






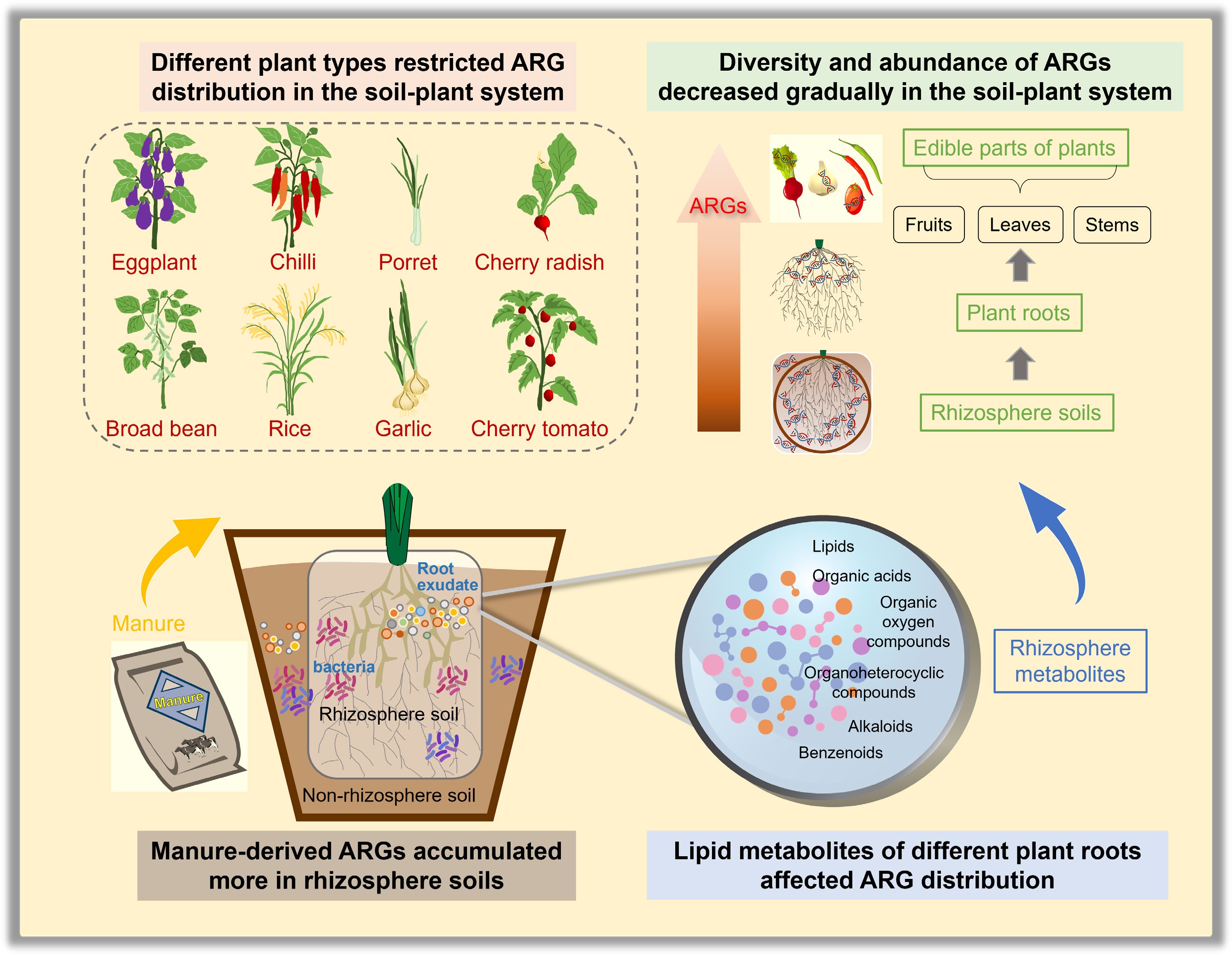



-
The widespread misuse of antibiotics has accelerated the dissemination of antibiotic resistance genes (ARGs) and antibiotic-resistant bacteria (ARB) into various environmental media, particularly soil[1−3]. The application of livestock manure to fields is a key pathway for ARGs and ARBs to enter soils[4−6]. It is particularly noteworthy that the cultivation of crops in soils amended with manure could accelerate the migration of ARGs along the food chain to humans[7,8]. Recent studies have demonstrated that the application of livestock manure can enhance the prevalence of antibiotic resistome within the rhizosphere and phyllosphere of crops[9,10]. Farmlands play a significant role in facilitating the transmission of ARGs to humans through food chains involving crops, vegetables, and fruits. Still, current technological measures exhibit limited capacity to control ARG contamination in soil[11−13]. Moreover, ARGs exhibit substantial horizontal gene transfer (HGT) capacity, potentially spreading within agricultural ecosystems through pathways such as soil pore water transport, soil fauna activities, and plant roots[14−16].
Several studies have shown that manure amendments can drive ARG evolution and dissemination in soil-plant systems[17,18]. Studies tracking the transmission of ARGs from swine manure to the soil-plant system revealed that Proteobacteria and Actinobacteria facilitated the dissemination of tetracycline ARGs (tet-ARGs) from manure. At the same time, the filtration of Firmicutes in the rhizosphere restricted the entry of tet-ARGs into plants[19]. Furthermore, plant species can significantly influence the prevalence of ARGs, which is attributable to variations in their physiological characteristics and microbiome profiles. Studies have shown that the abundance of ARGs in leafy vegetables (Shanghai greens) was significantly higher than in root vegetables (carrots), particularly within leaf and root endophytes, where differences of up to five to ten times were observed[9]. Under antibiotic selective pressure, ARGs can transfer from soil to root endophytes and subsequently to the phyllosphere, with cultivated species such as pakchoi and wild plants like barnyard grass demonstrating particularly strong capacities for ARG transfer[20]. The distinct distribution of ARGs in the soils and endophytes of different plant species may correlate with the rhizosphere metabolites emitted by these plants[16]. It is noteworthy that the variation in the abundance and diversity of ARGs across edible parts of different crop species indicates a potential preferential accumulation of antibiotic resistance in specific food crops, which could ultimately elevate the risk of human exposure through the food chain and pose a serious threat to public health[10,15,21,22].
The close bacterial interaction between soil and plant roots facilitates the HGT of ARGs into plants[15,23,24], while also enabling plant roots to regulate the variability of soil ARGs through their secretions[25,26]. Recent studies have shown that plant ARGs mainly originate from rhizosphere soil, with cooperative bacteria mediating migration—key Proteobacteria taxa like Clostridium-sensu stricito and Pseudomonas transferring ARGs to shoots[27]. Xiao et al.[28] demonstrated that 20.34% of ARGs were significantly shifted between rhizosphere and bulk soils, with rhizosphere-enriched ARGs associated with bacterial functions in signaling and degradation pathways, while rhizosphere-depleted ARGs were linked to biosynthetic pathways, nutrient cycling, and ABC transporters. Additionally, organic fertilization also influences the interaction between crop roots and microorganisms, thereby shaping the distinct characteristics of non-rhizosphere and rhizosphere environments. Rhizosphere metabolites, specific to different plant varieties, serve as chemical signals that indirectly promote the succession of microbial communities by regulating soil habitats[29,30], further affecting the migration and spread of ARGs in the soil-plant system[31−33]. However, how different plant root activities drive the distribution and spread of ARGs in soil-plant systems remains unclear. Certain metabolites in various crops could modulate ARG-host bacteria interactions, thereby influencing ARG transfer within soil-plant systems. However, the underlying mechanisms and key metabolites involved remain poorly understood and need further research.
