








-
Watermelon [Citrullus lanatus (Thunb.) Matsum. & Nakai] has high economic and nutritional value. It is widely grown as an important horticultural crop worldwide[1]. A report by FAOSTAT indicates that, worldwide in 2018, approximately 3.24 million hectares were used for watermelon production (
www.fao.org ). The yield of watermelons in 2018 was 103.93 million tons worldwide and the consumption per person was 13.62 kg, making it among the top five most consumed fresh fruits. China had approximately 1.51 million hectares of watermelon plantation and 43.32 kg of consumption per person in 2018. China is the largest watermelon producer and consumer globally. Demand for protected watermelon cultivation is increasing due to the increased economic benefits and increasing annual consumption of watermelon. In China, the area of protected watermelon cultivation currently accounts for over 50% of the total area used for watermelon cultivation.Watermelon crops are native to tropical and subtropical regions of Africa[2]. They are highly susceptible to low temperatures[3]. The optimum air temperature for watermelon growth and development ranges from 20 to 32 °C. Watermelon growth is inhibited by temperatures below 10 °C. Low temperatures below optimum, but above 0 °C, result in chilling stress[4]. Chilling stress inhibits photosynthesis and respiration, promotes membrane damage, and hormonal imbalance, leading to growth retardation, delay in flowering and fruiting, and results in decreases in total yield and fruit quality[5−7]. Therefore, chilling stress during winter and early spring significantly limits off-season production of watermelons under controlled environments[5].
Continuous mono-cropping of watermelon is widely applied worldwide; however, it results in significant continuous cropping obstacles (also known as the replanting disease). Continuous cropping obstacle is characterized by leaf yellowing, plant stunting, wilting and death, and reduced fruit yield and quality[8,9]. Fusarium wilt is the most common and damaging soil-borne disease that significantly limits watermelon production under a continuous cropping system globally[10,11]. Fusarium wilt is caused by the fungus Fusarium oxysporum f. sp. niveum (FON). Currently, four races (0, 1, 2, and 3) of FON have been described based on their virulence on watermelon genotypes with varying disease resistance[12]. Race 3 is the most virulent, followed by race 2, 1, and 0[13,14]. Race 1 and race 2 have been reported in most of the watermelon growing regions in the world[15−17]. FON is a soil saprophyte and can survive in the chlamydospore state for more than 10 years in the soil in the absence of a host under various environmental conditions. This characteristic of FON limits control and treatment of Fusarium wilt[18]. There is still a shortage of recommended chemical products to control this disease. Control of Fusarium wilt by planting watermelon in areas with low incidence of the disease is a less sustainable practice. Bio-control strategies used for management of the pathogen are not fully effective[19]. Crop rotation is a commonly used strategy for controlling the disease; however, it is limited by the ability of FON to survive without a host for several years[10]. Therefore, improving root resistance is the most cost-effective method for controlling Fusarium wilt.
Grafting on resistant rootstock is a fast, cost-effective and easy method compared with breeding methods and is widely used to control Fusarium wilt[20]. In addition, grafting of watermelon onto cold-tolerant root systems, such as figleaf gourd and pumpkin, improves watermelon tolerance to chilling stress[21,22]. The most commonly used rootstocks for watermelon grafting in China are pumpkin and bottle gourd. However, these rootstocks often cause adverse effects on fruit quality, such as low Brix, increased number of yellowish bands in the red flesh, increased rind thickness, insipid taste, and internal flesh breakdown[23−26]. Currently, demand for better fruit quality has increased due to the development of the economy and increase in population and the desire to improve the living standards of the population. Screening and breeding of rootstocks that alleviate Fusarium wilt and chilling stress, but have no adverse effects on fruit quality, are imperative for protected watermelon production.
Cucurbit germplasms present high genetic variability for resistance against FON and the ability to withstand low temperatures[6,27]. FON-resistant and chilling tolerant watermelon germplasms have good graft compatibility with watermelon and are potentially used as rootstocks[28]. Thick-skin muskmelon (Cucumis melo L.) has a vigorous root system and is relatively resistant to soilborne diseases[29]. However, the effects of use of thick-skin muskmelon as a rootstock for watermelon production have not been fully explored. In the current study, we evaluated the chilling and FON1 resistance of seven Cucurbit germplasms, including watermelon, thick-skin muskmelon, and pumpkin. Moreover, the effects of the use of these germplasms as rootstocks on watermelon resistance to chilling stress and FON1 were explored. Furthermore, the effects of these germplasms as rootstocks on Fusarium wilt, yield, and fruit quality of watermelon grown in greenhouses with continuously cropped soils were evaluated.
-
Chilling treatment at 7/4 °C (day/night) caused leaf wilting, significant decrease in root vitality and significant increase in REC and MDA content in all eight genotypes (Fig. 1). The root vitality of N5, QZ1, M08, TC, Yl, MY, JT1, and JSM germplasms was reduced by 60.04%, 11.78%, 48.63%, 31.03%, 52.08%, 56.41%, 37.31%, and 40.43%, respectively, after chilling treatment compared with the root vitality rates in the control groups. QZ1 germplasm presented the highest root vitality rate followed by TC, JT1, and JSM under chilling stress. N5 exhibited higher REC and MDA content relative to the other genotypes under chilling conditions. QZ1 and JT1 plants had the lowest REC followed by the REC of M08, TC, MY, and JSM germplasms. TC and M08 had the lowest MDA content followed by the MDA contents for QZ1, JSM, JT1, MY, and YL germplasms.
