








-
Ulcerative colitis (UC) is a chronic immune-mediated inflammatory bowel disease that is recurrent and difficult to cure[1]. According to current research, the pathogenesis of UC may be associated with the disruption and imbalance in intestinal homeostasis by four mechanisms: dysbiosis of the gut microbiota, defects in the mucus layer, defects in the epithelium, and inflammatory responses in the lamina propria[2]. Currently, conventional treatments for patients with UC include the use of drugs such as steroids and thiopurines or surgery. However, these treatments have limitations, such as an increased risk of intestinal infections, reactivation of herpes zoster, and pulmonary embolism[3,4]. Therefore, it is urgent to develop and explore safe and effective therapeutic agents for preventing and treating UC.
Increasing evidence demonstrated that the interaction of the gut microbiota with the body's oxidative stress and inflammatory response is important for maintaining the normal barrier function of the gut[5,6]. Under normal conditions, mechanical and biochemical barriers exist between the microbiota and the gut mucosa. When the barrier is breached, the microbiota can be transferred from the intestinal lumen to the lamina propria and promote the emission of a range of inflammatory factors, resulting in an inflammatory response[7]. Remodeling the structure of the gut microbiota and restoring flora homeostasis and intestinal barrier function by changing external factors such as diet may be effective ways to combat UC.
β-ionone (ION) is an important ketone aromatic substance found in plants and is often used as a flavor enhancer in food. It has been demonstrated that ION possesses diverse biological activities including anticancer, chemopreventive, anti-inflammatory, and antimicrobial effects[8,9]. The precursors of ION, carotenoids can modify the gut microbiota. For example, Tu1927 carotenoids affect the levels of intestinal tight junction proteins, and this effect is related to modifying the composition of gut microbiota[10]. Ma et al. reported the anti-inflammatory activity of ION-curcumin hybrid derivatives in improving UC[11]. However, the preventive efficacy of ION against UC and its effects on the gut microbiota has not been studied. Accordingly, this study was designed to investigate whether ION could inhibit the development of UC in the dextran sulfate sodium (DSS)-induced model. Furthermore, this study attempted to illuminate the potential mechanisms underlying the action of ION on the development and progression of UC, including the potential regulation of the gut microbiota and gut barrier integrity.
-
DSS (CAS: 9011-18-1, 98% pure) was acquired from Yuanye Biotechnology Co. Ltd. (Shanghai, China). ION (CAS: 14901-07-6, 97% pure) was obtained from Sigma-Aldrich (St. Louis, MO, USA).
Animals and experimental design
-
Twenty-four male C57BL/6J mice aged 7 weeks, weighing 22 ± 1 g (Vital River Laboratories, Beijing, China) were housed in a specific pathogen-free animal facility at the Animal Center (SYXK (Jing) 2020-0052) and placed in a room with a light-dark (12 h-12 h) cycle, a temperature of 22 ± 2 °C, and a humidity of 40% to 70%. All experimental animals had free access to normal Huafukang feed.
The 24 mice were classified into three groups (n = 8) after 7 d of acclimatization: the CON group, the DSS group, and the ION group. The mice in the DSS and ION groups drank 1.5% (w/v) DSS solution, while the mice in the ION group were gavaged daily with 200 mg/kg ION dissolved in corn oil. The DSS solution was changed every 2 d. The food intake, water intake, body weight, and feces characteristics of the mice were observed and recorded during the experiment. After 10 d of intervention, experimental mice were fasted for 8 h and cervically dislocated and sacrificed. The mice were dissected from the cecum to the intestinal segment near the anus, and the colon length was photographed and recorded. The colons were collected and stored at –80 °C.
Disease activity index (DAI) score
-
The DAI score was assessed by evaluating changes in body weight, fecal traits, blood in the feces, and overall status of each mouse according to the literature[12].
Hematoxylin-eosin (H&E) staining and histopathological scoring of the colon
-
Approximately 1 cm of the colon was collected and then fixed in the 4% paraformaldehyde solution. The collected tissues were made into paraffin sections, stained with H&E, and then microscopically photographed. The histologic score of the colon was computed based on: inflammation level, range of inflammation, crypt gland damage, and depth of injury according to the literature[13].
Determination of serum lipidemia indexes
-
The collected blood was centrifuged at 3000 r/min for 15−20 min, and then the upper layer of serum was placed in a blood biochemistry analyzer for analysis. The measured blood biochemical indices included the levels of cholesterol (CHO), triglyceride (TG), high-density lipoprotein cholesterol (HDL-C), and low-density lipoprotein cholesterol (LDL-C).
Real-time quantitative PCR (RT-qPCR)
-
Total RNA was extracted from colon tissue using TRIzol reagent, and cDNA was reverse-transcribed using a kit from Tiangen Biotech (Beijing, China). RT-qPCR was performed using the CFX96 Touch Real-Time PCR Detection System (Bio-Rad, USA) and SYBR Green Supermix (Tiangen, Beijing, China). The primer sequences used for RT-qPCR are shown in Table 1. The mRNA expression levels of tumor necrosis factor-alpha (TNF-α) and interleukin-8 (IL-8) were normalized to that of glyceraldehyde-3-phosphate dehydrogenase (GAPDH).
Table 1. RT-qPCR experiment primer information comparison table.
Gene Forward primer 5'-3' Reverse primer 5'-3' GAPDH TGTGTCCGTCGTGGATCTGA CCTGCTTCACCACCTCTTGAT TNF-α CAGGCGGTGCCTATGTCTC CGATCACCCCGAAGTTCAGTAG IL-8 ATGACTTCCAAGCTGGCCGTGGCT TCTCAGCCCTCTTCAAAAACTTCTC Due to the inhibitory effect of DSS on the PCR process, the RT-qPCR mixture was adjusted by adding 0.2 g/L spermine[14]. The reaction protocol for RT-qPCR was as follows: initial denaturation at 95 °C for 15 min, followed by 40 cycles of denaturation at 95 °C for 10 s, annealing at 60 °C for 20 s, and extension at 72 °C for 25 s.
Detection of oxidative stress-related indices in the colon
-
The superoxide dismutase (SOD) activity was detected by assaying the absorbance at 450 nm with a SOD assay kit (Nanjing Jiancheng, #S0101S, Nanjing, China). For the lipid peroxidation assay, the malondialdehyde (MDA) was reacted with thiobarbituric acid (TBA) to form the MDA-TBA adducts with strong absorption at 532 nm (Nanjing Jiancheng, #S0131S, Nanjing, China). The catalase (CAT) activity was calculated by measuring the absorbance at 240 nm following the CAT kit instructions (Nanjing Jiancheng, #S0051, Nanjing, China).
Alcian blue periodic acid-Schiff (AB-PAS) staining of the colons
-
The colon sections were deparaffinized and oxidized for 5 min in the 0.5% periodic acid solution. After washing, the colon sections were incubated for 15 min in the Schiff reagent (Sigma, St. Louis, MI, USA), rinsed with warm water, and restained with hematoxylin for 1 min[15]. After staining with 3,3'-diaminobenzidine, the sections were examined microscopically. Finally, the positive area was quantified by using ImageJ (version 1.53).
Detection of tight junction protein expression in the mouse colon
-
The colon sections were incubated overnight at 65 °C and deparaffinized with xylene and gradient ethanol solutions. The sections were subjected to antigenic thermal repair using sodium citrate buffer. After successive blocking with serum and hydrogen peroxide, the primary antibodies: rabbit anti-zonula occludens-1 (ZO-1) (1:75; Servicebio, GB111402, Wuhan, China), murine anti-occludin (1:100; Servicebio, GB111401, Wuhan, China), and phosphate buffer were added to the negative control group. Horseradish peroxidase (HRP)-labeled murine and rabbit universal secondary antibodies (goat anti-rabbit IgG, 1:200, Servicebio, GB23303, Wuhan, China) were added, and after staining with hematoxylin and 3,3'-diaminopropylamine, the sections were observed microscopically. Finally, the positive area was quantified by using ImageJ (version 1.53).
