








-
Anthocyanins belong to the most relevant group of flavonoids, possessing a 2-phenyl-1-benzopyrylium skeleton with hydroxyl and methoxyl moieties that render them with the most prominent antioxidant activity. They are water soluble pigments present in vacuoles of plant cells, which contribute to red, purple, and blue colour on leaves, roots, flowers, fruits and seeds[1]. They are synthesized in glycosylated forms, in which the anthocyanidin molecule is coupled with a sugar[2,3]. Anthocyanins make a substantial nutritional contribution to the human diet and health because of their antioxidant activity[3]. The biosynthesis pathway of anthocyanins is conserved among higher plants and has been well characterized in Solanaceous vegetables[4]. The structural genes in anthocyanin biosynthesis pathway are categorised into two groups, namely early and late biosynthetic genes (EBGs and LBGs) (Fig. 1)[4]. Among the two groups, the LBGs specifically control the biosynthesis of limited flavonoids including anthocyanins[5]. The pathway for anthocyanin biosynthesis starts with phenylpropanoid metabolism that involves three key enzymes, phenylalanine ammonia-lyase (PAL), cinnamate 4-hydroxylase (C4H) and 4-coumarate:CoA ligase (4CL), and the biosynthesis of 4-coumaroyl-CoA that triggers the flavonoid metabolism. The 4-coumaroyl-CoA undergoes condensation with three malonyl-CoA producing naringenin chalcone (Fig. 1). This step is catalysed by chalcone synthase (CHS), the foremost enzyme in flavonoid biosynthesis (Fig. 1). The chalcone scaffolds thus formed contain two phenyl rings which are precursors to many flavonoids. The naringenin-chalcone is converted to the flavanone called naringenin under the catalytic action of chalcone isomerase (CHI) (Fig. 1). Subsequently, a hydroxyl group is added to naringenin to produce dihydroflavonols, under the activity of flavanone-3-hydroxylase (F3H) (Fig. 1). The anthocyanidin molecules that exist in sugar free state are ultimately formed from dihydroflavonol through the catalytic action of dihydroflavonol 4-reductase (DFR) and anthocyanidin synthase (ANS) (Fig. 1). Finally, the enzymes anthocyanidin 3-O-glucosyltransferase (UFGT) modifies the structures with glucose to form their glycosidic forms or anthocyanins (Delphinidin, Malvidin, Petunidin-Glycosides)[6]. Apart from the structural genes, there are few transcription factors (TFs) identified to control anthocyanin biosynthesis. The TFs include two members of the R2R3-MYB family (ANT1 and AN2-like), two bHLH proteins (AN1 and JAF13), and a WD40 protein (AN11) (Fig. 1)[7−10]. The MBW (MYB–bHLH–WDR) complex formed from the combination of the above three proteins regulates the anthocyanin biosynthetic genes, which in turn are governed by internal (hormones) and external signals (biotic or abiotic factors) that control redox reaction[11]. The transcriptional control of anthocyanin biosynthetic genes dictates the final level of secondary metabolite anthocyanins.
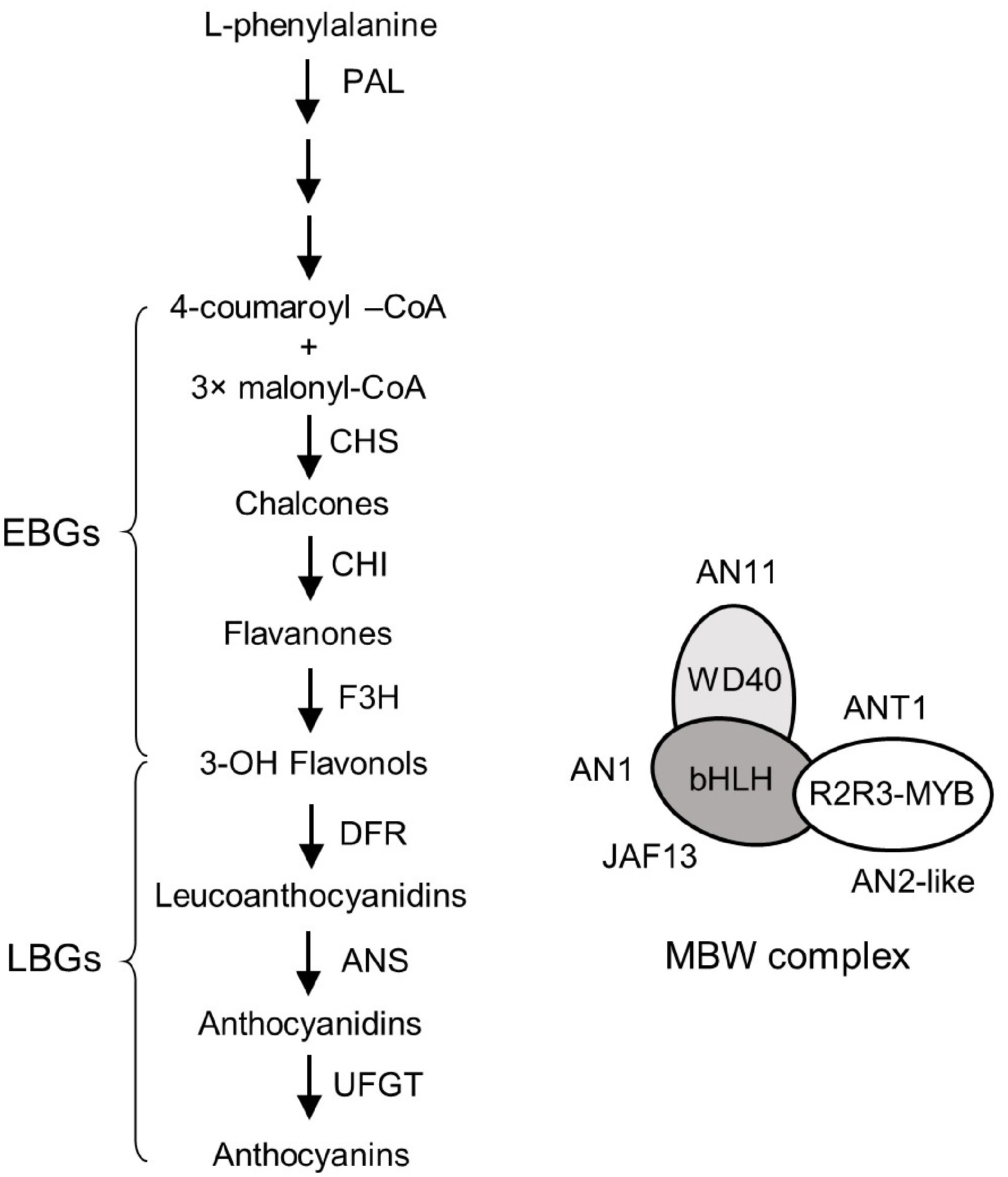
Figure 1.
Anthocyanin biosynthetic pathway and its regulatory genes in tomato. The anthocyanin biosynthetic pathway (ABP, left panel) consists of six core structural genes (CHS, CHI, F3H, DFR, ANS and UFGT) that can be divided into early biosynthetic genes (EBGs) (CHS, CHI and F3H) and late biosynthetic genes (LBGs) (DFR, ANS and UFGT). The phenylalanine ammonia lyase (PAL) gene, which is upstream from the ABP, is included in this figure. The identified regulatory genes of ABP encodes six transcription factors, including two members of the R2R3-MYB family (ANT1 and AN2-like), two bHLH proteins (AN1 and JAF13) and a WD40 protein (AN11), and they form a MBW complex (right panel).