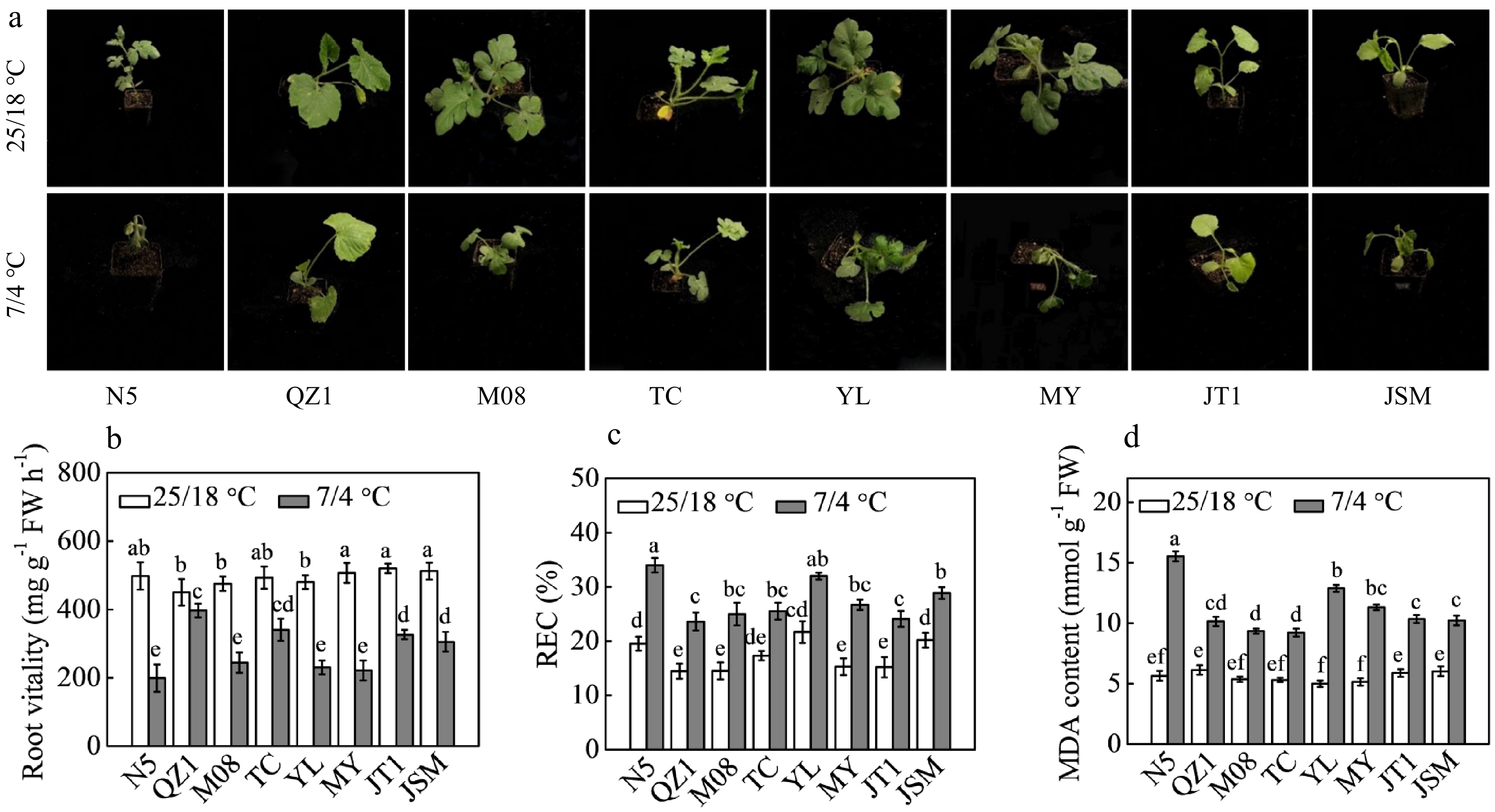
Figure 1.
Responses of eight different Cucurbit germplasms to chilling stress. (a) Plant phenotypes. (b) The maximum photochemical efficiency of PSII (Fv/Fm). (c) Relative electrical conductivity (REC). (d) Malonaldehyde (MDA) content in the eight Cucurbit germplasms. Seedlings at three- or four-leaf stage were transferred into growth chambers maintained at 25/18 °C (day/night) for the control groups or the 7/4 °C (day/night) for chilling treatment groups for 48 h. Data are presented as the means (± SDs) of three replicates. Different superscripts denote significant differences at P < 0.05.
The Pn of N5, QZ1, M08, TC, YL, MY, JT1, and JSM was reduced by 54.36%, 8.86%, 44.85%, 10.64%, 23.72%, 34.51%, 24.66%, and 23.78%, respectively, after chilling treatment compared with that of the respective controls (Fig. 2a). TC germplasm had the highest Pn under chilling conditions followed by QZ1, JSM, and JT1, whereas N5 plants had the lowest Pn. Chlorophyll SPAD value was significantly lower for N5, QZ1, and JSM germplasm after chilling treatment compared with the controls but not in the other groups. Fv/Fm decreased in all eight genotypes after exposure chilling stress. M08, TC, YL, and JT1 plants showed similar and higher Fv/Fm relative to that of the other genotypes. Activities of SOD, POD, and CAT enzymes were significantly increased by chilling stress in all genotypes (Fig. 2b). N5 showed the lowest activities of these enzymes. The highest SOD and POD activities were observed in QZ1 and TC germplasms, whereas TC had the highest CAT activity.
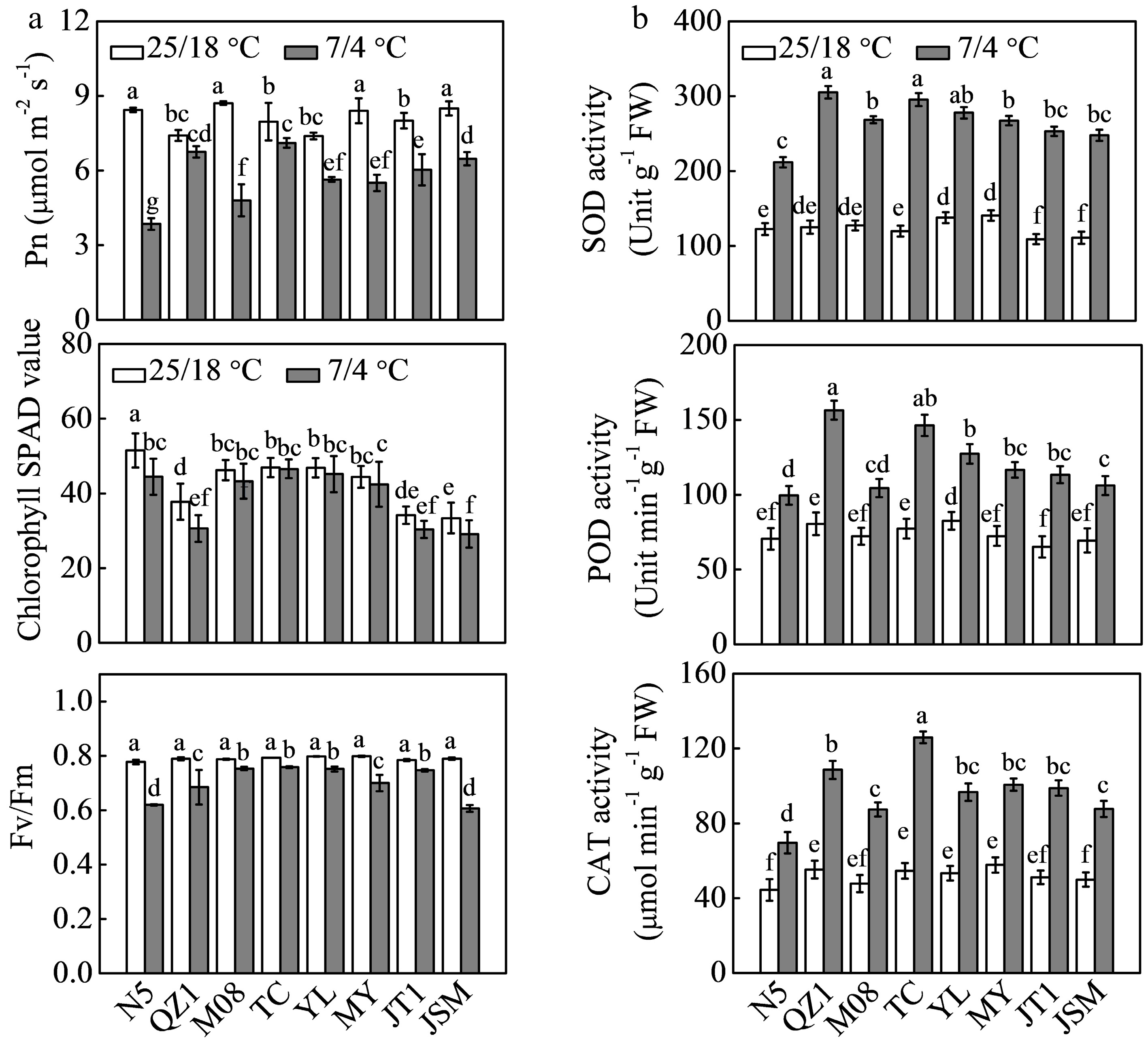
Figure 2.