Analysis of ION in the serum and colon contents of mice
-
The colon contents and serum were ultrasonicated with 10 mL of dichloromethane solution (ACS, Shanghai, China, CAS: 75-09-2). After centrifugation, the residue was extracted with dichloromethane solution and then mixed with the supernatant. The supernatant was dried with nitrogen at 40 °C. The residue was redissolved in hexane (ACS, Shanghai, China, CAS: 110-54-3) for gas chromatography-mass spectrometry (GC-MS) analysis. The samples were separated by a 30 m × 0.25 mm × 0.50 μm HP-5 column using the Agilent 6890N-G5795B (Agilent, Santa Clara, CA, USA).
Fecal DNA extraction and 16S rRNA gene sequencing
-
The extraction of fecal DNA was followed by the assessment of the purity and concentration of the DNA samples (n = 4) utilizing a NanoDrop-300 spectrophotometer. The primers 338F (5'-ACTCCTACGGGAGGCAGCAG-3') and 806R (5'-GGACTACHVGGGTWTCTAAT-3') were employed to amplify the V3-V4 regions of 16S rRNA gene fragments from the DNA samples. The PCR amplification was executed for a total of 27 cycles, with each cycle comprising denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s. Subsequently, paired-end sequencing of the amplified fragments was conducted using PE300 chemistry from Majorbio, facilitated by the Illumina MiSeq sequencing platform.
Analysis of the gut microbiota
-
Following demultiplexing and rigorous quality screening with fastp (version 0.19.6), the resulting sequences underwent merging with FLASH (version 1.2.11). Subsequently, the high-quality sequences were subjected to denoising utilizing the DADA2 plugin within the QIIME pipeline (version 2020.2), ensuring the accuracy and precision of the downstream analysis. Amplicon sequence variant (ASV) is a common term for DADA2 denoising sequences. The classification status of the ASVs was determined utilizing a Naive Bayes consensus classifier constructed within QIIME (version 2020.2), referencing the SILVA 16S rRNA database (version 138). Furthermore, the α-diversity and β-diversity indices were calculated and analyzed using QIIME, providing insights into the microbial community structure and diversity. Nonmetric multidimensional scaling (NMDS) analysis was performed using unweighted UniFrac distance-based analysis of similarities (ANOSIM) and visualized using the Majorbio cloud platform.
The linear discriminant analysis (LDA) effect size (LEfSe) analysis was performed with two stringent filters: an LDA score threshold of > 4 and a statistical significance level of p < 0.05. Since a potentially high rate of false positives of using LEfSe was previously described[16], the analysis composition of microbiomes (ANCOM) were also analyzed using the ANCOM 4.0.2 package in R to improve the discrimination between bacterial taxa enriched in different groups. W > 0.7 was considered to indicate statistical significance. Furthermore, the functional profiles of the gut microbiota were anticipated through PICRUSt. For statistical analysis, the STAMP software (
https://beikolab.cs.dal.ca/software/STAMP ) was utilized. Spearman correlation analysis and heat mapping were performed to observe the correlation of the presented differential gut bacteria taxa with body weight, the DAI, inflammation-related indicators, and intestinal barrier-related indicators on the Majorbio Cloud Platform (www.majorbio.com ).Data statistics
-
All the experimental data were analyzed using Microsoft Office Excel 2019, and the results are shown as the mean ± standard error of the mean (SEM). One-way ANOVA was used to analyze the differences with GraphPad Prism (version 9.0.0) software. Differences were considered statistically significant at p < 0.05.
-
In the current investigation, colitis was elicited through the administration of 1.5% (w/v) DSS (Fig. 1a), and the DSS group exhibited significant weight loss, diarrhea, hematochezia, and reduced intake of food and water (Fig. 1b−d). The ION-treated group had a significantly lower DAI, delayed weight loss, and attenuated colon shortening (Fig. 1e−h). Histological analysis further showed that ION significantly attenuated the inflammatory cell infiltration and mucosal injury, and reduced the overall histological score of the colon tissues (Fig. 1i & j). To gain a deeper understanding of ION's influence on intestinal inflammatory responses, the mRNA expression of proinflammatory cytokines were assessed. The results revealed that DSS treatment significantly upregulated the relative expression of TNF-α (Fig. 1k) and IL-8 (Fig. 1l). Notably, these increases were mitigated upon ION supplementation, indicating a potential regulatory role of ION in modulating intestinal inflammation.
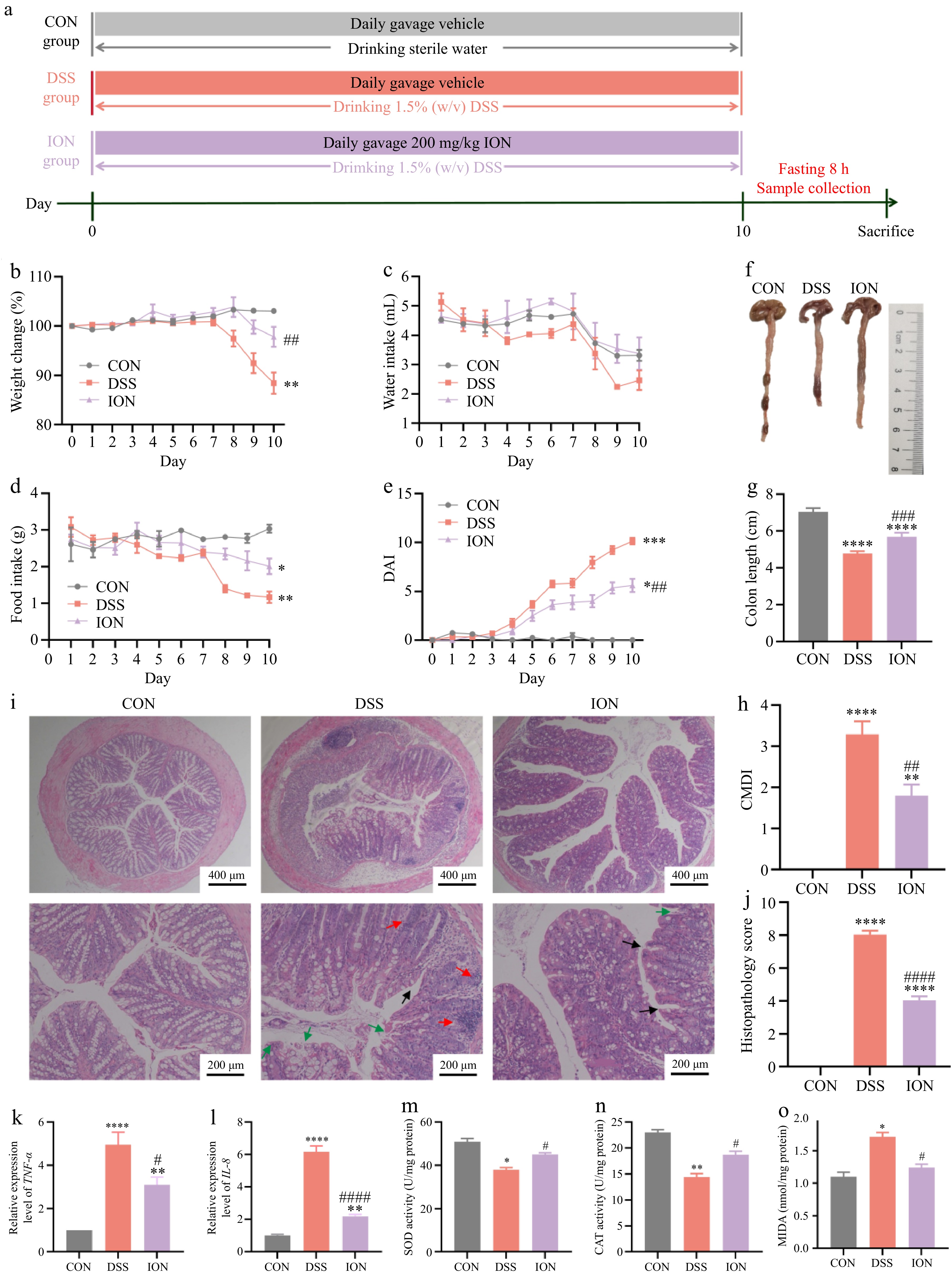
Figure 1.