Tomato (Solanum lycopersicum) is a juicy fruit vegetable, rich in secondary plant metabolites, predominantly carotenoids and phenolic compounds. However, owing to the partial activation of the flavonoid pathway, anthocyanins are generally absent in cultivated tomato varieties[5]. Anthocyanin-enriched tomatoes were obtained by employing breeding or transgenic approaches[7,12−16] . In recent years, the tomato cultivar ‘Indigo Rose’ gained attention due to its purple-skinned fruits rich in anthocyanins[17]. Indigo Rose was bred from the wild tomato species through the introgression of the Anthocyanin fruit (Aft) locus from Solanum chilense and the atroviolacium (atv) locus from Solanum cheesmaniae into cultivated tomato[12,13]. The Aft gene encodes an R2R3 MYB transcription factor named AN2-like that positively regulates anthocyanin biosynthesis in a light-dependent manner[7,18]. The dominant allele of the Atv gene encodes an R3 MYB repressor MYBATV that negatively regulates anthocyanin biosynthesis, whereas the recessive atv allele in Indigo Rose encodes a non-functional version[7,19,20]. The anthocyanin content in Indigo Rose fruits is mainly restricted to their fruit peels[7,21].
Singapore has a hot and humid tropical climate with abundant rainfall throughout the year, which is unfavourable for growing tomato in an open field. The tomato cultivar Indigo Rose is relatively new to the tomato plantation. It was developed by Oregon State University (USA), where the climate is milder in summer than in Singapore[17]. Hence, in the present study, we grew Indigo Rose in greenhouses (GH) to test its adaptability to the local climate in Singapore. In the meantime, we compared anthocyanin content and the expression of anthocyanin biosynthetic and regulatory genes in fruits grown under GH and growth chamber (GC) growth conditions. We then investigated the influence of light and temperature on anthocyanin content and gene expression in Indigo Rose under GC growth conditions and sought optimal light and temperature to boost anthocyanin production in Indigo Rose in indoor farming.
-
The seeds of the tomato cultivar Indigo Rose were purchased online (
www.plant-world-seeds.com/store/vegetable_seeds/tomato_seeds ). Tomato plants were grown in GCs and GHs, respectively. The day length in Singapore is about 12 h. The length of direct sunlight in the GHs was about 9 h with light intensity from 200 to 900 µmol m−2 s−1. The temperature at the GHs ranged from 25 °C at night to 35 °C in the early afternoon on a sunny day. The relative humidity in the GHs was at about 80%−85%, which was similar to the open environment in Singapore. The growth conditions in the GCs were 16-h light with a temperature of 25 °C and relative humidity of 65%. To test the effect of light intensity and room temperature on anthocyanin biosynthesis and gene expression in tomato fruits, tomato plants were allowed to grow in the GCs with light intensity at 80 µmol m−2 s−1 (Low), 160 µmol m−2 s−1 (Moderate) or 240 µmol m−2 s−1 (High), and temperature at 20 °C, 25 °C or 30 °C, respectively.Measurement of total anthocyanins in tomato fruits
-
Tomato fruits at about 5 and 8 weeks post anthesis, which were at the mature green (MG) and red ripe (RR) stages, respectively[22], were harvested for anthocyanin content measurement. The fruit peel and flesh were separated and weighed. Samples were frozen in liquid nitrogen and stored at −80 °C. The frozen samples were manually ground into powder using a pestle and mortar and transferred into 50-ml centrifuge tubes. Anthocyanin extraction was performed according to the published protocol with minor modifications[21]. Briefly, in each tube containing the powdered sample, anthocyanins were extracted by adding 500 μl of acidic methanol (1% v/v HCl) per 100 mg fresh weight (FW) of the frozen tissues and vortexed to mix well. The tubes were wrapped with aluminium foil and incubated at 4 °C overnight. After 12 h of extraction, the samples were centrifuged at 3,000 rpm for 2 min and then 500 μl of supernatant was collected. Next, 250 μl of distilled water and 250 μl of chloroform were added to the extract and vortexed. The mixture was centrifuged at 3000 rpm for 2 min at room temperature. Lastly, 200 μl of supernatant containing anthocyanins was pipetted into a 96-well plate and absorbance was measured spectrophotometrically (Spark®, TECAN, CHE) at 535 nm and 650 nm. Anthocyanin quantification was performed using the following equation: Q = (A535 − A650) / FW (100 mg) [Q = total anthocyanins; A535 = absorption at 535 nm; A650 = absorption at 650 nm; FW = fresh weight of fruit tissues (100 mg)]. Data for each sample were presented as the mean of triplicate.
Total RNA extraction, cDNA synthesis and qRT-PCR
-
The peel and flesh tissues of tomato fruits at the MG and RR stages were separated and stored in liquid nitrogen. Total RNA was isolated from fruit samples utilizing the FavorPrep Total RNA Isolation Mini Kit (FATRK001-1; Favorgen). In brief, the frozen fruit tissues were ground using a pestle and mortar. The total RNA was extracted from about 100 mg powdered samples. The concentration of RNA and its quality (260/280 ratio, 260/230 ratio) were checked using a Nanodrop spectrophotometer (ND-1000 from NanoDrop Technologies USA). The cDNA was synthesized from 1 µg of total RNA by using the iScriptTM cDNA synthesis kit (Bio-Rad, USA). The cDNA samples were utilized as the templates for quantitative real-time PCR (qRT-PCR) analysis on a real-time PCR system (Bio-Rad Laboratories, CFX96). The qRT-PCR mixtures included cDNA, gene-specific oligonucleotide primers (Table 1) and SYBR FAST qPCR Master Mix (2X) (KAPA Biosystems). The DNA sequences of all the genes involved in anthocyanin biosynthesis were downloaded from the NCBI and the Solanaceae Genomics Network (
https://solgenomics.net ). The gene-specific primers were designed using the primer QuestTM tool from Integrated DNA Technologies (https://sg.idtdna.com/Primerquest/Home/Index ). The qRT-PCR reactions were conducted with the following settings: 95 °C for 3 min followed by 40 cycles of 95 °C for 10 s, 60 °C for 30 s, and 65 °C for 10 s. The cycle threshold value (CT) was analysed using CFX ManagerTM software. The average CT was used for the expression analyses. The expression levels for the respective genes were normalized to the tomato Actin gene (Accession number: Solyc03g078400). The normalized expression was determined using with the comparative CT method (ΔΔCT)[23]. The genes of interest were measured with three replicates in each PCR run.Table 1. DNA oligo primers for qRT-PCR analysis.
Gene Accession No. DNA oligo primer (5' to 3') Actin Solyc03g078400 F: TGGATTGGAGGATCCATTCTTG R: CCTTAGAAGCACTTTCTGTGGA AN1 Solyc09g065100 F: GTGGAAGTTGTAGCAATCCAATC R: CACTTCCAATATGCTTGCCTTC AN11 Solyc03g097340 F: GGTTGGCTTGGAACAAACAGC R: GCCCAAGCAATAGCATTCAC AN2-like Solyc10g086290 F: GAGGAAGAAGCAACAGCAGTAT R: CTCCATGTTGCATGGTTGTTG ANS Solyc08g080040 F: TGTTGCTGAGCACATAGTCC R: CTGCTCATCCTCCTTGAGAAC ANT1 Solyc10g086260 F: CAACGTAGATCAATGGTGGATAA R: GCATGGAGTTACCTTCACCA CHI Solyc05g010320 F: CTCAATCACCGCTTGGATCA R: CTCCGTGCTCGCCAATTAT CHS Solyc09g091510 F: GGCCTAGGAACTACTGGTGAA R: TGTAAGCCCAGCCCACTAA DFR Solyc02g085020 F: TGCTCGTCCCATCATGCTATC R: CACTACGGGCAAGTCCTTATC F3H Solyc02g083860 F: TTCAGCCCGTTGAAGGTG R: TGGATTCTGGAATGTGGCTATC JAF13 Solyc08g081140 F: CTTCCAACCTTGATGGGATTCTA R: CATACTACTCTCTGAAGTGCTTCTC PAL NM_001320609.1 F: GATCCATTGTTGGAGTGTCTCA R: GCACTAGACAAGCATTCACAAC UFGT Solyc10g083440 F: GATGGAACTCGATTCTGGAAGG R: CCACCTTCAATTTGCAAACCA F, forward primer; R, reverse primer. Statistical analysis
-
Data of anthocyanin content and qRT-PCR analysis were analysed by one-way ANOVA followed by comparisons of means using Tukey’s test at p < 0.05. Statistical analysis was performed by the software of IBM SPSS Statistics 19.