Effects of chilling stress on (a) photosynthetic ability and (b) antioxidant enzyme activities in eight different Cucurbit germplasms. Seedlings received the same treatments as described in Fig. 1. Data are expressed as the means (±SDs) of three replicates. Different superscripts indicate significant differences at P < 0.05. Pn, net photosynthetic rate; Fv/Fm, maximum photochemical efficiency of PSII; SOD, superoxide dismutase; POD, peroxidase; CAT, catalase.
The membership function values of the eight germplasms based on their variation percentages in growth and physiological indexes are presented in Table 1. The chilling tolerance abilities of the eight genotypes were preliminarily ranked as follows: TC > QZ1 > JT1 > M08 > MY > YL > JSM > N5, based on the total value.
Table 1. Membership function values of eight materials based on relative change rate of growth and physiological indices.
Item N5 QZ1 M08 TC YL MY JT1 JSM Pn 0.00 0.89 0.29 1.00 0.55 0.51 0.67 0.80 SPAD 0.68 0.22 0.76 1.00 0.92 0.77 0.00 0.02 REC 0.00 1.00 0.87 0.81 0.19 0.70 0.95 0.49 MDA 0.00 0.85 0.98 1.00 0.42 0.67 0.82 0.84 Root vitality 0.00 1.00 0.23 0.71 0.16 0.11 0.64 0.54 SOD 0.00 1.00 0.61 0.90 0.71 0.60 0.44 0.39 POD 0.00 1.00 0.09 0.82 0.49 0.30 0.24 0.12 CAT 0.00 0.69 0.31 1.00 0.48 0.55 0.52 0.32 Fv/Fm 0.09 0.53 0.96 1.00 0.95 0.62 0.91 0.00 Average value 0.09 0.80 0.57 0.92 0.54 0.54 0.58 0.39 Tolerance rank 8 2 4 1 6 5 3 7 Data in the table was calculated based on variation in indexes relative to the control samples. The variation percentage (VP, %) for each index was calculated using the following formula: VP (%) = (Tr − CK)/CK × 100%, where Tr represents the value for the given parameter and CK indicates the corresponding value in the control plants. Then, the membership function value of REC and MDA was calculated as 1 − (X − Xmin)/(Xmax − Xmin), whereas that of the other physiological indicators was calculated as (X − Xmin)/(Xmax − Xmin). Effects of different rootstocks on watermelon tolerance to chilling stress
-
Grafted plants including N5/N5, N5/QZ1, N5/M08, N5/TC, N5/YL, N5/MY, N5/JT1, and N5/JSM were exposed to 7/4 °C (day/night) to explore the effects of chilling-tolerant genotypes used as rootstock on watermelon tolerance to chilling. N5/QZ1 and N5/JSM plants showed higher Pn under normal conditions at 25/18 °C (day/night) compared with the other grafted seedlings (Fig. 3). Chilling stress caused significant decrease in Pn and Fv/Fm but increased REC and MDA content in all grafted seedlings. N5/N5 had the lowest Pn and Fv/Fm under chilling conditions, but exhibited the highest REC and MDA content. The highest Pn was observed in N5/QZ1 plants followed by that of N5/TC, N5/JSM, N5/JT1, and N5/YL plants. N5/QZ1 showed the highest Fv/Fm followed by that of N5/JT1, N5/TC, and N5/M08 plants. The lowest REC was observed in N5/QZ1 followed by that in N5/M08, N5/TC, and N5/JT1 plants. N5/QZ1 had the lowest MDA followed by that in N5/JT1 and N5/TC grafted plants.
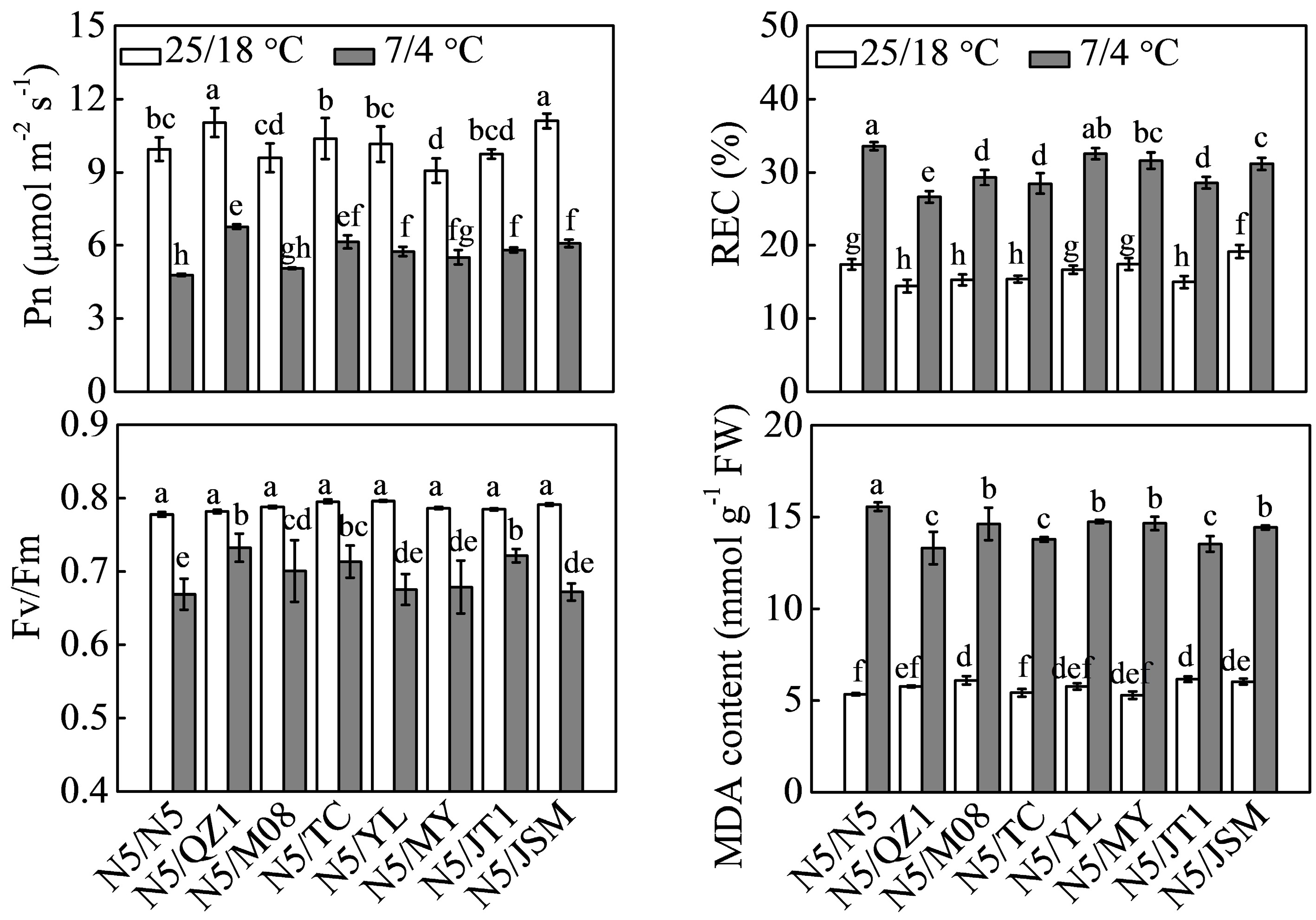
Figure 3.
