









-
Wilms tumor (WT), or nephroblastoma, is the leading renal malignancy in children, composing roughly 5%–6% of all childhood cancers[1]. With an incidence of 4.3 per million annually, WT is less prevalent in East Asia than the incidence of 8–9 per million seen in North America or Europe[2]. Its incidence peaks at the age of three years[3]. The incidence is about 3.2 cases per million annually in China[4]. This tumor results from failure of early nephrogenesis. Wilms tumors generally have a relatively favorable prognosis after multimodal treatment, with a 5-year survival rate exceeding 90%. However, a subset of high-risk patients frequently encounters treatment resistance and eventual mortality[5]. Moreover, ~20% of patients will face relapse, with the majority occurring within two years postdiagnosis[6]. Early diagnosis remains a challenge, especially in patients with asymptomatic tumors[1].
Environmental factors could play a role in the etiology of Wilms tumor, yet not necessarily, including prenatal exposure to pesticides, maternal smoking during pregnancy, and preterm birth[1]. In contrast, genetic factors play a more substantial role in predisposing individuals to Wilms tumor. Approximately 10% of Wilms tumors are related to known WT predisposition congenital syndromes, such as genitourinary anomalies and mental retardation (WAGR), alongside Beckwith–Wiedemann syndrome, and Denys–Drash syndrome[2]. In addition, candidate gene association studies and genome-wide association studies (GWASs) have revealed many Wilms tumor predisposition variants[7], such as single nucleotide polymorphisms (SNPs) in the survivin, MTHFR, BARD1 METTL14, and WTAP genes, as well as 2p24 (rs375513 and rs80762) and 11q14 (rs790356)[7−12], yet much of the genetic landscape remains unknown. Understanding these genetic variations is critical for improving preventive and treatment strategies.
tRNA methyltransferase 10 homolog C (TRMT10C), referred to as mitochondrial ribonuclease p protein 1 (MRPP1), is a gene that encodes an essential enzyme participating in the processing and maturation of mitochondrial tRNA (mt-tRNA)[13,14]. Moreover, TRMT10C belongs to the methyltransferase family and is responsible for the maturation and stability of mt-tRNA by catalyzing the N1-methylation at the 9th adenosine position (m1A)[15,16]. This methyltransferase activity is crucial for mitochondrial function and energy production because it maintains proper mt-tRNA folding for the synthesis of mitochondrial proteins, which are components of oxidative phosphorylation complexes[17,18]. Given the importance of mitochondrial function in cellular homeostasis, TRMT10C may play a significant role in tumorigenesis, where mitochondrial dysfunction is a hallmark of tumors[19,20]. Dysregulation of TRMT10C contributes to several cancers, including ovarian and cervical cancer, hepatocellular carcinoma[21], and lung cancer[22,23]. Wilms tumors often exhibit alterations in metabolic pathways[24,25]. However, the impact of TRMT10C on Wilms tumor risk needs to be clarified. This study aimed to investigate the association of Wilms tumor susceptibility with potentially functional TRMT10C polymorphisms that may affect gene function and expression abundance.
-
We collected genotyping information from 398 Wilms tumor cases and 1,198 frequency-matched controls among the 414 Wilms tumor cases and 1,199 controls. The control group's genotype distributions followed HWE (p > 0.05 for all four polymorphisms), supporting the accuracy of the genotyping for further statistical evaluation. In the overall analysis, none of the four SNPs demonstrated a statistically significant association with Wilms tumor susceptibility in any of the genetic models (Table 1). We observed no significant risk associated with the rs7641261 C > T, rs2303476 T > C, rs4257518 A > G, or rs3762735 C > G polymorphism in the Wilms tumor population compared with controls. Moreover, based on the observed genotype frequencies, we defined rs7641261 TT, rs2303476 CC, rs4257518 GG, and rs3762735 GG as high-risk genotypes. However, when we grouped the children based on the presence of genotypes 1–4 of these high-risk genotypes, there was no significant increase in Wilms tumor susceptibility compared with controls lacking any of these genotypes (OR = 1.14, 95%; CI = 0.88–1.48; p = 0.324). These results suggest that these SNPs and their combinations do not significantly contribute to Wilms tumor risk in the general population.