-
Indigo Rose is a tomato variety with fruits rich in anthocyanins[17]. To test anthocyanin biosynthesis and accumulation in fruits, Indigo Rose plants were initially planted in the GHs. The GHs used in this study was a semi-closed plant growth system with sunlight illumination and equipped with electric fans for air ventilation. As no cooling system was installed, the temperature in the GHs ranged from 25 °C at night time to 35 °C in the early afternoon on sunny days, which was similar to that in the open field. The semi-closed GHs were used to test the adaptation of Indigo Rose to a tropical climate with warm temperature, short-day photoperiod and high humidity. Indigo Rose plants exhibited normal vegetative growth and development in the GHs. However, they produced a significant proportion of small and seedless fruits with a dark red colour at the reproductive stage (Fig. 2a & b). The small and seedless fruits might have developed through parthenocarpy, possibly due to the high temperature stress[24]. The flowers on the Indigo Rose plants grown in the GHs had protruding stigmas, exhibiting stigma exsertion (Fig. 2a). The stigma exsertion might hamper pollination and cause failure in setting normal fruits. To investigate if this is the main reason that led to the production of parthenocarpy fruits, the flowers with exserted stigmas were then artificially pollinated with pollen collected from the flowers of Indigo Rose plants grown in the GHs. Most of the artificially pollinated flowers still could not develop normal fruits and produced only parthenocarpic fruits. However, the flowers with exserted stigmas could produce normal fruits when they were artificially pollinated with pollen collected from the flowers of Indigo Rose grown in the GCs with temperature set at 25 °C (Fig. 2a). The results of the artificial pollination study suggest that the high temperature in the GHs might have caused the defect in pollen development in Indigo Rose, whereas the development of female reproductive organs was not affected. The infertile pollens could not perform normal pollination and the unfertilized ovules developed into parthenocarpic fruits. Indigo Rose plants grown in the GCs developed uniform medium-sized dark purple or black fruits and had higher fruit yield than those grown in the GHs (Fig. 2c). The fruits developed under the GC growth conditions contained normal seeds (Fig. 2d). The flowers developed under the GC growth conditions had normal flower structure with stigma recessed within their anthers (Fig. 2c). The plantation tests under the GH and GC growth conditions demonstrate that Indigo Rose is not a thermotolerant variety and couldn’t adapt well to the warm tropical climate; however, it is a good candidate variety for indoor plantation in urban farming.

Figure 2.
Fruits developed on Indigo Rose plants grown in GH and GC. (a) Fruits developed on Indigo Rose plants grown in GH and cross section of flower with exserted stigma. Black and red arrows indicate normal and parthenocarpic fruits, respectively. (b) Cross section of a parthenocarpic Indigo Rose fruit grown in GH. (c) Fruits developed on Indigo Rose plants grown in GC and cross section of flower with recessed stigma. (d) Cross section of a normal Indigo Rose fruit grown in GC.
Anthocyanin biosynthesis is enhanced in Indigo Rose fruits grown in growth chambers than in greenhouses
-
The anthocyanin biosynthesis in Indigo Rose is light-dependent and mainly synthesized and stored in fruit peels[7]. The fruits set on Indigo Rose plants grown in the GHs or GCs displayed a significant difference in peel colour. Under the GH growth conditions, Indigo Rose fruits at the MG stage were a light purple and green colour, whereas the fruits at the RR stage were a mahogany and wine colour (Fig. 3a). Under the GC growth conditions, both MG and RR fruits displayed dark colour ranging from dark purple to black, suggesting that more anthocyanins were produced and accumulated in these fruits (Fig. 3a). Total anthocyanin content was then determined in the peels and flesh of Indigo Rose fruits. The anthocyanin accumulation occurred mainly in the fruit peels rather than in the flesh, regardless of the fruit’s developmental stage or growth conditions (Fig. 3b), consistent with the previous studies[6,25]. The anthocyanin accumulation in the fruit peels increased with the development of fruits under both growth conditions (Fig. 3b). At the same developmental stage, Indigo Rose fruits with darker purple colour grown under the GC growth conditions produced more anthocyanins than those with lighter purple colour grown under the GH growth conditions (Fig. 3b). At the RR stage, the total anthocyanin content in the peels of Indigo Rose fruits grown in the GCs [Q = 2.1 ± 0.6; Q = (A535 − A650)/FW (100 mg)] was about 5-fold higher than that of Indigo Rose fruits grown in the GHs (Q = 0.4 ± 0.0) (Fig. 3b). The results clearly indicate that the GC growth conditions enhanced biosynthesis and accumulation of anthocyanins in the peels of Indigo Rose fruits.
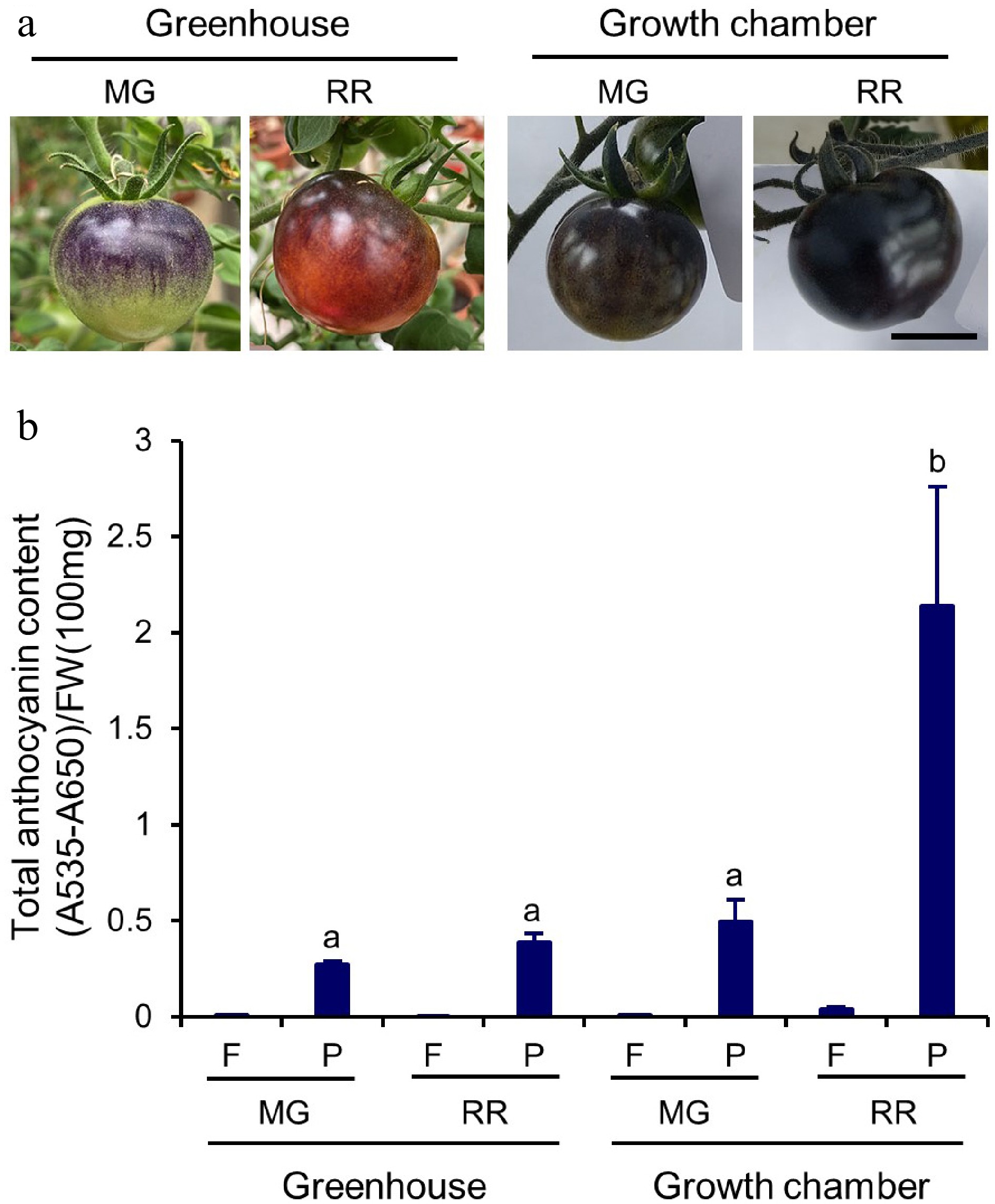
Figure 3.