ION prevents colitis symptoms, colonic damage, and oxidative stress in DSS-treated mice. (a) Design of experimental procedures for the prevention of ION on DSS-induced colitis. (b) Percentage of change in body weight. (c) Daily water intake. (d) Daily food intake. (e) The disease activity index (DAI) value. (f) Representative colon pictures. (g) Colon length. (h) Colon mucosa damage index (CMDI) value. (i) Representative images of colonic H&E staining (Upper: original magnification 40×, scale bar = 400 μm; bottom: original magnification 100×, scale bar = 200 μm. The red arrow represents the infiltration of inflammatory cells, the green arrow represents the damage of lumen surface, and the black arrow represents the destruction and bifurcation of intestinal crypt). (j) Histological scores of colons. The relative expression levels of (k) TNF-α and (l) IL-8 in colon tissues. The activities of (m) SOD and (n) CAT, and levels of (o) MDA in colon tissues. Data are shown as mean ± SEM. Statistical significance was determined by using one-way ANOVA followed by the Tukey test. * p < 0.05 vs CON, ** p < 0.01 vs CON, *** p < 0.001 vs CON, **** p < 0.0001 vs CON, # p < 0.05 vs DSS, ## p < 0.01 vs DSS, ### p < 0.001 vs DSS, #### p < 0.0001 vs DSS.
To evaluate the impact of ION on oxidative stress, the levels of SOD, CAT, and MDA were quantified within the colon tissues. As previously reported in the literature[17,18], the occurrence of UC is accompanied by elevated production of oxygen-free radicals, exemplified by an increase in MDA levels, and a concomitant decline in the levels of antioxidant enzymes SOD and CAT, leading to oxidative stress-induced injury of the intestinal mucosa. In line with these findings, the present study observed significantly reduced activities of SOD and CAT in the DSS-treated group compared to the CON group. Notably, ION supplementation effectively reversed this decline in enzyme activities (Fig. 1m & n). Furthermore, the DSS group exhibited a marked elevation in MDA content, which was also reversed upon ION treatment, as shown in Fig. 1o.
Numerous studies have documented the link between DSS-induced colitis and dyslipidemia[19]. Therefore, in the present study, the analysis of serum lipid parameters were investigated, including CHO, TG, HDL-C, and LDL-C, in mice subjected to DSS-induced colitis. The potential modulating effect of ION on DSS-mediated disruptions in lipid metabolism were further explored. Consistent with previous findings, DSS treatment led to a notable elevation in serum TG and LDL-C levels, coupled with a decrease in HDL-C levels. Notably, ION supplementation was effective in reversing these adverse changes, as illustrated in Fig. 2, suggesting a favorable impact on lipid homeostasis.
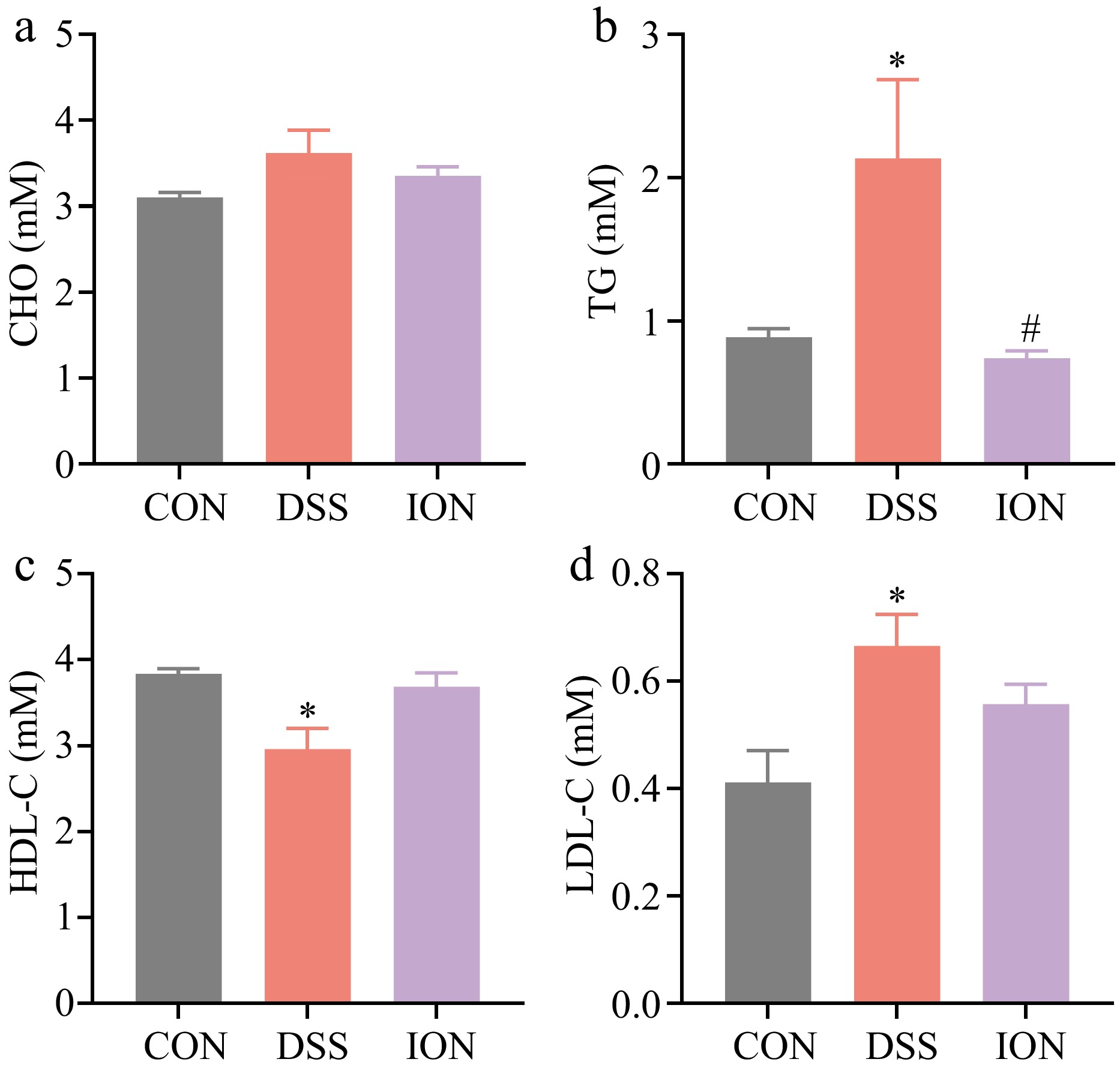
Figure 2.
Effect of ION on serum lipidemia indexes in DSS-treated mice. (a) CHO levels in serum. (b) TG levels in serum. (c) HDL-C levels in serum. (d) LDL-C levels in serum. Data are presented as mean ± SEM (n = 4). Statistical significance was determined using one-way ANOVA, followed by the Tukey test. * p < 0.05 vs CON, # p < 0.05 vs DSS.
Overall, these results demonstrated that ION could prevent and relieve DSS-induced colitis symptoms, colonic injury, and oxidative stress.