Hence, it is hypothesized that crop-specific rhizosphere metabolites drive the distribution of manure-derived ARGs in soil-plant systems. To verify this, a pot experiment involving diverse crops was performed to investigate the differences in ARGs, metabolites, and microbial communities across different soil compartments (non-rhizosphere and rhizosphere) and plant tissues (roots and edible parts) via high-throughput quantitative PCR (HT-qPCR) combined with untargeted metabolomics. This study aimed to: (1) systematically compare variations in ARG profiles between non-rhizosphere and rhizosphere soils of different crops, as well as their distribution in plant roots and edible parts; (2) elucidate the drivers of ARG enrichment in rhizosphere soils, by clarifying the interactions among rhizosphere metabolites, mobile genetic elements (MGEs), and host bacterial communities; (3) reveal the driving mechanisms underlying the spread of ARGs in soil-plant systems mediated by rhizosphere metabolites, particularly between plants categorized as high-ARG-abundance and low-ARG-abundance groups. This study is the first to reveal, from the perspective of rhizosphere metabolites, the mechanisms underlying the variation in ARG profiles across rhizosphere soil, plant roots, and edible parts across different soil-plant systems, which will help in further evaluating the impact of plant rhizosphere metabolites on ARG risks in soil and food.
-
The pot experiment was performed in a Yunnan Province greenhouse (Dali; 25°53′ N, 100°10′ E) between June and October 2022. The temperature was maintained at 26 ± 5 °C under ambient light and humidity conditions. Two treatments were set up: no fertilization (CK); livestock manure application (M). All treatments were performed simultaneously in the greenhouse under the same environmental conditions, using eight plant species with different edible parts: broad bean (B) (Vicia faba L.), garlic (G) (Allium sativum L.), rice (R) (Oryza sativa L.), cherry tomato (CT) (Lycopersicon esculentum var. cerasiforme A.Gray), chilli (C) (Capsicum annuum L.), eggplant (E) (Solanum melongena L.), cherry radish (CR) (Raphanus sativus L.) and porret (P) (Allium schoenoprasum L.). Two-week-old seedlings of all crops were transplanted into pots. All crops were cultivated with three plants per pot and harvested simultaneously on day 120 post-transplantation, ensuring consistent soil growth duration while encompassing critical developmental stages. At harvest, a range of samples were collected, comprising roots (G), the edible parts of the plants, non-rhizosphere soil (NR), rhizosphere soil (R), and soil solution. Three replicate pots were collected for each, totaling 48 pots. The root and edible parts of the eight plant species with different edible parts were collected, including the roots and stems of garlic, the roots and grains of rice, the roots and fruits of cherry tomato, the roots and fruits of chilli, the roots and fruits of eggplant, the roots and leaves of broad bean, the main roots and taproots of cherry radish, and the roots and leaves of porret. Each pot (19 cm in height × 18 cm in diameter) was filled with 2 kg of soil. Manure was applied at 100 g per pot for the M treatment, while no manure was added to the CK treatment. The initial properties of the soil used in the pot experiment were consistent across all pots, with a pH of 7.19 ± 0.16; organic matter content of 56.81 ± 2.35 g/kg; total nitrogen content of 2.38 ± 0.07 g/kg; and total phosphorus content of 0.89 ± 0.03 g/kg. Each pot was equipped with 200-mesh nylon rhizosphere bags (diameter 8 cm, height 18 cm) to ensure that the rhizosphere bags covered the roots of the plants. Rhizosphere soil was collected from inside the bags, and non-rhizosphere soil from outside. Soil solution was collected using soil solution samplers with hydrophilic microporous membranes (0.12–0.18 μm), preserving soil structure while preventing contamination. Plant tissues were surface-sterilized with 2% (w/v) available chlorine NaClO solution, followed by 70% ethanol, rinsed five times with sterile distilled water, and aseptically dried. Sterility was confirmed by culturing 0.1 mL of the final rinse water in LB medium at 30 °C for 5 d, during which no bacterial growth was observed. Soil solution (80 mL) and soil samples (1 kg) were stored at –80 °C for metabolite analysis and DNA extraction.
DNA extraction and high-throughput quantitative PCR
-
Each soil sample was weighed at 0.5 g (dry weight) for DNA extraction using the Fast DNA® Spin Kit for soil (MP Biomedicals, LLC, Santa Ana, USA). Each plant sample was weighed at 0.2 g for DNA extraction using the Fast DNA® Spin Kit (MP Biomedicals, LLC, Santa Ana, USA). DNA concentration and purity were measured using a NanoVue Plus spectrophotometer (GE Healthcare, USA). DNA concentrations exceeded 30 ng/μL, with A260/A280 ratios between 1.8–2.0 and A260/A230 > 2.0, ensuring extraction quality suitable for downstream analyses. DNA obtained from plants, rhizosphere, and non-rhizosphere soil samples was then stored at –80 °C for analysis. HT-qPCR was conducted on the Wafergen SmartChip Real-time PCR system (MicroAnaly Genetech Co., Ltd, China) to detect 88 ARG subtypes, seven MGE subtypes, and the 16S rRNA gene, which served as the reference gene for calculating relative abundance. The primers used are presented in Supplementary Table S1. The 100 nL reaction system included 2 ng/μL DNA template, 1 × LightCycler 480 SYBR Green I Master, and 500 nM of each primer. The PCR amplification protocol involved an initial denaturation step at 95 °C for 10 min, followed by 40 cycles of denaturation at 95 °C for 30 s and annealing at 60 °C for 30 s.
16S rRNA gene amplicon sequencing
-
The forward primer 338F (5'-ACTCCTACGGGAGGCAGCAG-3') and the reverse primer 806R (5'-GGACTACNNGGGTATCTAAT-3') were used to amplify the 16S rRNA gene. PCR was performed using an ABI 9700 PCR instrument (Applied Biosystems, USA) with a total reaction volume of 25 μL, including 2 μL template DNA, 3 μL BSA (2 ng/μL), 5.5 μL ddH2O, 12.5 μL 2× Taq PCR MasterMix (Vazyme Biotech Co., Ltd, China), 1 μL of forward primer (5 μM), and 1 μL of reverse primer (5 μM). The amplification process involved an initial denaturation at 95 °C for 5 min, followed by 28 cycles of denaturation at 95 °C for 45 s, annealing at 55 °C for 50 s, and extension at 72 °C for 45 s, concluding with a final extension at 72 °C for 10 min. The PCR products were purified using the Agencourt AMPure XP Kit (Beckman Coulter, Inc., USA), and sequencing libraries were constructed using the NEB Next Ultra II DNA Library Prep Kit (New England Biolabs, Inc., USA). PEAR (v0.9.6) software[34] was used to filter and concatenate the raw data, and then VSEARCH (v2.7.1) software[35] was used to filter out sequences shorter than 230 bp. Qualified sequences were clustered into operational taxonomic units (OTUs) using the UPARSE algorithm[36] in VSEARCH (v2.7.1) software, with a similarity threshold of 97%.
