










-
As a type of non-reducing soluble sugar, sucrose (Suc) is mainly biosynthesized by photosynthesis and can be transported over long distances through sieve tubes in the phloem to provide carbon and energy for various reservoir tissue cells in plants[1]. In the Suc metabolism pathway, the dynamic equilibrium between Suc synthesis and hydrolysis plays an important role in regulating cell turgor pressure, sink-source relationship, growth and development and stress responses[2]. At present, two Suc biosynthesis-related enzymes, SPS and SPP, have been reported as key regulators of Suc metabolism in plants[3]. In the Suc synthesis pathway, SPS catalyzes fructose-6-phosphate (F6P) and UDP-glucose (UDPG) to produce sucrose-6-phosphate, and then SPP further hydrolyzes sucrose 6-phosphate to yield Suc. Among them, SPPs contain the conserved and catalytic phosphonic acid hydrolase domain (S6PP, PF05116) in higher plants, which can further form S6PP-SPP_C structures with the carbon-terminal SPP_C (PF08472) domain[4,5]. SPSs possess three conserved domains, including sucrose synthetase domain (Sucrose-synth, PF00862), glycosyltransferase domain (Glycos-transf-1, PF00534) and phosphohydrolase domain (S6PP). Meanwhile, three phosphorylation sites of SPSs, including Ser-158, Ser-229 and Ser-424, were relatively conserved in higher plants[6−8].
Due to the important roles of SPP and SPS, more and more SPS and SPP genes have been identified and functionally verified in different plant species[9−13]. Functional analysis studies found that SPSs were widely involved in plant growth and development and stress responses. In Arabidopsis, mutation of SPS gene inhibited Suc synthesis, the development of rosettes, flowers, and horn fruit, and also the seed germination both in spsa1 and spsc mutants[14]. A similar result was also found by Bahaji et al.[15] where they found the growth of rosettes, flowers and siliques in spsa1/spsc and spsa1/spsa2/spsc mutants were hindered, respectively. Besides, spsa1/spsb/spsc and spsa1/spsa2/spsb/spsc mutants also exhibited poor seed germination and produced abnormal and sterile plants[15]. In apple, the expression of MdSPSA2.3 was positively correlated with Suc accumulation, and silencing of MdSPSA2.3 significantly decreased the Suc content in fruit, suggesting that MdSPSA2.3 plays a dominant role in Suc synthesis of apple[16]. Western-blot analysis found that the abundance of SPS increased in the leaves of Miscanthus × giganteus and chilling-sensitive Zea mays line under low temperature conditions, but not in chilling-tolerant Zea mays. Meanwhile, SPS labelling was significantly increased in the leaves of chilling-sensitive Zea mays line, especially in mesophyll cells, under low temperature conditions[17].
SPP is a rate-limiting enzyme that catalyzes the SPS reaction product, Suc-6-phosphate, to dephosphorylation and release Suc[4,5]. In plants, the number of SPPs is less than that of SPSs. Currently, partial SPPs have also been identified from different plant species[18,19]. In recent years, the functional research of SPP has been gradually developed. In wheat, 1-bp insertion-deletion (InDel) and three single nucleotide polymorphisms (SNPs) mutation events occurred in the coding region of TaSPP-5A, resulting in two haplotypes of TaSPP-5Aa and P-5Ab. Expression analysis found that the expression level of TaSPP-5Aa in the leaves of seedling wheat was higher than that of TaSPP-5Ab, which was also positively correlated with the increase of Suc content and thousand-grain weight. Besides, the expression of TaSPP-5A and Suc content in TaSPP-5Aa haplotype were higher than those in TaSPP-5Ab haplotype under 20% PEG-6000 and 100 μM ABA conditions, respectively[20]. In tobacco, inhibition of SPP expression directly decreased SPP activity by 10%, retarded the chlorophyll content, photosynthesis and growth rate of transgenic tobacco, and also reduced the contents of Suc, Glc and Fru in RNAi lines[21]. In sorghum, the expression of Sobic.009G040900 was down-regulated 30% by PEG treatment, but up-regulated by Glc and Suc treatments; Sobic.009g041000 was upregulated 30% by PEG, NaCl, cold, Glc and Suc treatments. Overexpression of Sobic.009G040900 significantly reduced the seed germination rate of the transgenic Arabidopsis under 150 mM NaCl conditions, suggesting a negative role of Sobic.009G040900 in dealing with salt stress[12].
Tea plant (Camellia sinensis) is a typical perennial evergreen plant, the growth and development of tea plant usually affected by various biotic and abiotic stresses. Previous studies have demonstrated that Suc metabolism is widely involved in vegetative and reproductive growth, and also in responding to low temperature, drought, and salt stresses[22,23]. With the completion of tea plant genome sequencing, a series of genes involved in Suc metabolism, such as CsINVs[24,25], hexokinase (CsHXKs)[26] and sugar transporters (CsSWEETs)[27,28], have been identified and functionally verified. In terms of the study on the molecular mechanism of Suc biosynthesis in tea plant, a SPS gene (CsSPS) was cloned previously, which showed differential transcriptions in different tea plant tissues and under cold treatment conditions[29,30]. Besides, Yang et al. found that a SPS gene was significantly induced by low temperature treatment, meanwhile, exogenous γ-aminobutyric acid (GABA), green algae powder and bamboo vinegar could further significantly induce the expression of CsSPS under low temperature conditions[31]. Despite the above studies, how many SPS and SPP genes are contained in tea plants, and what roles do they play? These questions have not yet been resolved. Therefore, based on the sequenced tea plant genomes, the present study carried out the genome-identification of CsSPSs and CsSPPs, and analyzed their biological information, tissue-specific and spatio-temporal expression patterns. In addition, the roles of CsSPSs and CsSPP in two tea plant cultivars, 'Shuchazao' ('SCZ') and 'Baiye 1' ('BY1'), were investigated under cold stress conditions. For these tea plant cultivars, 'SCZ' was reported as a cold-resistant tea plant cultivar[32], which has completed genome sequencing[33]; 'BY1', as a type of temperature-sensitive albino mutant, was reported to be a cold-sensitive tea plant cultivar[34]. The results will further enrich the theory of the molecular mechanisms of Suc metabolism regulation, and lay a foundation for further investigation of the function of Suc in tea plants.
-
One bud and two leaves, mature leaves, senescent leaves, flower buds, flowers, young fruits, immature stems, mature stems and roots of the ten-year-old tea plant cultivar 'SCZ' were sampled for tissue-specific analysis. The detailed sampling method was carried out as reported by Wang et al.[35].
The two-year-old clonal cuttings of 'SCZ' and 'BY1' were used to perform cold treatment. Before cold treatment, all tea cuttings were firstly cultured in the plant growth chamber for one week with the following growth parameters: 25 °C, 85% relative humidity, 100 μmol·m−2·s−1 light and 16 h light/8 h darkness. Then, the temperature of the climate chamber was dropped to 0 °C for cold treatment without changing the other parameters, and a total of 5 d of cold treatment was carried out. Finally, the temperature was turned up to 25 °C for 3 d of recovery. The third to fifth mature leaves of each tea plant cultivar were collected after 0 d and 5 d of 0 °C treatment, and also 3 d of 25 °C recovery. For abiotic stress treatments, 10% (w/v) polyethylene glycol (PEG), 150 mmol·L−1 NaCl and 4 °C were performed respectively to simulate drought, salt and cold stresses as described by Wang et al.[35]. Briefly, one-year-old tea cuttings of 'SCZ' cultivar with a similar growth state were cultured in the climate chamber for one week, some of these were then fed with 10% PEG and 150 mmol·L−1 NaCl to simulate drought and salt stresses, respectively. In addition, some of the tea cuttings were moved into a 4 °C climate chamber for cold treatment. After 0, 12, 24, and 48 h of each stress treatment, the third to fifth mature leaves of tea cuttings were collected for expression analysis. Each treatment processed three biological replicates, and each replicate contained ten tea cuttings with similar growth states. All collected samples were quickly frozen in liquid nitrogen and stored at −80 °C until use.
Genome-wide identification of CsSPSs and CsSPP genes
-
The genome-identification procedure of CsSPSs and CsSPP was carried out following the method as described by Li et al.[36]. Firstly, the conserved Hidden Markov Models (HMM) of SPS and SPP, including PF05116, PF08472, PF00862, and PF00534 were obtained from protein families (Pfam) database[37]. Following, the above domains were respectively matched to the protein databases of the 'SCZ'[33], 'Tieguanyin' ('TGY')[38] and 'Huangdan' ('HD')[39] by using HMMER 3.0 software. Subsequently, the obtained sequences were respectively submitted to the simple modular architecture research tool (SMART) server[40] and the conserved domain database of national center for biotechnology information (NCBI)[41] for confirming whether they belong to the SPP and SPS families. Finally, the above qualified sequences containing the SPS and SPP functional domains were used for bioinformatics and expression analysis.