Table 1. Association between TRMT10C gene polymorphisms and Wilms tumor risk.
Genotype Cases (N = 398) Controls (N = 1198) Pa Crude OR (95% CI) P Adjusted OR (95% CI)b Pb rs7641261 C > T
(HWE = 0.425)CC 225 (56.53) 674 (56.26) 1.00 1.00 CT 143 (35.93) 456 (38.06) 0.94 (0.74−1.20) 0.611 0.94 (0.74−1.19) 0.604 TT 30 (7.54) 68 (5.68) 1.32 (0.84−2.08) 0.230 1.31 (0.83−2.07) 0.246 Additive 0. 653 1.04 (0.87−1.26) 0.653 1.04 (0.87−1.25) 0.676 Dominant 173 (43.47) 524 (43.74) 0.924 0.99 (0.79−1.24) 0.924 0.99 (0.79−1.24) 0.908 Recessive 368 (92.46) 1130 (94.32) 0.180 1.36 (0.87−2.12) 0.182 1.34 (0.86−2.10) 0.195 rs2303476 T > C
(HWE = 0.797)TT 245 (61.56) 740 (61.77) 1.00 1.00 TC 134 (33.67) 405 (33.81) 1.00 (0.78−1.27) 0.996 0.99 (0.78−1.27) 0.964 CC 19 (4.77) 53 (4.42) 1.08 (0.63−1.87) 0.774 1.09 (0.63−1.88) 0.756 Additive 0.867 1.02 (0.84−1.24) 0.867 1.02 (0.84−1.24) 0.878 Dominant 153 (38.44) 458 (38.23) 0.940 1.01 (0.80−1.27) 0.940 1.01 (0.80−1.27) 0.964 Recessive 379 (95.23) 1145 (95.58) 0.771 1.08 (0.63−1.85) 0.771 1.09 (0.64−1.87) 0.748 rs4257518 A > G
(HWE = 0.416)AA 121 (30.40) 364 (30.38) 1.00 1.00 AG 186 (46.73) 580 (48.41) 0.97 (0.74−1.26) 0.790 0.97 (0.74−1.26) 0.806 GG 91 (22.86) 254 (21.20) 1.08 (0.79−1.48) 0.642 1.07 (0.78−1.47) 0.667 Additive 0.691 1.03 (0.88−1.21) 0.691 1.03 (0.88−1.21) 0.714 Dominant 277 (69.60) 834 (69.62) 0.995 1.00 (0.78−1.28) 0.995 1.00 (0.78−1.28) 0.996 Recessive 307 (77.14) 944 (78.80) 0.485 1.10 (0.83−1.45) 0.485 1.09 (0.83−1.44) 0.519 rs3762735 C > G
(HWE = 0.489)CC 304 (76.38) 903 (75.38) 1.00 1.00 CG 85 (21.36) 271 (22.62) 0.93 (0.71−1.23) 0.616 0.93 (0.70−1.23) 0.602 GG 9 (2.26) 24 (2.00) 1.11 (0.51−2.42) 0.786 1.11 (0.51−2.41) 0.800 Additive 0.790 0.97 (0.77−1.23) 0.791 0.97 (0.76−1.22) 0.771 Dominant 94 (23.62) 295 (24.62) 0.685 0.95 (0.73−1.24) 0.685 0.94 (0.72−1.23) 0.669 Recessive 389 (97.74) 1174 (98.00) 0.754 1.13 (0.52−2.46) 0.754 1.12 (0.52−2.44) 0.767 Risk genotypesc 0 295 (74.12) 919 (76.71) 1.00 1.00 1−4 103 (25.88) 279 (23.29) 0.294 1.15 (0.89−1.49) 0.294 1.14 (0.88−1.48) 0.324 OR, odds ratio; CI, confidence interval; HWE, Hardy-Weinberg equilibrium. a χ2 test for genotype distributions between Wilms tumor patients and cancer-free controls. b Adjusted for age and sex. c Risk genotypes were rs7641261 TT, rs2303476 CC, rs4257518 GG, and rs3762735 GG. Stratified analysis
-
To explore potential subgroup-specific effects, we performed stratified analyses regarding age, sex, and clinical stage, which were crucial for uncovering associations that may not be apparent in the overall population owing to the effects of confounding variables on disease susceptibility (Table 2). Interestingly, in males, the TRMT10C rs7641261 TT genotype emerged as a significant risk factor. Compared with those with the CC or CT genotype, those with the TT genotype presented a more than twofold enhanced Wilms tumor predisposition (OR = 2.15, 95%; CI = 1.20–3.86; p = 0.011). This association was not observed in females, suggesting a potential sex-specific effect of rs7641261 on Wilms