Soil metabolites detection and differential metabolite analysis
-
The rhizosphere metabolites were analyzed using a UHPLC (Vanquish, Thermo Fisher Scientific) coupled to an Orbitrap Exploris 120 mass spectrometer (Orbitrap, Thermo Fisher Scientific) at Beijing Allwegene Co., Ltd (Beijing, China). From each sample, 100 μL of soil solution was mixed with 400 μL of extraction solution (methanol and acetonitrile in a 1:1 volume ratio). The mixture was vortexed thoroughly and then subjected to ultrasonication in an ice-water bath. After settling, the mixture was centrifuged, and the supernatant was transferred to an autosampler vial for instrumental analysis. Additionally, equal volumes of supernatant from all samples were pooled to prepare a quality control (QC) sample, which was also analyzed using the same procedure. The QC sample method was used to normalize the non-targeted metabolite data. The relative standard deviation (RSD) of feature peaks was calculated across QC samples, with acceptance criteria set at ≤ 30% RSD for ≥ 70% of compounds in each sample. The three-dimensional data (sample name, peak number, and normalized peak area) were input into the 'MetaboAnalystR' package in R software for principal component analysis (PCA) and orthogonal partial least squares discriminant analysis (OPLS-DA). Differentially expressed metabolites (DEMs) were identified based on the following criteria: (1) a variable importance in the projection (VIP) score > 1 from the OPLS-DA model; (2) a p-value < 0.05 using a two-tailed Student's t-test; and (3) a fold change ≤ 0.67 or ≥ 1.5[37]. The metabolites were annotated using the KEGG (
www.kegg.jp ) and MetaboAnalyst (www.metaboanalyst.ca ), followed by an analysis of the enrichment degree in relevant metabolic pathways[38].Statistical analysis
-
Heatmaps, histograms, and line charts were created with Origin 2021 software to visualize the diversity and abundance of microbial communities, MGEs, and ARGs. Network analysis was visualized using Gephi 0.9.2, utilizing significant Spearman correlations (r > 0.8, p < 0.05) identified among ARGs, metabolites, microbial taxa, and MGEs. Mantel tests and structural equation models (SEMs) were implemented in R 4.4.0 to assess the persistence and spread of ARGs as shaped by bacterial communities, MGEs, and metabolites, employing the 'linkET' and 'piecewiseSEM' packages, respectively[39]. Model fit was evaluated with Fisher's C test, and the model was iteratively refined according to pathway significance (p < 0.05) and model fit criteria (0 ≤ Fisher's C/df ≤ 2 and 0.05 < p ≤ 1.00).
-
Across the rhizosphere and non-rhizosphere soils of various crops, 81 subtypes of ARGs conferring resistance to eight antibiotic classes were identified. Among them, multidrug ARGs (MDR-ARGs) accounted for the largest proportion (total abundance of 7.44 × 10−1 copies/16S rRNA copies), followed by β-lactamase ARGs (bla-ARGs), sulfonamide ARGs (sul-ARGs), tet-ARGs, and MLSB ARGs, whereas aac(6')-Iy, aph6ia, mecA, tetD-01, tetH, tetK and vanB-02 were not detected in all samples (Fig. 1a). For most abundant ARG subtypes were mexF, oprD, yceL/mdtH-01 (MDR-ARGs), and blaSHV-01, penA (bla-ARGs) in different crop soils. The diversity and abundance of ARGs in rhizosphere and non-rhizosphere manured soils were significantly higher than those in unmanured soils (p < 0.05) (Fig. 1), implying that manure is likely to release a large number of ARGs into the soil environment. For instance, the total abundance of ARGs in cherry radish and rice increased by 1.50 times and 1.28 times, respectively, while in chilli, the increase was the least, at only 0.10-fold compared with that in unmanured soils (p > 0.05).
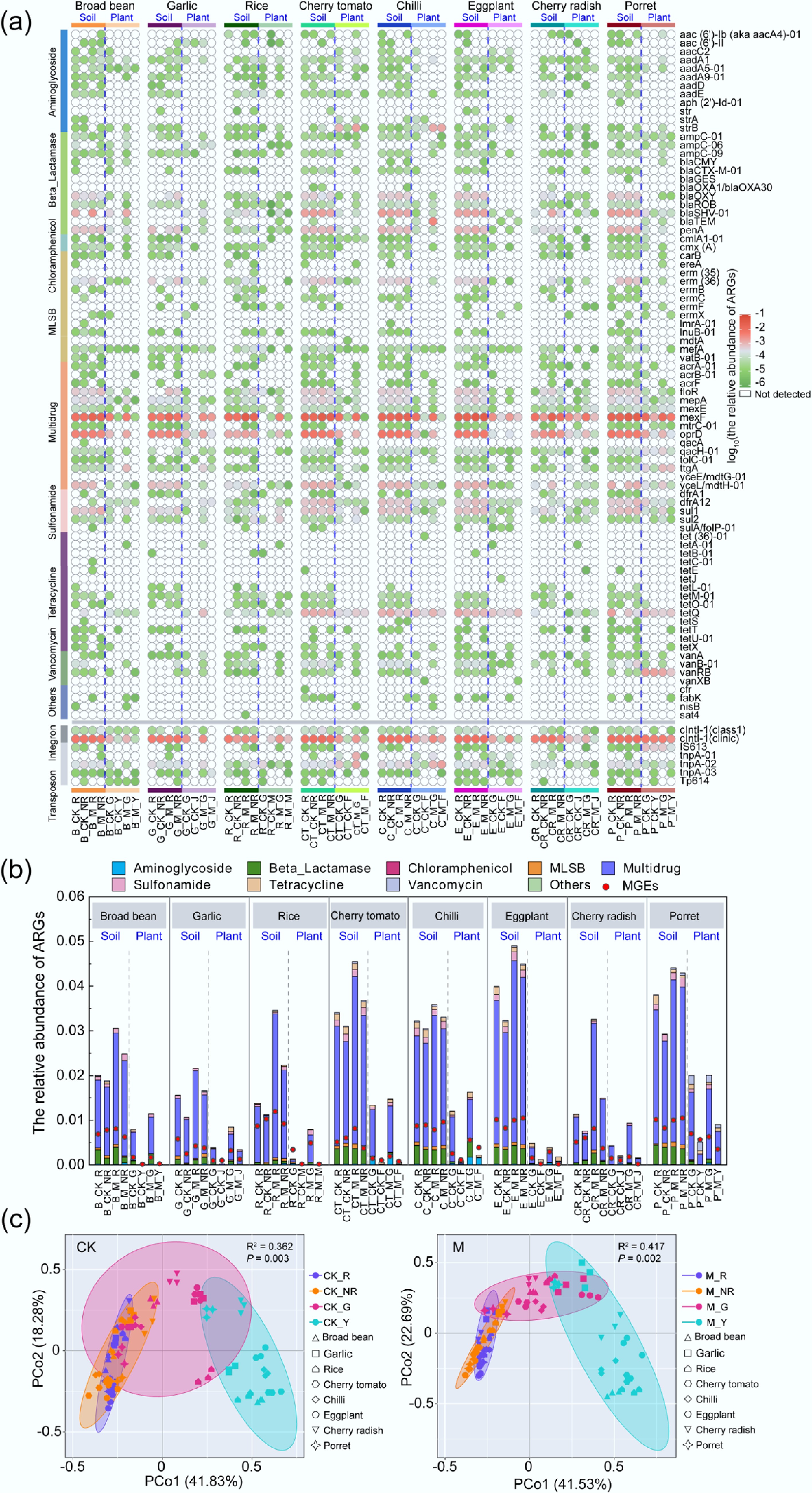
Figure 1.
Distribution of ARGs in the soil-plant system of different plants. (a) Detection of ARGs in all samples. (b) Relative abundance of ARGs in all samples. (c) Distribution of ARGs in all samples. (B, broad bean; G, garlic; R, rice; CT, cherry tomato; C, chilli; E, eggplant; CR, cherry radish; P, porret; CK, treatments without fertilizer; M, application of manure treatments; R, rhizosphere soils; NR, non-rhizosphere soils; G, plant roots; Y or J or F or M, edible parts of plants).