Total anthocyanin content in Indigo Rose fruits grown in GH and GC. (a) Phenotype of Indigo Rose fruits grown in GH and GC. Scale bar = 2 cm. MG, mature green; RR, red ripe. (b) Total anthocyanin content in flesh (F) and peel (P) of Indigo Rose fruits in (a). FW, fresh weight. Data are means ± SD of three biological replicates. Statistical comparison was calculated by one way ANOVA followed by the Tukey’s test. Different letters indicate significant difference in anthocyanin content in peel at p < 0.05.
To further characterize the relationship between growth conditions and the anthocyanin biosynthesis in Indigo Rose fruits, a comparative study was conducted to detect the gene expression of the structural and TF genes involved in anthocyanin biosynthetic pathway in MG and RR fruits grown in the GHs and GCs, respectively. The PAL gene, the early biosynthetic genes (CHS, CHI, and F3H), and the late biosynthetic genes (DFR, ANS, and UFGT) were mainly expressed in peel tissues (Fig. 4). At both MG and RR stages, the expression levels of the PAL gene and the six structural genes were higher in the fruits grown in the GCs than that in the fruits grown in the GHs, which accounted for the higher anthocyanin content in the peel tissues of the fruits grown in the GCs (Figs 3 & 4). Under both growth conditions, the PAL and CHI genes had higher expression levels in MG fruits than in RR fruits, whereas the CHS, F3H, DFR, ANS and UFGT genes showed comparable expression levels in fruits at both MG and RR stages (Fig. 4). The latter results suggests that the biosynthesis of anthocyanins should still be active in Indigo Rose fruits at the RR stage, especially under the GC growth conditions. Like the structural genes, the TF genes ANT1, AN2-like, AN1, JAF13 and AN11 were mainly expressed in the peel tissues (Fig, 4). However, lower levels of transcripts of the ANT1, AN1, JAF13 and AN11 genes were also detected in the flesh tissues (Fig. 4). The expression levels of the TF genes ANT1, AN1, JAF13 and AN11 in the peel tissues were comparable under the two growth conditions at MG and RR stages (Fig. 4). The latter results indicate that the four TF genes were consistently expressed during fruit ripening stages. However, the AN2-like gene showed different expression patterns at the MG and RR stages under two different growth conditions. Under the GH growth conditions, the AN2-like gene was highly expressed in the peel tissues of the RR fruits but was low expressed in the peel tissues of the MG fruits (Fig. 4). However, under the GC growth conditions, the AN2-like gene was highly expressed in the peel tissues of the fruits at both MG and RR stages (Fig. 4). As the AN2-like gene encodes a strong and positive R2R3 MYB transcription factor for anthocyanin biosynthesis[7,18], the high expression of the AN2-like gene during fruit ripening stages, especially at the MG stage, should have contributed to high anthocyanin content in fruits grown under the GC growth conditions. To better control environmental factors, we conducted subsequent experiments in the GCs to investigate the influence of light and temperature on anthocyanin biosynthesis and accumulation in Indigo Rose.
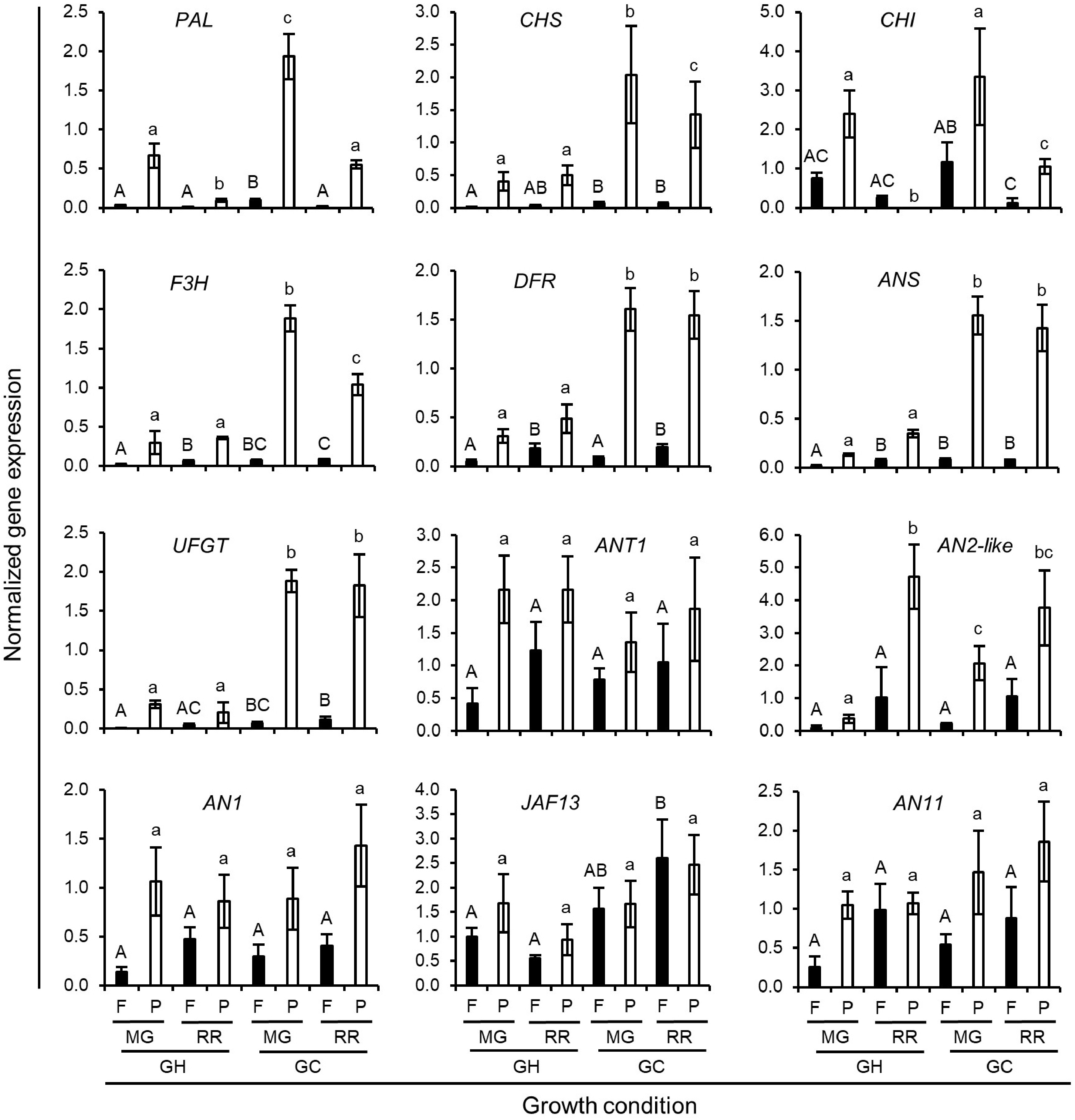
Figure 4.
Normalized expression level of anthocyanin biosynthetic genes in Indigo Rose fruits grown under GH and GC growth conditions. Data are means ± SD of three biological replicates. Statistical comparison was calculated by one way ANOVA followed by the Tukey’s test. Different letters indicate significant difference in gene expression in flesh (uppercase) or peel (lowercase) at p < 0.05. F, flesh; P, peel; MG, mature green; RR, red ripe.