tumor susceptibility. Furthermore, the rs7641261 TT genotype was shown to elevate the risk of developing early-stage Wilms tumor (clinical stages I and I + II) but not advanced-stage Wilms tumor (III, IV, or III + IV) (OR = 1.81, 95% CI = 1.12–2.93; p = 0.016). Our findings indicate that the rs7641261 TT may primarily contribute to the initiation of Wilms tumor rather than its progression. Similarly, we observed that carrying 1–4 high-risk genotypes predisposed early-stage Wilms tumor, particularly in stages I and I + II, but a decreased risk of advanced-stage tumors (III or III + IV). These findings suggest that the accumulation of high-risk genotypes may drive the early stages of tumorigenesis but does not affect tumor progression to more advanced stages. The lack of association between high-risk genotypes and advanced-stage Wilms tumors may be explained by the hypothesis that additional molecular events, beyond the presence of germline polymorphisms, are necessary for tumor progression. These findings highlight the importance of understanding how specific genetic variants contribute to distinct phases of tumor development.
Table 2. Stratification analysis of TRMT10C protective genotypes and Wilms tumor risk.
Variables rs7641261 (cases/controls) AOR (95% CI)a Pa Risk genotypes (cases/controls) AOR (95% CI)a Pa CC/CT TT 0 1−4 Age (month) ≤ 18 127/438 9/27 1.18 (0.54−2.58) 0.679 103/363 33/102 1.17 (0.74−1.83) 0.503 > 18 241/692 21/41 1.47 (0.85−2.54) 0.169 192/556 70/170 1.13 (0.82−1.56) 0.465 Sex Females 176/484 10/37 0.75 (0.36−1.53) 0.423 136/386 50/135 1.05 (0.72−1.54) 0.788 Males 192/646 20/31 2.15 (1.20−3.86) 0.011 159/533 53/144 1.24 (0.86−1.77) 0.253 Clinical stages I 126/1130 11/68 1.39 (0.72−2.71) 0.331 93/919 44/279 1.53 (1.04−2.25) 0.030 II 97/1130 14/68 2.35 (1.27−4.34) 0.006 76/919 35/279 1.50 (0.98−2.29) 0.059 III 88/1130 3/68 0.58 (0.18−1.89) 0.366 79/919 12/279 0.51 (0.27−0.95) 0.033 IV 41/1130 2/68 0.80 (0.19−3.38) 0.759 35/919 8/279 0.75 (0.34−1.63) 0.462 I + II 223/1130 25/68 1.81 (1.12−2.93) 0.016 169/919 79/279 1.52 (1.13−2.05) 0.006 III + IV 129/1130 5/68 0.65 (0.26−1.65) 0.364 114/919 20/279 0.58 (0.35−0.95) 0.031 AOR, adjusted odds ratio; CI, confidence interval. a Adjusted for age and sex, omitting the corresponding stratification factor. Values were in bold if the p values less than 0.05 or the 95% CI excluding 1.00. eQTL analysis
-
To explore the functions of the rs7641261 polymorphism, we performed an eQTL analysis (Fig. 1). The analysis revealed that rs7641261 was significantly correlated to the level of SENP7, a neighboring gene, in several tissue types, including cultured fibroblasts, EBV-transformed lymphocytes, and whole blood. Specifically, cells harboring the rs7641261 TT genotype presented significantly reduced SENP7 expression compared with those harboring the CC or CT genotypes (p < 0.01). SENP7 encodes a SUMO-specific protease that regulates protein sumoylation, a critical posttranslational modification that modulates protein stability, localization, and activity.