Rhizosphere soils exhibited a higher total abundance of ARGs compared to non-rhizosphere soils, showing a 1.24-fold increase for most detected subtypes (Fig. 1b). In different crops from unmanured treatments, ARG abundance in cherry radish and garlic rhizosphere soils was significantly higher than that in non-rhizosphere soils, increasing by 49.43% and 46.39%, respectively. Furthermore, all ARGs except vancomycin ARGs (van-ARGs) were higher in unmanured rhizosphere soils of eggplant and garlic, while in cherry tomato, the abundance of other ARGs except MDR-ARGs and van-ARGs was lower in rhizosphere soils than in non-rhizosphere soils. Otherwise, among different crops in manured soil, the abundance of ARGs in cherry radish and rice rhizosphere soils was 2.18 and 1.55 times that of non-rhizosphere soils, respectively, and the accumulation of ARGs in rhizosphere soils was higher. Specifically, the abundance of MDR-ARGs was consistently elevated in the rhizosphere compared to non-rhizosphere soils across all crops, with bla-ARGs demonstrating a similar pattern of enrichment in both garlic and chilli. However, except for eggplant and cherry radish, aminoglycoside ARGs (aac-ARGs) showed a tendency to be higher in non-rhizosphere soils than in rhizosphere soils. It is worth noting that all types of ARGs in cherry radish accumulated in the rhizosphere soils, and ARGs accumulated significantly in the rhizosphere soils under manure-amended conditions.
Diversity and abundance of ARGs gradually decreased from root to edible part
-
For the eight crops with different edible parts, this study measured the diversity and abundance of ARGs in various plant endophytes (the roots and edible parts), with a total of 69 ARG subtypes detected. The subtypes of ARGs detected in plant endophytes were 12 fewer than those in soil, including two aac-ARGs (aph(2')-Id-01 and str), two bla-ARGs (blaGES and blaOXA1/blaOXA30), three MLSB ARGs (erm(35), ermC and lmrA-01), three tet-ARGs (tetC-01, tetL-01 and tetS), and two other ARGs (cfr and sat4). Principal coordinates analysis (PCoA) revealed distinct segregation patterns of ARGs between plant endophytes and soils, and significant differences were also observed in ARG distribution between the roots and the edible parts (leaves, stems, and fruits) (Fig. 1c). Similar to soils, MDR-ARGs also had the highest abundance in plant endophytes (total abundance of 1.35 × 10−1 copies/16S rRNA copies), followed by bla-ARGs and tet-ARGs. Still, their total abundance levels were significantly decreased compared with rhizosphere soil (p < 0.05), by 81.83%, 83.50% and 38.17%, respectively.
Figure 2 shows the detection of ARGs in non-rhizosphere and rhizosphere soils of different crops, as well as endophytes in plant roots and edible tissues. Across four sample types from the eight crop species, shared ARGs were detected. Porret exhibited the highest number of shared ARGs (n = 17) in both soil compartments and plant tissues, followed by rice (n = 10) and cherry tomato (n = 8). Chilli and cherry radish each contained six shared ARGs, while broad bean showed the lowest count (n = 4). Moreover, the abundance heatmap of shared ARGs in Fig. 2 reveals a decreasing trend of most ARGs from soil to plant, suggesting that the shared ARGs from manure gradually migrated and spread to the edible parts of plants. These results suggested that different plant species may also determine the difference in ARG abundance between endophytes, rhizosphere, and non-rhizosphere soils. Figure 2 identifies mexF as the most abundant ARG subtype and a core shared ARG across soil and plant endophytes in all eight crops, which had a higher risk of transferring resistance as a multidrug ARG. mexF abundance was significantly higher in soils than in plant endophytes across all crops, with rhizosphere soils exhibiting greater accumulation than non-rhizosphere soils (p < 0.05). The relative abundance of mexF in edible plant parts decreased by more than 80% compared to rhizosphere soils. Reductions reached 99.98% in cherry tomato fruits, while porret leaves showed a smaller decline (80.84%).

Figure 2.
Detected number and abundance of shared ARGs in rhizosphere, non-rhizosphere soil, plant roots, and edible part samples of different crops. The columns represent the number of detected ARGs, and the data in the heatmap represent the abundance of ARGs. (a) Broad bean, (b) garlic, (c) rice, (d) cherry tomato, (e) chilli, (f) eggplant, (g) cherry radish, (h) porret. Refer to Fig. 1 for abbreviations.
ARG enrichment was lower in fruit plants and higher in leaf plants
-
There were differences in the spread of ARGs to various parts of the plant endophytes. Therefore, crops and vegetables with different edible parts were selected in this study, which showed a diverse distribution of ARGs in endophytic bacteria with edible parts. As shown in Fig. 3a, the diversity and abundance of ARGs in endophytes of plant roots were significantly higher than those in edible parts across different crops planted in manured soils and unmanured soils (p < 0.05). In the unmanured soils, the abundance of ARGs in edible parts of cherry radish showed the smallest decrease (53.1%), compared with that in root endophytes, while broad bean leaves showed the greatest decrease (96.9%) relative to their roots. Under manure application, ARGs in broad bean leaves decreased by 98.5% compared with those in the root endophytes, while those in porret leaves decreased by 55.2% relative to the roots.

Figure 3.
(a) ARG spread through the roots and edible parts of the plant, and the migration rate from the roots to the edible parts. (b) Distribution of ARGs in non-rhizosphere soil, plant roots, and aboveground part samples. (c) Distribution of MGEs in non-rhizosphere soil, plant roots, and aboveground part samples. (d) Regression analysis of cIntI-1(clinic) and ARGs (n = 32), with a 95% confidence interval. Refer to Fig. 1 for abbreviations.
Among the different edible parts of different crops, the distribution of ARGs varied significantly under the same manure application rate. Specifically, the total abundance of ARGs in the leaves of porrets on the edible parts was the highest among the eight plants, followed by that in the stems of garlic, with abundance levels of 9.00 × 10−3 and 3.39 × 10−3 copies/16S rRNA copies, respectively. That is, the risk of ARGs was higher in the leaves of porret, and the detection rate of MDR-ARGs reached up to 60%. Nevertheless, for plants whose edible parts are fruits and endosperm, the abundance of ARGs in their edible parts was low, and the total abundance of ARGs in cherry tomato fruits and in rice endosperm was only 6.82 × 10−4 and 2.56 × 10−4 copies/16S rRNA copies, respectively, which was an order of magnitude lower than that in porret leaves. Among the ARG subtypes detected in all plants, mexF was identified in every species studied. The highest abundance was observed in the leaves of porret (5.47 × 10−3 copies/16S rRNA copies), followed by the fleshy roots of garlic (10−3 copies/16S rRNA copies) and the stems of cherry radish (10−4 copies/16S rRNA copies). In contrast, rice and cherry tomato exhibited significantly lower abundances. The abundance of mexF in porret leaves was three orders of magnitude higher than that in cherry tomato, which had the lowest abundance. Therefore, the spread of ARGs from root endophytes to other plant tissues varied, and different parts of crops and vegetables consumed as food may pose different resistance risks.