The influence of light on anthocyanin biosynthesis in Indigo Rose fruits grown in growth chambers
-
The significant difference between GH and GC growth conditions in this study was that the GHs had short day growth conditions (12 h light : 12 h dark) with strong light intensity (200−900 µmol m−2 s−1). In contrast, the GCs were set with a long day photoperiod (16 h light : 8 h dark) with relatively low but consistent light intensity (160 µmol m−2 s−1). Under both growth conditions, Indigo Rose fruits developed purple to black colour on the area of peel facing the light (Figs 3a & 5a), indicating the importance of light on anthocyanin biosynthesis. To further investigate the influence of light intensity on anthocyanin accumulation in the fruit peel, Indigo Rose was allowed to grow in the GCs with light intensity at 80 µmol m−2 s−1 (Low), 160 µmol m−2 s−1 (Moderate) and 240 µmol m−2 s−1 (High), respectively. Based on the peel colour of the fruits, the high light intensity enhanced anthocyanin biosynthesis and accumulation in the fruits at both fruit developmental stages (Fig. 5a). The RR fruits grown under moderate (Q = 3.7 ± 0.1) and high light (Q = 3.5 ± 0.0) intensities contained similar levels of total anthocyanin content in the peel tissues (Fig. 5b). They were higher by 2-fold than in the peel tissues of the RR fruits grown under low light intensity (Q = 1.8 ± 0.1) (Fig. 5b). The expression levels of the structural and TF genes in anthocyanin biosynthetic pathways were examined by qRT-PCR analysis. In general, the expression levels of the structural genes were slightly upregulated with the increase in light intensity, which reached the highest expression levels under moderate or high light intensity (Fig. 6). The increase in light intensity also upregulated the expression of the ANT1, AN2-like, AN1 and AN11 genes (Fig. 6). They had higher expression levels under moderate and high light intensity than those under low light intensity (Fig. 6). The ANT1, AN2-like and AN1 genes showed their highest expression levels in RR fruits under moderate light intensity, whereas the AN11 genes reached its highest expression level in RR fruits under high light intensity (Fig. 6). Unlike other TF genes, the expression of the JAF13 gene, which encodes a positive bHLH TF for anthocyanin synthesis[26], seemed not to be significantly affected by the change of light intensity in this study (Fig. 6). Nevertheless, the qRT-PCR results collectively indicate that there was a positive correlation between light intensity and gene expression in Indigo Rose fruits and high light intensity enhanced the structural and TF gene expression. It should be noted that, under the long day photoperiod (16 h light:8 h dark), the moderate intensity of light at 160 µmol m−2 s−1 was sufficient for Indigo Rose to synthesize and accumulate anthocyanins in the peel tissues of fruits, achieving a performance efficiency similar to high light intensity (Fig. 5b). In addition, although lesser anthocyanins were accumulated in fruits, Indigo Rose could grow and set fruits normally under low intensity of light at 80 µmol m−2 s−1 (Fig. 5a). These results further suggest that Indigo Rose is a good candidate variety that can adapt well to artificial low light intensity under indoor growth conditions.
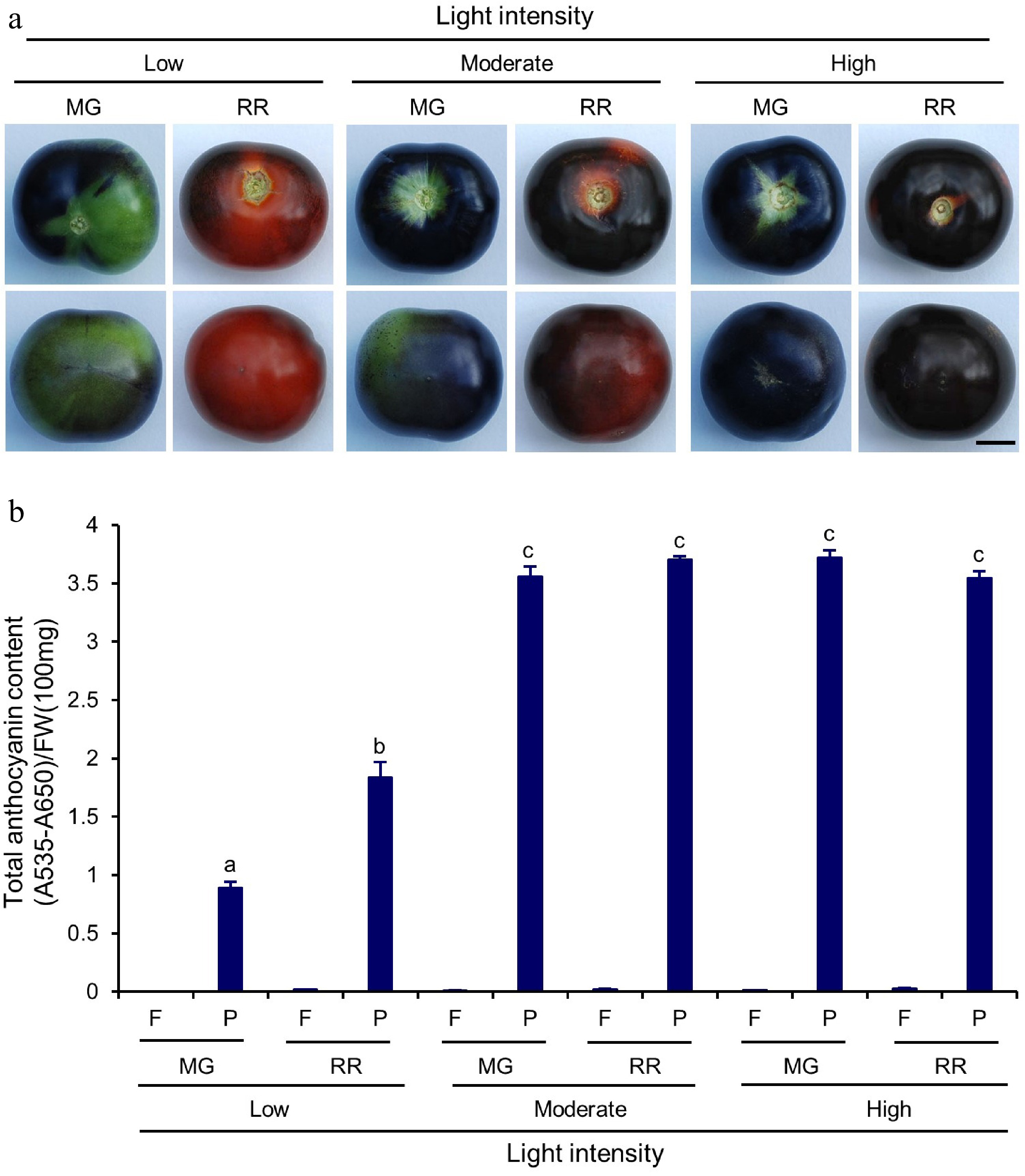
Figure 5.
Total anthocyanin content in Indigo Rose fruits grown in the GCs under different light intensity. (a) Top (upper row) and bottom (lower row) view of Indigo Rose fruits grown under low, moderate and high light intensity. Scale bar = 1 cm. MG, mature green; RR, red ripe. (b) Total anthocyanin content in flesh and peel of Indigo Rose fruits in (a). Statistical comparison was calculated by one way ANOVA followed by the Tukey's test. Different letters indicate significant difference in anthocyanin content in peel p < 0.05. FW, fresh weight; F, flesh; P, peel. Data are means ± SD of three biological replicates.