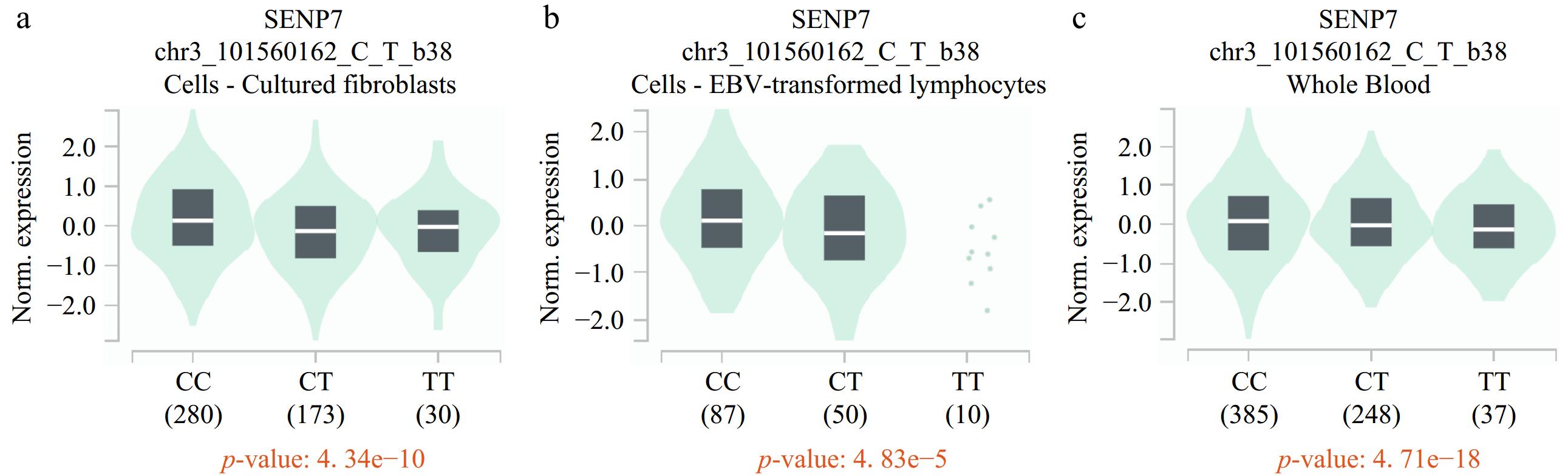
Figure 1.
eQTL analysis of rs7641261 in the GTEx database revealed a negative correlation with SENP expression. The rs7641261 variant shows significant eQTL effects, where the presence of the T allele is associated with (a) reduced SENP expression levels in cultured fibroblasts, (b) EBV-transformed lymphocytes, and (c) whole blood.
-
In childhood cancers, somatic mutations are relatively rare, whereas germline gene mutations dominantly predispose individuals to cancer.[26] Wilms tumors likely have a median somatic mutation rate of 0.17 per million bases (Mb), which is markedly lower than the 1–10 mutations per Mb observed in adult papillary and clear cell renal cell carcinomas[27,28]. In this context, common, low-penetrance genetic variants such as SNPs and their contributions to Wilms tumor risk have attracted attention, as SNPs can affect gene function and expression, leading to variations in tumor predisposition. Recent research has highlighted the role of genes involved in developmental pathways, tumor suppressors, and DNA repair mechanisms in the pathogenesis of Wilms tumor[3]. However, the genetic basis of Wilms tumor remains incompletely understood, and further investigation is needed to elucidate the specific genetic alterations that drive tumorigenesis.
In this study, the overall analysis unveiled no significant associations between TRMT10C variants and susceptibility to Wilms tumor. However, stratified analyses identified the rs7641261 TT as a risk factor in boys and early-stage Wilms tumor patients. The eQTL findings further support a biological role for rs7641261 in modulating Wilms tumor susceptibility through its influence on SENP7 expression. Consistent with the current results, we demonstrated that TRMT10C SNPs significantly enhanced hepatoblastoma in our previous research.[29]. These findings show that TRMT10C polymorphisms may confer susceptibility to different pediatric tumors. However, we identified significant associations between all four TRMT10C SNPs (rs7641261, rs2303476, rs4257518, and rs3762735) and increased hepatoblastoma risk[29]. In this Wilms tumor study, only one of these four SNPs (specifically rs7641261) was significantly associated with cancer risk, and this effect was specific to male patients. This discrepancy in findings between hepatoblastoma and Wilms tumors highlights the complexity and potential tissue-specific roles of TRMT10C in oncogenesis. Pediatric liver and kidney tumors may involve different genetic and epigenetic regulatory pathways that influence how TRMT10C polymorphisms affect tumor risk. Wilms tumors are among the few pediatric cancers with a higher incidence in girls (~10%) than in boys[30]. Additionally, the sex-specific association observed in Wilms tumors suggests that hormonal or sex-linked genetic factors may interact with TRMT10C polymorphisms, particularly influencing male susceptibility.