Effects of different rootstocks on watermelon tolerance to chilling stress. Watermelon plants grafted onto watermelon, pumpkin, and muskmelon rootstocks were subjected to chilling at 7/4 °C (day/night) for 48 h. Different superscripts indicate significant differences at P < 0.05. Pn, net photosynthetic rate; Fv/Fm, maximum photochemical efficiency of PSII; REC, relative electrical conductivity; MDA, malonaldehyde.
Evaluation of FON1 resistance of different Cucurbit germplasms
-
Seedlings were artificially infected with FON1 spores through the root-dip method to explore the resistance of different genotypes to FON1. N5 plants showed the most severe leaf wilting and root rot, the highest fusarium wilt incidence and disease index, but the lowest root vitality after FON1 inoculation for 21 d compared with the other grafted seedlings (Fig. 4). QZ1 exhibited the best growth performance and the highest root vitality. JT1 and JSM groups showed the lowest fusarium wilt incidence and disease index. M08, TC, YL, and MY watermelon germplasms exhibited higher resistance to FON1 relative to that of N5, but lower resistance compared with that of QZ1, JT1, and JSM groups. N5 showed the highest REC and MDA content and the lowest SPAD value after FON1 inoculation compared with the other groups (Fig. 5).
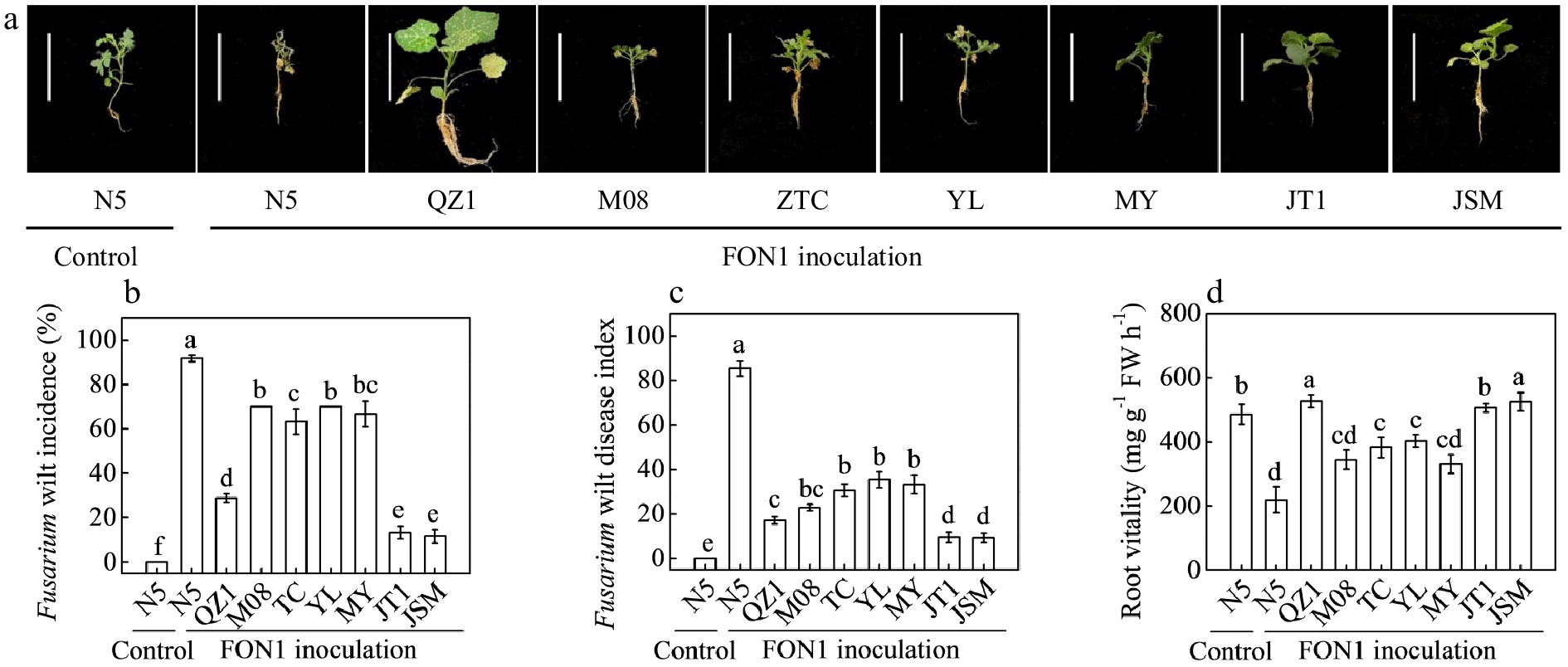
Figure 4.
Responses of eight different Cucurbit germplasms to artificial infection with Fusarium oxysporum f. sp. niveum race 1 (FON1). (a) Plant phenotypes. (b) Fusarium wilt incidence. (c) Fusarium disease index. (d) Root vitality. Seedlings at the two-leaf stage were infected with FON1 using a root-dipping method. Inoculated plants were replanted in sterile peat-based compost for 21 d. In (b−d), different superscripts denote significant differences at P < 0.05.