Bioinformatics analysis of CsSPSs and CsSPP
-
The NCBI ORF finder website (
www.ncbi.nlm.nih.gov/orffinder ) was used to predict the opening reading frame (ORF) lengths of CsSPS and CsSPPs. The protein parameter (ProtParam) tool[42] was used to calculate the molecular weights, theoretical pI and aliphatic index of CsSPP and CsSPSs. The signal peptide (SignalP) server[43] and the transmembrane protein topology with a Hidden Markov Model (TMHMM) Server v.2.0[44] were respectively used to predict the signal peptides and transmembrane regions (TMHs), and TargetP 2.0[45] was used to predict the sub-cellular location of CsSPSs and CsSPP.Phylogenetic analysis of CsSPSs and CsSPP
-
There are 46 SPSs and 18 SPPs originating from tea plant, Arabidopsis thaliana, Oryza sativa, Spinacia oleracea, Nicotiana tabacum, Cucumis melo, Citrus unshiu, Vitis vinifera, Triticum aestivum, Solanum tuberosum, Litchi chinensis, Sorghum bicolor Solanum lycopersicum, and Solanum lycopersicum, were used to construct a phylogenetic tree by using the neighbor-joining method of MEGA 7.0 software[46]. The detailed parameters were as follows: 1000 repeated bootstrap tests, p-distance method and pairwise deletion treatment. Then, Evolview[47] was used to annotate and manage the phylogenetic tree.
Chromosomal distribution and synteny analysis of CsSPSs and CsSPP
-
The chromosomal of CsSPSs and CsSPP, and the inter-species collinearity analysis between 'SCZ' cultivar and Arabidopsis, 'HD' and 'TGY' genomes were performed and visualized by using TBtools software respectively[48]. The genome data of Arabidopsis was downloaded from the NCBI website (
www.ncbi.nlm.nih.gov ). The genomes of 'TGY'[38] and 'HD'[39] were respectively obtained from national genomics data center (NGDC)[49].Gene structure, protein domain distribution and cis-acting element analysis
-
TBtools software was used to predict and display the exon-intron structures, protein domain distribution[48]. In order to explore the types and quantities of cis-acting elements in CsSPSs and CsSPP promoter regions, TBtools was used to extract 2000-bp upstream non-coding region sequence of the translation initiation site (ATG) in each CsSPSs and CsSPP genome sequence, then each sequence was submitted to plant cis-acting regulatory element (PlantCARE) web server[50] for predicting putative cis-acting elements involved in responding to stresses and hormones. Finally, TBtools was used to visualize the prediction results in the form of a heatmap[48].
Co-expression network analysis of CsSPSs and CsSPP
-
Firstly, the TPIA online website (
http://tpia.teaplants.cn/geneIdConvert.html ) was used to convert the version 2 'SCZ' genome IDs of CsSPP and CsSPSs into the version 1 'SCZ' genome IDs[51]. Then, the target IDs were uploaded to the TeaCoN website using 261 high quality RNA-Seq data, for constructing the gene co-expression network with Pearson correlation coefficients (PCC-values) > 0.7 and statistical p-values < 0.05 following the method as described by Zhang et al.[52]. Finally, the resulting CSV annotation file was downloaded and submitted to the Graphbiol website (www.graphbio1.com/ ) for further embellishment and beautification.qRT-PCR analysis
-
Total RNA was isolated from different samples as mentioned above by using the RNA extraction kit (Bioflux, Hangzhou, China). Immediately, the first-strand cDNA was synthesized using the reverse transcription kit (Takara, Dalian, China). The programs and systems used for qRT-PCR were conducted as described by Wang et al.[35]. Polypyrimidine tract-binding protein (CsPTB) of tea plant[53] was used as the reference gene to quantify the relative expression of each CsSPSs and CsSPP. The results were calculated by the 2−ΔCᴛ or the 2−ΔΔCᴛ method[54], and visualized as the mean values ± standard error (± SE). The qRT-PCR primers are shown in Supplemental Table S1.
Physiological indicators determination
-
To compare the cold tolerance of 'SCZ' and 'BY1' cultivars after 2 d of 0 °C treatment, the relative electrolytic leakage (EL), malondialdehyde (MDA) content, the maximum quantum yield of PSII (Fv/Fm) and net photosynthetic rate (NP) were measured in this study. The EL was determined following the method described by Wang et al.[55]. The FluorPen FP 110 (Photon Systems Instruments, spol.sr.o., Drásov, Czech Republic) and LI-6400XT (LI-COR, USA) were used to measure Fv/Fm and NP following the instructions of the instrument, respectively. Three biological replicates were performed, and each replicate contained six tea cuttings with similar growth state. The SPS activity, MDA content, and the contents of total soluble sugar (TSS), Suc, Glc and Fru in 'SCZ' cultivar and 'BY1' cultivar were respectively measured using the corresponding measurement kits following the introduction of the reagent kits (Suzhou Comin Biotechnology, Suzhou, China).
Statistical analysis
-
The statistical differences were analyzed by One-way Analysis of Variance (ANOVA) followed by Duncan's test. Correlation heatmaps were drawn using online websites (
www.chiplot.online/correlation_heatmap.html ). Bar charts were drawn by using GraphPad Prism 6.0 (www.uone-tech.cn/graphpad-prism.html ). -
In this study, five CsSPSs (CsSPS1-5) and one CsSPP were identified from three tea plant genomes by using the conserved HMM models of SPS (PF00862, PF00534 and PF05116) and SPP (PF05116 and PF08472), respectively. As shown in Table 1, CsSPP is highly conserved among three tea plant cultivars, except for two and one non-synonymous mutations in 'SCZ' and 'HD' genomes, respectively (Supplemental Fig. S1). In terms of CsSPSs, although the amino acid sequence length of the same SPS may be varied in different tea plant cultivars, each CsSPS was also highly conserved among these three tea plant cultivars. In particular, the amino acid sequence of CsSPS5 was identical in these three cultivars except for a non-synonymous mutation in the 'SCZ' genome. Besides, CsSPS1 was only identified in the 'SCZ' genome, which may be the product of the tandem repeat of CsSPS2 in the 'SCZ' genome, as CsSPS1 and CsSPS2 shared 99.4% amino acid sequence identity, located on the same chromosome, and only separated by six genes. Subcellular localization further predicted that CsSPP and CsSPSs were located in cytoplasm. In brief, these results indicate that CsSPP and CsSPSs in different tea plant cultivars possess the same function as the 'switch' of Suc biosynthesis.
Table 1. Basic information of CsSPP and CsSPSs.
Gene Accession number ORF (bp) AA MW (KDa) pI Aliphatic index Loc SignalP TMHs CsSPP CSS0017072.1('SCZ')
GWHPASIV039206 ('TGY')
GWHPAZTZ037371 ('HD')
GWHPBAUV077964 ('HD'-HB)
GWHPASIX044577 ('TGY'-HA)
GWHPASIX046144 ('TGY'-HB)1,275 424 48.13
48.11
48.10
48.11
48.11
48.115.55
5.62
5.55
5.62
5.62
5.6281.37
82.29
82.29
82.29
82.29
82.29Cytoplasm NO NO CsSPS1 CSS0047114.1('SCZ') 2,988 995 111.15 5.65 86.04 Cytoplasm NO NO CsSPS2 CSS0020276.1 ('SCZ')
GWHPASIV037217 ('TGY')
GWHPAZTZ035335 ('HD')
GWHPBAUV071117 ('HD'-HA)
GWHPBAUV073449 ('HD'-HB)3,057
2,796
2,916
2,916
2,9161018
931
971
971
971113.16
103.18
107.81
107.81
107.835.71
5.76
5.76
5.76
5.7686.58
87.65
86.86
86.86
86.86CsSPS3 CSS0009603.1 ('SCZ')
GWHPASIV029409 ('TGY')
GWHPAZTZ027893 ('HD')
GWHPBAUV055399 ('HD'-HA)
GWHPBAUV058548 ('HD'-HB)
GWHPASIX032838 ('TGY'-HA)
GWHPASIX034616 ('TGY'-HB)3,111
3,120
2,916
3,192
3,120
3,120
3,1201036
1039
1039
1039
1039
1039
1039116.78
117.77
117.66
215.79
117.66
117.66
117.775.92
6.32
6.26
6.53
6.26
6.21
6.3290.52
88.59
88.21
94.36
88.21
88.59
88.59Cytoplasm NO NO CsSPS4 CSS0024623.1('SCZ')
GWHPAZTZ027407('HD')
GWHPASIV029106('TGY')
GWHPBAUV054918('HD'-HA)
GWHPBAUV058088 ('HD'-HB)3,192
2,916
3,864
3,237
3,1921063
1063
1287
1078
1063119.71
119.64
144.79
119.19
119.646.05
6.00
6.01
6.10
6.0083.57
83.47
86.83
83.40
83.47Cytoplasm NO NO ORF, Opening reading fame; AA, The numbers of amino acid residues; MW, Molecule weight; pI, Theoretical isoelectric point; Loc, Subcellular location; SignalP, Signal peptide; TMHs, Transmembrane helices. 'SCZ', 'TGY' and 'HD' mean 'Shuchazao', 'Tieguanyin', and 'Huangdan', respectively. 'HA' and 'HB' represent haplotype A and haplotype B genomes of 'Huangdan' and 'Tieguanyin' cultivars, respectively. Phylogenetic analysis of CsSPSs and CsSPP
-
To explore the phylogenetic relationship among different SPSs and SPPs in different plant species, a phylogenetic tree was constructed. As shown in Fig. 1, all of these SPSs could be divided into four subfamilies (I−IV). As a typical dicotyledonous plant, CsSPS1 and CsSPS2 of tea plant were clustered into subfamily I and showed the closest relationship with MD02G1022300 and MD15G1164900. CsSPS5 was also clustered into subfamily I and showed the closest relationship with StSPS. CsSPS3 belonged to subfamily III, and showed the closest relationship with NtSPS3 and AtSPS4, while CsSPS4 belonged to subfamily II and showed the closet relationship with NtSPS2 and SlSPS2. In addition, the phylogenetic analysis of SPPs showed that the unique CsSPP presented the closest relationship with MD12G1045400 and MD14G1044300.