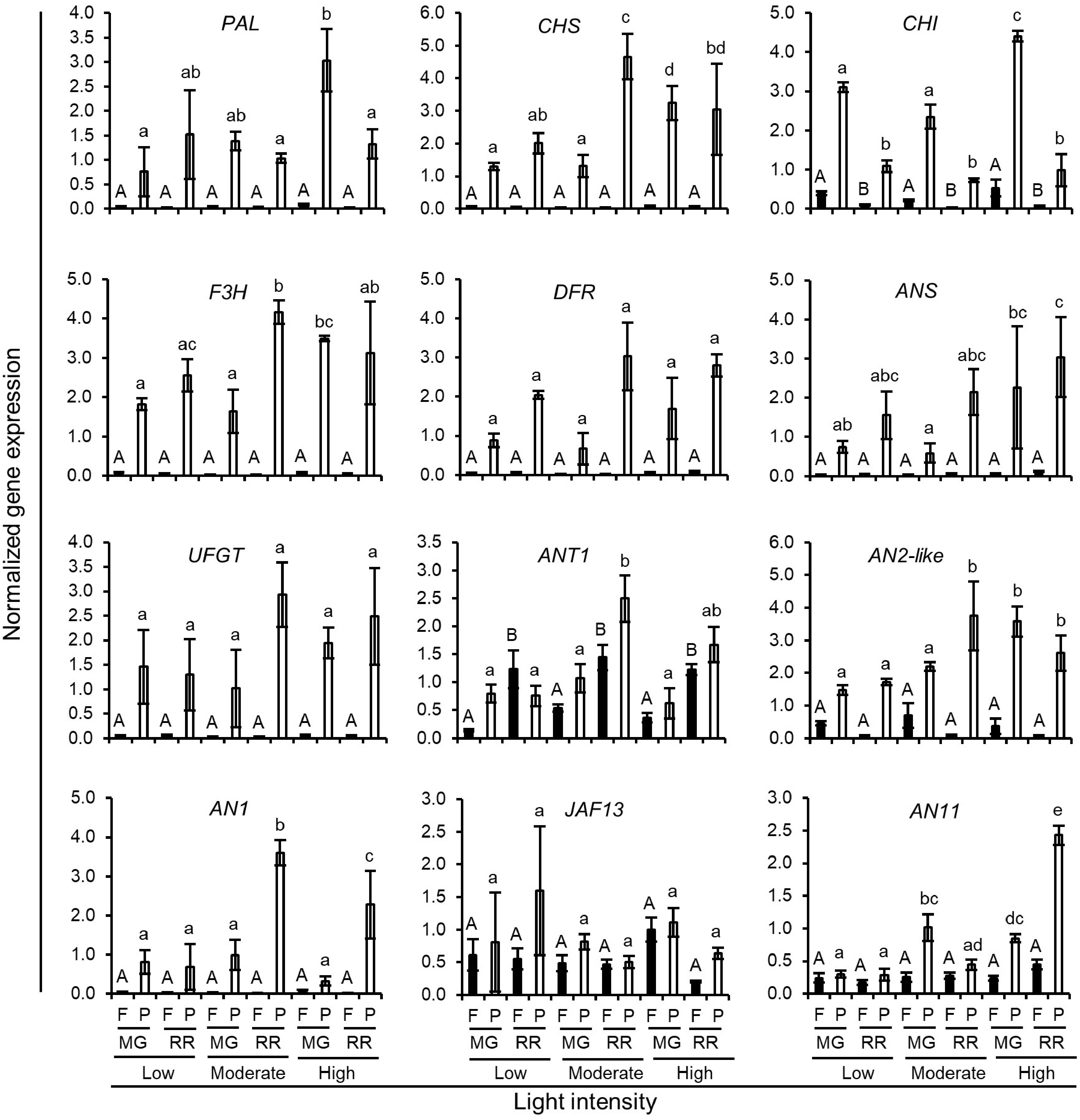
Figure 6.
Normalized expression level of anthocyanin biosynthetic genes in Indigo Rose fruits grown in the GCs under different light intensities. Data are means ± SD of three biological replicates. Statistical comparison was calculated by one way ANOVA followed by the Tukey's test. Different letters indicate significant difference in gene expression in flesh (uppercase) or peel (lowercase) at p < 0.05. F, flesh; P, peel; MG, mature green; RR, red ripe.
The influence of temperature on anthocyanin biosynthesis in Indigo Rose fruits grown in growth chambers
-
A notable difference between GH and GC growth conditions in this study was that the temperature in the GHs varied from 24 °C at night to 35 °C in the afternoon on sunny days, whereas an optimal temperature was consistently maintained in the GCs at 25 °C. Indigo Rose plants were grown in the GCs to investigate whether the temperature would affect the biosynthesis of anthocyanins and its accumulation in the fruit peel with temperatures set at 20, 25 and 30 °C. The colour of fruits grown at 20 and 25 °C was darker than that of fruits grown at 30 °C (Fig. 7a). The total anthocyanin content in the peel tissues of the RR fruits grown at 20 °C (Q = 3.6 ± 0.0) and 25 °C (Q = 3.7 ± 0.0) were also higher than that of the RR fruits grown at 30 °C (Q = 1.8 ± 0.1) (Fig. 7b). Notably, the total anthocyanin content in the peel tissues of the MG fruits grown at 30 °C (Q = 3.5 ± 0.2) was comparable to that of the MG fruits grown at 20 °C (Q = 3.7 ± 0.0) or 30 °C (Q = 3.3 ± 0.1). However, it decreased to a lower level in the RR fruits grown at 30 °C (Fig. 7b). The influence of temperature on the gene expression of the genes involved in the anthocyanin biosynthetic pathway was investigated. The PAL gene, which had a higher expression level in the peel tissue of the MG fruits than in the peel tissues of the RR fruits, was not significantly affected by temperature change (Fig. 8). The expression levels of the structural genes (CHS, CHI, F3H, DFR, ANS and UFGT) were generally decreased with the increase of temperature at both MG and RR stages (Fig. 8). The expression of the TF genes in the peel tissues was also enhanced by low temperature treatment. The ANT1 and AN11 genes, which were mainly expressed in the peel tissues of the RR fruits, showed higher expression levels in the RR fruits at 20 °C or 25 °C than that in the RR fruits at 30 °C (Fig. 8). The AN2-like and AN1 genes showed comparable expression levels in peel tissues of fruits at the MG and RR stages (Fig. 8). The two genes had the highest expression levels at 20 °C (Fig. 8). Finally, the JAF13 gene, which was expressed in both flesh and peel tissues, showed slightly higher expression in MG or RR fruits at 20 or 25 °C than that in the fruits at 30 °C (Fig. 8). In summary, the expression of both structural and TF genes was generally upregulated by the temperature treatments at 20 and 25 °C. The results suggests that anthocyanin biosynthesis in Indigo Rose fruits is stimulated by a relatively lower temperature at 20 to 25 °C. The consistent high temperature at 30 °C significantly affects the transcription of anthocyanin biosynthetic genes as well as the anthocyanin accumulation in the peel tissues of the fruits at the RR stage.