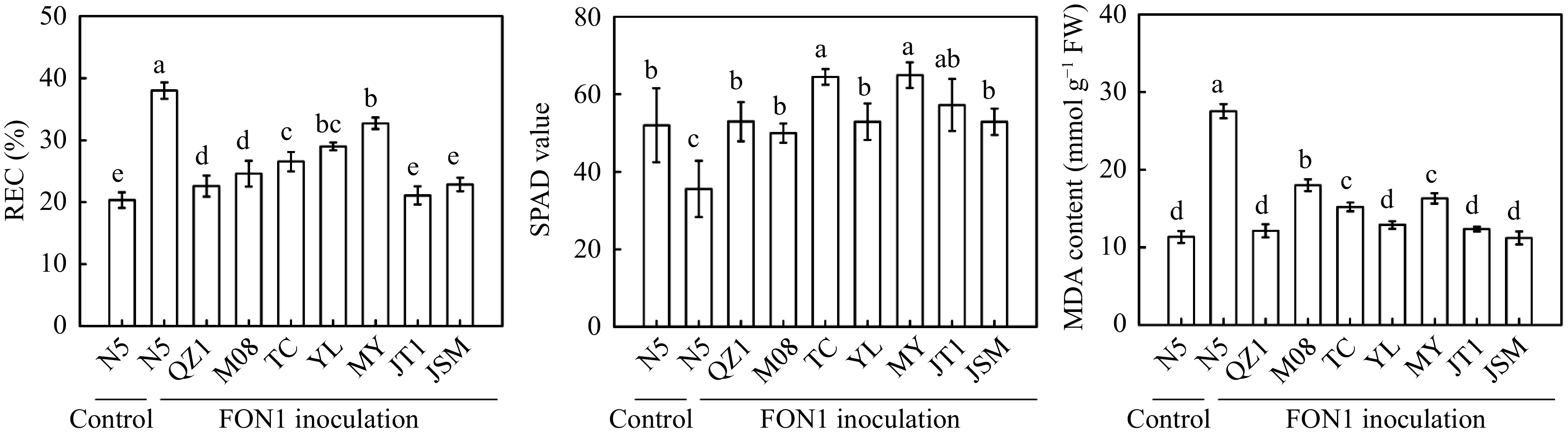
Figure 5.
Effects of artificial infection with Fusarium oxysporum f. sp. niveum race 1 (FON1) on the relative electrical conductivity (REC), chlorophyll SPAD value, and malonaldehyde (MDA) content in eight different grafted watermelons. Seedlings received the same treatments as described in Fig. 4. Data are expressed as the means (± SDs) of three replicates. Different superscripts denote significant differences at P < 0.05.
Effects of different rootstocks on wilt, yield, and fruit quality of watermelon grown on soils continuously cropped for 11 years
-
The grafted plants, including N5/N5, N5/QZ1, N5/M08, N5/TC, N5/YL, N5/MY, N5/JT1, and N5/JSM were planted on soils where watermelons had been continuously cropped for 11 years to evaluate the effects of different rootstocks on watermelon tolerance to successive cropping conditions. The grafted plants especially in the N5/N5 and N5/M08 groups withered or even died except plants in N5/QZ1 and N5/TC groups (Fig. 6). Plants in N5/N5 and N5/M08 groups showed the smallest fruits whereas N5/QZ1 plants had the largest fruits. Yellowish bands in the red flesh were observed in fruits in N5/QZ1 and N5/YL groups. The Fusarium wilt incidence of N5/N5 was higher (96.33%) relative to that of the other grafted plants (Fig. 7). N5/QZ1 plants showed the lowest wilt incidence (2.05%) followed by N5/JT1 (14.85%), N5/YL (40.82%), and N5/TC (40.90%) plants. The economic yield of the eight groups of grafted plants ranged from 1.96 (N5/N5) to 10.16 (N5/QZ1) kg m−2.

Figure 6.
(a) Plant growth and (b) fruit appearance and section plane of eight different grafted watermelons planted in greenhouses where watermelons were continuously mono-cropped for 11 years.
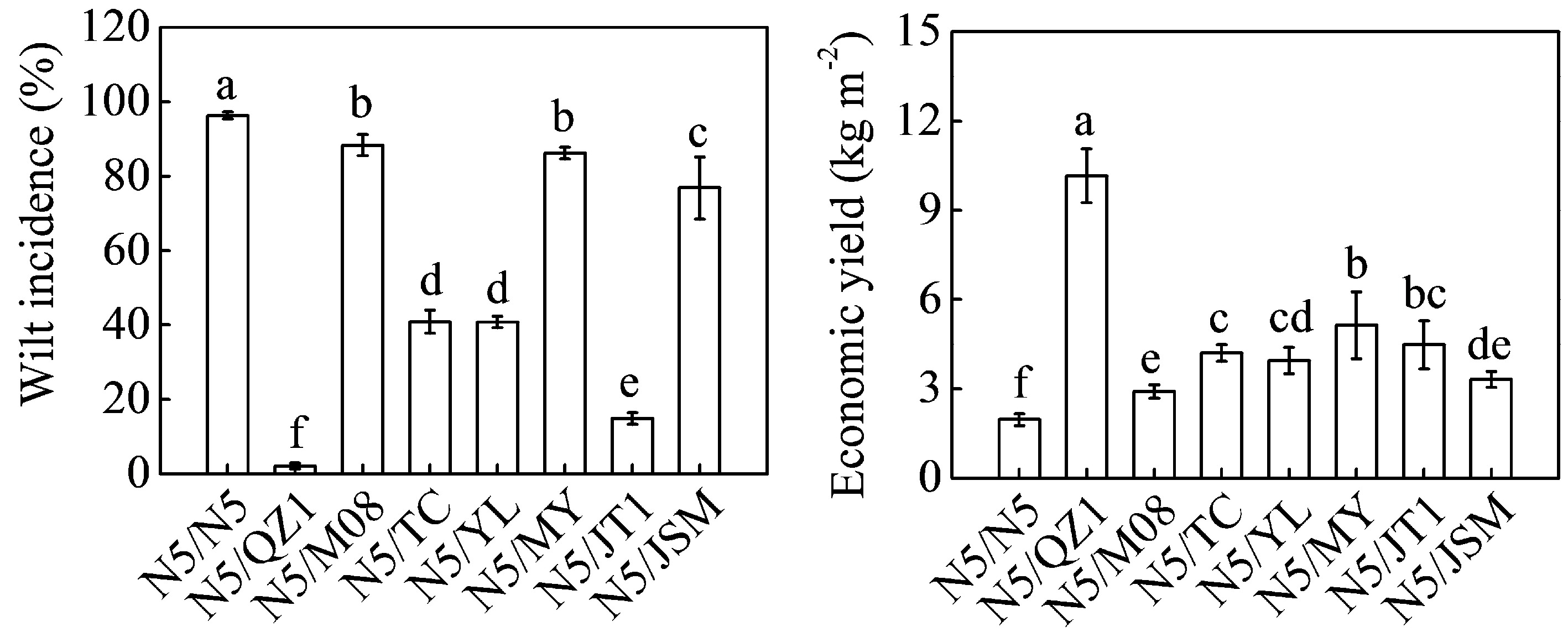
Figure 7.
Wilt incidence and economic yield of eight different grafted watermelons planted in greenhouses where watermelons were continuously mono-cropped for 11 years. Data are presented as the means (± SDs) of three replicates. Different superscripts denote significant differences at P < 0.05.
Analysis of fruit quality showed that fruits of N5/TC, N5/N5, N5/YL, and N5/M08 groups had similar and relatively higher central soluble solid (Table 2). Fruits of N5/N5, N5/QZ1, N5/M08, N5/TC, and N5/YL had similar and relatively higher marginal soluble solid compared with fruits from the other groups. Fruits from N5/N5, N5/M08, N5/TC, N5/MY, and N5/YL plants showed similar and relatively higher lycopene content compared with the other groups. Fruits from N5/M08, N5/TC, N5/JT1, and N5/JSM plants showed relatively higher vitamin C levels compared with the level in other groups. Fruits of N5/N5, N5/M08, N5/TC, and N5/JT1 groups showed relatively higher soluble protein content relative to the levels in the other groups. The fruit quality from different grafted plants was preliminarily ranked as follows: N5/TC > N5/M08 > N5/N5 > N5/JSM > N5/JT1 > N5/YL > N5/MY > N5/QZ1, according to the membership function values of fruit qualities based on their variation percentages (Table 3).