The tumor stage represents the severity of disease progression and may impact genetic expression, clinical outcomes, and possibly the genetic predisposition of the tumor. Wilms tumors are diagnosed at different stages, which could inadvertently affect genetic associations if not controlled. For example, advanced stages may involve genetic mutations not present in earlier stages, introducing potential bias in the analysis. Stratified analysis by tumor stage revealed that the rs7641261 TT genotype was a risk factor for early-stage Wilms tumor (stages I and I + II), but this effect did not extend to more advanced stages, indicating that the rs7641261 TT may drive Wilms tumor initiation. Moreover, carrying 1–4 high-risk genotypes conferred an enhanced risk of early-stage (stages I and I + II) Wilms tumors but a decreased risk of advanced-stage tumors (III or III + IV). These findings suggest that the accumulation of high-risk genotypes may drive the early stages of tumorigenesis. The protective association between high-risk genotypes and advanced-stage Wilms tumors may result from complex genetic alterations in late-stage diseases. Possession of 1–4 high-risk genotypes significantly elevated the risk for early-stage Wilms tumor, yet this risk diminished for advanced stages. These findings indicate that these genotypes may be involved in early tumor development, whereas the protective relationship in advanced stages might be attributed to intricate genetic changes as the tumor progresses.
Recent studies have revealed correlations between abnormal TRMT10C expression and the progression of several cancers[21−23], although its exact role in renal tumors, including Wilms tumors, remains underexplored. Disruption of TRMT10C activity may impair mitochondrial function, resulting in various pathophysiological conditions, particularly those linked to energy metabolism. Currently, research on TRMT10C is limited, and its pathogenic mechanisms are poorly understood. Several lines of evidence suggest that aberrant expression of TRMT10C is related to mitochondrial disorders. First, TRMT10C facilitates the conversion of mitochondrial polycistronic transcripts to mRNAs, rRNAs, and tRNAs as part of the MRPP1/2/3 complex[13]. Second, the MRPP1/MRPP2 complex acts as a pivotal site for tRNA maturation within human mitochondria[13]. Recessive TRMT10C gene mutations disrupt mitochondrial RNA synthesis and maturation and lead to several respiratory chain impairments and lactic acidosis[14]. Third, TRMT10C introduces m1A and/or m1G at position 9 in mt-tRNA in collaboration with SDR5C1, promoting mt-tRNA biogenesis[15]. In Alzheimer's disease, the overexpression of the TRMT10C protein enhances m1A methylation of ND5 mRNA, a key component of complex I of the respiratory chain, suppressing its translation and resulting in mitochondrial dysfunction[31]. TRMT10C dysregulation may contribute to the metabolic reprogramming of cancer cells, promoting their survival and proliferation. Zhao et al. reported that circFAM126A m7G modification catalyzed by TRMT10C effectively suppresses lung cancer cell proliferation by modulating key glycolytic pathways, suggesting that TRMT10C affects mitochondrial disorders in tumors[22].
There are several possible limitations to this study. First, although the study included multiple centers, the sample size is moderate, which could still limit the statistical power, particularly in subgroup analyses, and increase the possibility of false positives in sex- and stage-specific analyses. Second, our study focused exclusively on TRMT10C polymorphisms, whereas other genetic, environmental, or epigenetic factors may also contribute to Wilms tumor predisposition. Thus, this study may not capture the broader complexity of genetic predisposition for Wilms tumors. Third, as a retrospective case‒control study, it may be prone to selection and recall biases, potentially affecting the accuracy of the relationships drawn between TRMT10C SNPs and the risk of this pediatric tumor. Fourth, functional validation is crucial to confirm the regulatory role of rs7641261 on SENP7 expression and its potential involvement in Wilms tumor pathogenesis. Finally, these SNPs' biological relevance remains to be explored. Our results should be explained cautiously prior to rigorous validation.