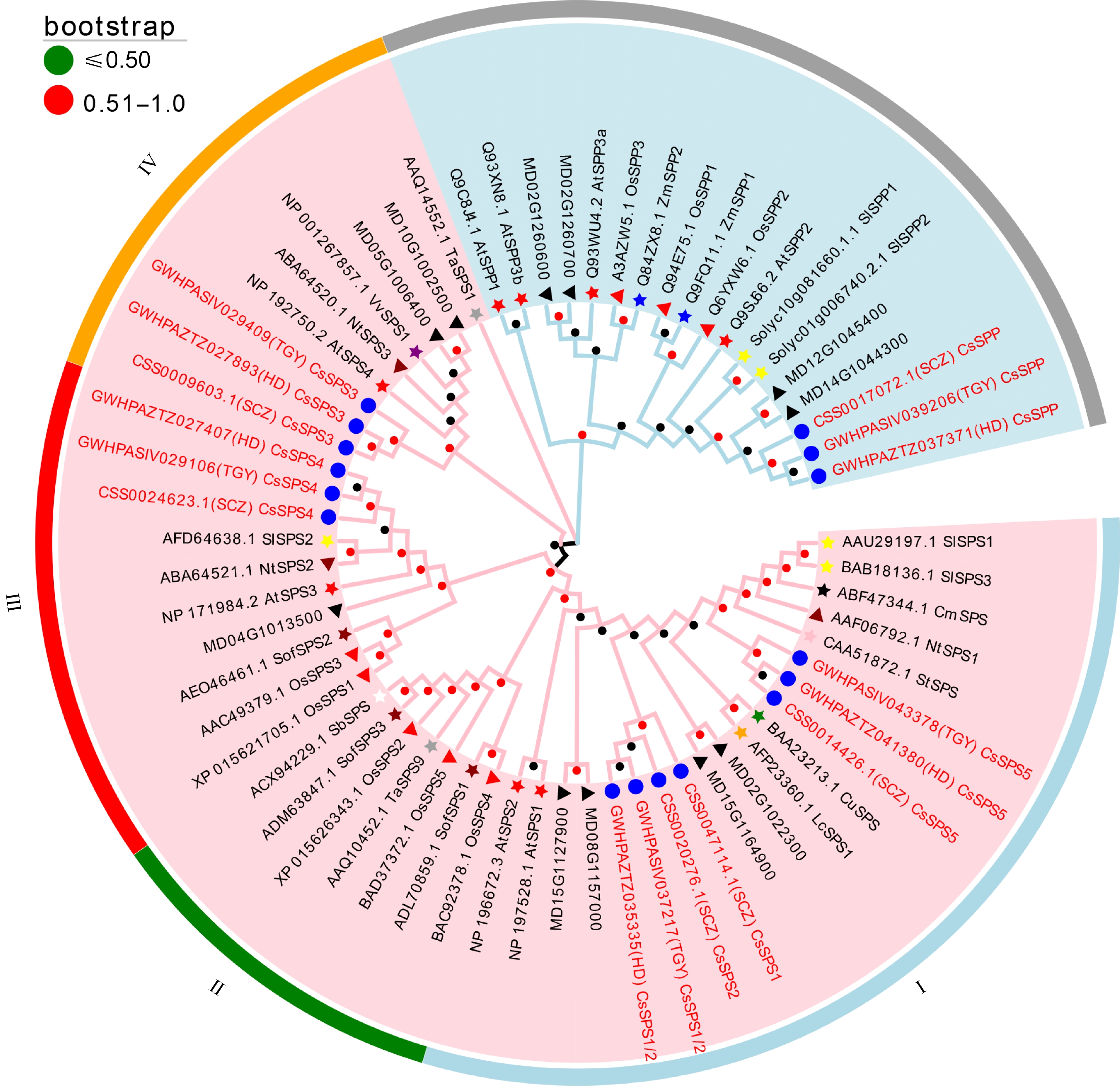
Figure 1.
Phylogenetic analysis of SPPs and SPSs originating from 15 different plant species. Pink area: SPS family; Light blue area: SPP family. Blue circle: tea plant; red star: Arabidopsis; red triangle: rice; blue star: maize; yellow star: tomato; dark red star: spinach; dark red triangle: tobacco; black star: melon; green star: citrus; purple star: grape; gray star: wheat; pink star: potato; orange star: litchi; white star: sorghum; black triangle; apple. Bootstrap values of all branches are above 50%.
Chromosomal distribution and collinearity analysis of CsSPSs and CsSPP
-
The chromosomal distribution of CsSPP and CsSPSs in three tea plant genomes was predicted and visualized by TBtools software. As shown in Fig. 2a and Supplemental Fig. S2, CsSPSs and CsSPP shared same chromosomal distribution in these three tea plant genomes, respectively. In detail, CsSPP located on Chr13, CsSPS1 and CsSPS2 co-located on Chr12, CsSPS3 and CsSPS4 co-located on Chr9, and CsSPS5 located on Chr14.
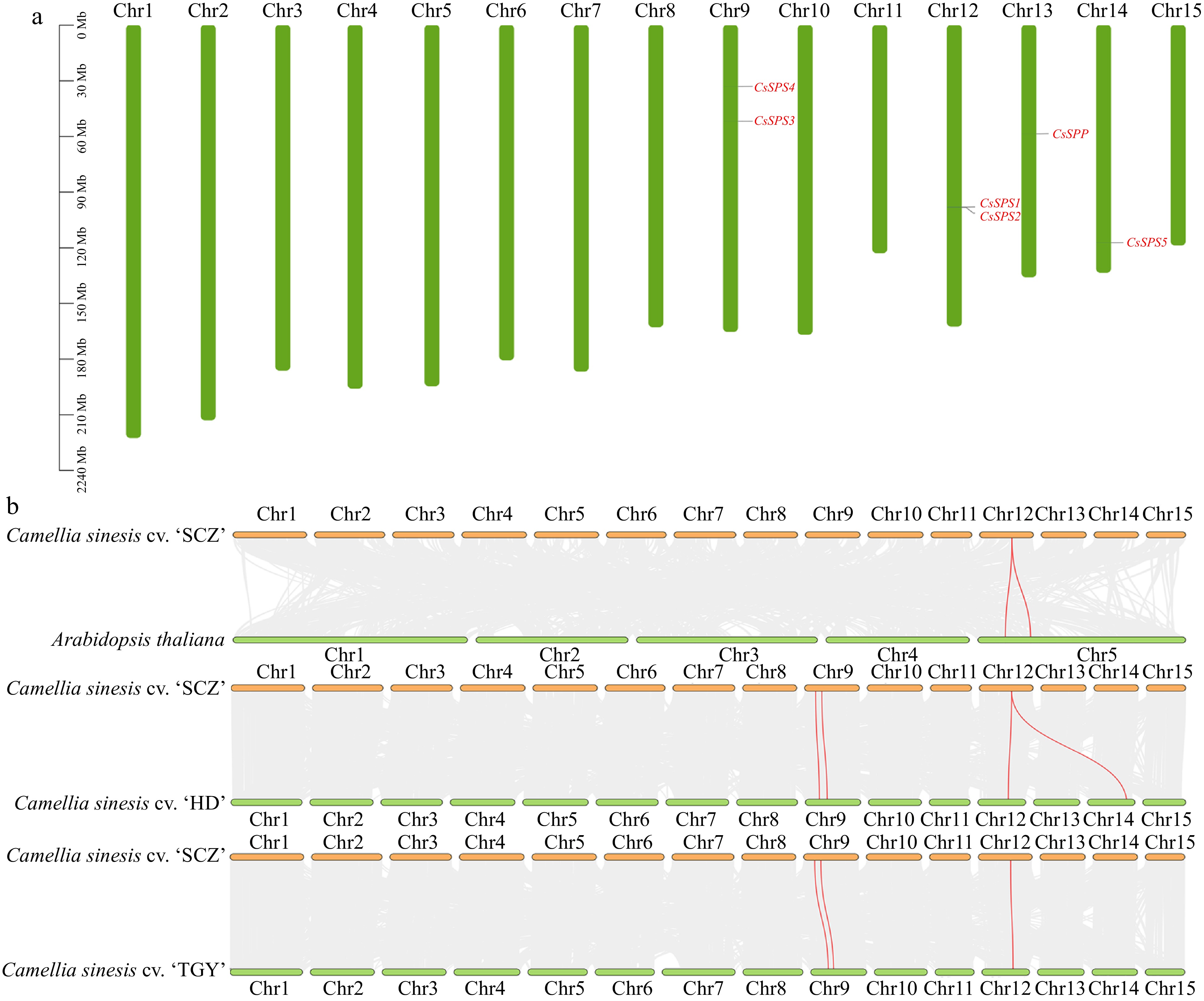
Figure 2.
Chromosomal location and collinearity analysis of CsSPP and CsSPSs. (a) Chromosomal distribution of CsSPP and CsSPSs in 'Shuchazao' genome. (b) Interspecies synteny analysis of CsSPP and CsSPSs in 'Shuchazao' associated with Arabidopsis, 'Huangdan' and 'Tieguanyin' genomes.