ION alleviates DSS-induced impairment of colonic barrier function
-
To gain insights into the effect of ION on the colonic mucosal barrier function, the colonic mucin secretion levels and the expression of tight junction proteins, notably occludin and ZO-1 were assessed. This analysis aimed to elucidate the potential role of ION in maintaining the integrity and functionality of the colonic barrier. When the mucosa is damaged, colonic mucins are expressed by epithelial cells and function as chemical barriers[20]. After AB-PAS staining, the mucins in the colonic mucosa showed a bluish-violet color, and the level of the colonic mucins was evaluated by observing the color and distribution of the stained areas. In the present study, AB-PAS staining revealed a significant decrease in optical density and this inhibitory effect was significantly attenuated by ION supplementation, suggesting that ION increases mucin secretion.
The present findings indicated that ION possesses the potential to enhance the intestinal mucosa's repair capabilities, as evidenced by the results presented in Fig. 3a & b. Notably, ZO-1 and occludin, two crucial tight junction proteins essential for regulating cell proliferation, differentiation, and growth in intestinal epithelial cells are known to exhibit reduced expression in inflammatory bowel disease[21]. The immunohistochemical analysis revealed a significant decrease in the expression of both ZO-1 and occludin in the DSS-treated group compared to the CON group. ION supplementation mitigated this loss to a considerable extent, as depicted in Fig. 3c−f. These observations suggested that ION effectively alleviates the DSS-induced impairment of colonic barrier function.
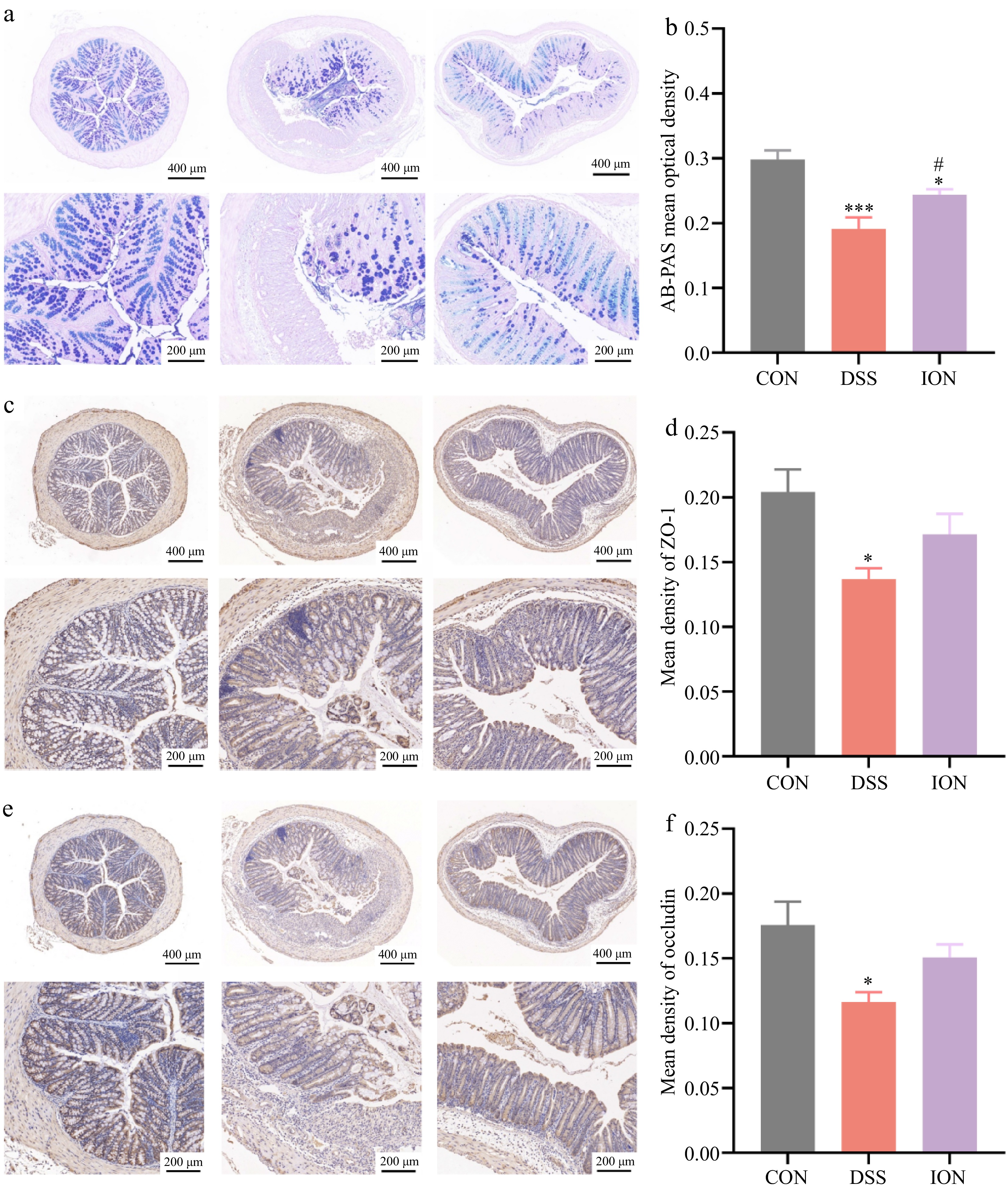
Figure 3.
ION alleviates DSS-induced impairment of colonic barrier function. (a) Representative AB-PAS-stained sections of the colon. (b) AB-PAS-stained optical density values. (c) Immunohistochemistry-stained sections of colons were stained for ZO-1. (d) Optical density values of immunohistochemistry-stained sections of ZO-1. (e) Immunohistochemistry-stained sections of the colon were stained for occludin. (f) Optical density values of immunohistochemistry-stained sections of occludin. (Upper: original magnification 40×, scale bar = 400 μm; bottom: original magnification 100×, scale bar = 200 μm). Data are shown as mean ± SEM. Statistical significance was determined by using one-way ANOVA followed by the Tukey test. * p < 0.05 vs CON, *** p < 0.001 vs CON, # p < 0.05 vs DSS.
ION regulates the composition and function of the gut microbiota
-
Employing an independent cohort of mice, GC-MS was employed to quantify ION levels in both serum and colonic contents. This approach aimed to elucidate the tissue distribution pattern of ION within the mice, providing valuable insights into its bioavailability and potential sites of action. The results revealed that the average concentration of ION in the serum was 1.65 μM, and the average concentration of ION in the colonic contents was 5.71 mM/kg (Table 2), suggesting that ION may directly interact with and affect colon tissues and the gut microbiota.
Table 2. Content of ION in serum and colonic contents of mice.
Tissue Serum (μM) Colonic contents (mM/kg) ION levels 1.65 ± 0.70 5.71 ± 1.95 To investigate the potential of ION to modulate the composition and function of the gut microbiota, fecal DNA was extracted from mice belonging to three distinct groups and subjected to 16S rRNA sequencing. The ASV analysis revealed that DSS treatment resulted in an augmentation of gut microbiota α-diversity, as evidenced by notable increases in the Shannon index, Shannoneven index, Simpsoneven index, and Pd index, along with a significant decrease in the Simpson index. Notably, ION intervention effectively reversed these DSS-induced alterations (Fig. 4a−e). Furthermore, an NMDS plot at the genus level highlighted significant variations in the gut microbiome composition among the three groups, indicating that ION significantly reshaped the gut microbiota structure in mice with UC (Fig. 4f).