Table 2. Fruit quality of eight different grafted watermelons planted in greenhouses where watermelons were continuously mono-cropped for 11 years.
Grafted plants Central soluble
solid (%)Marginal soluble
solid (%)Reducing sugar
(mg g−1)Lycopene
(mg g−1)Vitamin C
(μg g−1)Soluble protein
(mg g−1)Organic acid
(%)N5/N5 9.47 ± 0.19a 7.51 ± 0.25a 9.6 ± 1.87ab 1.53 ± 0.82ab 3.43 ± 1.06e 0.28 ± 0.07ab 0.12 ± 0.01b N5/QZ1 8.93 ± 0.36b 7.04 ± 0.14ab 8.08 ± 2.26b 0.97 ± 0.14b 4.88 ± 1.12de 0.25 ± 0.04bc 0.12 ± 0.03b N5/M08 9.64 ± 0.60ab 7.80 ± 0.22a 11.21 ± 2.25ab 1.44 ± 0.06ab 8.97 ± 0.97bc 0.33 ± 0.05ab 0.13 ± 0.01b N5/TC 10.11 ± 0.32ac 8.04 ± 0.17a 14.64 ± 4.46ab 1.25 ± 0.20ab 7.74 ± 0.57c 0.41 ± 0.10a 0.11 ± 0.01b N5/YL 9.84 ± 0.17a 7.09 ± 0.23ab 8.66 ± 1.24b 1.03 ± 0.16b 2.93 ± 0.64e 0.12 ± 0.05cd 0.12 ± 0.01b N5/MY 8.89 ± 0.46b 6.36 ± 0.95bb 12.81 ± 1.90ab 1.36 ± 0.23ab 5.84 ± 0.41d 0.25 ± 0.07bc 0.16 ± 0.02a N5/JT1 8.93 ± 0.22b 6.29 ± 0.39b 8.59 ± 4.20b 1.05 ± 0.03b 10.03 ± 1.46b 0.32 ± 0.12ab 0.13 ± 0.03b N5/JSM 8.13 ± 0.09c 6.18 ± 0.17b 11.36 ± 0.82ab 1.66 ± 0.00a 13.62 ± 1.27a 0.08 ± 0.05d 0.14 ± 0.00ab Table 3. Membership function values of eight different grafted watermelons based on the relative change rate of fruit quality indices.
Grafted
plantsCentral
soluble solidMarginal
soluble solidReducing
sugarLycopene Vitamin C Soluble
proteinOrganic
acidAverage
valueQuality
rankN5/N5 0.68 0.72 0.23 0.81 0.05 0.61 0.94 0.60 3 N5/QZ1 0.40 0.46 0.00 0.00 0.18 0.52 0.80 0.35 8 N5/M08 0.76 0.87 0.48 0.68 0.57 0.78 0.63 0.66 2 N5/TC 1.00 1.00 1.00 0.41 0.45 1.00 1.00 0.71 1 N5/YL 0.86 0.49 0.09 0.08 0.00 0.11 0.91 0.34 6 N5/MY 0.38 0.10 0.72 0.56 0.27 0.50 0.00 0.52 7 N5/JT1 0.40 0.06 0.08 0.11 0.66 0.72 0.64 0.44 5 N5/JSM 0.00 0.00 0.50 1.00 1.00 0.00 0.52 0.51 4 Data in the table was calculated based on variation in indexes relative to the control samples. The variation percentage (VP, %) for each index was calculated using the following formula: VP (%) = (Tr − CK)/CK × 100%, where Tr represents the value for the given parameter and CK indicates the corresponding value in the control plants. Then, the membership function value of organic acid content was calculated as 1 − (X − Xmin)/(Xmax − Xmin), whereas that of the other physiological indicators was calculated as (X − Xmin)/(Xmax − Xmin). -
Cucurbitaceae species undergo significant genetic changes to adapt to low temperatures. Cucurbita ficifolia and Cucurbita maxima are chilling-tolerant species, whereas C. sativus, Citrullus lanatus, Cucumis melon, Luffa cylindrica, and Benincasa hispida are chilling-sensitive varieties[3,30,31]. Notably, different germplasms of the same species, such as watermelon, also exhibit great variability in chilling tolerance[6]. The findings of the present study showed that pumpkin (QZ1), watermelon germplasms (TC, YL, M08, and MY), and muskmelon (JT1 and JSM) showed higher tolerance to chilling stress relative to the watermelon cultivar N5 (Figs 1, 2). Moreover, TC variety exhibited higher chilling tolerance compared with QZ1 based on the comprehensive evaluation through membership function analysis (Table 1). These findings show that the wild-type watermelon TC is a potential germplasm source for breeding chilling-tolerant watermelon varieties.
Grafting onto tolerant rootstocks can enhance plant tolerance to various biotic or abiotic stresses such as soilborne diseases, cold, heat, drought, salinity, and flood[29]. Enhancement of plant tolerance by grafting is attributed to inherent resistance of the root systems of rootstocks, as well as to root-to-shoot communication that regulates shoot responses[32]. For instance, findings from our previous study showed that melatonin and ABA, which are root-to-shoot signals, play an important role in grafting-induced watermelon tolerance to cold stress[21,22]. In the current study, grafting onto chilling-tolerant genotypes, especially QZ1, TC, and JT1, improved N5 tolerance to chilling stress (Fig. 3). Therefore, QZ1, TC and JT1 varieties can be utilized as rootstocks to improve watermelon production under low temperatures during winter and early spring.
FON isolates cause disease only on watermelon but not on non-Citrullus varieties, including muskmelon, cucumber, pumpkin, and squash, implying that they are host-specific[12,13]. However, some studies report cross pathogenicity of individual FON isolates within cucurbits, despite the host specificity of FON. For instance, some FON isolates are pathogenic to muskmelon, cucumber, and summer squash[33,34]. The results of the present study showed that non-Citrullus varieties including QZ1, JT1, and JSM exhibited higher resistance to FON1 relative to Citrullus germplasms. Different watermelon genotypes showed varied resistance to FON. PI296341 is resistant to FON races 0, 1 and 2, whereas, watermelon cv. Sugar Baby is FON-susceptible[13,20]. The findings in the current study showed that M08, TC, YL, and MY germplasms exhibited higher resistance to FON1 relative to the N5 variety (Figs 4, 5). This finding indicates that these germplasms can be used for breeding watermelon rootstocks with high resistance to Fusarium wilt and can serve as tools for studying FON resistance mechanisms and the genetics of disease resistance.