To further understand the evolutionary relationships of CsSPP and CsSPSs among different plant species, the inter-species collinearity relationships between 'SCZ' and 'HD', 'TGY' and Arabidopsis were constructed, respectively. As shown in Fig. 2b, both CsSPS1 and CsSPS2 belong to orthologous genes with AtSPS1 (NP197528.1) and AtSPS2 (NP196672.3) in Arabidopsis, HD.09G0012280.t1 and HD.12GOO24590.t1 in 'HD' cultivar, and TGY103558.t1 in 'TGY' cultivar. Besides, CsSPS3 and CsSPS4 are orthologous genes of HD.10G0021440.t1 and HD.10G0017080.t1 in 'HD' cultivar, and TGY080122.t1 and TGY081105.t1 in 'TGY' cultivar, respectively. These results also corresponded to the results of chromosome localization and phylogenetic analysis. In addition, the distribution and numbers of CsSPP and CsSPSs homologous genes in 'SCZ' genome were further explored through intra-special collinearity analysis, while there has no genome replication or fragment replication events occurred between CsSPSs and CsSPP (data not shown), indicating that CsSPSs and CsSPP are highly conserved in different tea plant cultivars.
cis-acting elements prediction and co-expression network analysis of CsSPP and CsSPSs
-
To understand whether CsSPP and CsSPSs are involved in stress and hormone responses, the cis-acting elements contained in 2000-bp 5'-terminal untranslated region (UTR) sequences of CsSPP and CsSPSs were predicted. As shown in Fig. 3a, the type, number and distribution of cis-acting elements in UTR sequences of CsSPP and CsSPSs were varied among each other. Overall, all of them contain numerous light responsiveness related elements (data not shown). Besides, myeloblastosis (MYB) and myelocytomatosis (MYC) elements were also enriched in these promoter regions. Meanwhile, different numbers of anaerobic induction element (ARE) were also found in these promoters, indicating that CsSPSs and CsSPP play important roles in photosynthesis and respiration of tea plants. Besides, different numbers of hormone response elements, such as auxin-responsive element (TGA), MeJA-responsiveness (MeJA) element, abscisic acid responsiveness element (ABRE), and gibberellin (GA) element were predicted in these promoters, especially 3 MeJA elements were respectively enriched in the promoter regions of CsSPS2 and CsSPS4, suggesting their central roles in responding to hormones. Moreover, low-temperature response element (LTR) elements were enriched in the promoter regions of the CsSPP and CsSPS1/3/5, indicating these genes participate in cold stress response of tea plants. Furthermore, we found the numbers and types of cis-acting elements were most abundant in promoter of CsSPS4, which suggested that CsSPS4 may be widely involved in various stress responses of tea plants. In short, the above results showed that CsSPP and CsSPSs play important roles in mediating hormones and abiotic stress responses.
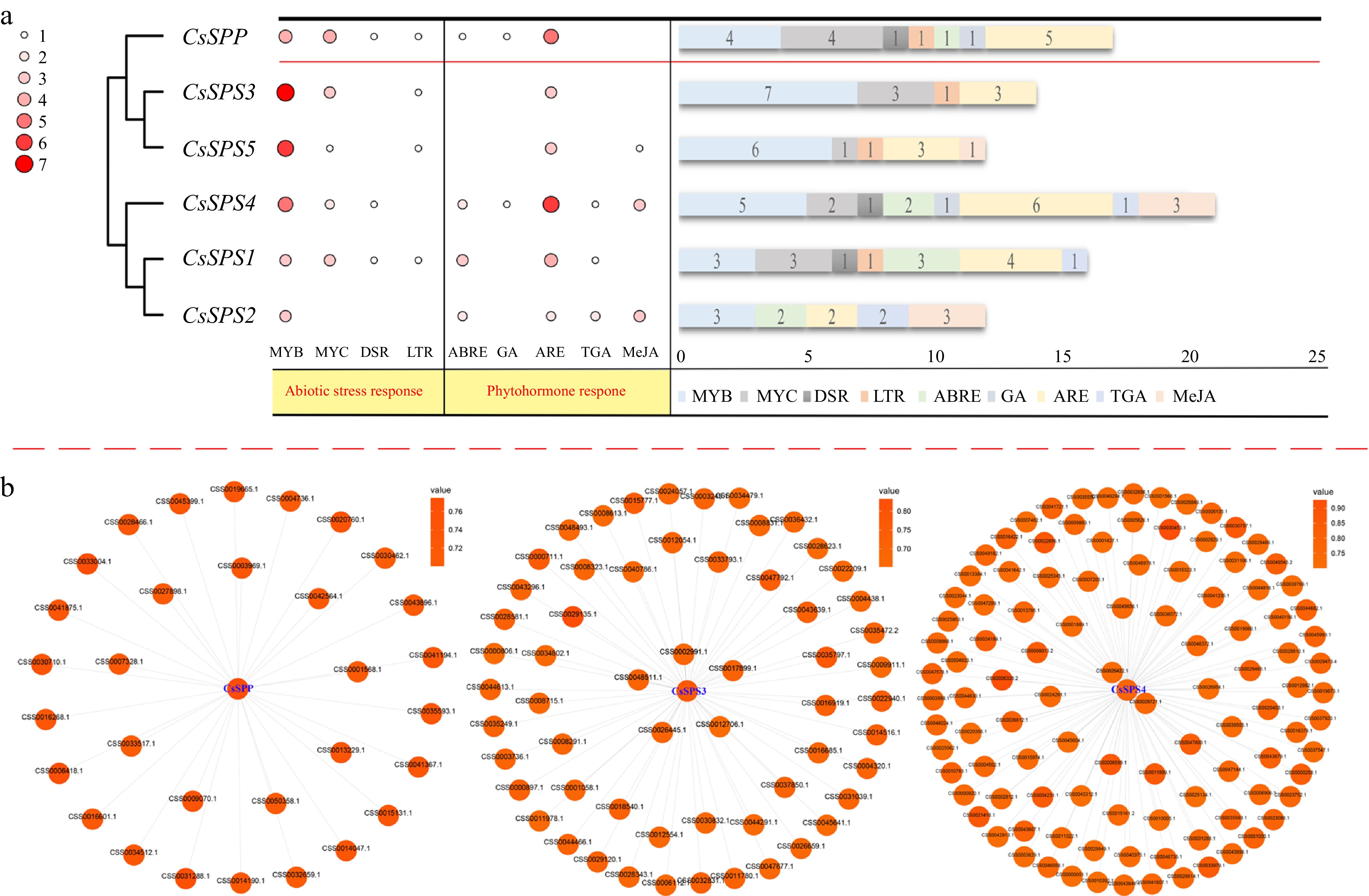
Figure 3.
The cis-acting elements in the promoters of CsSPP and CsSPSs, and co-expression networks of CsSPP and CsSPSs. (a) cis-acting elements in promoters of CsSPP and CsSPSs. The heat map displays the type and number of cis-acting elements and the bar chart displays the number of cis-acting elements. MYB: myeloblastosis; MYC: myelocytomatosis; DSR: defense and stress responsiveness; LTR: low-temperature responsiveness; ABRE: abscisic acid responsiveness; GA: gibberellin-responsiveness; ARE: anaerobic induction; TGA: auxin-responsive element; MeJA: MeJA-responsiveness. (b) Co-expression networks of CsSPP and CsSPSs 3/4.
Here, the co-expression networks of CsSPP and CsSPSs were also predicted with the help of the TeaCoN web server. As a result, only CsSPP, CsSPS3 and CsSPS4 predicted to contain 31, 70, and 110 co-expressed genes with strong associations (PCC-value > 0.7), respectively (Fig. 3b). Among them, most of the co-expressed genes of CsSPP are related to photosynthesis and respiration in plants. For example, a co-expressed gene of CsSPP, CSS0031288.1, encodes zeaxanthin epoxidase, which is involved in zeaxanthin synthesis and could adapt to different light intensity by controlling the amount of zeaxanthin accumulation in plant photosynthesis. Similar to CsSPP, the highly correlated genes of CsSPS3 were also related to photosynthesis and respiration. Besides, the expression profiles of two transcription factors, including CsMYB35 (CSS0014516) and CsGLOBOSA-like (CSS0022940), were highly correlated with CsSPS3, suggesting there may be a potential transcriptional regulatory relationship between them. Moreover, CSS0030453 (ATP synthase) and CSS0006328 (Peroxisomal membrane protein) are two highly correlated co-expressed genes in the co-expression network of CsSPS4. Among them, CSS0030453 plays an important role in cellular energy metabolism, plant photosynthesis and respiration, and CSS0006328 participates in scavenging free radicals.
DNA structures and conserved motifs of CsSPP and CsSPSs
-
The DNA structure analysis results showed that each of these genes contains more than 10 exons. Among them, CsSPP contains eight exons, CsSPS1 contains 14 exons, CsSPS3 contains 13 exons, CsSPS5 contains 11 exons, while CsSPS2 and CsSPS4 contain 12 exons, respectively (Fig. 4a). Based on the complex structures of these genes, we speculated that the functions of these genes may be irreplaceable in tea plants. Conserved motif analysis result showed that CsSPSs are highly conserved, and all of them contain 15 motifs except motif 14 which is missing in CsSPS3 (Fig. 4b). Besides, CsSPP is distinct from CsSPSs, indicating the different functions they played. This conclusion is further proved in Fig. 4c, where we found each CsSPS contains three conserved domains, including Sucrose_synth, Glycos_transf_1 and S6PP, while CsSPP contains the conserved S6PP and S6PP_C domains. In addition, the S6PP domain and the S6PP_C domain of CsSPP is closely connected. However, there are some amino acid sequences between the CsSPSs domains and a variable Linker between Glycos_transf_1 and S6PP. Moreover, some potential conserved serine phosphorylation sites were also identified in all five CsSPSs. Among them, two of the same phosphorylation sites, Ser191 and Ser385, were identified both in CsSPS1 and CsSPS2. Besides, Ser148 and Ser217 in CsSPS4, and Ser221 and Ser416 in CsSPS5 are also potential conserved phosphorylation sites, respectively. Moreover, three phosphorylation sites, Ser146, Ser220 and Ser409, were identified in CsSPS3. These results further confirmed that CsSPP and CsSPSs own Suc biosynthesis ability, and their activities are regulated by phosphorylation.
