absorption of potassium, as potassium precipitates tartrates in juice, resulting in a reduction in tartaric acid[34]. However, the potassium uptake capacity of grapevine is affected by the interaction of cultivars, rootstocks and soil conditions, and potassium with the greater availability in the granite soil could mask the differences in potassium uptake capacity of different rootstocks[9]. In this study, there was no significant difference in TiA and pH of juice between grafted grapes, which may be attributed to the great potassium availability of vineyard soil as we reported earlier[33].
Anthocyanins are distributed in the skin of red grapes and are the key substances for the coloration of red grapes and wines[35]. In agreement with previous studies[36], the main contributor to anthocyanin profile of grapes is the acetylated and methoxylated anthocyanins in Petit Verdot (Supplemental Table S2). However, the main contributor to total anthocyanins, in several cultivars such as Carignan and Garnacha, is the coumaroylated form[37,38]. This indicated that there were some differences in anthocyanin profiles among cultivars as expected because of the genetic distances existing for the different genotypes[39]. Interestingly, among the acylated anthocyanins, both the concentration and the proportion of caffeoylated anthocyanins were greatly influenced by the rootstock in this study, which was consistent with a recent study[33]. Of the rootstocks determined in this study, 1103P and Beta were beneficial to the accumulation of anthocyanin concentrations in grape skins. However, the viticultural literature often categorized 1103P as a very vigorous rootstock which tends to increase leaf-to-fruit ratio and attenuate the anthocyanin biosynthesis[21,31,40]. This inconsistent results mainly due to that Petit Verdot were more likely to be temperature sensitive and more leaves improved the microclimate of the bunch zone, alleviating the high temperature stress and promoting the synthesis of anthocyanins[41]. Beta is a commonly used rootstock in eastern China, because of its strong cold resistance[42]. It also has been reported that Summer Black grafted onto Beta rootstock increased the anthocyanin concentrations, which is consistent with the current study[43]. With few exceptions, the effect of 101-14MGt was strongly influenced for anthocyanin concentrations by the season factor. Besides, 5BB and SO4 did not significantly influence anthocyanin concentrations in agreement with other previous studies[20,33]. Notably, the effect of rootstocks on anthocyanin profiles was found to be more obvious in the growing season with less precipitation and more light intensity. This might be due to the fact that intense light upregulates the expression of key genes of the anthocyanin biosynthetic pathway[41] and therefore amplified the effect of rootstocks.
Flavonols are important copigments that could stabilize the color of the wine. In agreement with the previous study[33], 5BB did not significantly affect flavonols concentrations in mature berries in the present study. Besides, PV/101-14MGt and PV/1103P combinations had higher flavonol concentrations in skins in the two growing seasons. Quercetin, as the most abundant flavonol in Petit Verdot, was also statistically increased by 101-14MGt and 1103P. These results indicated that 101-14MGt and 1103P grafted onto Petit Verdot tended to promote the biosynthesis of flavonols. Notably, flavonols are particularly sensitive to light, and have ultraviolet protection in the grape skins. A study found that 1103P increased the pruning weight and resulted in insufficient light, thus attenuated the flavonol concentrations in Cabernet Sauvignon[21]. In addition, Beta significantly reduced the concentrations of flavonols in skins in this study, while a study reported that this rootstock induces low vigor to scions which could increase the exposure of the berries[42].
Most of flavan-3-ols accumulate in the grape berry seeds and skins. According to previous studies, there was no significant effect of rootstock on flavan-3-ol concentrations, indicating that flavan-3-ol concentrations were relatively stable and not easily influenced by cultivation practices[21,44,45]. Nonetheless, in this study, Petit Verdot grapevines grafted onto the 101-14MGt increased the concentrations of flavan-3-ol to some extent in the two seasons, which is consistent with the previous report[15]. Furthermore, the vintages remained the dominant factor for the difference in flavan-3-ol concentrations in grapes among the grafted and own-rooted vines. In this study, the flavan-3-ol concentrations of grapes were lower in 2017 with plenty light and less rain, and higher in 2016 with less light and more rain. This difference was mainly caused by the special accumulation pattern of flavan-3-ols. A recent study has shown that flavan-3-ols in grape berries usually reaches its highest concentration at the veraison and then shows a decreasing trend[46]. Plenty light and less rain during veraison accelerate the decomposition rate of flavan-3-ols, which in turn causes the low flavan-3-ol concentrations in mature berries in this vintage[47]. Furthermore, since the astringency of wines depends not only on the concentrations of proanthocyanidin but also on its structure, the mean degree of polymerization (mDP) is also an indicator that is often focused on. In agreement with the previous reports, the mDP of skin proanthocyanidins was higher than that of seed proanthocyanidins[48]. Gutiérrez-Gamboa et al.[38] revealed that 101-14MGt and SO4 increased the skin proanthocyanidin mDP of Merlot. However, this result was not clearly evidenced in Petit Verdot. Moreover, changes in the phytohormone content in grape can affect the content of several phenolic compounds such as procyanidi, terpenoid derivatives peonidin-3-glucoside, catechin and epicatechin[49].