MGEs significantly correlated with the spread of ARGs in the soil-plant system
-
A total of seven MGE subtypes belonging to integrase and transposase were detected in soil and plant endophytes of the eight different crops, showing similar trends to those of some ARGs. The cIntI-1(clinic) in MGEs had the highest abundance in soil (2.18 × 10−1 copies/16S rRNA copies), followed by tnpA-02 and cIntI-1(class1), while in plant endophytes, cIntI-1 (clinic) and tnpA-02 showed higher abundance, reaching 5.01 × 10−2 and 8.00 × 10−3 copies/16S rRNA copies, respectively. The diversity and abundance of MGEs under the manure treatment were significantly higher than those in the unfertilized treatment (p < 0.05). Compared with non-rhizosphere soil, MGE total abundance was significantly higher in rhizosphere soil (p < 0.05), indicating enrichment of MGEs and ARGs in rhizosphere soils. The abundance of integrons in endophytic bacteria was significantly lower than that of MGEs in soil (p < 0.05), consistent with changes in ARGs. Specifically, the total relative abundance of cIntI-1(class1) and cIntI-1(clinic) in endophyte bacteria decreased by 74.66% and 77.04%, respectively, compared with soil.
Meanwhile, MGE abundance in root endophytes was significantly lower than that in other plant parts (p < 0.05), indicating that MGEs in manure entered the plant through the roots and spread upward, though the abundance level gradually decreased. According to Fig. 3b, c, the distribution of MGEs and ARGs in rhizosphere soil, root endophytes, and aboveground endophytes showed a consistent pattern, and MGEs and ARGs were closely related to their transmission in soil-plant system. Particularly, the high proportion of cIntI-1(clinic) was very similar to the profile of high levels of MDR-ARGs and bla-ARGs. Therefore, the relative abundances of the seven MGEs and ARGs were further analyzed by linear regression (Supplementary Fig. S1). Notably, cIntI-1(clinic) in integron was closely related to ARGs with a significant linear correlation (r = 0.92, p < 0.001, Fig. 3d), suggesting a vital role of cIntI-1(clinic) in ARG spread from soil to plant endophytes.
Differences in soil and plant-associated bacterial communities
-
There were significant differences in soil and plant bacterial communities among different crops (Fig. 4 and Supplementary Fig. S2). As shown in Fig. 4a, the observed species and α-diversity in the soil bacterial community were significantly higher than those in the plant samples, and the diversity of the endophytic bacterial community in the root of plants was also significantly higher than that in the edible aboveground tissues (p < 0.05). The bacterial community composition of rhizosphere and non-rhizosphere soil of different crops and vegetables at the phylum and genus levels under livestock manure application and control treatment was shown in Fig. 4b, c. At the phylum level, Actinobacteriota (36.00%–46.05%), Chloroflexi (17.36%–23.52%), Proteobacteria (14.78%–19.41%), Acidobacteriota (7.95%–12.59%), and Gemmatimonadota (2.00%–3.32%) consistently emerged as the five dominant bacterial phyla in both manure-amended and unfertilized treatments, in both rhizosphere and non-rhizosphere environments. However, the characteristics of plant endophytic bacteria community are different, among which Proteobacteria occupied the highest abundance ratio, accounting for 14.78%–99.57%. Then, followed by Actinobacteriota (0.09%–54.36%), Chloroflexi, Acidobacteriota, and Gemmatimonadota (0.00%–3.33%). At the genus level, Nocardioides was the most abundant in soil, while Streptomyces was the most abundant in plants. When comparing rhizosphere and non-rhizosphere soil, the total abundance of Actinobacteriota in rhizosphere soil increased by approximately 2.17% relative to non-rhizosphere soil, while Acidobacteriota was 4.17% lower.
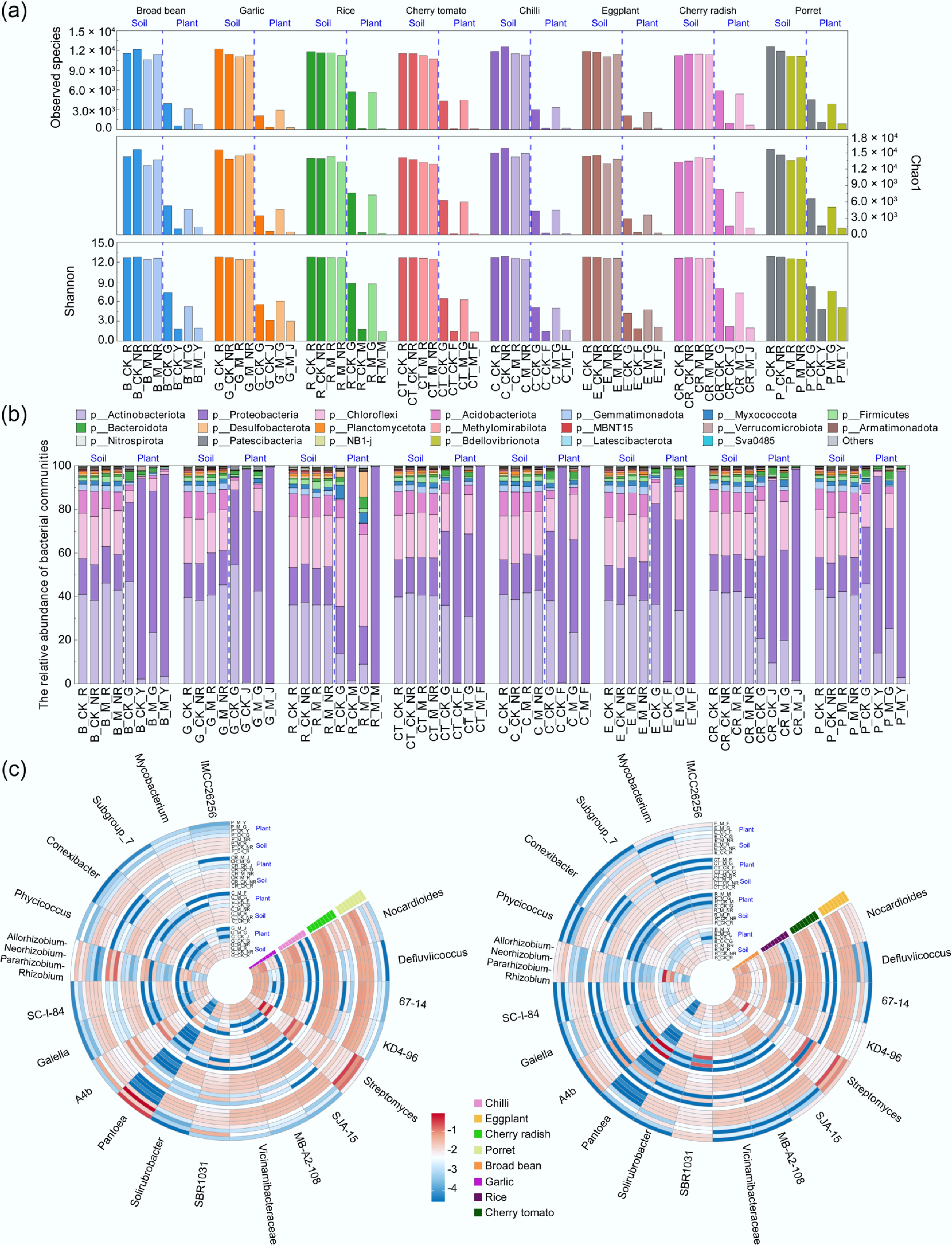
Figure 4.
Relative abundance of the dominant bacterial taxa at the phylum and genus levels (top 20). (a) Alpha diversity of bacterial communities in samples. (b) Proportion of bacterial communities at the phylum level. (c) Heatmap of bacterial communities at the genus level. Refer to Fig. 1 for abbreviations.