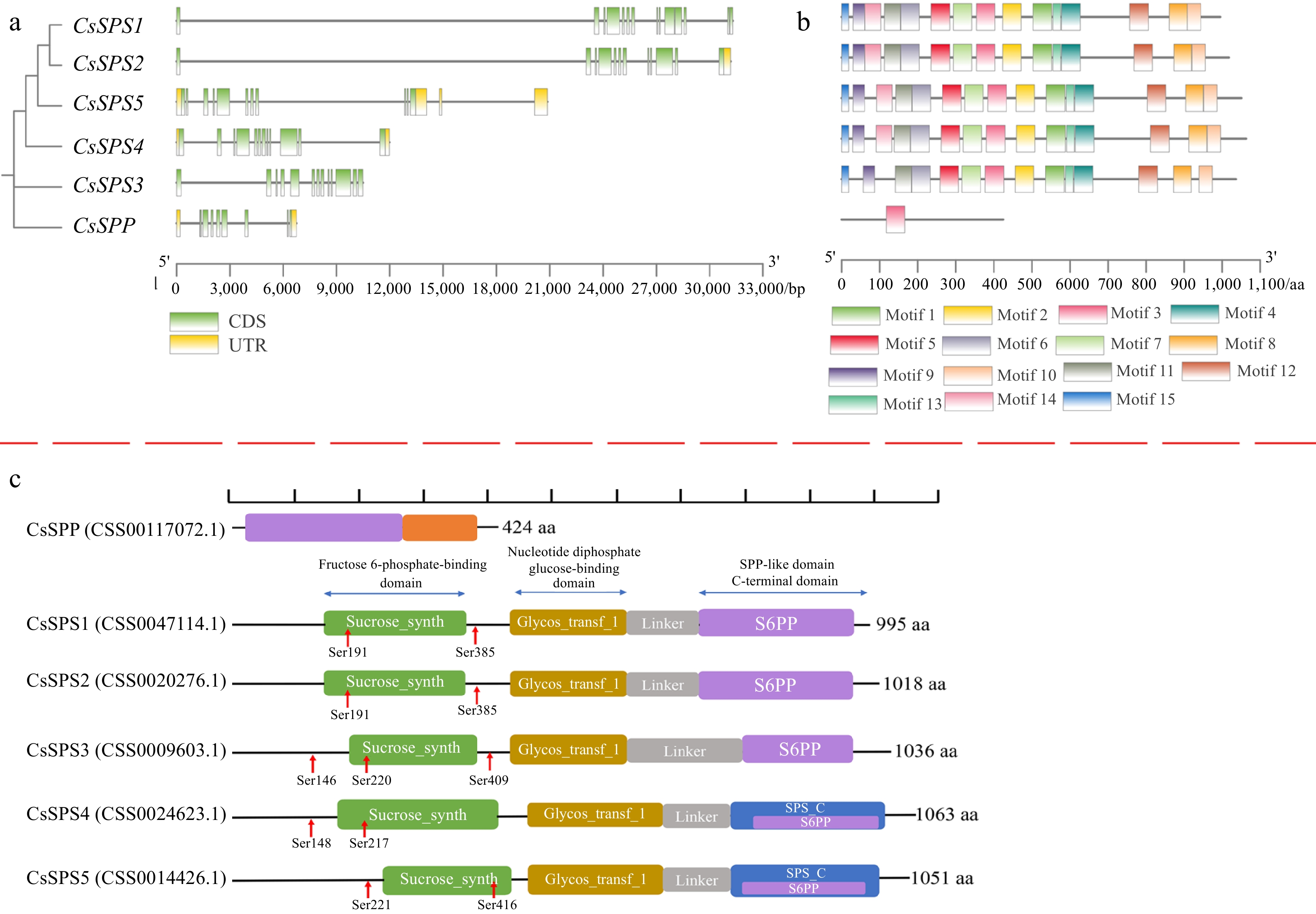
Figure 4.
The exon-intron structures CsSPP and CsSPSs, conserved motifs and domains of CsSPP and CsSPSs. (a) The exon-intron structures CsSPP and CsSPSs. Green boxes represent exons, yellow boxes represent untranslated upstream/downstream regions, and lines indicate introns. (b) Conserved motifs of CsSPP and CsSPSs. Different motifs are presented by different colored squares. (c) Conserved domains of CsSPP and CsSPSs. Different domains are shown in different colors.
Tissue-specific analysis of CsSPSs and CsSPP in different tea plant tissues
-
Tissue-specifics of CsSPSs and CsSPP were detected in nine different tissues of the 'SCZ' cultivar. As shown in Fig. 5, CsSPP and CsSPSs transcripts were detected in all tissues, but the transcription abundance of each gene varied among the detected tissues. Among them, the transcription abundance of CsSPP was highest in immature stem, while significantly lower in other tissues. Besides, all of CsSPSs showed highest transcription abundances in flower than that in other tissues, except for CsSPS2 that showed a similar expression level in senescent leaf. Meanwhile, all CsSPSs showed extremely low transcription abundances in root. In addition, the transcription abundance of CsSPS5 in each detected tissue was significantly higher than that in other CsSPSs, which speculated that CsSPS5 may play a leading role in Suc synthesis during the growth and development of tea plants. In brief, it followed that CsSPP and CsSPSs mediated entire vegetative and reproductive progress of tea plants. In particular, CsSPSs may play important roles in floral nectar production of flower, and CsSPP is necessary for the immature stem growth of tea plants.
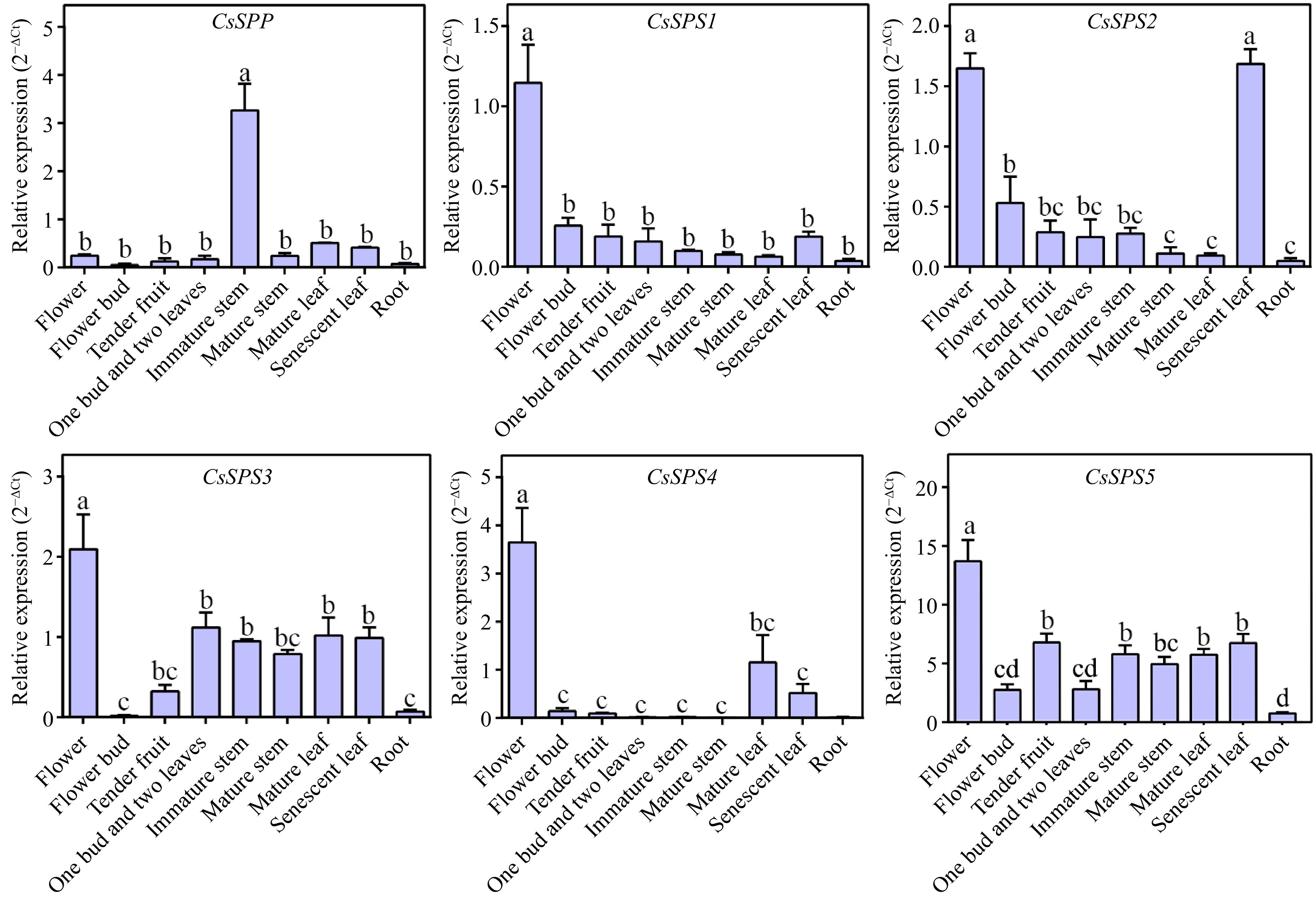
Figure 5.