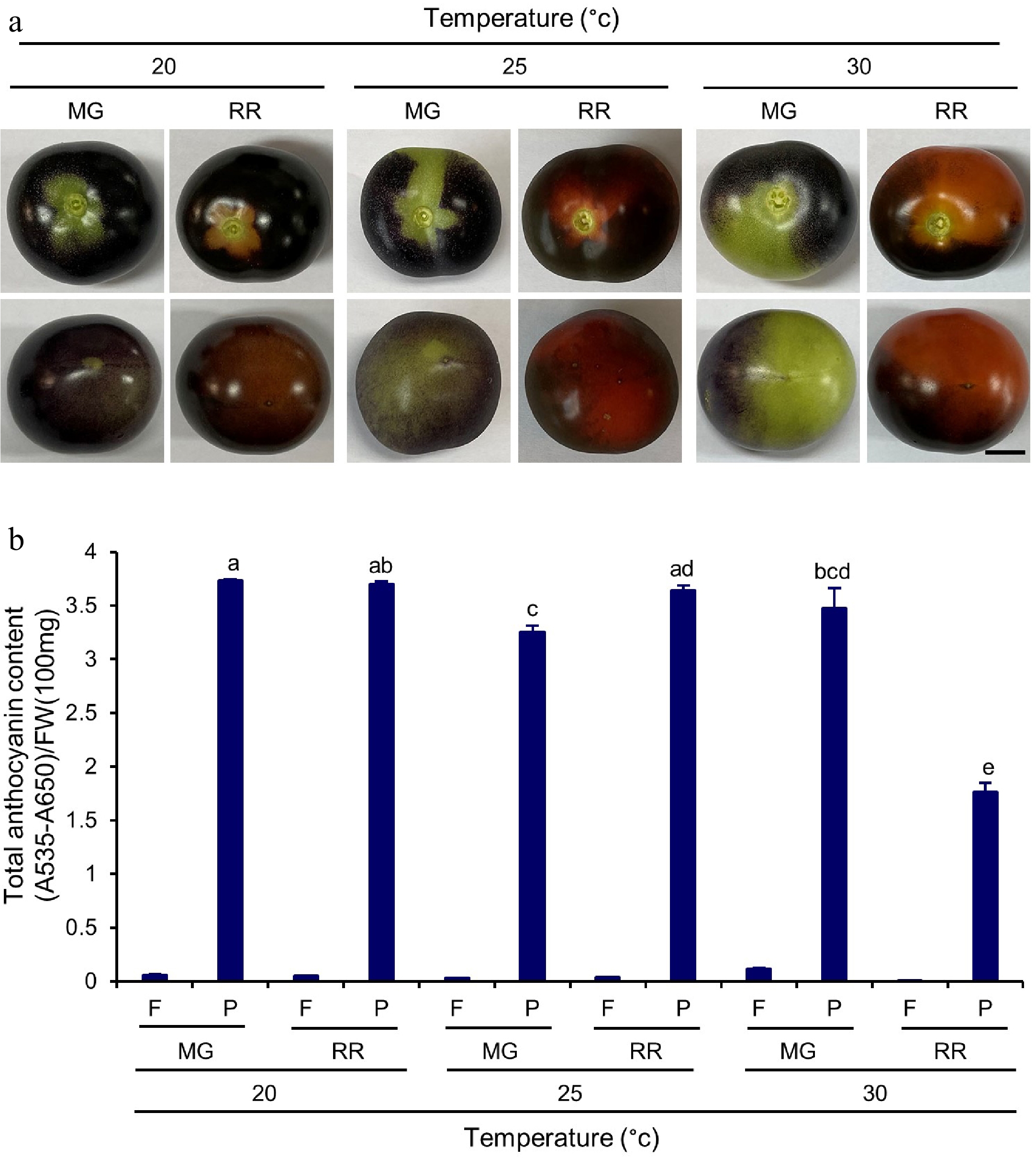
Figure 7.
Total anthocyanin content in Indigo Rose fruits grown in the GCs at different temperature. (a) Top (upper row) and bottom (lower row) view of Indigo Rose fruits grown in the GCs at different temperature. Scale bar = 1 cm. MG, mature green; RR, red ripe. (b) Total anthocyanin content in flesh and peel of Indigo Rose fruits in (a). Statistical comparison was calculated by one way ANOVA followed by the Tukey’s test. Different letters indicate significant difference in anthocyanin content in peel at p < 0.05. FW, fresh weight; F, flesh; P, peel.
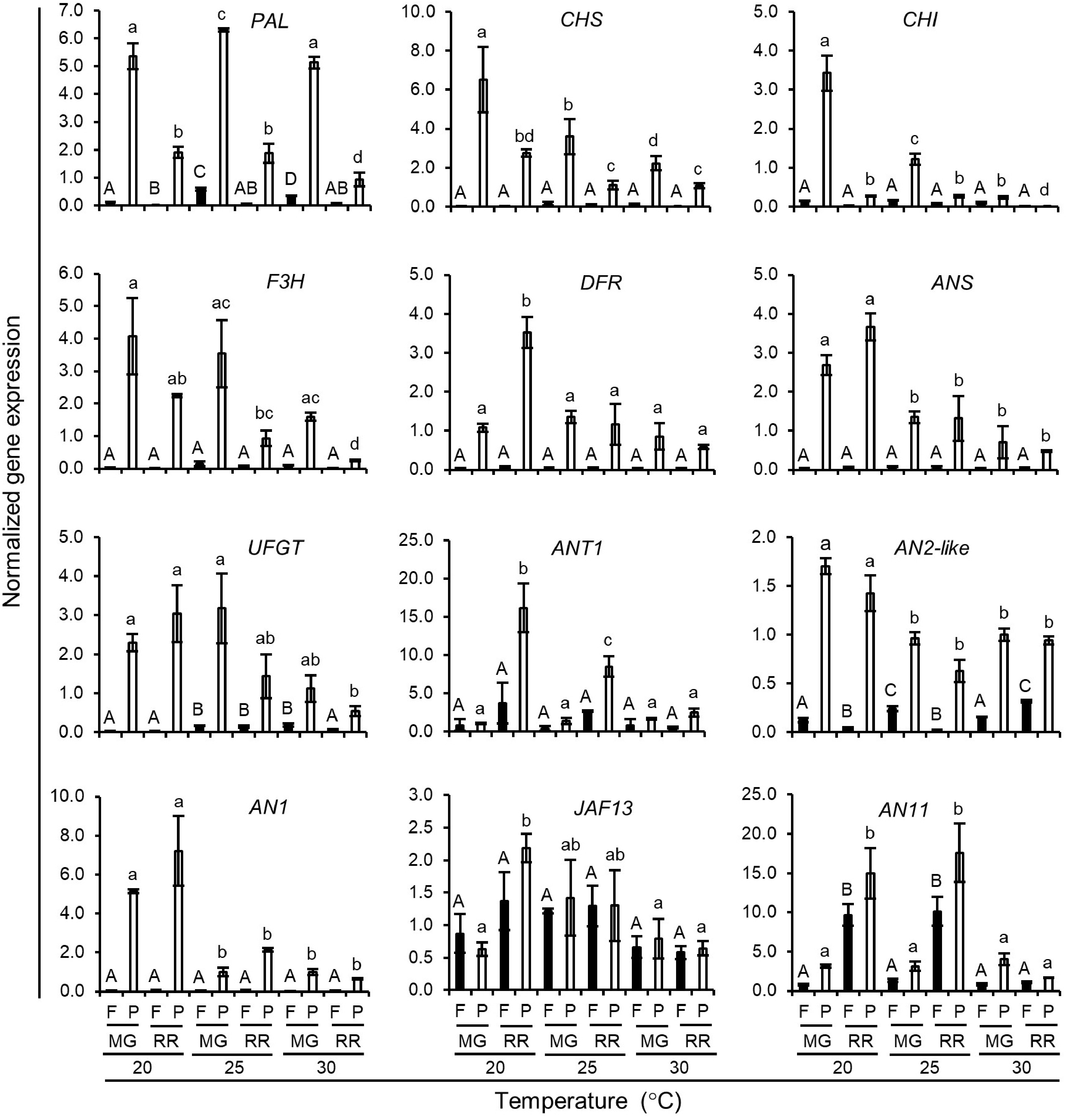
Figure 8.
Normalized expression level of anthocyanin biosynthetic genes in Indigo Rose fruits grown in the GCs at different temperature. Data are means ± SD of three biological replicates. Statistical comparison was calculated by one way ANOVA followed by the Tukey’s test. Different letters indicate significant difference in gene expression in flesh (uppercase) or peel (lowercase) at p < 0.05. F, flesh; P, peel; MG, mature green; RR, red ripe.
-
The initial objective of this research was to test the adaptation of the anthocyanin-rich tomato variety Indigo Rose to the tropical climate in Singapore. The preliminary results clearly indicate that Indigo Rose was not suitable for plantation in the GHs or open fields in Singapore, where the tropical climate with high temperatures causes low fertility and parthenocarpy in fruit setting. Furthermore, Indigo Rose fruits grown in the GHs accumulated less anthocyanins in fruits than that produced in the GCs. However, Indigo Rose displayed strong vegetative growth, normal fruit setting and high anthocyanin content in fruit peels when it was grown in the GCs. Indigo Rose showed the potential of being a good candidate variety of tomato for indoor farming. Therefore, the subsequent experiments of this study mainly concentrated on the optimalization of light and temperature in the GCs and their influence on the expression of genes involved in anthocyanin biosynthetic pathways in Indigo Rose.