Additionally, there were significant differences in the abundances of endophyte communities between different plant parts at both phylum and genus levels (p < 0.05). At the phylum level, the total abundance of Actinobacteriota and Chloroflexi in the edible parts of the plant was significantly lower than that in the root endophytes, while the total abundance of Proteobacteria was significantly higher than that of the root endophytes (p < 0.05). At the genus level, the total abundance of Streptomyces in the edible part of the plant was significantly lower than that of the root endophytes, while the total abundance of Pantoea was significantly higher than that of the root endophytes (p < 0.05). Specifically, the endophytic bacteria in the edible parts of rice germ and the Proteobacteria in tomato fruit increased most compared with that root, reaching 4.03-fold and 1.76-fold, respectively. Among all crops, Actinobacteriota in garlic stems showed the greatest abundance decrease relative to roots, while Chloroflexi in rice grains showed the greatest decrease compared with the roots.
Multiple factors driving ARGs transmission in the soil-plant systems
-
Rhizosphere metabolites affected the ARG distribution and spread in soil-plant systems of different crops. Three crops with high ARG abundances (garlic, cherry radish, and porret) and three crops with low ARG abundances (broad bean, rice, and cherry tomato) were selected and divided into two comparison groups. The rhizosphere metabolites were analyzed by OPLS-DA. Using a fold change ≤ 0.67 or ≥ 1.5 and VIP > 1 as thresholds, nine DEMs were identified in the high abundance group and the low abundance group. The categories of DEMs included lipids and lipid-like molecules, organic oxygen compounds, organoheterocyclic compounds, and benzenoids, of which lipids and lipid-like molecules accounted for the largest proportion (55.6%) (Fig. 5a). In the group with high ARG abundances (Fig. 5b), the content of Prostaglandin J2 (lipids), Gibberellin A14 (lipids), PA (16:0/15:0) (lipids), Erythromycin, and 3'-Deaminofusarochromanone was up-regulated. The content of the other five kinds of DEMs—PC (14:1[9Z]/16:0) (lipids), Methyl 2-(10-heptadecenyl)-6-hydroxybenzoate, PS (14:0/ 14:1[9Z]) (lipids), and coniferin—was down-regulated. Additionally, the VIP value of PA (16:0/15:0) (lipids) was the highest, which indicated that lipids in rhizosphere metabolites had a more obvious influence on endophytic bacteria spread and ARG distribution in different crops.
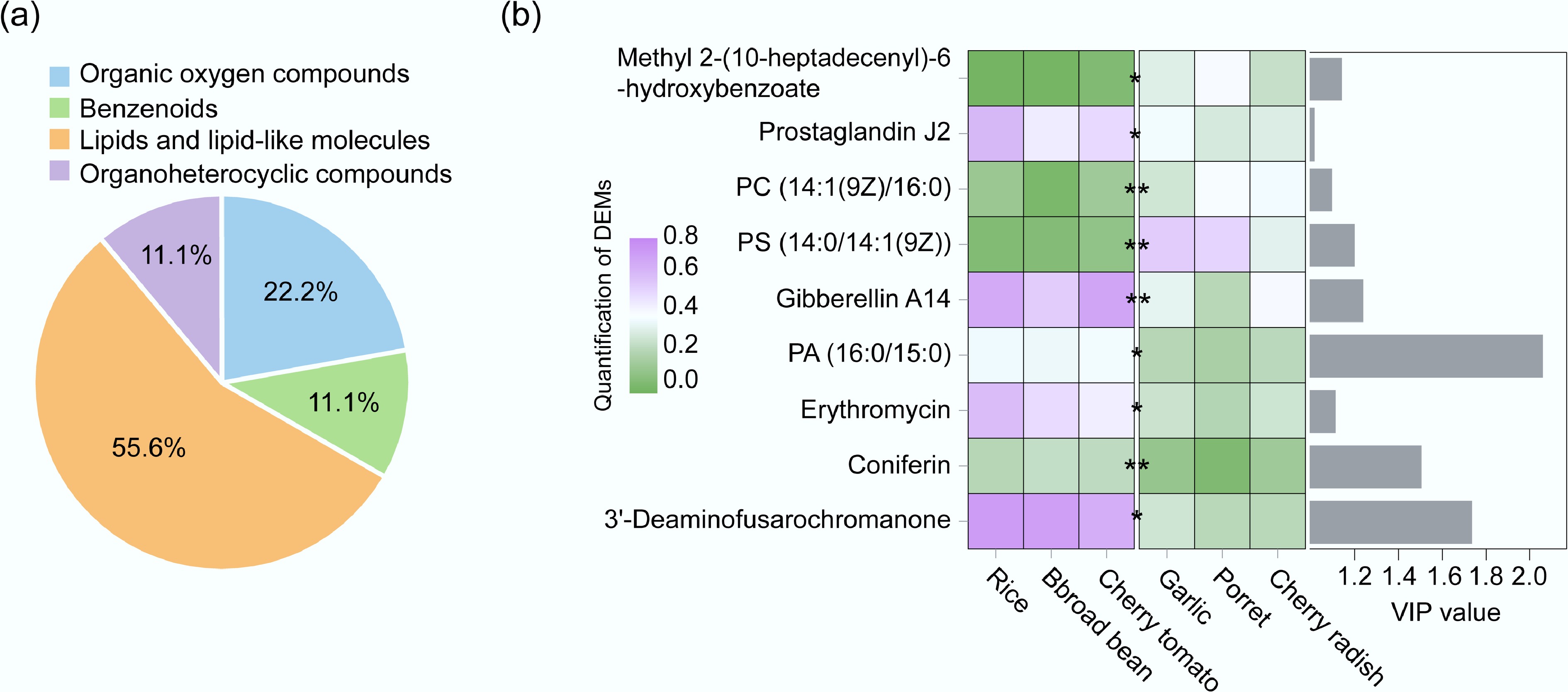
Figure 5.
(a) Percentage of DEMs in the high abundance crop group and the low abundance group in ARGs. (b) Content and VIP values of DEMs in the ARGs high-abundance crop group and low-abundance crop group.
Regarding the differences in ARG transmission among endophytic bacteria across different crops, this study further compared crops with high and low abundances of ARGs in edible parts. Mantel tests showed that most DEMs significantly affected bacterial community, with lipids PC (14:1[9Z]/16:0) and PA (16:0/15:0) showing significant correlations (p < 0.01, Fig. 6a). To further elucidate the relationship between rhizosphere metabolites and their potential bacterial hosts across plant species, a co-occurrence network was constructed integrating metabolites, bacterial communities, MGEs, and ARGs (Fig. 6b, c). In the high-abundance crop group, PA (16:0/15:0) and PS (14:0/14:1[9Z]) in lipids significantly affected key potential host bacteria Streptomyces, Phycicoccus, and Pantoea, and further affected mexF, mefA, ampC-06, qacH-01, dfrA12, and oprD in ARGs (p < 0.05). The potential host bacteria in ARGs (tetQ, dfrA12, mexF and mefA) in the low-abundance crop group were Nocardioides and Streptomyces, which were mainly influenced by PC (14:1[9Z]/16:0) and Gibberellin A14. Furthermore, SEMs analysis further described relationships between ARG abundance differences in endophytic bacteria of different plants and the main driving factors (Fig. 6d, e). In ARGs high-abundance crop group, MGEs (λ = 0.32, p < 0.05), microbial communities (λ = 0.43, p < 0.05), and DEMs (λ = 0.21, p < 0.05) positively affected ARGs, with DEMs also indirectly affecting ARGs via microbial communities (λ = 0.43, p < 0.05). In ARGs low-abundance crop group, MGEs (λ = 0.49, p < 0.05), microbial communities (λ = 0.50, p < 0.01), and DEMs (λ = 0.33, p < 0.05) also had significant positive effects on ARGs, with DEMs primarily affecting ARGs through microbial succession (λ = 0.76, p < 0.05). Hence, differential metabolites drive the difference in ARGs transport from different plant soils to endophytes by influencing the succession of microbial communities.