Interspecific grafting whereby the rootstock and the scion are derived from different species is common in cucurbits[20]. Most of the watermelons grown in China, especially those under protected cultivation, are grafted mainly on Cucurbita rootstocks (F1 hybrids of C. moschata × C. maxima). These rootstocks have high root growth rate and high resistance to a wide variety of soilborne pathogens and some abiotic stresses[35]. However, grafting onto Cucurbita rootstocks can result in undesirable fruit characteristics such as low Brix, less crispy and harder fruit flesh, increased white fibers or yellowish bands in the red flesh, changes in fruit size, fruit deformation, increased rind thickness, insipid taste, and internal flesh breakdown reducing fruit quality[23−26]. These features are more pronounced in mini watermelon cultivars[36,37]. The results of the present study showed that QZ1 rootstocks increased resistance to wilt and fruit yield, and significantly reduced fruit quality of watermelons grown on soils continuously cropped for 11 years (Figs 6, 7; Tables 2, 3). Muskmelon (JT1 and JSM) rootstocks showed similar effects as QZ1 rootstocks. The low quality of these varieties can be attributed to incompatibility between Cucurbita or muskmelon rootstocks and watermelon.
Intraspecific grafting that involves grafting watermelon on watermelon rootstocks can alleviate reduction of fruit quality caused by interspecific grafting due to the high compatibility between the scion and rootstock of watermelons[23,25]. Use of M08 and TC germplasms as rootstocks increased the resistance to wilt and fruit yield and improved fruit quality of watermelons grown on soils continuously cropped for 11 years (Figs 6, 7; Tables 2, 3). Although TC or M08 rootstock-induced wilt resistance and fruit yield were lower relative to those for plants grafted from QZ1 rootstock, the two varieties can be utilized for watermelon production on continuously cropped soils for < 10 years.
-
In summary, the seven rootstock materials explored in this study showed higher resistance to chilling stress and FON1 compared with the N5 variety. TC plants showed the highest tolerance to chilling stress, followed by QZ1, JT1, M08, MY, YL plants. Use of JSM, TC, QZ1, and JT1 as rootstocks increased chilling tolerance of watermelon scion. Watermelons grafted onto QZ1 rootstock showed the lowest wilt incidence, highest fruit yield, but the worst fruit quality when grown on soils continuously cropped for 11 years. On the contrary, watermelons grafted onto TC showed the best fruit quality although their wilt incidence and the fruit yield were higher and lower relative to that of N5/QZ1 plants, respectively. TC was the best variety for grafting watermelon production on continuously cropped soils for < 10 years. Notably, QZ1 was the best variety for grafting watermelon grown on continuously cropped soils for over 10 years to improve fruit yield and Fusarium wilt resistance.
-
Eight cucurbit germplasms were used in this study. An early-maturing watermelon cultivar (Nongkeda No. 5, N5) with medium-sized fruit (~5.9 kg per fruit) was used as a control. Qingyan Zhenmu No. 1 (QZ1) is a commercial and widely used pumpkin (F1 hybrids of Cucurbita maxima × C. moschata) rootstock variety for commercial watermelon production. TC is a wild-type watermelon variety[38]. M08 and YL are inbred watermelon lines with high resistance to FON1. MY is a F1 hybrid of M08 × YL. Jingtian No. 1 (JT1) and Jiashi muskmelon (JSM) is a commercial muskmelon cultivar. Watermelon germplasm seeds were provided by the Watermelon and Melon Research Group at Northwest A&F University (Yangling, China). QZ1 seeds were provided by Qingdao Agricultural University (Qingdao, China). JSM seeds were provided by the Xinjiang Academy of Agricultural Sciences (Wulumuqi, China). All seeds were sterilized with 2% sodium hypochlorite for 10 min, pre-soaked at 25 °C for 4−6 h, and maintained under dark conditions at 30 °C until germination. Germinated seeds were planted in plastic pots (7 cm × 7 cm × 7.5 cm) filled with commercial peat-based compost. The seedlings were maintained at 25/18 °C (day/night), a photosynthetic photon flux density of 500 μmol m−2 s−1, a 12-h photoperiod, and 70%−80% constant relative humidity.
Seedlings at three- or four-leaf stage were transferred into growth chambers maintained at 25/18 °C (day/night) for the control or 7/4 °C (day/night) for chilling treatment for 48 h to evaluate the chilling tolerance of different germplasms. Each treatment comprised three independent biological replicates. Each replicate consisted of 15 plants. Seedlings at two-leaf stage were inoculated with FON1 conidial using a root-dipping method to evaluate FON1 resistance of different germplasms. Each treatment was conducted in triplicate with 20 plants per replicate.
Germinated seeds of rootstocks were planted 7 d before planting the seeds for scions (N5) for grafting. Top insertion grafting was performed when the scion cotyledons expanded[29]. The resulting eight seedling groups were designated N5/N5, N5/QZ1, N5/M08, N5/TC, N5/YL, N5/MY, N5/JT1, and N5/JSM according to the rootstock species. N5/N5 plants were used as control group. After grafting, the seedlings were maintained at 26−30 °C, a photosynthetic photon flux density (PPFD) of 55 μmol m−2 s−1, and 90%–100% constant humidity for 7 d.
Seedlings at three- or four-leaf stage were exposed to 7/4 °C (day/night) for 48 h to explore the chilling tolerance of different grafted plants. Each treatment was performed in triplicates with 15 plants per replicate.
Grafted seedlings with two or three true leaves were transplanted into greenhouses, whereby watermelons had been continuously mono-cropped for 11 years, at Yangling, China to evaluate the effects of different rootstocks in alleviating continuous cropping obstacle of watermelon. The experiment comprised three replicates per treatment with 20 plants per replicate.
Analysis of gas exchange, chlorophyll fluorescence, and chlorophyll content
-
The net photosynthetic rate (Pn) was determined using a LI-6400 portable photosynthesis system (Li-6400; Li-Cor, Lincoln, NE, USA) under the following conditions: a temperature of 25 °C, CO2 concentration of 380 μmol mol−1, a relative humidity of 80%, and a PPFD of 500 μmol m−2 s−1.
An imaging pulse amplitude modulated (PAM) chlorophyll fluorometer was used for determination of chlorophyll fluorescence (Heinz Walz, Effeltrich, Germany) following a method described by Li et al.[39]. The plants were kept under dark conditions at 25 °C for 30 min before recording the parameters. The maximal quantum yield of photosystem (PS) II (Fv/Fm) was directly recorded.
Chlorophyll content in leaves were assayed using a Chlorophyll Meter Reading (Soil plant analysis development, SPAD-502 Plus, Japan).
Analysis of root vitality
-
A triphenyltetrazolium chloride method (TTC)[40] was used for root vitality determination. Fresh roots (0.3 g) were cut into 2-mm-long fragments and placed in 6 mL 0.06M Na2HPO4–KH2PO4 containing 0.6% (w/v) TTC at 37 °C for 1.5 h. Subsequently, 0.05% (v/v) Tween 20 was added and the samples were vacuum-infiltered for 15 min. Samples were washed twice with 5 mL of distilled water then extracted in 95% (v/v) ethanol at 80 °C for 15 min. Absorbance was then recorded at 520 nm.