-
In conclusion, this multiple-center study provides evidence that specific TRMT10C polymorphisms conferred enhanced predisposition to Wilms tumor, particularly in certain subpopulations. These findings indicate that TRMT10C may be associated with genetic susceptibility to Wilms tumor. However, extra studies are essential to validate these associations in larger, diverse populations and to investigate the underlying biological mechanisms by which TRMT10C influences Wilms tumor risk.
-
This research included 414 children who were diagnosed with Wilms tumor with pathological confirmation (Supplementary Table S1)[32]. Samples were collected before patients underwent any medical interventions. The participants were enrolled from five independent hospitals located in different regions of China, namely, Guangzhou (southern region), Zhengzhou (central region), Wenzhou (eastern region), Xi'an (northwest region), and Taiyuan (northern region), providing a geographically diverse representation of the Chinese population. The control group included 1199 healthy children who were recruited from the same hospitals and whose data were matched with the data of the Wilms tumor patients. Frequency matching of controls to cases was conducted based on age and sex to limit possible confounding variables. Informed consent was offered by the parents or legal guardians. The study protocol received approval from the Institutional Review Board of Guangzhou Women and Children's Medical Center (No. 202016601) and was conducted according to the ethical standards of the Declaration of Helsinki.
SNP selection and SNP genotyping
-
This study adopted a systematic approach to select candidate SNPs for the TRMT10C gene based on the criteria we described previously[29,33]. First, we queried the NCBI dbSNP database to identify all known SNPs in the TRMT10C gene region. These SNPs were filtered based on a minor allele frequency (MAF) greater than 5% in the Han Chinese population, specifically using data from the Beijing cohort (CHB) in the 1000 Genomes Project. This threshold ensures the selected SNPs are sufficiently frequent in the study population. Next, we used the SNPinfo Web Server (
https://snpinfo.niehs.nih.gov ) to identify potentially functional SNPs, such as nonsynonymous SNPs, as well as SNPs located in splicing sites, promoter or enhancer regions, and transcription factor binding sites. Finally, to ensure that the selected SNPs are not in strong linkage disequilibrium (LD), we performed LD analysis using the whole genome association analysis toolset PLINK. SNPs with high LD (R2 ≥ 0.8) were excluded to avoid redundancy in genetic information. Ultimately, four SNPs for genotyping that fit these criteria were selected: rs7641261 C > T, rs2303476 T > C, rs425758 A > G, and rs3762735 C > G.Peripheral blood samples were processed using the TIANamp Blood DNA Kit (TIANGEN Biotech Co. Ltd., Beijing, China). The DNA's concentration and purity were determined via spectrophotometry. DNA concentrations were normalized to the same level for all samples before genotyping with TaqMan real-time PCR assays[34−36]. The assays were prepared using specific probes for the selected SNPs, and the reaction mixture was dispensed into 384-well plates designed for high-throughput genotyping. The real-time PCRs were executed on an ABI Q6 instrument (Applied Biosystems, Foster City, CA, USA) to detect SNP variants. Several quality control measures have been implemented. First, each 384-well plate included both positive and negative controls to monitor assay performance. Second, 10% of the DNA specimens were randomly picked out for repeat genotyping, and the consistency between the initial and repeated genotyping results must be above 99%. Moreover, the laboratory personnel were blinded to the genotype.
Definition of expression quantitative trait loci (eQTLs)
-
We utilized the Genotype-Tissue Expression (GTEx) database to investigate whether the selected TRMT10C SNPs are eQTLs. GTEx is a comprehensive resource that links genetic variants with gene expression across various human tissues. We queried the SNPs of interest (rs7641261, rs2303476, rs4257518, and rs3762735) in the GTEx portal to evaluate their potential effects on the levels of TRMT10C and nearby genes.