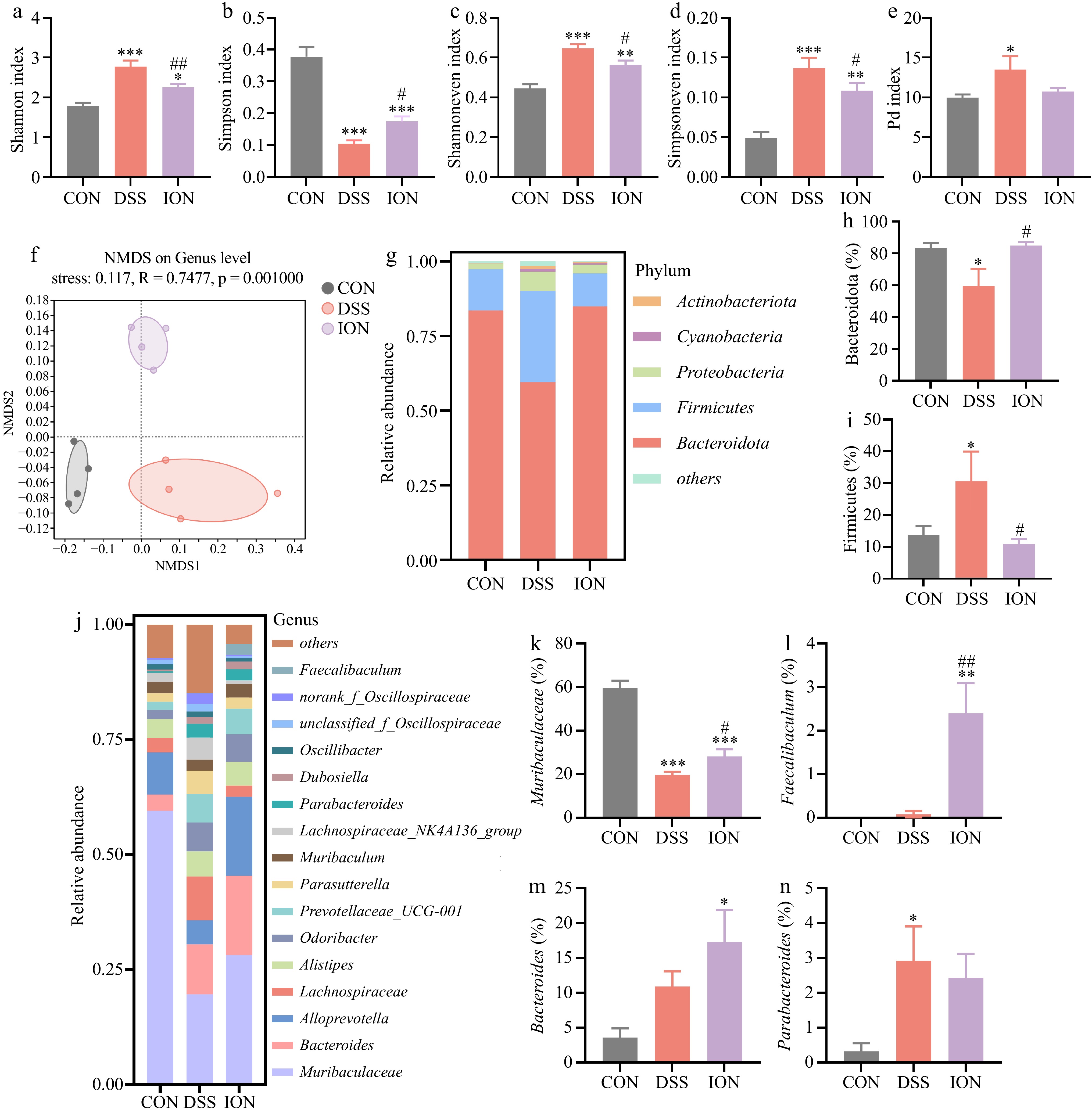
Figure 4.
ION affects the structure and relative abundance of gut microbiota in DSS-induced UC mice. (a) The level of the Shannon index. (b) The level of the Simpson index. (c) The level of the Shannoneven index. (d) The level of the Simpsoneven index. (e) The level of the Pd index. (f) The NMDS analysis on ASV level. (g) The relative abundance at the phylum level. (h) The relative abundance of Bacteroidota. (i) The relative abundance of Firmicutes. (j) The relative abundance at the genus level. (k) The relative abundance of Muribaculaceae. (l) The relative abundance of Faecalibaculum. (m) The relative abundance of Bacteroides. (n) The relative abundance of Parabacteroides. Data are shown as mean ± SEM (n = 4). Statistical significance was determined by using one-way ANOVA followed by the Tukey test. * p < 0.05 vs CON, ** p < 0.01 vs CON, *** p < 0.001 vs CON, # p < 0.05 vs DSS, ## p < 0.01 vs DSS.
A comprehensive analysis of the gut microbiota composition was undertaken at both the phylum and genus levels (Fig. 4g & j). At the phylum level, notable differences emerged, with the ION group exhibiting a significantly elevated relative abundance of the Bacteroidota phylum (Fig. 4h) and a reduced relative abundance of the Firmicutes phylum (Fig. 4i), in comparison to the DSS group. Delving deeper into the genus level, the ION group displayed significantly higher relative abundances of Muribaculaceae, Faecalibaculum, and Bacteroides, whereas the abundance of Parabacteroides was lower, effectively reversing the alterations induced by DSS treatment to a certain degree (Fig. 4k−n). These findings underscore the ability of ION to modulate the gut microbiota composition in UC mice.
To gain a nuanced understanding of the differences in gut microbial composition across the three groups, LEfSe analysis was conducted, spanning from the phylum to the genus level, with statistical significance set at p < 0.05 and an LDA score threshold of > 4. The findings illuminated distinct microbial signatures within each group. In the DSS group, Lachnospirale emerged as the predominant phylum, while Clostridia dominated at the class level, Lachnospiraceae and Tannerellaceae dominated at the family level, and Parabacteroides and Lachnospiraceae_NK4A136_group dominated at the genus level, respectively. The ION group was characterized by the dominance of Erysipelotrichales at the phylum level, Bacilli at the class level, Bacteroidaceae and Erysipelotrichaceae at the family level, and Bacteroides and Faecalibaculum prevailing at the genus level (Fig. 5a & b). The ANCOM method was further used to reduce false-positive results and increase the efficiency of the analysis compared to traditional LEfSe[22]. Upon setting a significance threshold of W > 0.7, the analysis revealed a notable enhancement in the abundance of Faecalibaculum in the ION group, in stark contrast to the DSS group (Table 3). Consequently, Faecalibaculum (LDA > 4) was found to be a key bacterium in the ION group according to the LEfSe and ANCOM analyses.