Analysis of malondialdehyde content and relative electric conductivity
-
Malondialdehyde (MDA) is a byproduct of lipid peroxidation and its level is an index of lipid peroxidative damage. MDA content was determined using 2-thiobarbituric acid following a method reported by Hodges et al.[41]. Relative electric conductivity (REC) was determined according to the method described by Zhou & Leul[42].
Antioxidant enzyme activity assays
-
A total of 0.3 g leaf samples was ground with 3 mL 25 mm HEPES buffer (pH 7.8) containing 2 mM ascorbic acid, 0.2 mM EDTA, and 2% polyvinyl pyrrolidone on ice. The samples were centrifuged at 12,000 g for 20 min at 4 °C and the resulting supernatants were used for determination of enzyme activity. Superoxide dismutase (SOD) activity was determined according to the method reported by Stewart & Bewley[43] based on photochemical reduction of NBT. Peroxidase (POD) activity was evaluated following a method described by Cakmak & Marschner[44]. Catalase (CAT) activity was determined by monitoring the decrease in A240 following a method reported by Patra et al.[45].
FON1 inoculation
-
FON1 was isolated from infected watermelon plants from the field and cultured on potato dextrose agar at 28 °C for 5−7 d. Inoculation was performed following a root-dip method as described by Li et al.[46]. Seedlings at two-leaf stage were carefully obtained from the soil and washed under running tap water. The roots were cut 1−2 cm from the root tip and then dipped in a spore suspension (106 conidia mL−1) of fungal conidia for 15 min. Control plants were dipped into sterile distilled water. Inoculated plants were replanted in sterile peat-based compost. The inoculated plants were grown under a temperature of 24−28 °C and a PPFD of 100 μmol m−2 s−1. The PPFD was set at 500 μmol m−2 s−1 after day 2. The resulting disease response of plants was rated on a scale of 0 to 5. The number of wilted plants was recorded and disease incidence was calculated after inoculation for 21 d[46].
Analysis of fruit quality and economic yield
-
Central and marginal soluble solid was determined using a digital refractometer (TD-45, Top Co. Ltd., Zhejiang, China). Reducing sugar content was determined following a 3,5-dinitrosalicylic acid method as previously described by Lindsay[47].
Lycopene content was determined according to a method reported by Adsule & Dan[48]. A sample of 0.2 g flesh fruit was ground with 5 mL acetone. Extraction was performed at room temperature for 30 min and the homogenates were centrifuged at 12,000 g for 5 min. Lycopene content was then determined by recording absorbance at 505 nm using a spectrophotometer.
Vitamin C (ascorbic acid) content was determined according to a method described by Law et al.[49]. A sample of 0.3 g flesh fruit was homogenized using 6% (w/v) trichloroacetic acid (TCA). Samples were centrifuged at 12,000g for 15 min and 400 μL 44% phosphoric acid (v/v), 400 μL 10% (w/v) TCA, 400 μL 70% (v/v) α’-dipyridyl in ethanol, and 200 μL 3% (w/v) FeCl3 were added to the supernatants. The mixture was incubated at 37 °C for 1 h and the absorbance recorded at 525 nm.
A total of 0.3 g flesh was ground in 3 mL ice-cold 25mM HEPES buffer (pH 7.8) containing 2% PVP and 0.2 mM EDTA for extraction of soluble protein. The samples were centrifuged at 12,000 g for 15 min at 4 °C and the protein contents were determined as described by Bradford & Williams[50]. Organic acid content was determined through titration with NaOH according to a method reported by Mitchell et al.[51].
The total number of fruits and weights of fruits were recorded for each harvest. Economic yield indicates the yields of fruits above 1.5 kg.
Membership function analysis
-
Chilling tolerance of different germplasms and fruit quality of different grafted watermelon plants were comprehensively evaluated using the membership function method reported by Zhang et al.[18]. The membership function value was calculated based on variation in indexes relative to the control samples. The values were then expressed as variation percentage (VP, %). The VP for each index was calculated using the following formula: VP (%) = (Tr − CK)/CK × 100%, where Tr represents the value for the given parameter and CK indicates the corresponding value in the control plants[52]. The membership function value was calculated as (X − Xmin)/(Xmax − Xmin) if the VP of a tested index was positively correlated with chilling tolerance or fruit quality. On the contrary, the membership function value was given by the formulae: 1 − (X − Xmin)/(Xmax − Xmin) if the VP of a tested index was negatively correlated with chilling tolerance or fruit quality. In both functions, X represents the VP of a given index in a certain material, and Xmin and Xmax represent the minimum and maximum VPs, respectively, for the given index in all examined materials. An average membership function value for the VPs of all indexes was then calculated and used to determine the chilling tolerance or fruit quality of each material.
Statistical analysis
-
All experiments were performed in a completely randomized design. Data were analyzed using analysis of variance (ANOVA) followed by Tukey’s test. P values < 0.05 were considered statistically significant.
The work was supported by the National Key Research and Development Program of China (2018YFD1000800), the National Natural Science Foundation of China (31972479), the Earmarked Fund for China Agriculture Research System (CARS-25), the Science and Technology Innovation Team of Shaanxi (2021TD-32), and the Tang Scholar of Northwest A&F University.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Kai Xu, Zhixiang Lan
- Copyright: © 2023 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Xu K, Lan Z, Liu Q, Xue Y, Yan J, et al. 2022. Screening of rootstocks with resistance to chilling and continuous cropping but without compromising fruit quality for protected watermelon production. Vegetable Research 2:10 doi: 10.48130/VR-2022-0010 

Screening of rootstocks with resistance to chilling and continuous cropping but without compromising fruit quality for protected watermelon production
- Received: 28 March 2022
- Accepted: 15 August 2022
- Published online: 26 August 2022
Abstract: Chilling stress and continuous cropping obstacles limit sustainable production of watermelons under controlled environments. Grafting of watermelon scions onto resistant rootstocks is an effective strategy currently used to overcome these environment limitations. However, currently used commercial rootstocks adversely affect watermelon fruit quality. The chilling tolerance and Fusarium oxysporum f. sp. niveum race 1 (FON1) resistance of seven Cucurbit germplasms, including four watermelon germplasms (M08, TC, YL, and MY), two muskmelon cultivars (JT1 and JSM), and one commercial Cucurbita rootstock (QZ1) of watermelon, were explored in the current study. The effects of the rootstocks of these germplasms on watermelon resistance to chilling stress and continuous cropping obstacles were evaluated. TC rootstock showed the highest chilling tolerance and increased chilling tolerance of watermelon scion. All Cucurbit germplasms showed higher resistance to FON1 than watermelon cultivar N5 (control). Watermelons grafted onto QZ1 showed the lowest wilt incidence and highest fruit yield but had the worst fruit quality after planting on soils continuously cropped for 11 years. Watermelons grafted onto TC showed higher resistance and yield and the best fruit quality. These findings indicate that TC has a large potential for use in grafting watermelon planted in continuously cropped soils (< 10 years). TC can also be used as breeding rootstocks to improve watermelon resistance against continuous cropping obstacles without compromising fruit quality.
-
Key words:
- Grafting /
- Rootstock /
- Chilling tolerance /
- Continuous cropping resistance /
- Fruit quality /
- Watermelon