Tissue-specific analysis of CsSPP and CsSPSs in tea plant.
Expression analysis of CsSPSs and CsSPP under various abiotic stress conditions
-
The spatial-temporal expression patterns of CsSPP and CsSPSs were analyzed under various abiotic stress conditions. As shown in Fig. 6, CsSPP and CsSPSs are differentially expressed under different stress conditions. Under salt treatment (ST) conditions, the expression of CsSPP was up-regulated nearly 3-fold after 1 d of ST, and then decreased when the treatment time continued. CsSPS4 was down-regulated within 2 d of ST. CsSPS2 was slightly up-regulated after 12 h and 48 h of ST, respectively, while down-regulated after 24 h of ST. Besides, CsSPS1/3/5 showed similar expression patterns under ST condition, which were highly induced within 12 h of ST, and then decreased when the treatment time continued. Under cold treatment (CT) condition, CsSPP and CsSPS1-3 showed similar expression patterns, all of them were down-regulated firstly within 12 h of CT, and then up-regulated by CT, of which CsSPS2/3 transcripts were respectively induced more than 3- and 2-fold after 48 h of CT as compared to 0 h. Besides, CsSPS4 was inhibited by CT within 2 d of CT, while CsSPS5 was continuously induced with the increased treatment time. Under drought treatment (DT) conditions, CsSPP and CsSPS2/4 showed similar expression patterns, all of them were constantly induced, and reached maximum expression levels after 24 h of DT, and then to some extent reduced. Similarly, CsSPS3/5 were constantly induced and reached maximum expression levels at 12 h. In addition, CsSPS1 was not significantly affected by DT. Briefly, CsSPP and CsSPSs participated in different stress responses of tea plants, but the time and degree of their effects were different.
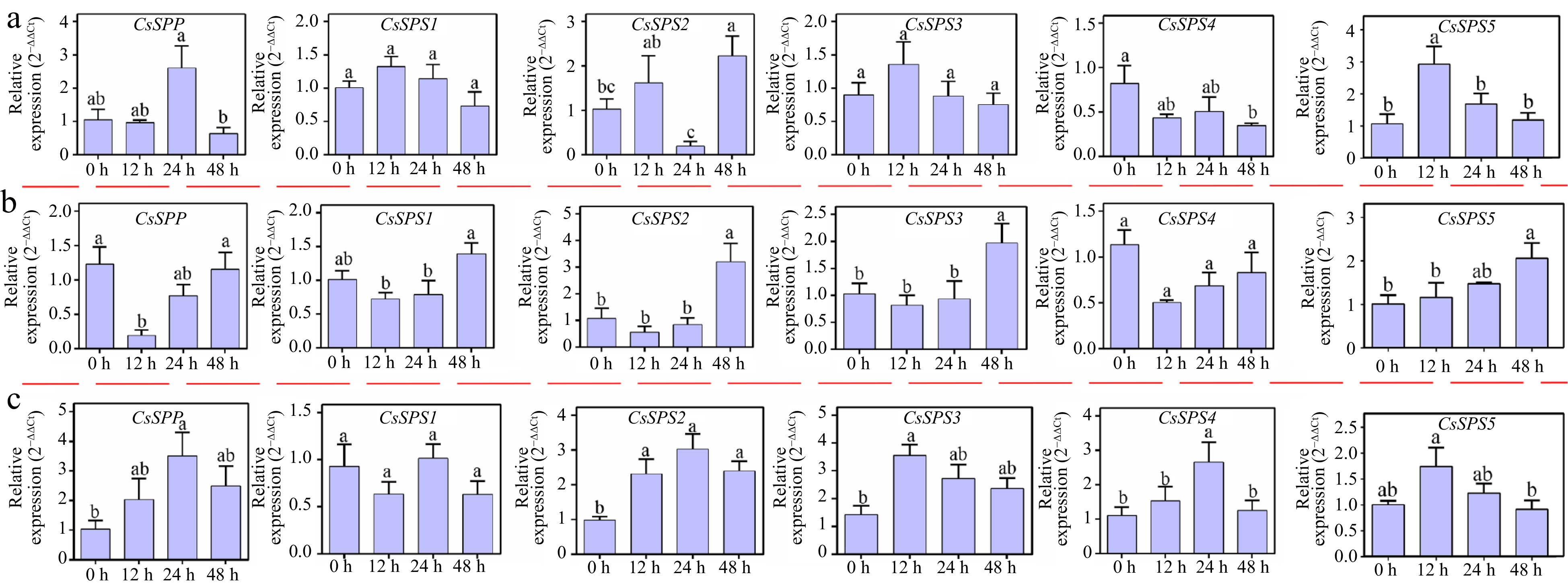
Figure 6.
Expression analysis of CsSPP and CsSPSs under different abiotic stress conditions. (a) Expression profiles of CsSPP and CsSPSs under salt stress conditions. (b) Expression profiles of CsSPP and CsSPSs under cold stress conditions. (c) Expression profiles of CsSPP and CsSPSs under drought stress conditions.
Expression analysis of CsSPSs and CsSPP in different tea plant cultivars under cold stress conditions
-
In this study, the cold tolerance of two tea plant cultivars, 'SCZ' and 'BY1', were compared with different physiological indexes. As shown in Fig. 7a, the Fv/Fm and NP values of 'SCZ' cultivar were higher than that of the 'BY1' cultivar, but the EL value and MDA content in 'SCZ' cultivar were lower than that of the 'BY1' cultivar (Fig. 7a), indicating that the cold resistance of the 'SCZ' cultivar was higher than that of the 'BY1' cultivar. Therefore, these two cultivars were further used to investigate the relationships among the expression of CsSPSs and CsSPP, SPS activity, soluble sugar content and cold stress. As shown in Fig. 7b, except for CsSPS1, all of them were up-regulated under 0 oC treatment for 5 d both in these two cultivars, and returned to normal levels after 3 d of recovery. Specifically, CsSPP and CsSPS2/4/5 transcripts were increased more than 2-fold after 5 d of CT, respectively. Besides, we found the expression level of CsSPS2 was significantly higher in the 'BY1' cultivar than that in the 'SCZ' cultivar, while the expression level of CsSPS5 was significantly higher in the 'SCZ' cultivar than that in the 'BY1' cultivar, indicating the different roles of CsSPSs in coping with cold stress in different tea plant cultivars. Moreover, we found the SPS activity was obviously increased in the 'SCZ' cultivar, but not significantly changed in the 'BY1' cultivar after 5 d of CT. After 3 d of recovery growth, the SPS activity was decreased in the 'SCZ' cultivar, while significantly increased in the 'BY1' cultivar. Furthermore, the contents of TSS, Suc, Glc and Fru were increased after 5 d of CT except for TSS in the 'BY1' cultivar, and then decreased to normal levels after 3 d of NT. Furthermore, the 'BY1' cultivar contained a relatively higher content of TSS, Suc and Fru under CT and recovery conditions. As shown in Fig. 7c, CsSPP and CsSPS2-5 were positively correlated with SPS activity and the contents of soluble sugar (TSS, Suc, Glc and Fru) in the 'SCZ' cultivar, especially CsSPS2/4/5, significantly correlated with Suc content, respectively. Besides, CsSPS3 was also significantly and positively correlated with Glc and Fru contents in the 'SCZ' cultivar, respectively. These results indicated that the high expressions levels of CsSPP and CsSPSs contributed to the accumulation of different types of soluble sugars in the 'SCZ' cultivar, thus improving the adaptability to low temperatures. Different from the 'SCZ' cultivar, CsSPS1 was positively correlated with SPS activity in the 'BY1' cultivar. Meanwhile, although CsSPS2/4/5 were positively correlated with Suc, Glc, and Fru, all of them were negatively correlated with SPS activity, which suggested that SPS activity in the 'BY1' cultivar may be regulated by post-transcriptional and post-translational regulation, as some potential phosphorylation sites were identified in the amino acid sequences of CsSPSs (Fig. 4c). Different from the 'BY1' cultivar, the higher cold resistance of the 'SCZ' cultivar may be due to the up-regulated expression of CsSPSs leading to the increase of SPS activity, which promotes the synthesis of soluble sugar content to improve cold tolerance.