-
In recent years, Petit Verdot grapes have been trialed and promoted in some production areas in eastern China, but there are limited reports on the effects of rootstocks on flavonoid profiles in Petit Verdot grapes. Commonly, different rootstocks could either delay ripening or accelerate ripening of the grape berries, apart from other effects, but there was no clear evidence in Petit Verdot in the current study. Vintage was the decisive factor in berry physiochemical parameters as already reported in several studies, whereas rootstocks had no significant effects on berry physicochemical parameters such as berry weight, titratable acidity, total soluble solids and pH, except for Beta which significantly increased the pH. The rootstocks played an important role in the accumulation of flavonoid compounds, especially anthocyanins and flavonols. 1103P increased the accumulation of anthocyanins and flavonols in berry skins in the two seasons. Beta significantly increased the anthocyanin concentrations of the berry skins in this study, while also showing a detrimental effect on the biosynthesis of flavonols except for syringetin. Flavan-3-ols were relatively stable in grapes and less influenced by rootstocks, with no significant differences observed between the two years. The effects of SO4 on anthocyanins and flavonols in grape skins were susceptible by seasonal factors and did not show consistent results in the two-successive years. The PCA showed that PV/5BB was closest to the own-rooted vines in terms of flavonoid compounds and the rest of the rootstocks were different from the own-rooted vines. Overall, the current study provided some interesting data for some rootstocks to be used for Petit Verdot in eastern China.
This research was supported by China Agriculture Research System of MOF and MARA (CARS-29).
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 Effects of five rootstocks (101-14MGt, 1103P, Beta, 5BB, SO4) on anthocyanin compounds in mature grapes of Petit Verdot during 2016 and 2017 (mg/kg FW).
- Supplementral Table S2 Effects of five rootstocks (101-14MGt, 1103P, Beta, 5BB, SO4) on flavonol compounds in mature grapes of Petit Verdot during 2016 and 2017 (mg/kg FW).
- Supplemental Table S3 Effects of five rootstocks (101-14MGt, 1103P, Beta, 5BB, SO4) on flavan-3-ol compounds in mature grapes of Petit Verdot during 2016 and 2017 (mg/kg FW).
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Shi N, Wang W, Li H, Han X, Lu H, et al. 2023. Modifications of Vitis vinifera L. cv. Petit Verdot grape flavonoids as affected by the different rootstocks in eastern China. Technology in Horticulture 3:14 doi: 10.48130/TIH-2023-0014 

Modifications of Vitis vinifera L. cv. Petit Verdot grape flavonoids as affected by the different rootstocks in eastern China
- Received: 02 March 2023
- Accepted: 20 May 2023
- Published online: 14 August 2023
Abstract: Rootstocks are widely used in viticulture because of their strong ability to resist biotic and abiotic stress. In two-successive years (2016−2017), the effects of five rootstocks (101-14MGt, 1103P, Beta, 5BB, and SO4) on the berry flavonoid profiles of Petit Verdot were evaluated by HPLC-MS. The results showed that there was limited influence of rootstocks affecting the technological ripening of Petit Verdot. With respect to flavonoid compounds, 1103P and Beta tended to increase the concentrations of anthocyanin in grape skins in the two seasons. 101-14MGt and 1103P enhanced the flavonol concentrations, whereas Beta showed a detrimental effect on the biosynthesis of flavonols except for syringetin. Additionally, despite the season factor dominating the effect on flavan-3-ols, 101-14MGt tended to show a slight increase in flavan-3-ol concentrations in skins and seeds. In conclusion, based on the evaluation of physicochemical indicators and flavonoid compounds of mature berries, the current study provided data for grafting of Petit Verdot on various rootstocks in eastern China.
-
Key words:
- Graft /
- Berry /
- Flavonoids /
- PCA