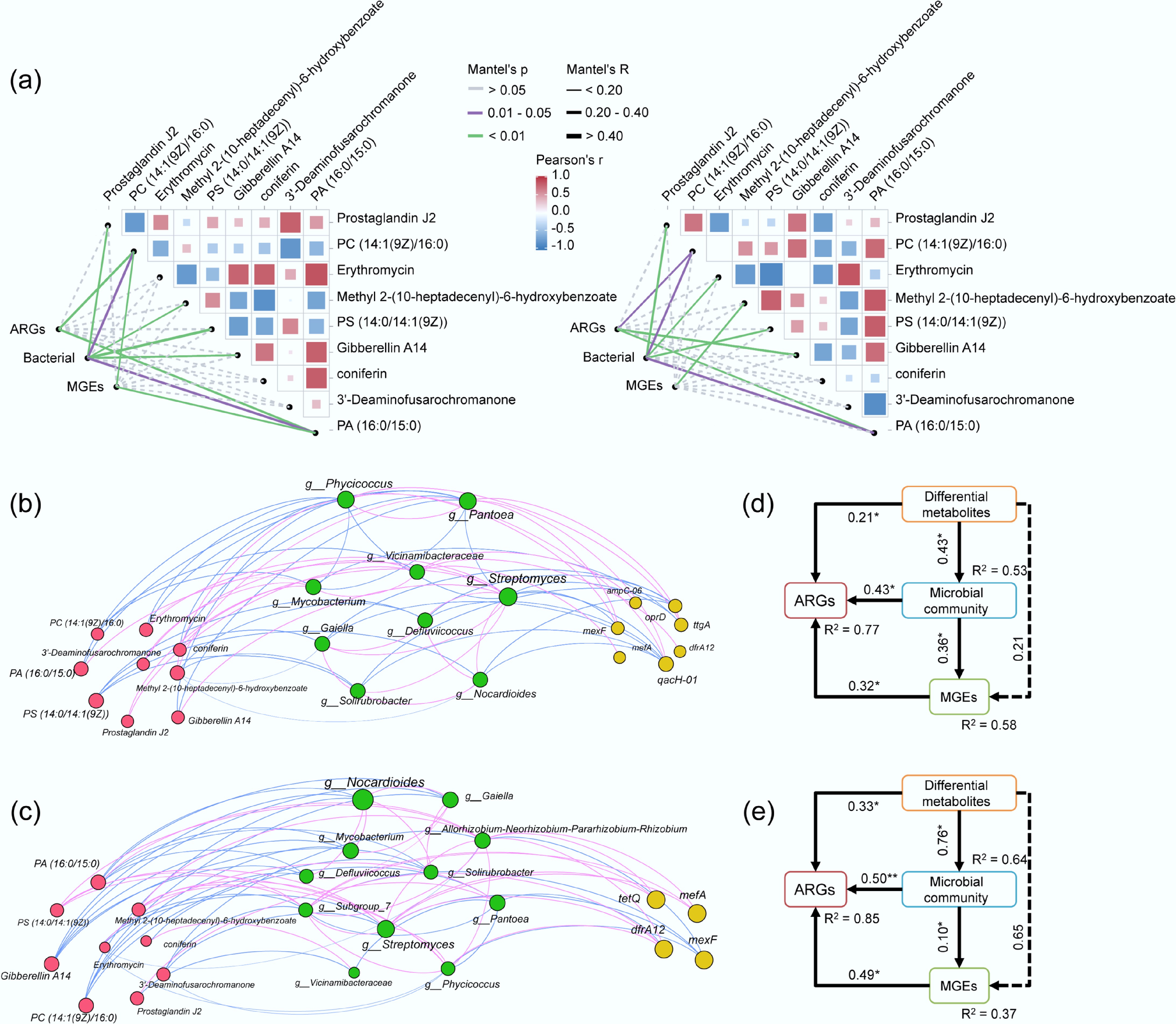
Figure 6.
Mantel test results showing the correlation between DEMs, MGEs, ARGs, and bacterial community in the (a)-left high-abundance crop group, and (a)-right the low-abundance crop group. (b) Co-occurrence networks based on Spearman's rank correlation coefficients in the high-abundance crop group, and (c) the low-abundance crop group. Nodes signify the abundance of individual ARGs, DEMs, or bacterial taxa. Positive and negative relationships are represented by pink and blue arrows, respectively. The SEMs in (d) and (e) depict the effects of MGEs, bacterial communities, and DEMs on ARGs for the high- and low-abundance crop groups, respectively.
-
The results of this study indicated that there are significant differences (p < 0.05) in the distribution of ARGs between rhizosphere and non-rhizosphere soils of different plants, with the total abundance of ARGs in rhizosphere soil being higher than that in non-rhizosphere soil, specifically 1.24 times that of non-rhizosphere soil. Previous studies have shown that both bacterial community composition and biomarkers differ between rhizosphere soil and non-rhizosphere soils under a certain concentration of antibiotic stress, and the absolute abundance of some ARGs in rhizosphere soil is higher than that in non-rhizosphere soil[34]. Yue et al.[7] found that a greater diversity and higher abundance of ARGs were detected in peanut rhizosphere soil, and were higher than those in non-rhizosphere soil, possibly because ARBs were contained in microorganisms responding to peanut rhizosphere metabolites. Additionally, the relationships between ARGs, MGEs, and microorganisms in rhizosphere soils may be more complex than those in non-rhizosphere soils (i.e., key regions of HGT)[40]. In this study, the results showed that the number and abundance of ARGs in endophytes in different crops were significantly lower than those in rhizosphere soil (p < 0.05). Previous studies have consistently demonstrated that the ARG abundance in plant endophytes is significantly lower than levels in rhizosphere soils[41,42]. The abundance level of manure-derived ARGs in the soil-plant system gradually decreased from the roots to the edible parts of plants. The ARG transmission may be associated with the colonization of plants by their potential microbial hosts[43,44]. That is, ARGs in many root endophytes are susceptible to the influence of ARGs in rhizosphere soil, while a few ARGs may further spread to the endophytes in the edible parts of plants.
In this study, it was observed that different crops had different subtypes of common ARGs in the soil-plant system, and the ARG profiles were also variable, indicating that different plant species affect the distribution and transmission differences of ARGs in the soil-plant systems. Previous studies have also shown that plant types determine the ARG profiles in the soil microbiome and endophyte microbiome[45]. In this study, the distribution of ARGs in different edible parts of plants showed that the abundance of ARGs in leaf plants was higher than that in fruit plants, with a difference of approximately one order of magnitude, indicating variations in ARG transmission from plant roots to edible tissues and differing resistance risks among different edible parts of plants. Crop types influence the distribution characteristics and transmission routes of ARGs in the soil-plant system. ARGs with stronger transmission may be more readily absorbed and colonized by endophytic bacteria in stems and leaves compared to fruits[46,47]. Additionally, compounds related to plants, such as rhizosphere metabolites, can regulate the microbial composition in the soil and the endophytic microbial communities of plants through microorganisms[48,49], which is also consistent with the results of this study. Therefore, different plant types limit ARG distribution in soil-plant systems, and plant-specific regulation of endophytic communities leads to differences in the potential risk of ARGs in different edible parts.