Light is one of the most important abiotic phenomena that is the energy source for photosynthesis and regulates plant growth and development. Light even acts as a biological stimulus that triggers the defence mechanisms in which several complex metabolic networks interact each other, and several genes are regulated at the transcription level that alters the content of secondary metabolites[27]. Sunlight is known to enhance the accumulation of anthocyanins, reflecting their role as UV protectants[28]. Several studies have demonstrated that high light intensities could trigger the biosynthesis of anthocyanins[29−31]. Indeed, anthocyanins accumulate only on those parts of the Indigo Rose fruit peel that are exposed to sunlight (Fig. 3a)[9]. In this study, the average light intensity of the sunlight at the GHs was higher than that in the GCs, yet less anthocyanins were produced in the peel tissues of the fruits grown in the GHs (Fig. 3a & b). One of the reasons for low anthocyanin content in Indigo Rose fruits grown in the GHs might be caused by the short-day photoperiod under the GH growth conditions, which had a light cycle of 12 h light and12 h dark but with less than 8 h direct sunlight. A long-day photoperiod (16 h light/8 h dark) was set to plant Indigo Rose in the GCs. The light intensity at 160 µmol m−2 s−1 was found to be sufficient for anthocyanin biosynthesis and accumulation in Indigo Rose fruits under the GC growth conditions. This result suggests that Indigo Rose is an ideal candidate for indoor farming, requiring relatively low illumination. The high light utilization efficiency of Indigo Rose allows to keep a balance between fruit production with nutrient accumulation and energy consumption as lighting is one of the biggest expenses for an indoor farm. Future study may need to investigate the influence of light quality, photoperiod and their combination on anthocyanin biosynthesis and accumulation in Indigo Rose fruits.
Tomato is a thermophilic crop. The ideal temperature for tomato growth and fruit production ranges between 20 and 24 °C. Temperatures below 13 °C or above 32 °C adversely affect the fruit set and development[32,33]. Both low and high temperature induce reactive oxygen species (ROS) in plants, which causes oxidative damage to cellular functions and threatens plant growth and development. The hydrogen atom from the anthocyanins neutralizes the free radical in ROS. Hence, anthocyanins act as effective antioxidants by counterbalancing the negative factors and protecting plants against oxidative damage. Several studies found that low-temperature could induce anthocyanin biosynthesis and accumulation in maize (Zea mays L.)[34], red orange [Citrus sinensis (L.) Osbeck][35], grape (Vitis vinifera L. × Vitis labrusca L.)[36], apple (Malus domestica)[37], tomato[9] and Arabidopsis (Arabidopsis thaliana ecotype Col-0)[38]. In this study, the purpose of treating Indigo Rose with different temperatures was to screen for an optimal and economical temperature for indoor plantation, but not an abiotic stress study with extremely low or high temperature treatment. The anthocyanin content in the RR fruits grown at 20 and 25 °C was about two times higher than that in the RR fruits grown at 30 °C (Fig. 7). Meanwhile, the expression of most of the structural and TF genes in the RR fruits grown at 20 and 25 °C were also higher than that in the RR fruits grown at 30 °C (Fig. 7). It was observed that the anthocyanin content in the MG fruits grown at 30 °C was similar to that in the MG fruits grown at 20 and 25 °C, which was indeed higher than that in the RR fruits grown at 30 °C. The results demonstrate that the high temperature at 30 °C not only decreased anthocyanin biosynthesis by downregulating TF and structural gene expression but also reduced anthocyanin accumulation in ripened fruits. High temperature could be another significant reason that caused low anthocyanin content in Indigo Rose fruits grown in the GHs. Both the expression levels of the structural genes and the anthocyanin accumulation were significantly reduced in Indigo Rose fruits grown under the GH growth conditions (Figs 3b & 4). Based on the results derived from the experiments with temperature treatment, we recommend setting the temperature at 25 °C for Indigo Rose in indoor farming.
Under long-day photoperiod growth conditions in the well-controlled GCs, a general positive correlation between the expression of the structural and TF genes and the anthocyanin content in fruits was observed when they were grown under different light and temperature treatments (Figs 5b, 6, 7b & 8). Under the GH growth conditions with a short-day photoperiod, the expression levels of the structural genes in the fruits were lower than that in the fruits grown in the GCs (Fig. 4). It was important to note that the AN2-like gene, the master regulator for anthocyanin biosynthesis[7], was found to be highly expressed at both MG and RR stages under the GC growth conditions, whereas it was mainly expressed at the RR stage under the GH growth conditions (Fig. 4). The high expression of the AN2-like gene in Indigo Rose fruits at the MG stage positively regulates the expression of structural genes in the anthocyanin biosynthetic pathway through the MBW complex at the early ripening stages[11]. Considering the fact that the AN2-like gene positively regulates anthocyanin biosynthesis in a light-dependent manner[7], the expression of the AN2-like gene in peel tissues of fruits throughout the fruit ripening stages would enhance anthocyanin biosynthesis and accumulation in Indigo Rose fruits grown under the long-day photoperiod GC growth conditions.
-
Our findings in this report demonstrate that although the purple tomato ‘Indigo Rose' is not suitable for the semi-open GHs in tropical region due to high temperature, it is a good tomato cultivar for indoor plantation and urban farming, in which the moderate to high intensity of artificial light (160−240 µmol m−2 s−1) and relatively low temperature (20−25 °C) promote anthocyanin production and accumulation in fruits.
The authors thank Dongsheng Tian for technical support and useful discussion. This research was supported by the Agency for Science, Technology and Research (A*STAR) of the Ministry of Trade and Industry of Singapore under its Industry Alignment Fund Prepositioning (IAF-PP) with Grant No. A19E4a0101.
-
The authors declare that they have no conflict of interest.
-
# These authors contributed equally: Joanne Teo, Raji Mohan, Shilu Zhang
- Copyright: © 2022 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Teo J, Mohan R, Zhang S, Gui Y, Sng BJR, et al. 2022. Optimization of light and temperature in indoor farming to boost anthocyanin biosynthesis and accumulation in Indigo Rose tomato. Vegetable Research 2:18 doi: 10.48130/VR-2022-0018 

Optimization of light and temperature in indoor farming to boost anthocyanin biosynthesis and accumulation in Indigo Rose tomato
- Received: 19 September 2022
- Accepted: 17 November 2022
- Published online: 02 December 2022
Abstract: Anthocyanins are a group of flavonoids that are found ubiquitously in fruits and vegetables. They act as antioxidants in plants and are considered to have potential benefits to human health. The tomato cultivar 'Indigo Rose' produces purple-skinned fruits that are rich in anthocyanins in fruit peel. The current study focussed on the detection of anthocyanins and examined the expression profiling of anthocyanin biosynthetic and regulatory genes in Indigo Rose fruits grown under semi-closed greenhouse and fully controlled growth chamber conditions. The anthocyanin content in Indigo Rose fruits grown under growth chamber conditions with a long-day photoperiod and ambient temperature was higher than those grown in the greenhouse with a short-day photoperiod and high temperature. Under indoor growth conditions, moderate to high intensity of artificial light (160−240 µmol m−2 s−1) and relatively low temperature (20−25 °C) promoted anthocyanin production and accumulation in Indigo Rose fruits. The comparative studies on gene expression under different growth conditions also demonstrate that the expression levels of the structural and regulatory genes in the anthocyanin biosynthetic pathway have a positive correlation with the anthocyanin content in the fruit peel. The growth conditions identified in this study could be used to grow Indigo Rose in indoor farming in Singapore, where the tropical climate makes it challenging to set up a tomato plantation in an open field or greenhouse.
-
Key words:
- Tomato /
- Anthocyanin /
- Indigo Rose /
- Gene expression /
- Indoor farming