Statistical analysis
-
All the statistical analyses were implemented via SAS v9.4 (SAS Institute Inc., Cary, NC). The chi-square (χ2) test or Fisher's exact test was applied to compare allele and genotype distributions for Wilms tumor patients and controls. We first evaluated the genotype distribution in control subjects to check whether they adhered to Hardy‒Weinberg equilibrium (HWE) via a goodness-of-fit χ2 test. We subsequently employed multiple inheritance models (homozygous, heterozygous, additive, dominant, and recessive) to test the associations of each SNP with Wilms tumor risk. Unconditional logistic regression was applied to assess odds ratios (ORs) and 95% confidence intervals (CIs) for each SNP, adjusted for potential confounders such as age and sex. To assess potential heterogeneity in genetic associations, we performed stratified analyses based on age, gender, and clinical stages of the Wilms tumor. Subgroup-specific analyses were conducted to explore whether TRMT10C polymorphisms exhibit differential effects across these variables. This approach accounted for demographic and clinical stratification to identify patterns potentially masked in the overall analysis. We applied a standard p-value threshold of 0.05 to determine nominal statistical significance.
This study was funded by grants from the Youth Medical Innovation and Practice Research Program of Guangzhou (Grant No: 2023QNYXYB010), Guangzhou Science and Technology Project (GrantNo: SL2024A04J02146, SL2024A03J01616), and the Major Science and Technology Special Project of Wenzhou (Grant No: ZY2020021). We appreciate Jiabin Liu and Linqing Deng for their assistance in DNA extraction and genotyping.
-
The study was conducted in accordance with the Declaration of Helsinki, and the protocol was approved by the Institutional Review Board of Guangzhou Women and Children's Medical Center (No. 202016601) on November 30, 2020.
-
The authors confirm contribution to the paper as follows: all authors contributed significantly to this work. Samples and data collection, research study performed: Deng C, Chen M, Zhou H, Zhang J, Cheng J, Li S, Fu W, Liu G, He J; data analysis: Hua RX, He J; study design: Niu H; draft manuscript preparation: Deng C, Zhu J, Duan F, He J; table and figure preparation: Hua RX, He J. All authors have read and approved the final manuscript to be published.
-
All the data are available upon request from the corresponding author.
-
The authors declare that they have no conflict of interest.
-
# Authors contributed equally: Changmi Deng, Jinhong Zhu, Fei Duan
- Supplementary Table S1 Frequency distribution of selected variables in Wilms tumor patients and cancer-free controls.
- Copyright: © 2025 by the author(s). Published by Maximum Academic Press, Fayetteville, GA. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Deng C, Zhu J, Duan F, Chen M, Zhou H, et al. 2025. TRMT10C polymorphisms confer Wilms tumor predisposition: a five-center study. Epigenetics Insights 18: e001 doi: 10.48130/epi-0024-0006 

TRMT10C polymorphisms confer Wilms tumor predisposition: a five-center study
- Received: 04 November 2024
- Revised: 07 December 2024
- Accepted: 16 December 2024
- Published online: 20 January 2025
Abstract: Wilms tumor is a prevalent childhood kidney cancer, which is known to be influenced by genetic predisposition. TRMT10C plays a role in RNA modification and mitochondrial function. This study explored the associations of four TRMT10C variants with susceptibility to Wilms tumor in Chinese children. This case‒control study included 414 cases diagnosed with Wilms tumor and 1199 controls. Four TRMT10C variants (rs7641261 C > T, rs2303476 T > C, rs4257518 A > G, and rs3762735 C > G) were genotyped by the TaqMan method. Logistic regression analyses were applied under multiple genetic models to determine the contributions of these genetic variants to Wilms tumor risk. None of the four TRMT10C variants was significantly associated with susceptibility to Wilms tumor. However, subgroup analysis revealed that the rs7641261 TT genotype enhanced the risk of Wilms tumor in boys and susceptibility to early-stage Wilms tumors. Moreover, individuals carrying multiple high-risk genotypes presented an elevated early-stage Wilms tumor predisposition. eQTL analysis demonstrated a significant correlation between the rs7641261 TT genotype and reduced transcript levels of SENP7. This study indicates that the TRMT10C rs7641261 polymorphism may confer Wilms tumor risk, particularly in boys and those with early-stage disease. These findings contribute to our comprehension of Wilms tumor's genetic basis.
-
Key words:
- Wilms tumor /
- Susceptibility /
- TRMT10C /
- Polymorphism