Lipid metabolites in plant roots coordinated the transport differences of ARGs in different crops by regulating microbial communities
-
The spread of ARGs in the soil-plant system is influenced by multiple factors, and the difference in ARGs is influenced by bacterial communities, MGEs, and differential metabolites. In this study, a significant positive correlation (r = 0.92, p < 0.001) between cIntI-1 (clinic) and ARGs in rhizosphere soils, plant roots, and edible parts indicated that MGEs play an important role in the spread of ARGs from soil to plant. The abundance of MGEs followed trends similar to those of ARGs, exhibiting enrichment in rhizosphere soil while progressively decreasing in plants. Integrons may increase ARG dissemination risks between humans and the environment by facilitating the spread of ARGs[50,51]. The capacity of MGEs to facilitate the spread and transfer of ARGs to plants varies across different soil-plant systems, thereby potentially impacting human food safety to varying extents[52−54]. Thus, rhizosphere soil and endophytic bacteria across plant species harbor MGE-carrying microorganisms that promote the dissemination of ARGs within plant-associated microbial communities.
By analyzing the driving factors of ARGs transmission difference between the high-abundance and low-abundance ARGs groups in edible endophytic bacteria, it was found that lipids and lipid-like molecules were highly correlated with the main host bacteria of ARGs. The results of this study indicated that within high-abundance crops, lipid metabolites PA (16:0/15:0) and PS (14:0/14:1[9Z]) significantly influenced key potential host bacteria (Streptomyces, Phycicoccus, Pantoea), consequently modulating ARGs including mexF, mefA, ampC-06, qacH-01, dfrA12, and oprD (p < 0.05). The potential host bacteria in ARGs (tetQ, dfrA12, mexF, and mefA) in the low abundance crop group were Nocardioides and Streptomyces, which were mainly affected by PC (14:1[9Z]/16:0) and Gibberellin A14. These findings demonstrated that lipid substances within rhizosphere metabolites critically regulate ARG transmission across plant species through plant-microbe interactions. Lipids play an essential role in the physiological functions of the plant plasma membrane, including the regulation of hormone signaling and transport, responses to abiotic stress, and interactions between plants and microbes[55−57]. In the rhizosphere microzone, plant-microbe signaling is a critical underground life activity, with lipids being highly involved[58,59], potentially further affecting the succession process of microorganisms. Lipids are the main components of plant plasma that can create physical barriers by affecting the surface of living cells[60]. Variations in lipid distribution across crop species drive differential plant-microbe signaling[56,61], thereby shaping microbial colonization patterns and ARG distribution. Furthermore, the relatively low abundance of ARGs observed in the edible fruits of cherry tomato may be attributed to their secretion of antimicrobial compounds such as tomatine and other glycoalkaloids. Studies have found that various metabolites derived from vitamin C-rich fruits, including tomatoes, exhibit potent antibacterial activity[62]. Some of these antimicrobial substances may selectively inhibit host bacteria associated with ARGs and reduce the potential for transmission and dissemination of ARGs in the rhizosphere environment[63,64]. Therefore, the variability of root metabolites in different crops regulates microbial community succession and further leads to the differential distribution of ARGs in the soil-plant system by affecting the colonization process of host microorganisms.
-
This study explored the influence of rhizosphere metabolites of different crops on ARG distribution in soil-plant systems. It was found that manure-derived ARGs mainly accumulated in rhizosphere soils and spread to the edible part of the plant, with the lowest abundance detected in the edible fruit plants and the highest in the edible leaf plants. Variations in rhizosphere metabolites among crops regulated ARG distribution, with lipid metabolites in the rhizosphere particularly influencing interactions between rhizosphere microorganisms and plants. The cIntI-1(clinic) of MGEs exhibited a significant correlation with the abundance of ARGs in endophytic bacteria in plant roots and edible parts, which notably affected the spread of ARGs in plants. These results underscore how plant root systems and their associated metabolites shape the dissemination of ARGs, providing a scientific basis for developing targeted strategies to mitigate antibiotic resistance propagation in agricultural soil environments.
-
It accompanies this paper at: https://doi.org/10.48130/aee-0025-0005.
-
Not applicable.
-
The authors confirm their contributions to the paper as follows: study conception and design: all authors; conceptualization: Zhang K, Ding Y, Yang F; manuscript review and editing: Wu Q, Ding Y, Yang F; formal analysis: Han B, Xu H, Shen S, Yang F; visualization: Han B, Shen S; methodology: Han B, Xu H; project administration, funding acquisition: Yang F; investigation, original draft preparation: Han B. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets used or analyzed during the current study are available from the corresponding author on reasonable requests.
-
This study was financially supported by the National Natural Science Foundation of China (Grant Nos 42277033 and 42307543), Basic Research Foundation of Yunnan Province of China (Grant No. 202401AT070304), the Youth Innovation Program of the Chinese Academy of Agricultural Sciences (Grant No. Y2024QC28), Tianjin Municipal Natural Science Foundation (Grant No. 23JCYBJC00250), and Natural Science Foundation of Shandong Province (Grant No. ZR2023QD008).
-
The authors declare that they have no conflict of interest.
-
Full list of author information is available at the end of the article.
- Supplementary Table S1 The primers of ARGs used for high-throughput quantitative PCR.
- Supplementary Fig. S1 Regression analysis of the relative abundance of MGEs and ARGs.
- Supplementary Fig. S2 Relative abundance of the top 20 genus level bacteria with the highest abundance. Abbreviation: B, broad bean; G, garlic; R, rice; CT, cherry tomato; C, chili; E, eggplant; CR, cherry radish; P, porret; CK, treatments without fertilizer; M, application of manure treatments; R, rhizosphere soils; NR, non-rhizosphere soils; G, plant roots; Y or J or F or M, edible parts of plants.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Han B, Xu H, Shen S, Wu Q, Zhang K, et al. 2025. Rhizosphere metabolites drive crop-specific distribution of manure-derived antibiotic resistance genes. Agricultural Ecology and Environment 1: e005 doi: 10.48130/aee-0025-0005 

Rhizosphere metabolites drive crop-specific distribution of manure-derived antibiotic resistance genes
- Received: 11 June 2025
- Revised: 04 September 2025
- Accepted: 12 September 2025
- Published online: 29 September 2025
Abstract: Rhizosphere metabolites profoundly affect the microorganisms in soil-plant systems. However, how this process drives the distribution and propagation of manure-derived antibiotic resistance genes (ARGs) in soil-plant systems remains unclear. Herein, a pot experiment with eight crop species was conducted to explore the effects on ARGs and bacterial communities mediated by rhizosphere metabolites. Most ARGs were primarily enriched in rhizosphere soils, with an abundance that was 1.24 times higher than that in non-rhizosphere soils, while their diversity and abundance in plant roots and edible parts were significantly reduced (p < 0.05). Plants with edible leaves (or stems), such as porret, tended to enrich more ARGs than those with fruits, like cherry tomato. Significant differences were observed in rhizosphere metabolites among different crops, among which lipid metabolites from plant roots were a key factor that regulates ARG spread in different crops by affecting the interactions between rhizosphere microorganisms and plants. Meanwhile, cIntI-1(clinic) was positively correlated with ARGs in plant roots and edible parts of endophytes (p < 0.05), promoting the spread of ARGs in plants. This study highlighted the co-regulatory role of differential metabolites induced in ARG distribution, offering a novel perspective on the dynamic regulation mechanism of ARGs in different soil-plant systems.