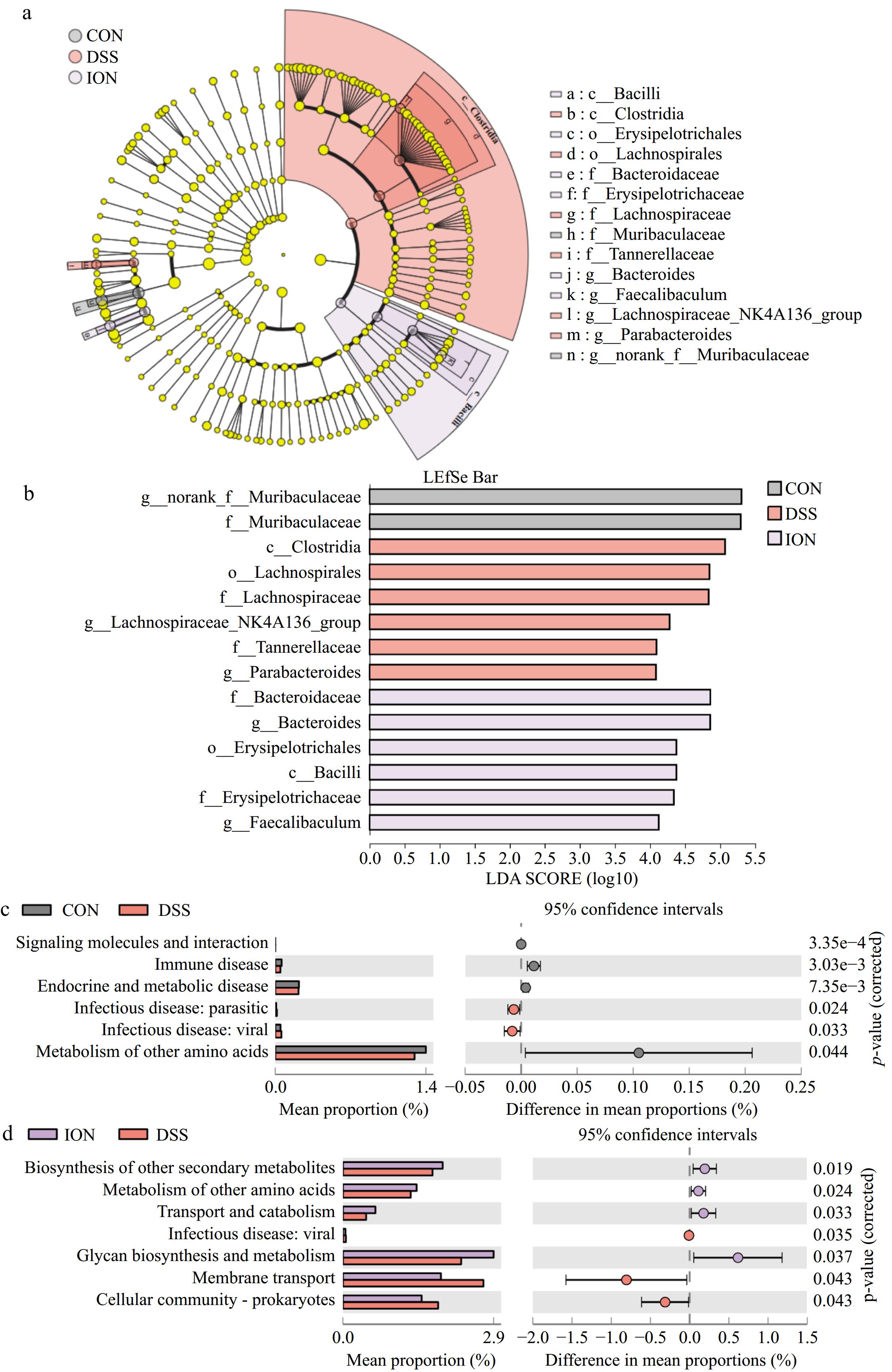
Figure 5.
ION supplementation changes the key phylotypes and function of the gut microbiota of UC mice. (a) LEfSe analysis (the rings, from inner to outer: phylum, class, order, family, and genus). (b) Gut microbiota between CON, DSS, and ION groups with LDA score > 4 and p < 0.05. (c) Comparison of KEGG pathways in the CON and DSS groups. (d) Comparison of KEGG pathways in the ION and DSS groups. Statistical differences (p < 0.05) were analyzed by using the unpaired two-tailed t-test in STAMP (n = 4).
Table 3. ANCOM analysis among the CON, DSS, and ION groups.
Levels Species name W-value Significance
(detected_0.7)CON vs DSS Phylum Deferribacterota 11 Insignificant Eubacterium_
siraeum_group113 Significant Genus Marvinbryantia 102 Significant Mucispirillum 86 Insignificant ION vs DSS Phylum Deferribacterota 11 Insignificant Verrucomicrobiota 10 Insignificant Genus Faecalibaculum 117 Significant Mucispirillum 85 Insignificant To explore how the microbial changes induced by DSS and ION in mice affects metabolism, PICRUSt was used to predict potential functional pathways that may be affected by the gut microbiota. These results suggested that various pathways were significantly modified by DSS and ION. The metabolism of other amino acids tends to be reduced, and infectious disease : viral tends to occur in the DSS group compared to the CON group. Moreover, ION significantly reverses these metabolic pathways, indicating that ION can reverse the dysfunction of the gut microbiota induced by DSS to some degree. In addition, ION also regulates certain functional pathways. For example, ION upregulated glycan biosynthesis and metabolism functional pathways, transport and catabolism, and biosynthesis of other secondary metabolites, as well as downregulated the membrane transport and cellular community-prokaryotes functional pathways (Fig. 5c & d).
In conclusion, the preventative effects of ION in mice with DSS-induced UC likely stems from its ability to fortify the gut barrier and modulate the gut microbiota. Notably, genera including Faecalibaculum and Mucispirillum emerge as key players in this protective mechanism, contributing significantly to ION's ability to safeguard mice against UC.
Correlation analysis of the gut microbiota and UC-related indices
-
To delve deeper into the intricate link between genus-level bacterial characteristics and disease status, a Spearman correlation analysis was performed. The outcomes robustly demonstrated a strong association between alterations in the gut microbiota and UC-related indices. Specifically, Lachnospiraceae_UCG_001 and norank_f_Muribaculaceae abundances were positively associated with favorable markers such as body weight changes, colon length, and elevated ZO-1 and occludin expression levels, while inversely correlated with disease severity indicators like DAI, CMDI score, and TNF-α and IL-8 expression levels (Fig. 6). Conversely, the abundance of Parabacteroides showed a positive correlation with markers of disease severity (DAI, CMDI score, and IL-8 expression) and a negative correlation with colon length and body weight, underscoring its potential role in disease progression.
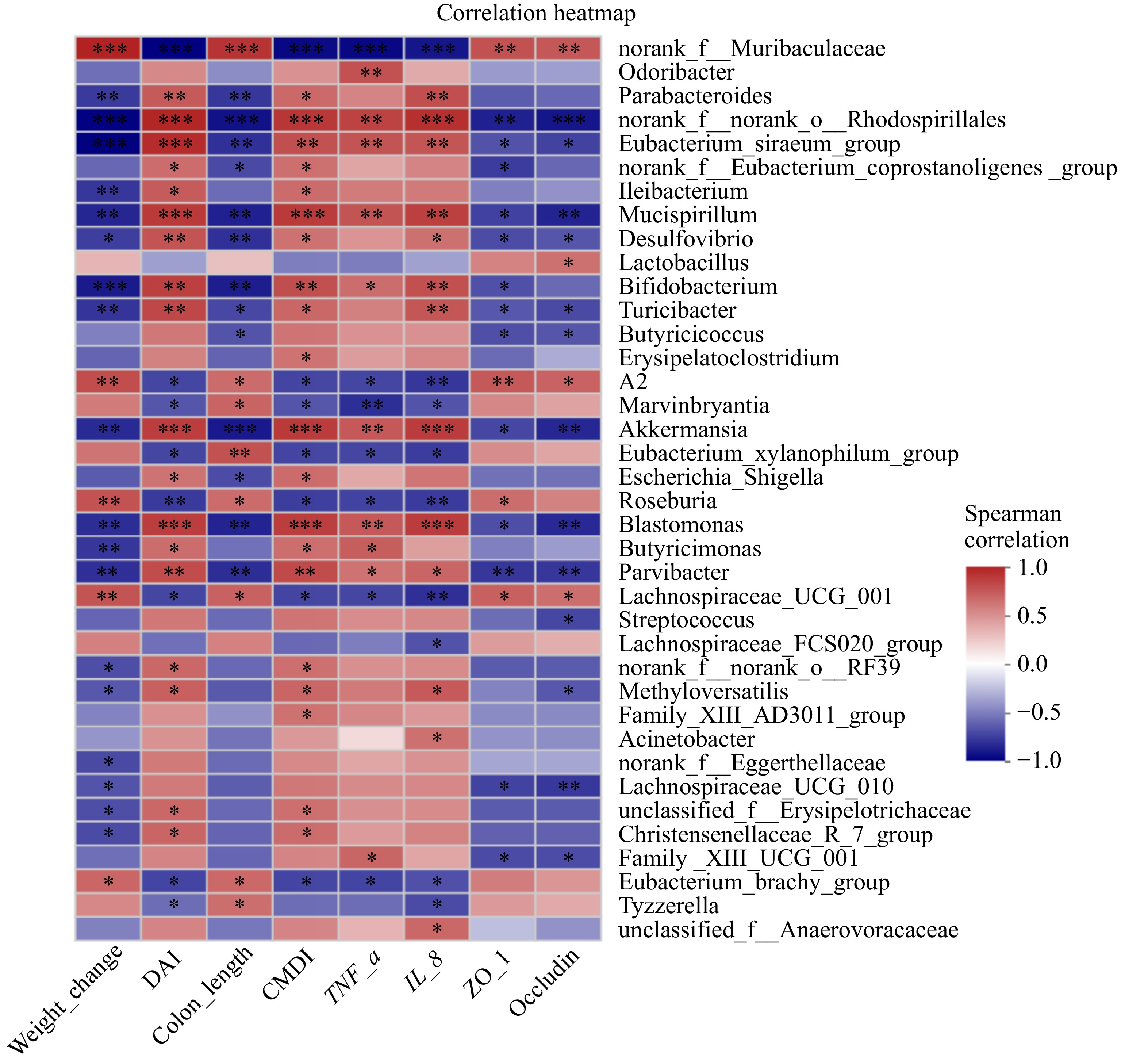
Figure 6.