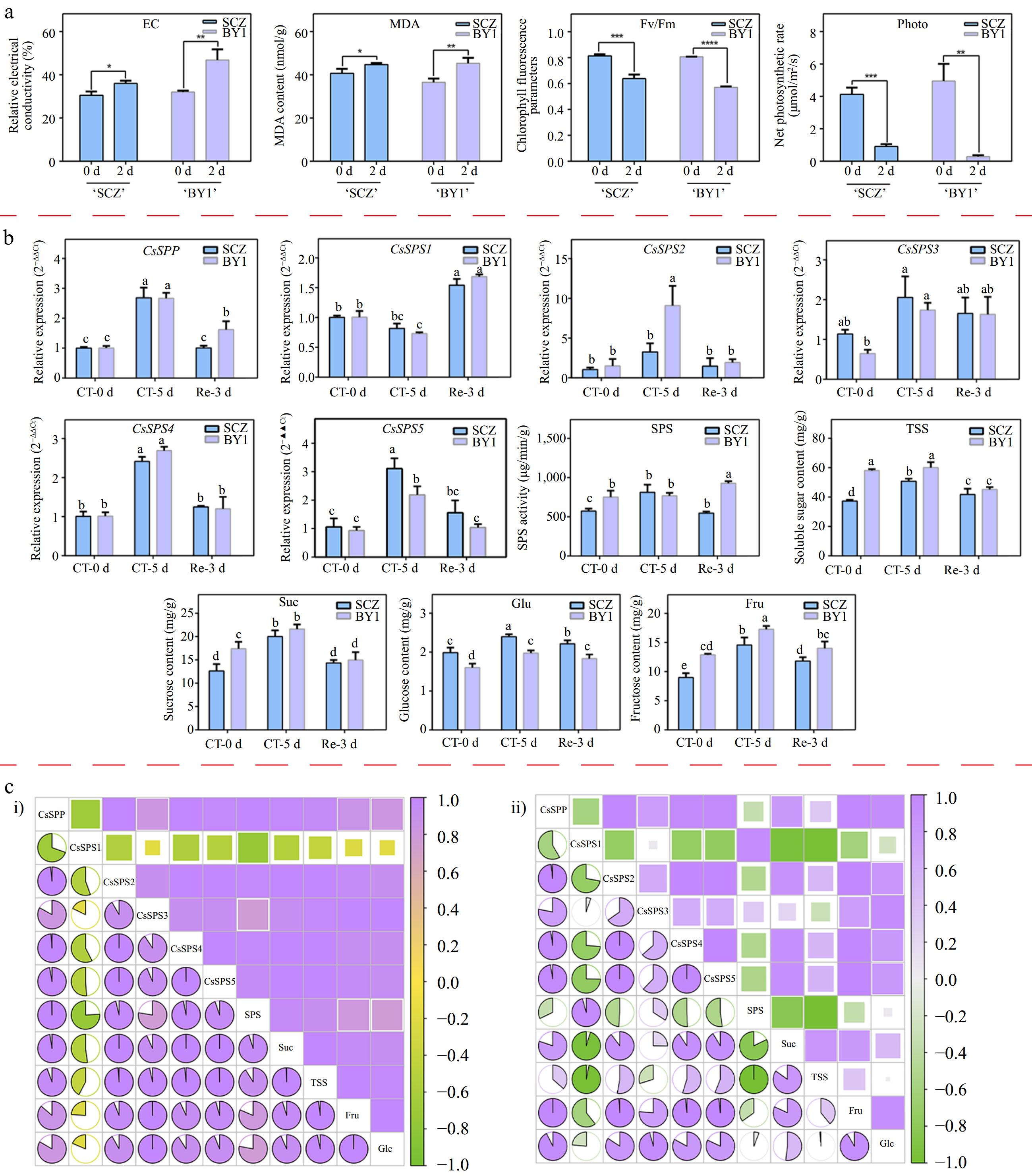
Figure 7.
Expression analysis of CsSPP and CsSPSs in different tea plant cultivars under cold stress conditions. (a) Relative electrolytic leakage, malondialdehyde and photosynthetic parameters of 'Shuchazao' and 'Baiye1' under cold stress conditions. (b) Expression levels of CsSPP and CsSPSs, SPS activity and different types of soluble sugar content. (c) Correlation analysis of CsSPP and CsSPSs, SPS activity and different types of soluble sugar components in 'Shuchazao' and 'Baiye1' cultivars, respectively. (c-i) Correlation analysis of CsSPP, CsSPSs, SPS activity and soluble sugars in the 'Shuchazao' cultivar. (c-ii) Correlation analysis of CsSPP, CsSPSs, SPS activity and soluble sugars in the 'Baiye1' cultivar. Green color means negative correlation, purple color means positive correlation.
-
As the main product of photosynthesis, Suc plays an important role in plant growth and development, yield and quality formation, and stress responses. Previous studies found that Suc biosynthesis is mainly regulated by SPS and SPP[56]. In recent years, the protein structures and the roles of SPSs and SPPs have been explored in many plant species. Among them, SPSs mainly contain a D-fructose 6-phosphate (F6P)-binding domain, nucleotide diphosphate glucose (NDPGlc)-binding domain, and a SPP-related C-terminal domain[57]. Besides, SPS activity was regulated by multisite protein phosphorylation[6,58]. For example, Ser-158 may be mainly responsible for light/dark modulation, Ser-229 may be a binding site for 14-3-3 inhibitory proteins, and Ser-424 is thought to be responsible for osmotic stress activation of SPS[59,60]. In this study, five CsSPSs were identified from three tea plant genomes, among which CsSPS1 was a tandem duplication product of CsSPS2 in the 'SCZ' genome. All of these CsSPSs contain the conserved F6P-binding domain, NDPGlc-binding domain, and the SPP-related C-terminal domain (Fig. 4c), indicating that these CsSPSs possess the Suc biosynthesis ability. Meanwhile, some potential phosphorylation sites, such as Ser191 and Ser385 in CsSPS1/2, Ser146, Ser220 and Ser409 in CsSPS3, Ser148 and Ser217 in CsSPS4, and Ser221 and Ser416 in CsSPS5, were also identified (Fig. 4c), suggesting that SPS activity was also influenced by light, osmotic stress, and also 14-3-3 inhibitory proteins in tea plants.
In addition to SPS, there are fewer SPPs than SPSs in most plant species, and some SPP tandem duplication events occurred in plants[61]. Previous study found that SPP protein structure mainly consists of S6PP (PF05116) and S6PP_C (PF08472) domain, and SPP depends on Mg2+ to specifically dephosphorylate S6P to produce Suc[62]. Besides, SPP formed homodimer in some vascular plants, and the molecular weight of the SPP monomer is usually 50 KDa, while the molecular weight of the homodimer is about 120 KDa[18,62,63]. Moreover, since SPS contains the SPP-related C-terminal domain, SPP and SPS could interact to improve the efficiency of Suc synthesis[64]. Maloney et al.[65] found that SPS could directly interact with SPP to affect the soluble carbohydrate pool and the allocation of carbon to starch. Meanwhile, co-overexpression of AtSPS-AtSPP and AtSPP-AtSPS chimera increased the content of soluble carbohydrates and also promoted the growth rates both in Arabidopsis and hybrid poplar, respectively[19]. In the present study, a unique and conserved CsSPP with 48.11 KDa MW was identified from three tea plant genomes. Meanwhile, both the conserved S6PP domain and S6PP_C domain were contained in CsSPP, suggesting that CsSPP participates in Suc biosynthesis of tea plants, and this process is highly conserved in different tea plant cultivars. In addition, CsSPP may also interact directly with CsSPSs to participate in Suc accumulation of tea plants due to the conserved SPP-related C-terminal domain observed in CsSPSs. However, this hypothesis will be further certificated in the future.
CsSPSs and CsSPP mediate vegetative and reproductive progress of tea plants
-
Previous studies stated that SPS and SPP widely participated in the flowering, plant growth, seed germination and pollen activity in plants[66−68]. In tobacco, inhibition of NtSPP expression of tobacco significantly reduced the SPP activity, Suc and Hex contents, but dramatically increased starch content, and thus reducing the photosynthesis, chlorosis and growth rate of transgenic plants[21]. In this study, we found that the expressions of CsSPSs and CsSPP could be detected in all tissues, indicating that Suc is inseparable from all stages of vegetative and reproductive growth of tea plants. However, the expression levels of CsSPP and CsSPSs were lowest in roots, which indicated that Suc may be mainly synthesized from the source tissue of the above-ground part and then transported to the underground part. Besides, all CsSPSs showed the highest transcriptions in flower, which suggested that Suc plays an important role in floral nectar production of flower. However, our previous study found that Suc content of flowers was not the highest compared with other tissues, instead, the contents of TSS, Glc and Fru were highest in flowers[69]. Meanwhile, the vacuole INV activity and the transcription abundance of a vacuole INV gene (CsINV5), were highest in flower[24]. Here, we further performed the correlation analysis among the expressions of Suc-related genes (CsSPP, CsSPSs, and CsINV5), VIN activity and the contents of soluble sugars (TSS, Suc, Glc, and Fru) (Supplemental Fig. S3), where we found each component was positively correlated with each other except CsSPP, which speculated that the SPP/SPS-VIN module mediated the Suc metabolism in the flower of tea plants. In detail, after the biosynthesis of Suc by SPS and SPP in the flower of tea plants, part of the Suc needs to be further hydrolyzed by INV to form two monosaccharides, Glc and Fru, and then participates in floral nectar production, pollination, fertilization and fruit formation. In addition to mediating reproductive growth, previous studies reported that Suc accumulation might be regarded as one of the key indicators of leaf senescence[70,71]. A similar phenomenon was also found in our previous study, where we found Suc content was highest in senescent leaf compared with the other tissues[69], suggesting a great role of Suc in the aging process of tea plant leaves. Here, we further found the transcription abundance of CsSPS2 was higher in senescent leaf than the other tissues except for flower, indicating that CsSPS2 may be an important regulator of leaf senescence through mediating Suc accumulation in tea plants.