Correlation analysis of gut microbiota at the genus level and UC-related indicators. Spearman correlation, showing only the genera having significant correlations with UC-related indicators using two filters: |correlation coefficient| > 0.6 and p < 0.05 (n = 4). Scale (right legend) indicates the level of positive (red) or negative (blue) correlation. The significant differences between groups are expressed as: * p < 0.05; ** p < 0.01; *** p < 0.001.
In summary, ION demonstrated a potent ability to mitigate DSS-induced colonic inflammation and damage, thereby preventing the onset of UC in mice. These preventive effects are likely underpinned by its dual mechanisms of action: reinforcing the intestinal barrier and modulating the gut microbiota.
-
Imbalance of intestinal homeostasis is a recently accepted theory in the academic community that explains the pathogenesis of UC. The disruption of the intestinal barrier, characterized by malfunctioning of the mucus layer and intestinal epithelium, coupled with an imbalance in gut microbiota homeostasis, can precipitate inflammatory cascades and oxidative stress, ultimately fostering the progression of UC[2,23]. This study underscored the efficacy of ION in preventing DSS-induced UC in mice by synergistically safeguarding the mucus layer, fine-tuning the composition and function of the gut microbiota, preserving intestinal homeostasis, and mitigating inflammatory processes within the intestinal tract.
The DSS-induced mouse model is an ideal animal model for UC because of its simplicity, high success rate, and reproducibility, especially because the induced symptoms match the clinical characteristics of UC patients. Numerous researchers have induced the formation of an acute colitis model by administering a 1% (w/v) to 6% (w/v) solution of DSS to rodents. For example, Peng et al. showed that after 7 d of induction with 3% (w/v) DSS, mice exhibited watery feces and weight loss[24]. Moura et al. successfully induced a UC model using 2% (w/v) DSS[25]. Agista et al. reported that diarrhea and blood in the stool of mice appeared in mice on the 4th day after drinking 1.5% DSS[26]. Combining the results of previous studies and previous experimental explorations, a 1.5% (w/v) dose of DSS was selected for the UC model and the protective effect of ION evaluated. The results conclusively demonstrated the successful establishment of the UC model within 10 d of administering 1.5% DSS, as evidenced by the emergence of blood in stool, diarrhea, weight loss, colon shortening, pathological damage, and pronounced intestinal inflammation in the mice (Fig. 1).
The absorption of β-carotene, the precursor substance of ION, by the small intestine and its translocation to peripheral tissues and serum have been reported, but pharmacokinetic studies on ION are limited[27]. The levels of ION in the serum and colonic contents of mice after oral administration of ION for 6 h by GC-MS were determined to investigate the pharmacokinetics of ION. The tissue distribution results showed that ION could accumulate in the mouse colonic contents after oral administration (Table 2), indicating that ION could be effectively absorbed through the oral route and that most ION would be excreted from the body through the feces after oral administration, which may provide ION with direct access to colonic tissue and gut microbiota.
The causes of UC pathogenesis include the inflammatory response, imbalance of oxidative stress, and intestinal barrier dysfunction. ION is a natural ketone compound widely found in vegetables and fruits and is a recognized safe flavoring agent. Studies have shown that ION, both endogenous and exogenous, has anticancer, chemopreventive, melanogenic, anti-inflammatory, and antimicrobial effects[8]. Recently, Yin et al.[28] illuminated ION's potential in suppressing Aspergillus fumigatus growth and mitigating fungal keratitis in mice by modulating inflammation through hemagglutinin-like oxidized LDL receptors and activating the JNK/p38 MAPK pathway. Inspired by these physiological insights, it was postulated that ION could serve as a preventive agent against UC progression. The present study corroborates this hypothesis, revealing that ION significantly mitigated body weight loss, DAI, colonic shortening, CMDI, and histopathological injury in DSS-induced UC mice (Fig. 1), aligning with Ma et al.'s findings on ION-curcumin hybrids' protective effects against colonic damage[11]. Notably, DSS-induced elevations in TNF-α and IL-8 mRNA expressions (Fig. 1k & l), consistent with previous literature[29], were significantly attenuated by ION intervention at 200 mg/kg (Fig. 1k & l), echoing Yin et al.'s observations[28]. Furthermore, ION modulated oxidative stress by enhancing SOD and CAT activities while decreasing MDA levels (Fig. 1m−o), in line with ION's established antioxidant properties[29]. Collectively, these findings underscore ION's efficacy in reducing DSS-induced colonic inflammation and injury, thereby preventing UC in mice.
The potential preventive effect of ION on DSS-induced UC in mice may be intimately linked to its ability to safeguard the gut barrier. Notably, the mucus layer plays a pivotal role in demarcating the intestinal internal and external environments, facilitating mucosal lubrication, and impeding the infiltration of harmful substances into the intestinal lumen[30]. Thus, ION's protective action on this critical barrier could underlie its efficacy in mitigating UC development. As expected, DSS induced a decrease in mucin secretion and ION inhibited this decrease, thereby protecting the intestinal mucosal barrier (Fig. 3a & b). Moreover, epithelial cells form a tightly sealed epithelial barrier between them through tight junctions[31]. Abnormal expression of tight junction proteins is considered to be one of the key contributors responsible for the destruction of the intestinal epithelial barrier in patients with UC[32]. Herein, the results demonstrated that ION inhibited the DSS-induced reduction in occludin and ZO-1 expression (Fig. 3c−f), indicating that ION intervention significantly improved intestinal barrier function and enhanced the integrity of the mucosal barrier and tight junctions in DSS-induced UC mice.
A symbiotic relationship exists between the host and the gut microbiota and dysbiosis of the gut microbiota serves as one of the hallmarks of intestinal diseases. Changes in the gut microbial composition of UC patients have been reported many times and such alterations are closely linked to changes in bacterial composition, structure, function, and interactions[33]. For example, Hu et al. reported that VK2 attenuates DSS-induced colitis by enhancing the dominant gut microflora, such as the Faecalibaculum[34]. Microbial stability is negatively correlated with chronic diseases and metabolic dysfunction, so stable markers such as abundance and diversity are often used as indicators of gut health status[35]. The present study demonstrated that DSS increased the α diversity of the gut microbiota (Fig. 4a−e), which is in line with the study of Tian et al.[36]. However, there are also some reports of DSS-induced reductions in gut microbial diversity in UC mice[37]. This difference may be related to the mouse strain, intervention time, DSS intervention concentration, and feeding environment. Colonic mucosal barrier defects promote intestinal pathogen invasion[38]; thus, it was considered that the DSS-induced increase in gut microbial α diversity may result from the invasion of harmful microorganisms associated with UC with increased relative abundance. Moreover, it was observed that ION restored the disordered α diversity caused by DSS (Fig. 4a−e).