CsSPSs and CsSPP involved in various abiotic stress and hormone responses
-
Many studies found that multiple carbohydrate metabolism-related genes involved in carbohydrate biosynthesis, hydrolysis, and transport were differentially expressed under stress conditions[67]. Besides, there is increasing evidence that Suc metabolism is one of the key regulatory systems that confer stress tolerance in plants[72,73]. Changes in the activity of SPS and SPP significantly influenced Suc accumulation, thus affecting plant growth and development and stress tolerance[74−76]. In Arabidopsis, mutation of SPSA2 did not affect the seeds and plants of the mutant, but reduced the drought tolerance of the spsa2 mutant through the regulation of proline content, sugar accumulation and antioxidant response[77]. Under normal water condition, the reduction of SPS activity by 70%−80% resulted in a corresponding reduction of Suc synthesis by 30%−50%, while under water deficit condition, the reduction of SPS activity prevented dry-matter allocation to tubers, indicating that SPS is essential for adaptive changes in tuber metabolism and whole plant allocation process[74]. In terms of tea plant, previous studies revealed that carbohydrate metabolism plays an important role in cold[78,79], drought[80], and salt[81] stress responses. In particular, many sugar-related genes involved in cold acclimation of tea plant have been identified by Yue et al.[82]. Among them, CsINVs (e.g., CsINV2/5/10)[24,25], CsSWEETs (e.g., CsSWEET1a/16/17)[27,83], and CsHXKs (e.g., CsHXK3/4)[26] have been further demonstrated to be involved in the cold response of tea plants through Suc hydrolysis, sugar transport and sugar signaling, respectively. In this study, we further found the expressions of CsSPP and CsSPSs were induced by cold, drought and salt stresses at different treatment time points, respectively, indicating that SPP and SPS positively modulate abiotic stress responses of tea plants. Besides, CsSPP and CsSPSs differentially expressed in two tea plant cultivars with different cold tolerance under cold stress condition. Interestingly, we found SPS activity was higher in the 'BY1' cultivar under normal temperature conditions, while slightly lower under low temperature conditions compared with the 'SCZ' cultivar. Meanwhile, higher soluble sugar content except Glc found in 'BY1' cultivar both under normal and low temperature conditions. However, from these results, we found that the SPS activity and soluble sugar content of the 'SCZ' cultivar increased significantly more than that of the 'BY1' cultivar, indicating that the Suc synthesis ability of the 'SCZ' cultivar through photosynthesis was higher than that of the 'BY1' cultivar, and the low temperature adaptability of the 'SCZ' cultivar was higher than that of the 'BY1' cultivar. Based on the above studies, it can be seen that the increase of SPP and SPS activities in tea plant under stress conditions can promote Suc synthesis. Subsequently, partial Suc is transported to vacuoles, cell walls and other sub-organelles by sugar transporters, and is hydrolyzed by INV to form monosaccharides. Monosaccharides can be further phosphorylated by HXK to participate in the synthesis of other substances (e.g., inositol, trehalose and mannitol); finally, these synthetic Suc and monosaccharides participate in the stress responses of tea plants with carbon sources, osmoprotectants, reactive oxygen scavengers and sugar signaling molecules.
Although SPSs and SPPs are known to be involved in various stress responses, their transcriptional and post-translational regulation mechanisms have been poorly studied. As is mentioned above, the phosphorylation levels of SPSs were affected by different environmental factors, among which Ser158, Ser229 and Ser424 were three important phosphorylation sites[59,60,84]. Besides, previous study found that SPS was also phosphorylated by calcium-dependent protein kinase (CDPK) via calcium signaling pathway. In detail, the encoding protein of a cold-reduced gene, OsCPK17, could directly phosphorylate OsSPS4 in rice. Under low temperature conditions, the phosphorylation level of Ser170 residue in OsSPS4 was higher in wild type plant than that in oscpk17 mutant, indicating that the reduction of OsSPS4 activity possibly regulated by OsCPK17 through directly phosphorylating OsSPS4 during the early stages of cold stress[85]. In tea plant, a previous study found that CsCDPKs play important roles in coping with various stresses, among which CsCPK4/5/9/30 may be the main cold regulators of tea plants[86]. Combined with the expression profiles and the conserved Ser residues of CsSPSs, we speculated that SPS activity may also be regulated by CDPK phosphorylation, and participate in stress response through the calcium signaling pathway. On the other hand, there are few studies on the transcriptional regulation of SPSs and SPPs. Recently, seven GmSPSs genes were identified from Glycine max, and all of them were up-regulated by cold stress in soybean leaves, especially GmSPS8 and GmSPS18. Promoter analysis found that many potential inducers of CBF expression 1 (ICE1) binding sites were predicted in the promoter regions of GmSPSs. Electrophoretic mobility shift assay (EMSA) further proved that GmICE1 could regulate the transcription abundances of GmSPS8 and GmSPS18 in N. benthamiana[87]. In the present study, in addition to stress-related cis-acting elements, many hormone-related cis-acting elements, such as ABRE, GA, MeJA, and TGA, were contained in the promoter regions of CsSPSs and CsSPP, suggesting that CsSPPs and CsSPP participate in development and stress response of tea plant via hormone-signaling pathway. In particular, the promoter region of CsSPS4 contains three MeJA, two ABRE, one GA, and one TGA, which indicated that CsSPS4 may be a central component in the cascade of sugar signaling and hormone signaling in tea plants. Moreover, co-expression analysis results found that the expression profiles of CsMYB35 (CSS0014516) and CsGLOBOSA-like (CSS0022940) were positively correlated with CsSPS3, suggesting that CsMYB35 and CsGLOBOSA-like may be two candidate regulators of CsSPS3. However, the function and the regulation mechanism of these two transcription factors need to be further explored in the future.
-
In this study, a unique CsSPP and five CsSPSs genes were identified from three tea plant genomes. Bioinformatic analysis results showed that CsSPP and CsSPSs were highly conserved in different tea plant cultivars respectively, and all of them can participate in Suc biosynthesis in the cytoplasm of tea plants. Tissue-specific analysis found that CsSPP and CsSPSs are necessary for vegetative and reproductive growth of tea plants, especially for the floral nectar production of flower. In addition, CsSPP and CsSPSs were differentially expressed under various abiotic stress conditions, among which CsSPS2/3/5 were induced by cold, drought and salt stress treatments at different treatment time points. Under cold stress conditions, the SPS activity and the soluble sugar contents of the 'SCZ' cultivar increased more than that of the 'BY1' cultivar, indicating that the 'SCZ' cultivar owns higher photosynthetic capacity and Suc synthesis ability under cold stress conditions. This study will provide theoretical foundation for further exploring the function of SPP and SPS involved in abiotic stress responses of tea plants.
-
The authors confirm contributions to the paper as follows: study conception and design: Qian W, Ikka T; material preparation and data collection: Liang S, Lang X, Yue J, He S; data analysis: Liang S, Zhang S, Wang H, Fan K; draft manuscript preparation: Liang S, Qian W, Wang H; review and editing: Wang Y, Ding Z, Yamashita H, Ikka T; Partial funds and consultation: Qian W, Wang Y. All authors read and approved the final manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
This research was supported by the National Natural Science Foundation of China (32272767, 31800588), the Shandong Agricultural Elite Variety Project (2020LZGC010), and 'Provincial and School Joint Training Program' for Government-sent Overseas Visiting Scholars of Shandong Province in 2020.
-
The authors declare that they have no conflict of interest.
- Supplemental Table S1 qRT-PCR primers.
- Supplemental Fig. S1 Amino acid sequences alignment analysis of CsSPP and CsSPSs.
- Supplemental Fig. S2 Chromosomal location of CsSPP and CsSPSs in 'Tieguanyin' and 'Huandan' genomes, respectively.
- Supplemental Fig. S3 Correlation analysis of CsSPP, CsSPSs, CsINV5 and soluble sugar contents in different tea plant tissues.
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Liang S, Wang H, Yamashita H, Zhang S, Lang X, et al. 2024. Genome-wide identification and expression analysis of sucrose phosphate synthase and sucrose-6-phosphate phosphatase family genes in Camellia sinensis. Beverage Plant Research 4: e015 doi: 10.48130/bpr-0024-0007 

Genome-wide identification and expression analysis of sucrose phosphate synthase and sucrose-6-phosphate phosphatase family genes in Camellia sinensis
- Received: 25 December 2023
- Revised: 13 January 2024
- Accepted: 23 January 2024
- Published online: 07 May 2024
Abstract: Sucrose phosphate synthetase (SPS, EC 2.4.1.14) and sucrose phosphate phosphatase (SPP, EC 3.1.3.24) are two key enzymes for sucrose biosynthesis, which play essential roles in growth, development and stress responses of plants. However, the roles of SPS and SPP have not been illustrated well in tea plants until now. In this study, a unique CsSPP and five CsSPSs (CsSPS1-5) were identified from the tea plant genome. Bioinformatic analysis results found that CsSPP and CsSPSs were clustered together with the known SPPs and SPSs of other species, respectively, and their amino acid sequences contain the conserved domains required for sucrose biosynthesis. Tissue-specific analysis results showed that CsSPP and CsSPSs were widely involved in vegetative and reproductive growth of tea plant, among which the transcription levels of CsSPP was highest in immature stem, while CsSPSs were highest in flower. Spatio-temporal expression analysis results showed that all of these genes are involved in abiotic stress responses of tea plants. Meanwhile, SPS activity and the contents of sucrose, glucose, fructose, and total soluble sugar in 'Shuchazao' cultivar increased more than that of 'Baiye1' under low temperature conditions. Correlation analysis results showed that the expression profiles of CsSPS2/4/5 were significantly and positively correlated with sucrose content in 'Shuchazao' cultivars under low temperature conditions, suggesting the significant roles of these genes in sucrose accumulation. In conclusion, this study will provide a theoretical basis for further functional research of SPS and SPP in plants.
-
Key words:
- Camellia sinensis /
- SPP /
- SPS /
- Tissue-specific analysis /
- Spatio-temporal analysis