Dietary interventions are important external factors influencing the gut microbiota, and there is increasing evidence to support this view[39,40]. Therefore, the impact of ION on the gut microbiota was assayed. The experimental results demonstrated that gut microbial communities were altered after DSS treatment and that these alterations were reversed by ION intervention to a certain extent. In particular, after ION treatment, the abundance of Bacteroidota was remarkably enhanced, and the abundance of Firmicutes was significantly reduced (Fig. 4g−i). Moreover, ION effectively boosted the relative abundance of Muribaculaceae and Faecalibaculum in the intestine (Fig. 4k−l). Among them, Bacteroidota, Muribaculaceae, and Faecalibaculum are beneficial gut microbiota that play a variety of important physiological roles in the intestinal tract, including helping to digest food, maintain intestinal health, and regulate the immune system[41,42]. Spearman correlation analysis further confirmed that the changes in a variety of ION-induced gut microbiota constituents were correlated with the changes in pathophysiological indicators of UC (Fig. 6). The above results indicated that the preventive effect of ION on DSS-induced colitis may be related to the modification of the gut microbiota.
Several studies have indicated that UC patients have altered amino acid metabolism, and the amino acids that are altered include valine, tryptophan, glycine, and serine[43]. However, the gut microbiota can inhibit inflammatory responses by metabolizing branched-chain and aromatic amino acids, which regulate the immune system and promote intestinal mucosal cell growth and repair[44,45]. For example, the gut microbiota can transform tryptophan into other metabolites, such as indole-3-acetic acid, which also has good preventive effects on UC[46]. In this study, the functional pathway related to the metabolism of other amino acids was downregulated in UC mice, while ION upregulated this functional pathway (Fig. 5c & d ). These findings suggested that ION might be able to prevent DSS-induced UC by affecting the amino acid metabolism of the gut microbiota. On the other hand, the gut microbiota can reduce viral pathogenicity by occupying ecological niches in the gut, preventing viral invasion and reproduction, and interacting with other microorganisms to regulate the homeostasis of the intestinal microbial community, reduce the risk of viral infection, and alleviate conditions[47]. The present results showed that the incidence of infectious disease:viral tended to decrease in the ION group, indicating that ION improved intestinal immunity in mice and thus acted as a preventive agent against DSS-induced UC (Fig. 5d). These results suggest that the mechanism by which ION interferes with the onset and progression of ulcerative colitis may be associated with influencing the composition and structure of the gut microbiota, thus modulating the gut immune system and reducing inflammation by producing beneficial metabolites via the 'metabolism of other amino acids' pathway as well as regulating the balance of the gut immune system and microbial community via the 'infectious diseases : viruses' pathway.
In conclusion, ION was effective in preventing DSS-induced UC in mice and its mechanism may be associated with the protection of the intestinal barrier and regulation of the gut microbiota, among which Faecalibaculum and Mucispirillum may play a role in the prevention of UC in mice.
-
This study demonstrated that ION could prevent DSS-induced colitis by reducing colonic pathological damage, decreasing the levels of proinflammatory cytokines, relieving oxidative stress, and preventing intestinal barrier destruction. AB-PAS staining and immunohistochemical staining analyses showed that ION significantly increased the amount of mucin and enhanced the expression of ZO-1 and occludin in UC mice, suggesting that ION alleviated UC by protecting the mucus barrier and maintaining the integrity of the intestinal epithelium. In addition, 16S rRNA sequencing analysis revealed that ION significantly restored the composition, structure, and function of the disturbed gut microbiota in UC mice. Therefore, the mitigating effect of ION on DSS-induced UC might be related to protecting the intestinal barrier, regulating the gut microbiota, and restoring the bacteria to a healthy state. Thus, ION may be a valuable food supplement or nutraceutical for managing and preventing UC. However, the present study also has certain limitations. The animal model used mimics some typical histopathology and immunological characteristics of patients with colitis, but the model does not fully represent the complex features of the disease. Furthermore, the present study demonstrated a correlation between UC-related indicators and gut microbiota but did not further investigate the signaling pathways and detailed mechanisms by which the ION prevented UC. Therefore additional potential mechanisms are needed, and further clinical studies in patients with colitis are warranted. Overall, the present results are still preliminary and require further verification and clarification of the specific mechanism of ION in preventing DSS-induced colitis, as well as the causal relationship with the regulation of gut microbiota.
-
All experimental procedures were conducted according to the guidelines provided by the Animal Care Committee. The China Agricultural University Laboratory Animal Welfare and Animal Experimental Ethical Committee approved the animal experimental protocol used in this study (AW40702202-4-4).
-
The authors confirm contribution to the paper as follows: study conception and design: Tong T, Huang K, Kang SG; draft manuscript preparation: Fang J, Wang Y; experiments conduction: Liu T; analysis and interpretation of results: Fang J, Liu T; manuscript revision: Fang J, Tong T. All authors reviewed the results and approved the final version of the manuscript.
-
The datasets generated during and/or analyzed during the current study are available from the corresponding author upon reasonable request.
This research was funded by the Science and Technology project of Xizang Autonomous Region (XZ202401ZY0092) and Chinese Universities Scientific Fund (2024TC083).
-
The authors declare that they have no conflict of interest.
-
Authors contributed equally: Jingjing Fang, Tingting Liu
- Copyright: © 2024 by the author(s). Published by Maximum Academic Press on behalf of China Agricultural University, Zhejiang University and Shenyang Agricultural University. This article is an open access article distributed under Creative Commons Attribution License (CC BY 4.0), visit https://creativecommons.org/licenses/by/4.0/.
-
About this article
Cite this article
Fang J, Liu T, Wang Y, Kang SG, Huang K, et al. 2024. β-ionone prevents dextran sulfate sodium-induced ulcerative colitis and modulates gut microbiota in mice. Food Innovation and Advances 3(4): 320−331 doi: 10.48130/fia-0024-0031 

β-ionone prevents dextran sulfate sodium-induced ulcerative colitis and modulates gut microbiota in mice
- Received: 25 July 2024
- Revised: 25 September 2024
- Accepted: 25 September 2024
- Published online: 22 October 2024
Abstract: β-ionone has various biological activities, such as anti-inflammatory, antimicrobial, and anticancer effects. The pathogenesis of ulcerative colitis is correlated with immune dysfunction, intestinal barrier damage, and gut microbiota imbalance. However, whether β-ionone has preventive efficacy against ulcerative colitis is unknown. This study investigated the effect of β-ionone on dextran sulfate sodium-induced ulcerative colitis and the underlying molecular mechanisms involved. The ulcerative colitis mouse model was induced by 1.5% dextran sulfate sodium for 10 d. Meanwhile, 200 mg/kg β-ionone was administrated to the mice. Body weight, colon length, colon tissue pathology, colon tissue inflammatory cytokines, colonic oxidative stress, and barrier function were assessed. The composition and structure of gut microbiota were profiled using 16S rRNA sequencing. The results showed that β-ionone supplementation effectively prevented ulcerative colitis by ameliorating colonic tissue damage, reducing inflammatory phenomena, and protecting the colonic epithelial mucosal barrier. β-ionone also protected mice from dextran sulfate sodium-induced gut microbiota disturbance by modifying the overall structure and function of the gut microbiota community and increasing the relative abundance of beneficial gut microbiota. The Spearman correlation analysis revealed that the changes in abundance of the gut microbiota were correlated with ulcerative colitis-related indicators. Overall, this study demonstrated that β-ionone has a preventive effect on ulcerative colitis in mice, and the underlying mechanism may be associated with the protection of the gut barrier and regulation of the gut microbiota. These results are conducive to promoting clinical trials and product development of β-ionone for the prevention and treatment of ulcerative colitis.
-
Key words:
- Gut microbiota /
- Intestinal barrier /
- Ulcerative colitis /
- β-ionone